
Dual Acting Carbon Monoxide Releasing Molecules and Carbonic Anhydrase Inhibitors Differentially Modulate Inflammation in Human Tenocytes

 , , and
, , and 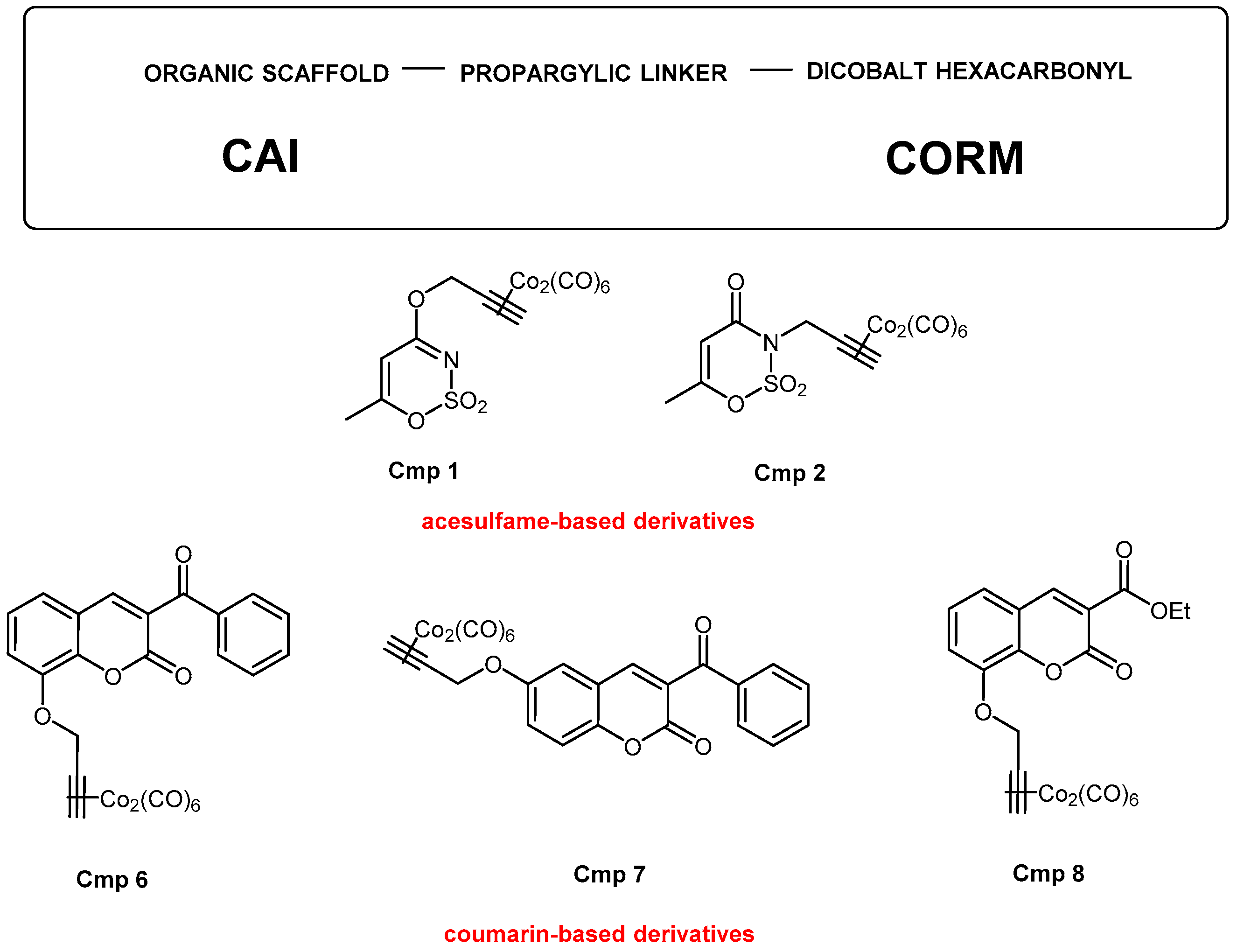{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Synthesis of CAI-CORM Hybrids
2.2. Cell Culture
2.3. Cell Metabolic Activity
2.4. Cytotoxicity Assay
2.5. Quantification of Protein
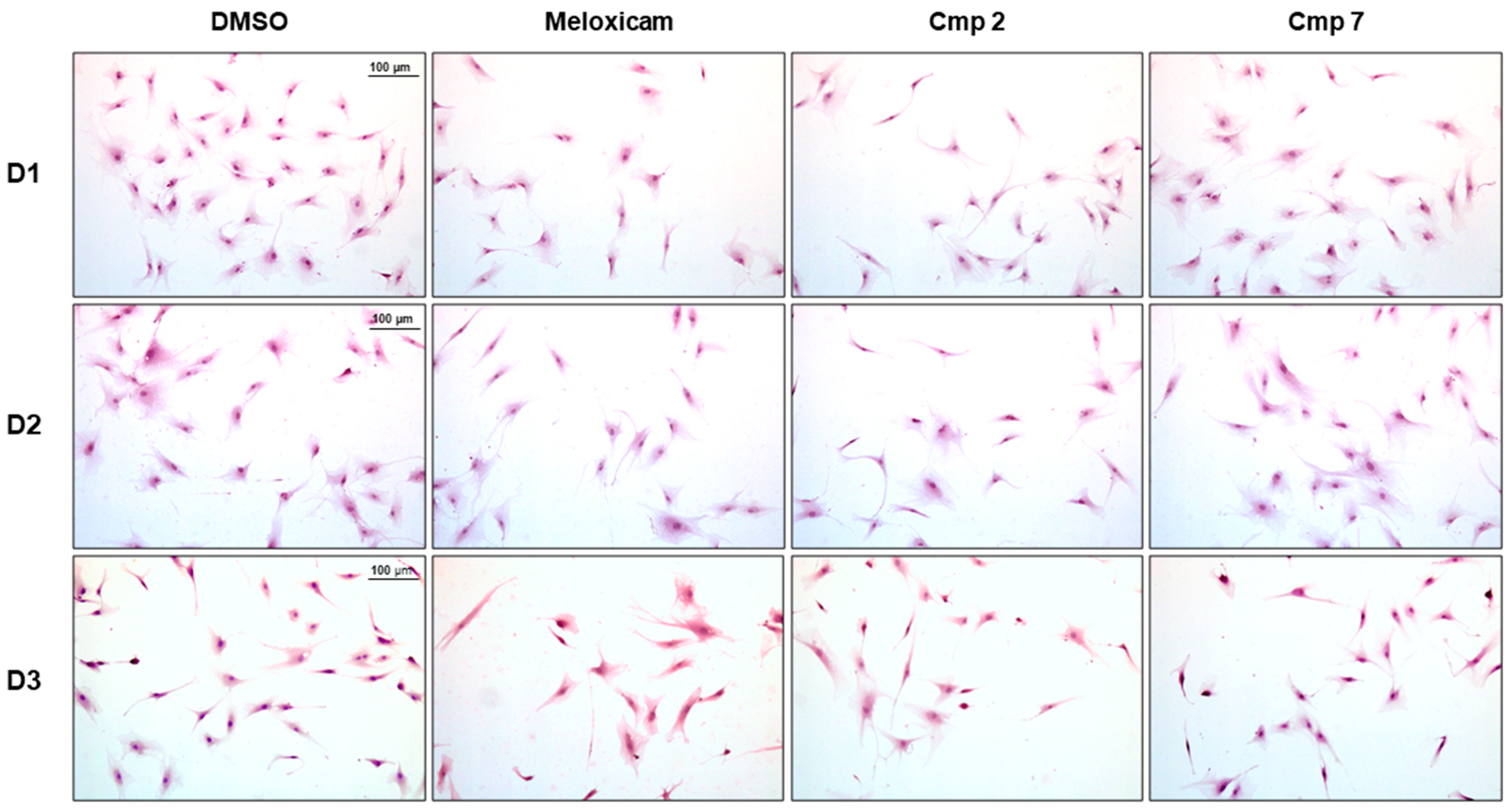
2.6. Hematoxylin/Eosin Staining
2.7. Cell Cycle Analysis
2.8. Collagen Type I Secretion
2.9. Detection of Mitochondrial Superoxide Anions by Flow Cytometry
2.10. Protein Expression by Western Blot Analysis
2.11. Immunofluorescence Staining of NF-kB
2.12. Statistics
3. Results and Discussion
3.1. Establishment of the Inflammatory Cell Model and Preliminary Investigations
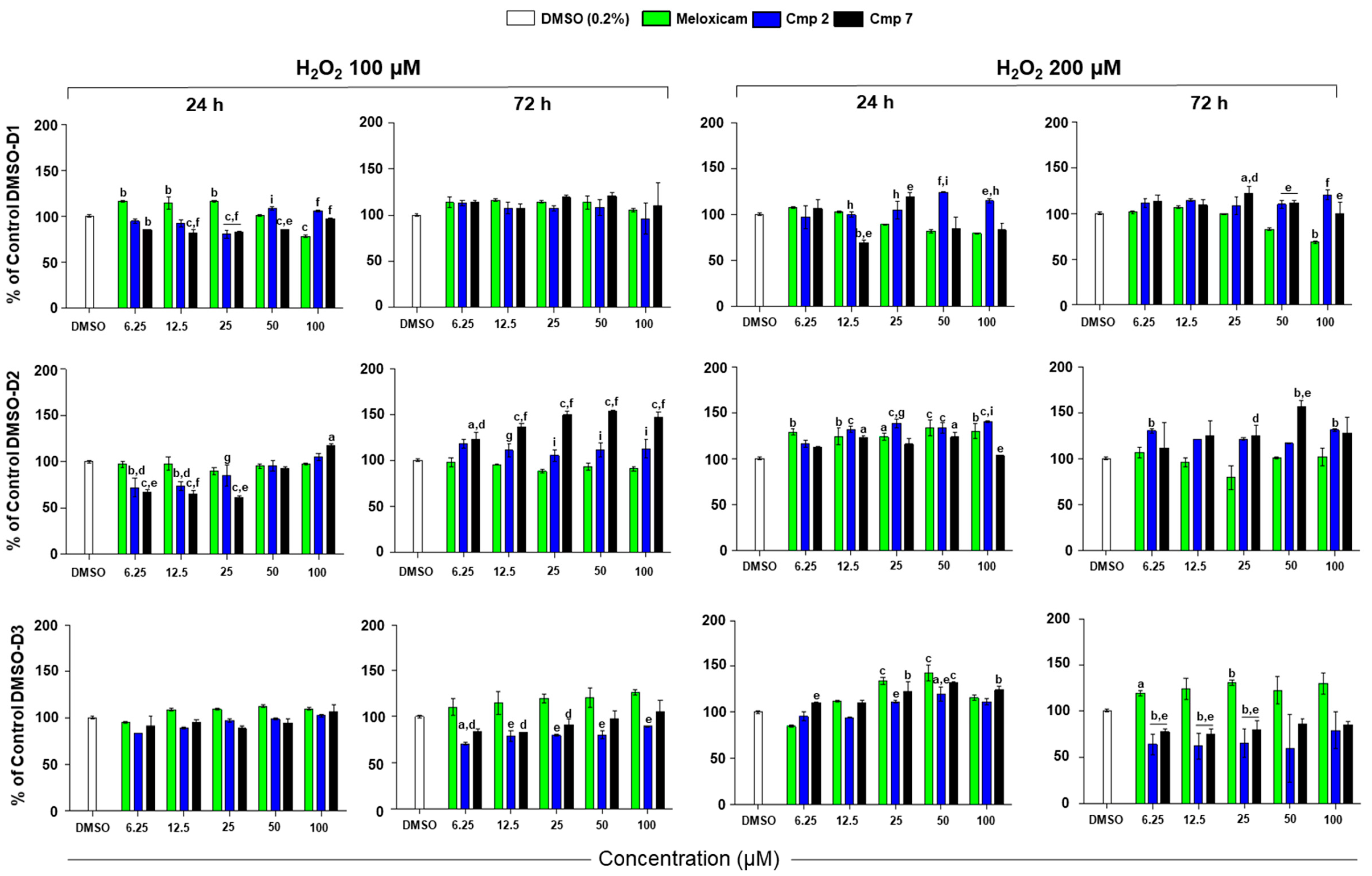
3.2. Cell Metabolic Activity of Tenocytes under H2O2 Stimulation in the Presence of Compounds
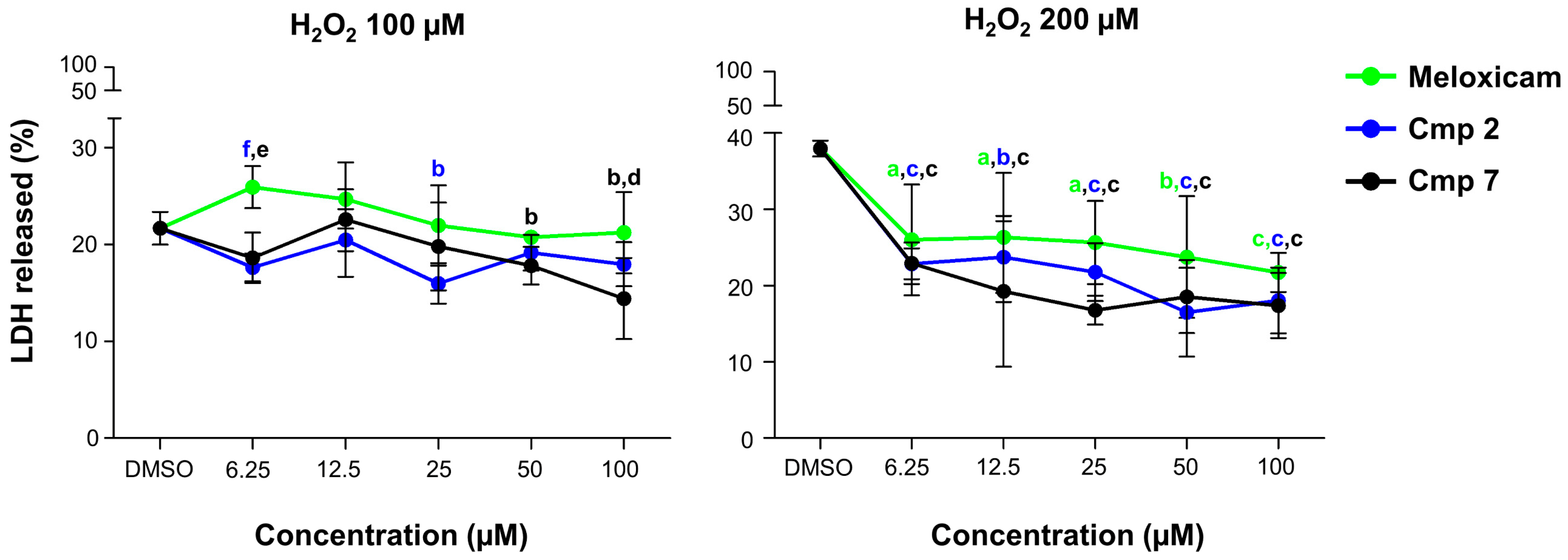
3.3. Counteraction of H2O2-Induced Cytotoxicity
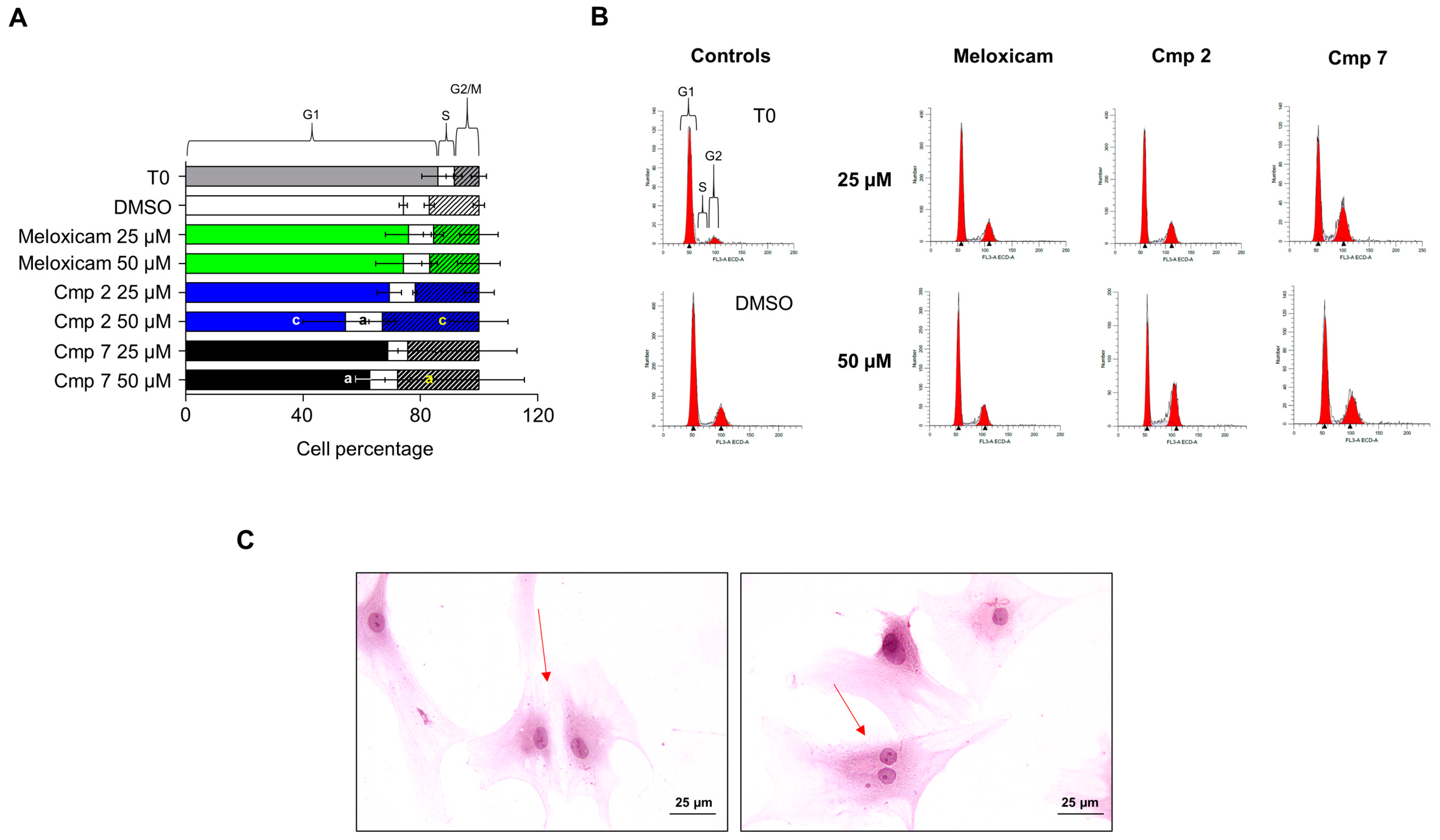
3.4. Compounds 2 and 7 Increase the Entrance in the G2 Phase of Tenocytes under Oxidative Stress Conditions
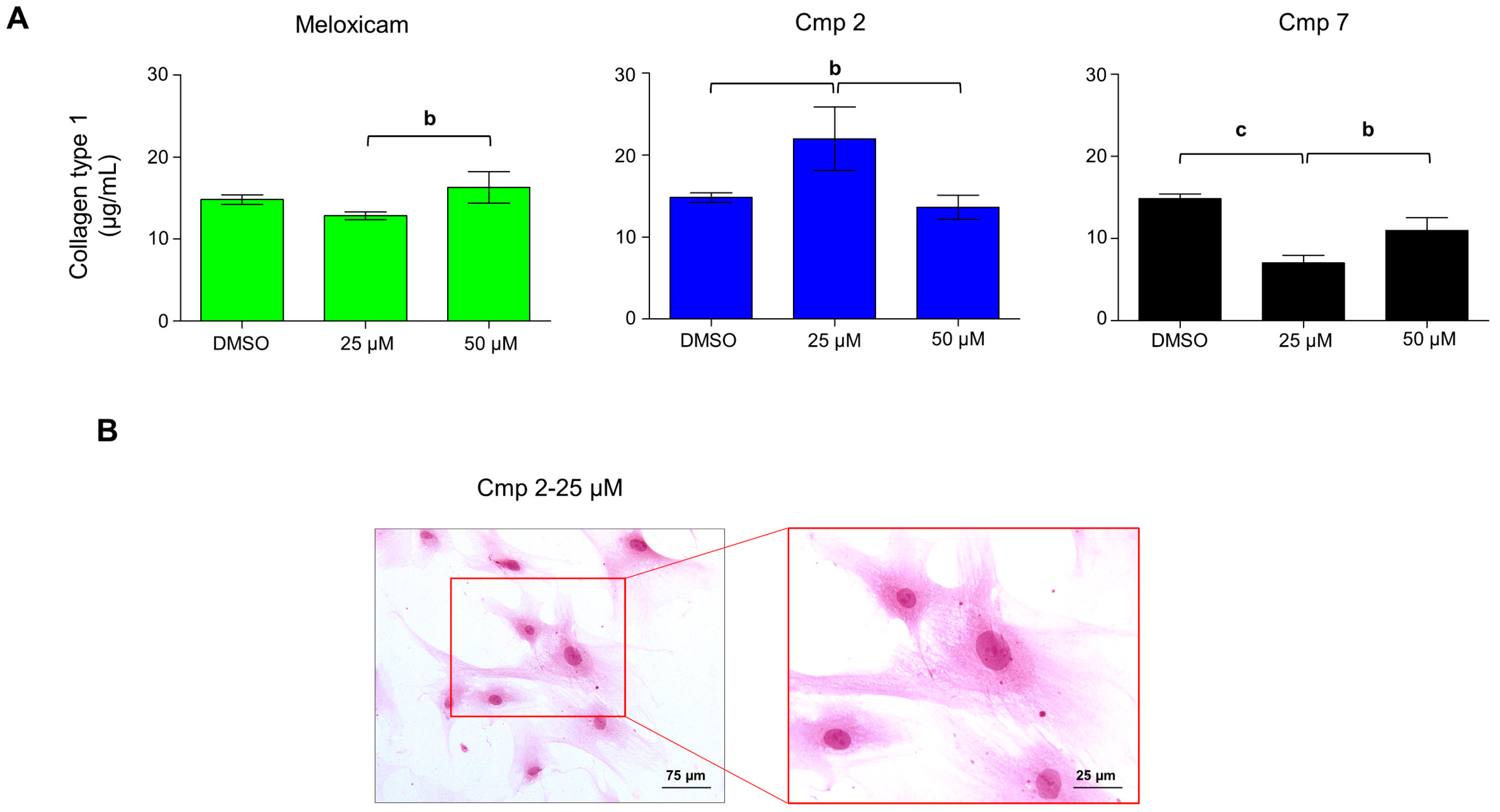
3.5. Compound 2 Enhances Collagen Type I Secretion in H2O2-Stimulated Tenocytes
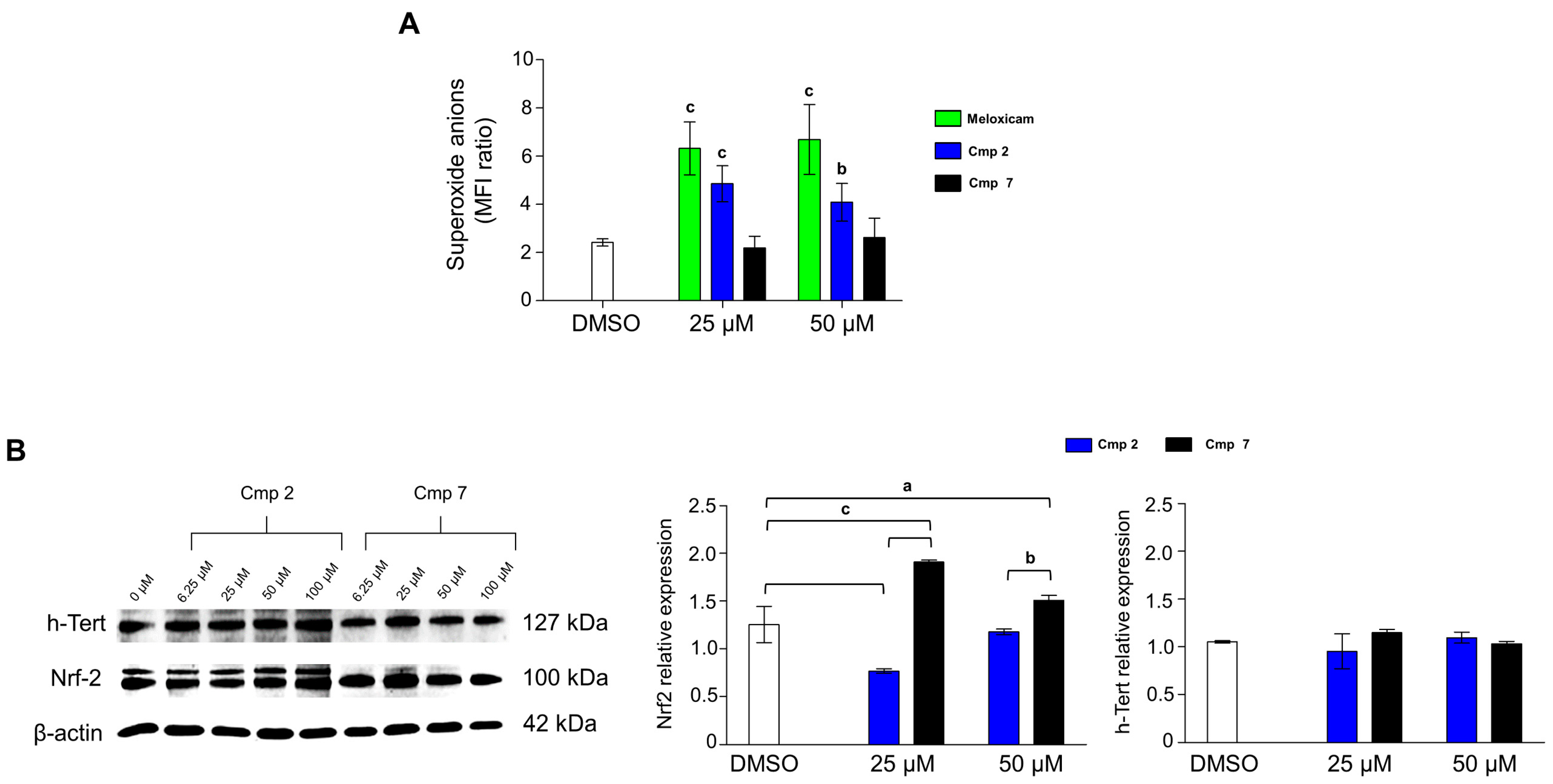
3.6. Compound 7 Triggers Nrf2 Expression and Acts as a Superoxide Scavenger
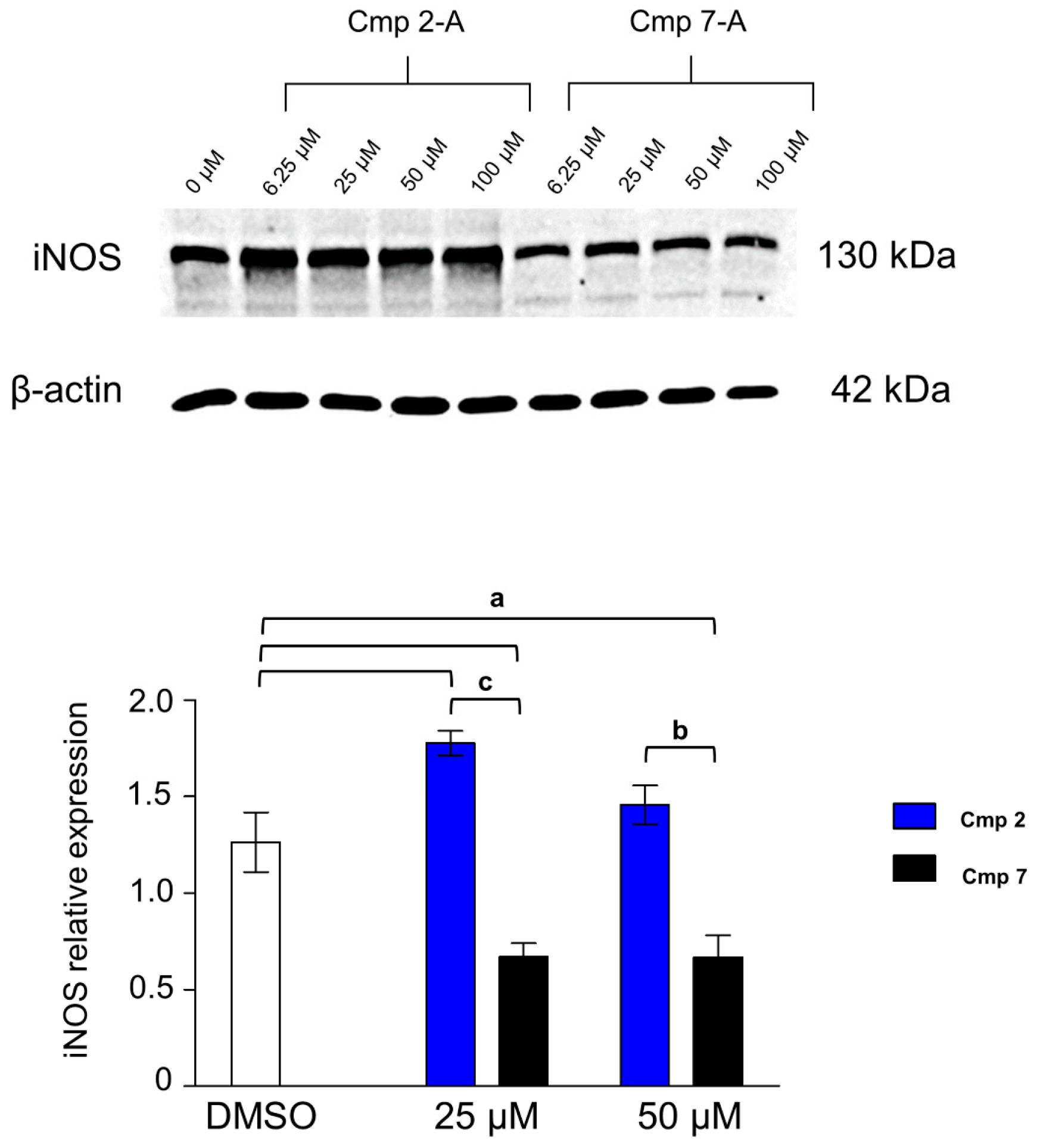
3.7. iNOS Is Differentially Expressed in the Presence of Compounds 2 and 7
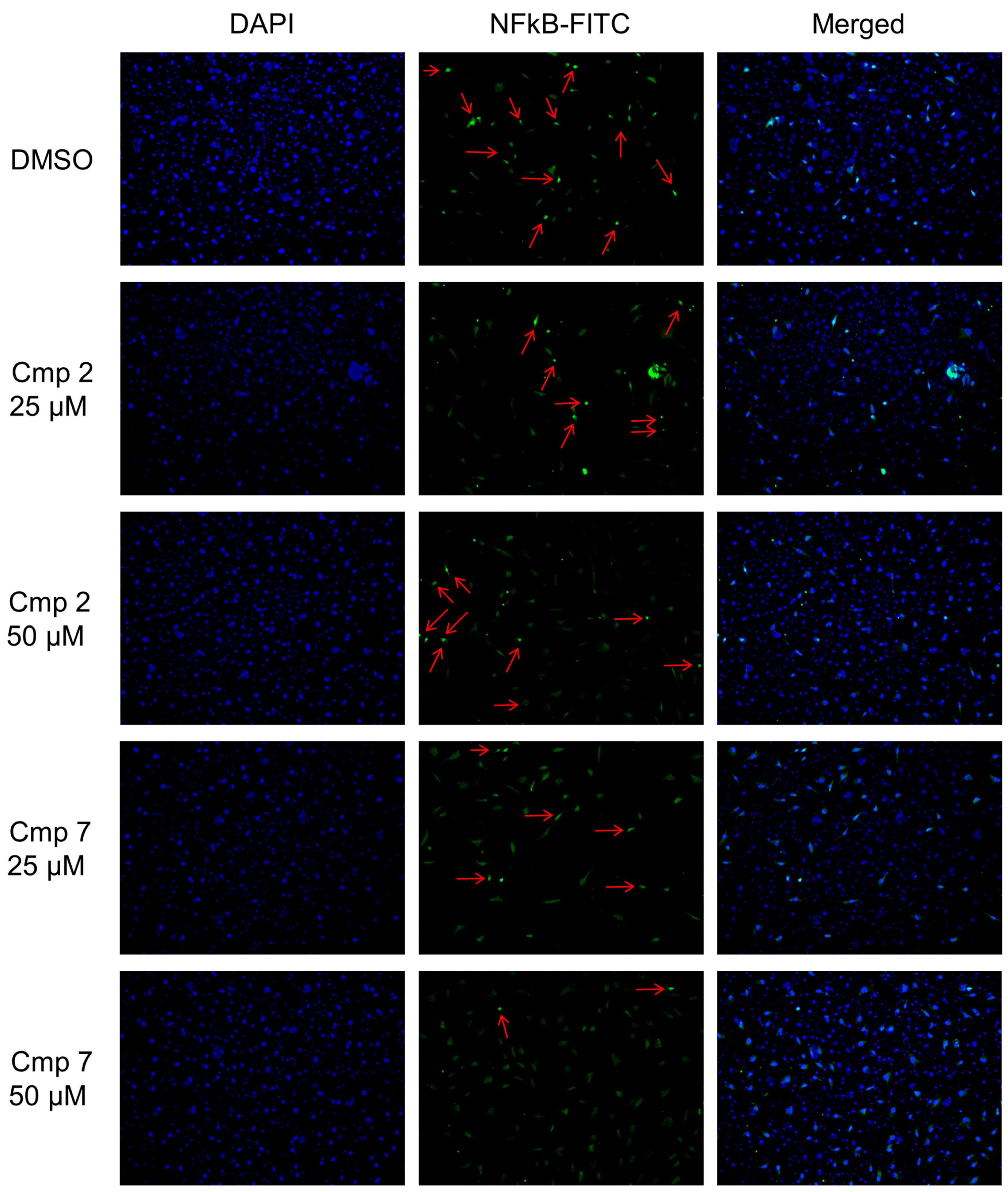
