Withanolide Metabolites Inhibit PI3K/AKT and MAPK Pro-Survival Pathways and Induce Apoptosis in Acute Myeloid Leukemia Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Viability Assays
2.3. Apoptosis Assay
2.4. Western Blot Analysis of Apoptosis Markers
2.5. Proteome Analysis
2.5.1. Cell Lysis and Protein Extraction
2.5.2. Digestion
2.5.3. Desalting
2.5.4. TiO2 Phosphopeptide Enrichment
2.5.5. Nanoflow-Liquid Chromatography Tandem Mass Spectrometry (LC–MS/MS)
2.5.6. MS Data Analysis
3. Results
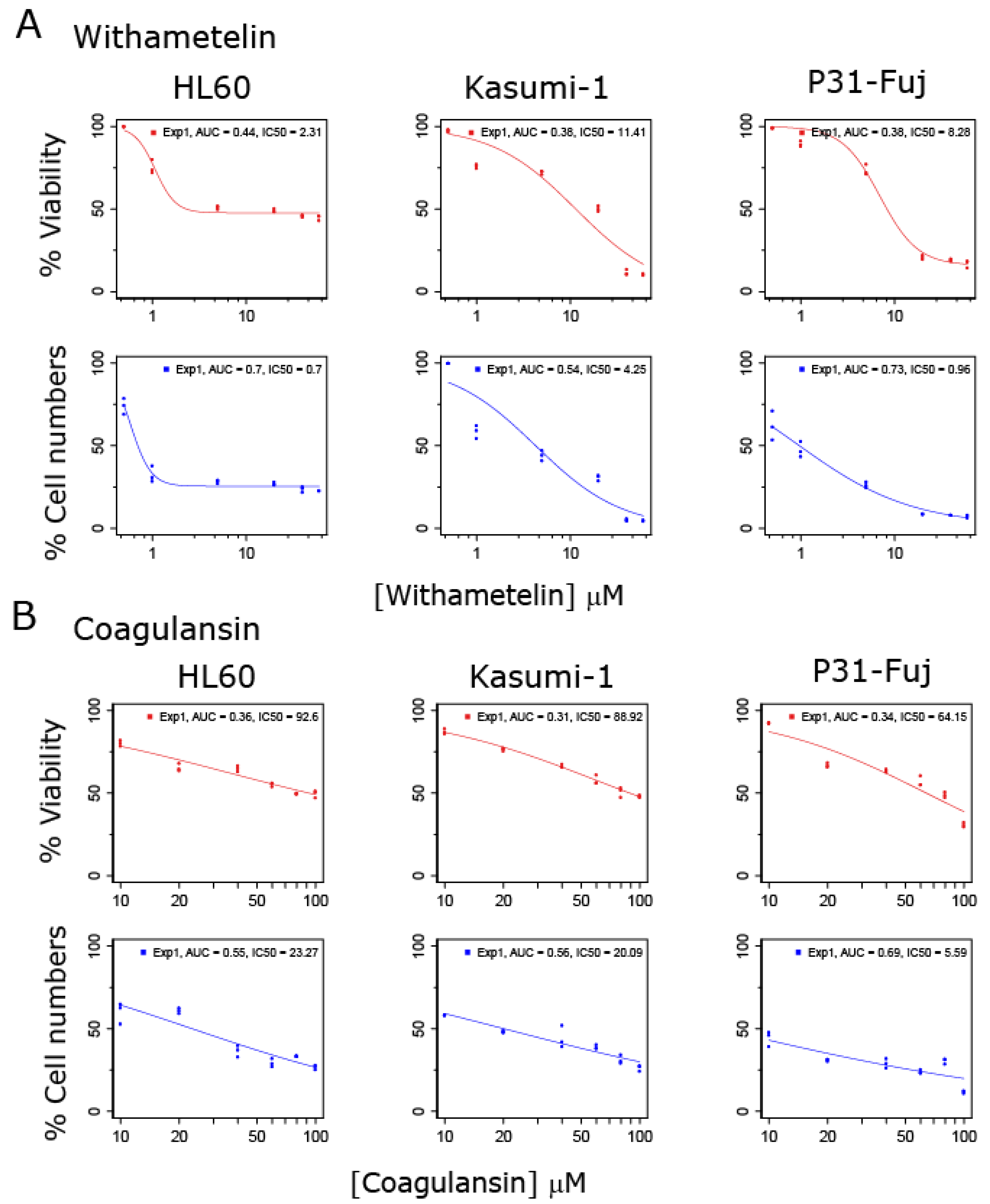
3.1. Withametelin and Coagulansin A Inhibit the Proliferation of AML Models
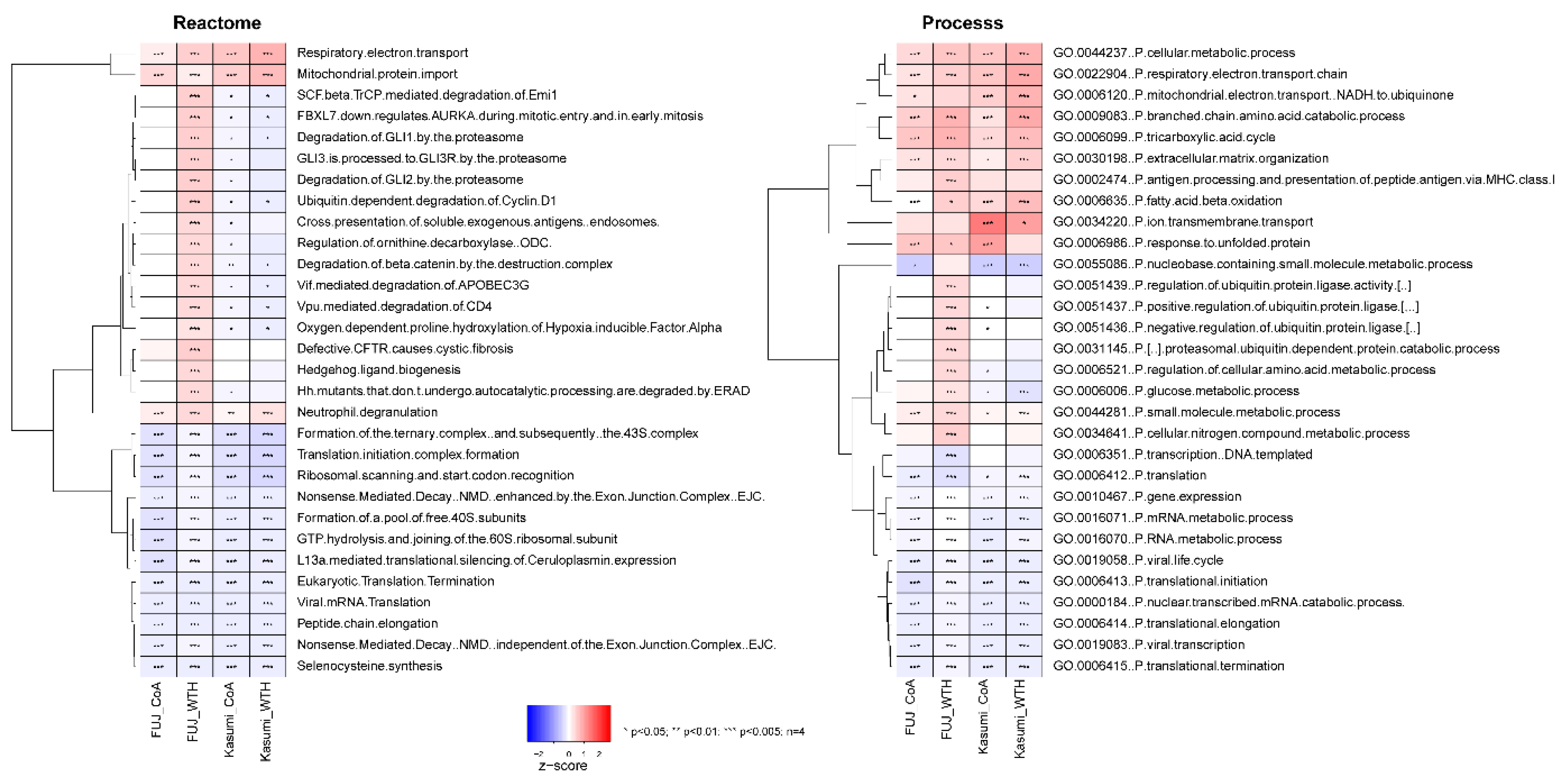
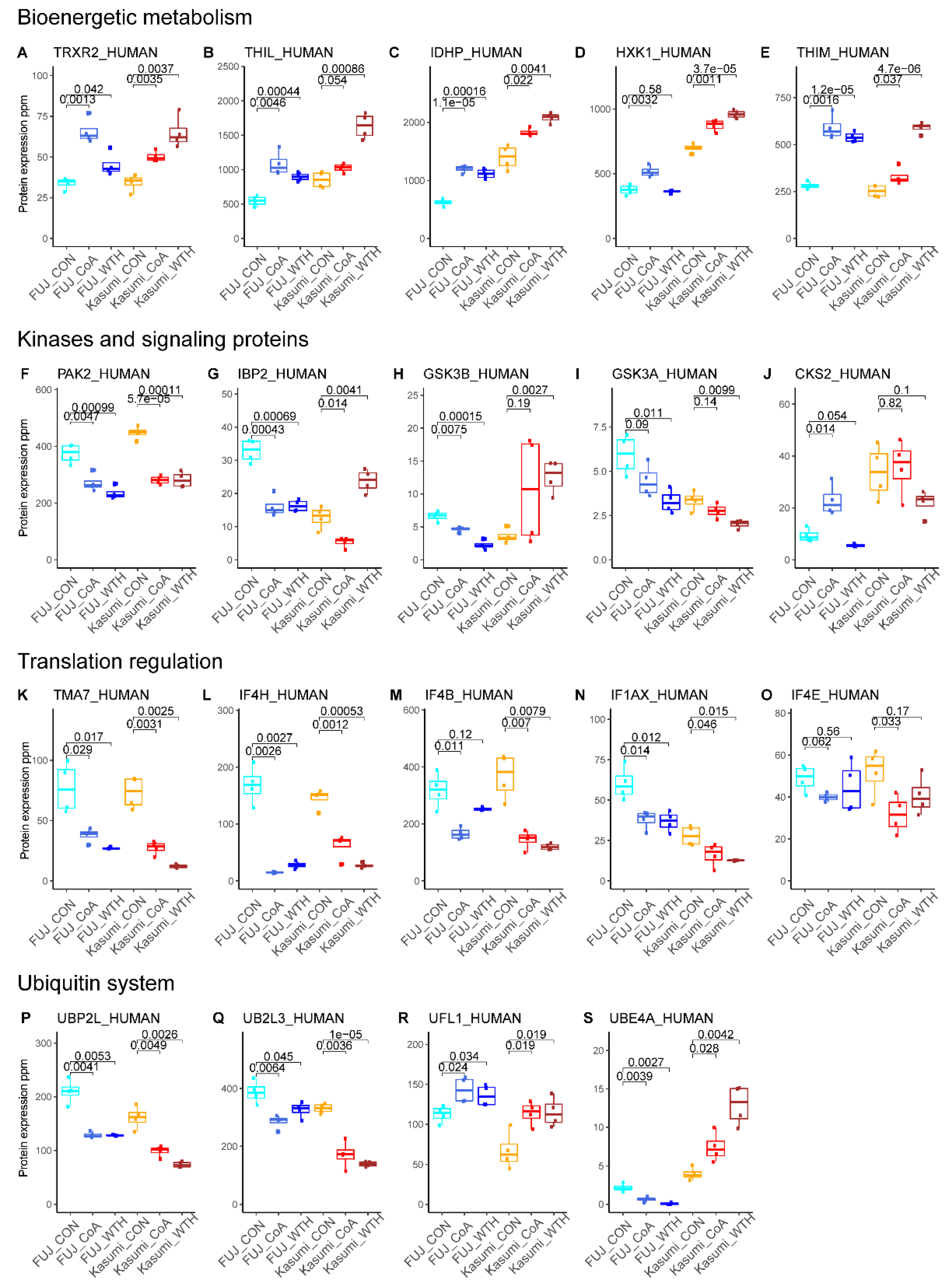
3.2. Withametelin and Coagulansin A Modulate Pathways Involved in Bioenergetic Metabolism, Signal Transduction and Regulation of Gene Expression