3.8. NF-kB Nuclear Translocation Is Enhanced in the Presence of Compound 2
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oliva, F.; Osti, L.; Padulo, J.; Maffulli, N. Epidemiology of the rotator cuff tears: A new incidence related to thyroid disease. Muscles Ligaments Tendons J. 2014, 4, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Cipollaro, L.; Sahemey, R.; Oliva, F.; Maffulli, N. Immunohistochemical features of rotator cuff tendinopathy. Br. Med. Bull. 2019, 1, 105–123. [Google Scholar] [CrossRef] [PubMed]
- Oliva, F.; Berardi, A.C.; Misiti, S.; Verga Falzacappa, C.; Iacone, A.; Maffulli, N. Thyroid hormones enhance growth and counteract apoptosis in human tenocytes isolated from rotator cuff tendons. Cell Death Dis. 2013, 7, e705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliva, F.; Gallorini, M.; Antonetti Lamorgese Passeri, C.; Gissi, C.; Ricci, A.; Cataldi, A.; Colosimo, A.; Berardi, A.C. Conjugation with Methylsulfonylmethane Improves Hyaluronic Acid Anti-Inflammatory Activity in a Hydrogen Peroxide-Exposed Tenocyte Culture In Vitro Model. Int. J. Mol. Sci. 2020, 21, 7956. [Google Scholar] [CrossRef]
- Gallorini, M.; Berardi, A.C.; Gissi, C.; Cataldi, A.; Osti, L. Nrf2-mediated cytoprotective effect of four different hyaluronic acids by molecular weight in human tenocytes. J. Drug Target. 2020, 2, 212–224. [Google Scholar] [CrossRef]
- Darrieutort-Laffite, C.; Soslowsky, L.J.; Le Goff, B. Molecular and Structural Effects of Percutaneous Interventions in Chronic Achilles Tendinopathy. Int. J. Mol. Sci. 2020, 19, 7000. [Google Scholar] [CrossRef]
- Gallorini, M.; Petzel, C.; Bolay, C.; Hiller, K.A.; Cataldi, A.; Buchalla, W.; Krifka, S.; Schweikl, H. Activation of the Nrf2-regulated antioxidant cell response inhibits HEMA-induced oxidative stress and supports cell viability. Biomaterials 2015, 56, 114–128. [Google Scholar] [CrossRef]
- Adach, W.; Olas, B. A comparison of multifunctional donors of carbon monoxide: Their anticoagulant, antioxidant, anti-aggregatory and cytotoxicity activities in an in vitro model. Nitric Oxide 2020, 97, 20–26. [Google Scholar] [CrossRef]
- Ryter, S.W. Therapeutic Potential of Heme Oxygenase-1 and Carbon Monoxide in Acute Organ Injury, Critical Illness, and Inflammatory Disorders. Antioxidants 2020, 11, 1153. [Google Scholar] [CrossRef]
- Wilson, J.L.; Fayad Kobeissi, S.; Oudir, S.; Haas, B.; Michel, B.; Dubois Randé, J.L.; Ollivier, A.; Martens, T.; Rivard, M.; Motterlini, R.; et al. Design and synthesis of new hybrid molecules that activate the transcription factor Nrf2 and simultaneously release carbon monoxide. Chemistry 2014, 45, 14698–14704. [Google Scholar] [CrossRef]
- Berrino, E.; Milazzo, L.; Micheli, L.; Vullo, D.; Angeli, A.; Bozdag, M.; Nocentini, A.; Menicatti, M.; Bartolucci, G.; di Cesare Mannelli, L.; et al. Synthesis and Evaluation of Carbonic Anhydrase Inhibitors with Carbon Monoxide Releasing Properties for the Management of Rheumatoid Arthritis. J. Med. Chem. 2019, 15, 7233–7249. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Carbonic anhydrases and metabolism. Metabolites 2018, 8, E25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, M.; Jin, W.; Waheed, A.; Moed, B.R.; Sly, W.; Zhang, Z. Expression profile of carbonic anhydrases in articular cartilage. Histochem. Cell Biol. 2011, 136, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Parkkila, S.; Shibata, S.; Fujimiya, M.; Murakami, G.; Cho, B.H. Expression of carbonic anhydrase IX in human fetal joints, ligaments and tendons: A potential marker of mechanical stress in fetal development? Anat. Cell Biol. 2013, 46, 272–284. [Google Scholar] [CrossRef] [PubMed]
- Margheri, F.; Ceruso, M.; Carta, F.; Laurenzana, A.; Maggi, L.; Lazzeri, S.; Simonini, G.; Annunziato, F.; Del Rosso, M.; Supuran, C.T.; et al. Overexpression of the transmembrane carbonic anhydrase isoforms IX and XII in the inflamed synovium. J. Enzyme Inhib. Med. Chem. 2016, 31, 60–63. [Google Scholar] [CrossRef] [Green Version]
- Ji, M.J.; Hong, J.H. An overview of carbonic anhydrases and membrane channels of synoviocytes in inflamed joints. J. Enzyme Inhib. Med. Chem. 2019, 34, 1615–1622. [Google Scholar] [CrossRef] [Green Version]
- Berrino, E.; Carradori, S.; Angeli, A.; Carta, F.; Supuran, C.T.; Guglielmi, P.; Coletti, C.; Paciotti, R.; Schweikl, H.; Maestrelli, F.; et al. Dual carbonic anhydrase IX/XII inhibitors and Carbon Monoxide Releasing Molecules modulate LPS-mediated inflammation in mouse macrophages. Antioxidants 2021, 10, 1–24. [Google Scholar] [CrossRef]
- Guglielmi, P.; Rotondi, G.; Secci, D.; Angeli, A.; Chimenti, P.; Nocentini, A.; Bonardi, A.; Gratteri, P.; Carradori, S.; Supuran, C.T. Novel insights on saccharin- and acesulfame-based carbonic anhydrase inhibitors: Design, synthesis, modelling investigations and biological activity evaluation. J. Enzyme Inhib Med. Chem. 2020, 35, 1891–1905. [Google Scholar] [CrossRef]
- Aspatwar, A.; Berrino, E.; Bua, S.; Carta, F.; Capasso, C.; Parkkila, S.; Supuran, C.T. Toxicity evaluation of sulfamides and coumarins that efficiently inhibit human carbonic anhydrases. J. Enzyme Inhib. Med. Chem. 2020, 35, 1765–1772. [Google Scholar] [CrossRef]
- Schattenkirchner, M. Meloxicam: A selective COX-2 inhibitor non-steroidal anti-inflammatory drug. Expert Opin. Investig. Drugs 1997, 6, 321–334. [Google Scholar] [CrossRef]
- Osti, L.; Berardocco, M.; di Giacomo, V.; Di Bernardo, G.; Oliva, F.; Berardi, A.C. Hyaluronic acid increases tendon derived cell viability and collagen type I expression in vitro: Comparative study of four different Hyaluronic acid preparations by molecular weight. BMC Musculoskelet Disord. 2015, 16, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallorini, M.; Berardi, A.C.; Berardocco, M.; Gissi, C.; Maffulli, N.; Cataldi, A.; Oliva, F. Hyaluronic acid increases tendon derived cell viability and proliferation in vitro: Comparative study of two different hyaluronic acid preparations by molecular weight. Muscles Ligaments Tendons J. 2017, 7, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Gissi, C.; Radeghieri, A.; Antonetti Lamorgese Passeri, C.; Gallorini, M.; Calciano, L.; Oliva, F.; Veronesi, F.; Zendrini, A.; Cataldi, A.; Bergese, P.; et al. Extracellular vesicles from rat-bone-marrow mesenchymal stromal/stem cells improve tendon repair in rat Achilles tendon injury model in dose-dependent manner: A pilot study. PLoS ONE 2020, 15, e0229914. [Google Scholar] [CrossRef] [PubMed]
- Frich, L.H.; Fernandes, L.R.; Schrøder, H.D.; Hejbøl, E.K.; Nielsen, P.V.; Jørgensen, P.H.; Stensballe, A.; Lambertsen, K.L. The inflammatory response of the supraspinatus muscle in rotator cuff tear conditions. J. Shoulder Elb. Surg. 2020, 20, 30710–30712. [Google Scholar] [CrossRef]
- Dakin, S.G.; Newton, J.; Martinez, F.O.; Hedley, R.; Gwilym, S.; Jones, N.; Reid, H.A.B.; Wood, S.; Wells, G.; Appleton, L.; et al. Chronic inflammation is a feature of Achilles tendinopathy and rupture. Br. J. Sports Med. 2018, 52, 359–367. [Google Scholar] [CrossRef]
- Benhar, M. Oxidants, Antioxidants and Thiol Redox Switches in the Control of Regulated Cell Death Pathways. Antioxidants 2020, 9, 309. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Maffulli, N. Biology of tendon injury: Healing, modeling and remodeling. J. Musculoskelet Neuronal Interact. 2006, 6, 181–190. [Google Scholar]
- Figueiredo-Pereira, C.; Dias-Pedroso, D.; Soares, N.L.; Vieira, H.L.A. CO-mediated cytoprotection is dependent on cell metabolism modulation. Redox Biol. 2020, 32, 101470. [Google Scholar] [CrossRef]
- Yuda, A.; Lee, W.S.; Petrovic, P.; McCulloch, C.A. Novel proteins that regulate cell extension formation in fibroblasts. Exp. Cell Res. 2018, 365, 85–96. [Google Scholar] [CrossRef]
- Russo, V.; Mauro, A.; Martelli, A.; Di Giacinto, O.; Di Marcantonio, L.; Nardinocchi, D.; Berardinelli, P.; Barboni, B. Cellular and molecular maturation in fetal and adult ovine calcaneal tendons. J. Anat. 2015, 226, 126–142. [Google Scholar] [CrossRef] [Green Version]
- Villegas, I.; Martín, M.J.; La Casa, C.; Motilva, V.; De La Lastra, C.A. Effects of oxicam inhibitors of cyclooxygenase on oxidative stress generation in rat gastric mucosa. A comparative study. Free. Radic. Res. 2002, 36, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Feng, H.; Lv, J.; Zhao, L.; Zhao, J.; Wang, L.A. Protective effect of coumarin-pi against t-BHP-induced hepatotoxicity by upregulating antioxidant enzymes via enhanced Nrf2 signaling. Mol. Cell Biochem. 2020, 475, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Bokhari, A.R.; Murrell, G.A. The role of nitric oxide in tendon healing. J. Shoulder Elb. Surg. 2012, 21, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Szomor, Z.; Wang, Y.; Murrell, G.A.C. Nitric oxide enhances collagen synthesis in cultured human tendon cells. J. Orthop. Res. 2006, 24, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Millar, N.L.; Murrell, G.A.C.; McInnes, I.B. Inflammatory mechanisms in tendinopathy—Towards translation. Nat. Rev. Rheumatol. 2017, 13, 110–122. [Google Scholar] [CrossRef]
- Swayampakula, M.; McDonald, P.C.; Vallejo, M.; Coyaud, E.; Chafe, S.C.; Westerback, A.; Venkateswaran, G.; Shankar, J.; Gao, G.; Laurent, E.M.N.; et al. The interactome of metabolic enzyme carbonic anhydrase IX reveals novel roles in tumor cell migration and invadopodia/MMP14-mediated invasion. Oncogene 2017, 36, 6244–6261. [Google Scholar] [CrossRef] [Green Version]
- Morgan, M.J.; Liu, Z.G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Schweikl, H.; Birke, M.; Gallorini, M.; Petzel, C.; Bolay, C.; Waha, C.; Hiller, K.A.; Buchalla, W. HEMA-induced oxidative stress inhibits NF-kB nuclear translocation and TNF release from LTA- and LPS-stimulated immunocompetent cells. Dental. Mater. 2021, 37, 175–190. [Google Scholar] [CrossRef]
- Sancilio, S.; Marsich, E.; Schweikl, H.; Cataldi, A.; Gallorini, M. Redox Control of IL-6-Mediated Dental Pulp Stem-Cell Differentiation on Alginate/Hydroxyapatite Biocomposites for Bone Ingrowth. Nanomaterials. 2019, 9, 1656. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallorini, M.; Berardi, A.C.; Ricci, A.; Antonetti Lamorgese Passeri, C.; Zara, S.; Oliva, F.; Cataldi, A.; Carta, F.; Carradori, S. Dual Acting Carbon Monoxide Releasing Molecules and Carbonic Anhydrase Inhibitors Differentially Modulate Inflammation in Human Tenocytes. Biomedicines 2021, 9, 141. https://doi.org/10.3390/biomedicines9020141
Gallorini M, Berardi AC, Ricci A, Antonetti Lamorgese Passeri C, Zara S, Oliva F, Cataldi A, Carta F, Carradori S. Dual Acting Carbon Monoxide Releasing Molecules and Carbonic Anhydrase Inhibitors Differentially Modulate Inflammation in Human Tenocytes. Biomedicines. 2021; 9(2):141. https://doi.org/10.3390/biomedicines9020141
Chicago/Turabian StyleGallorini, Marialucia, Anna C. Berardi, Alessia Ricci, Cristina Antonetti Lamorgese Passeri, Susi Zara, Francesco Oliva, Amelia Cataldi, Fabrizio Carta, and Simone Carradori. 2021. "Dual Acting Carbon Monoxide Releasing Molecules and Carbonic Anhydrase Inhibitors Differentially Modulate Inflammation in Human Tenocytes" Biomedicines 9, no. 2: 141. https://doi.org/10.3390/biomedicines9020141