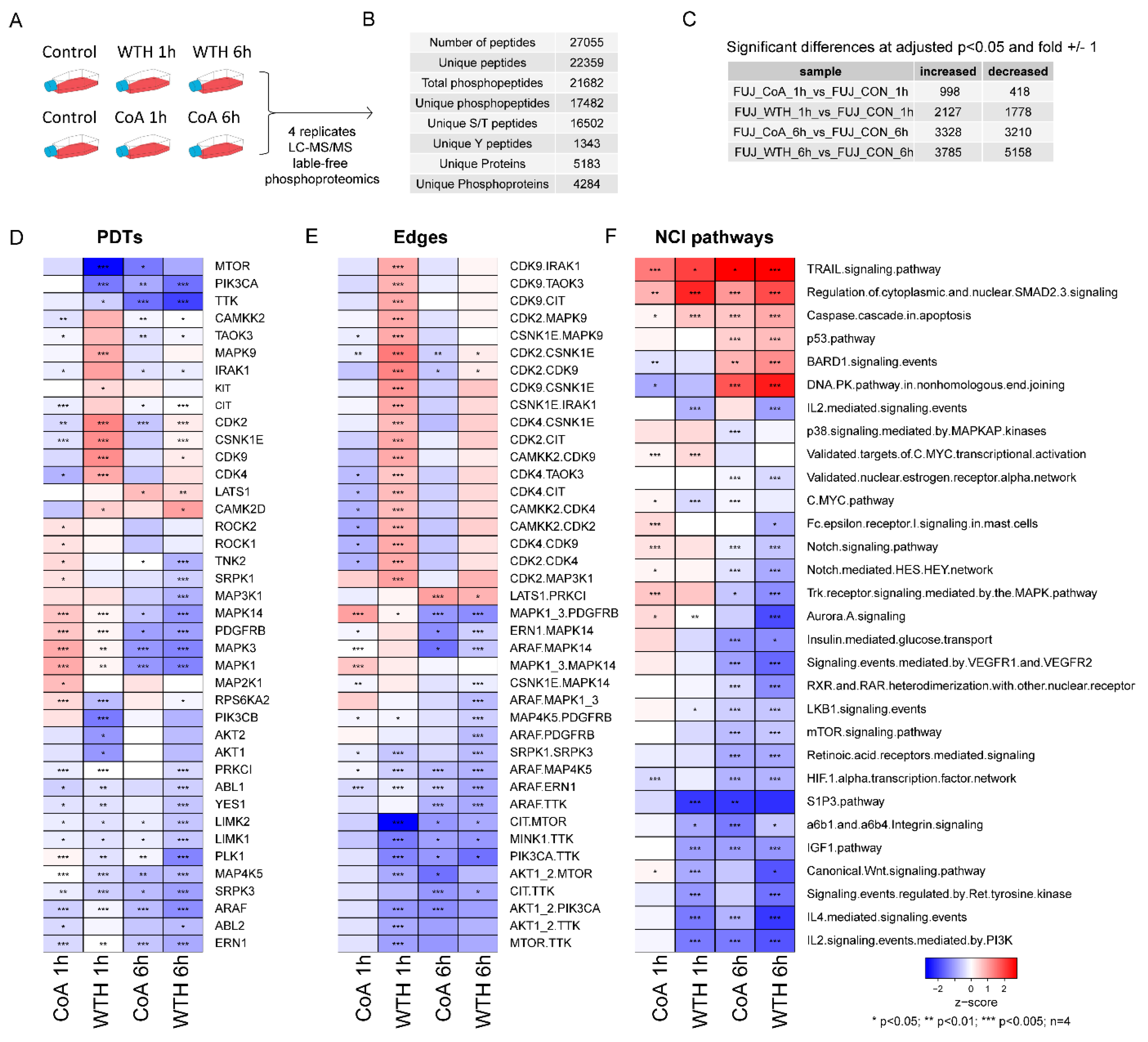
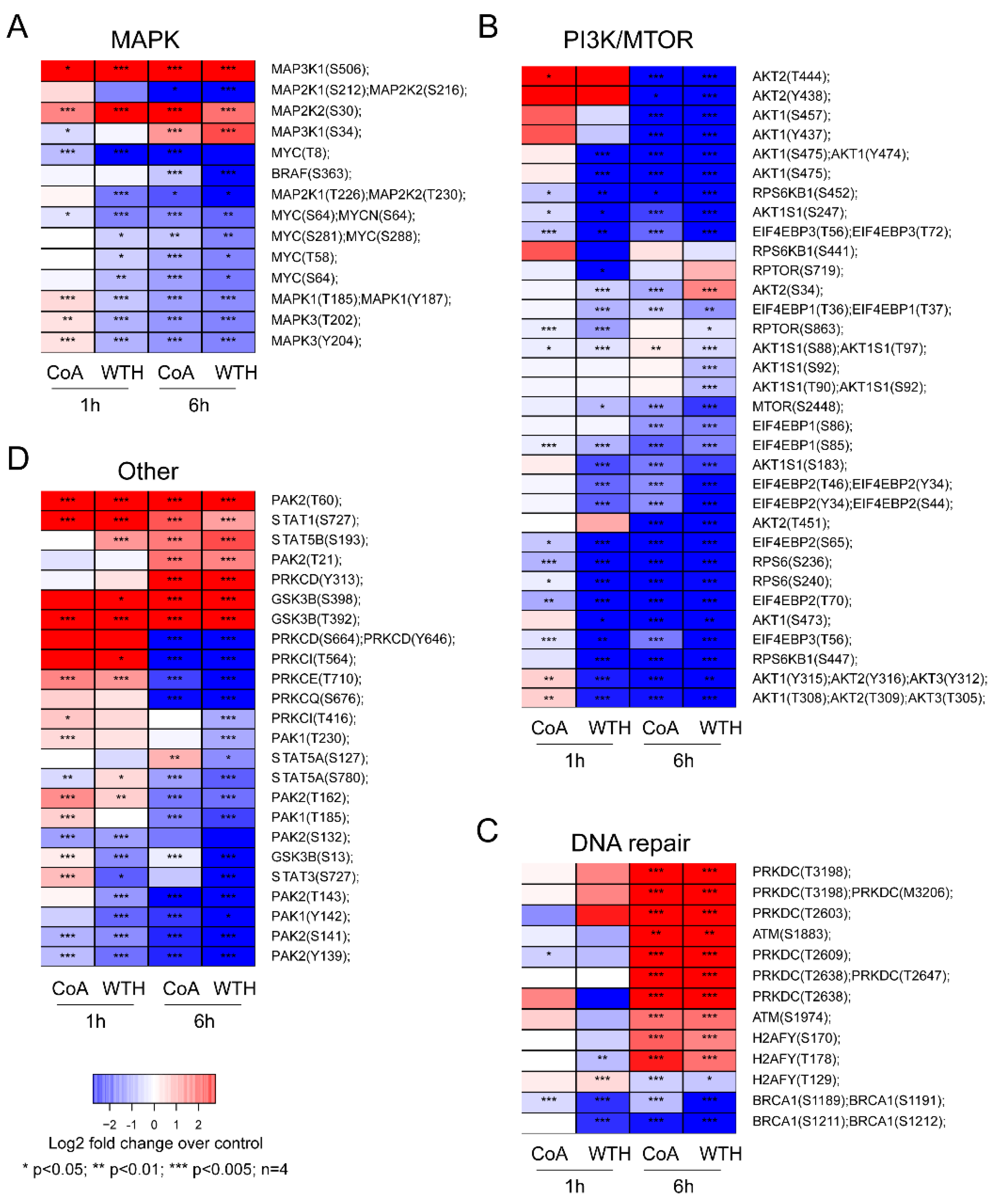
3.3. Phosphoproteomic Analysis Reveals That Withametelin and Coagulansin A Have an Impact on the Activity of Pro-Survival Kinase Signaling
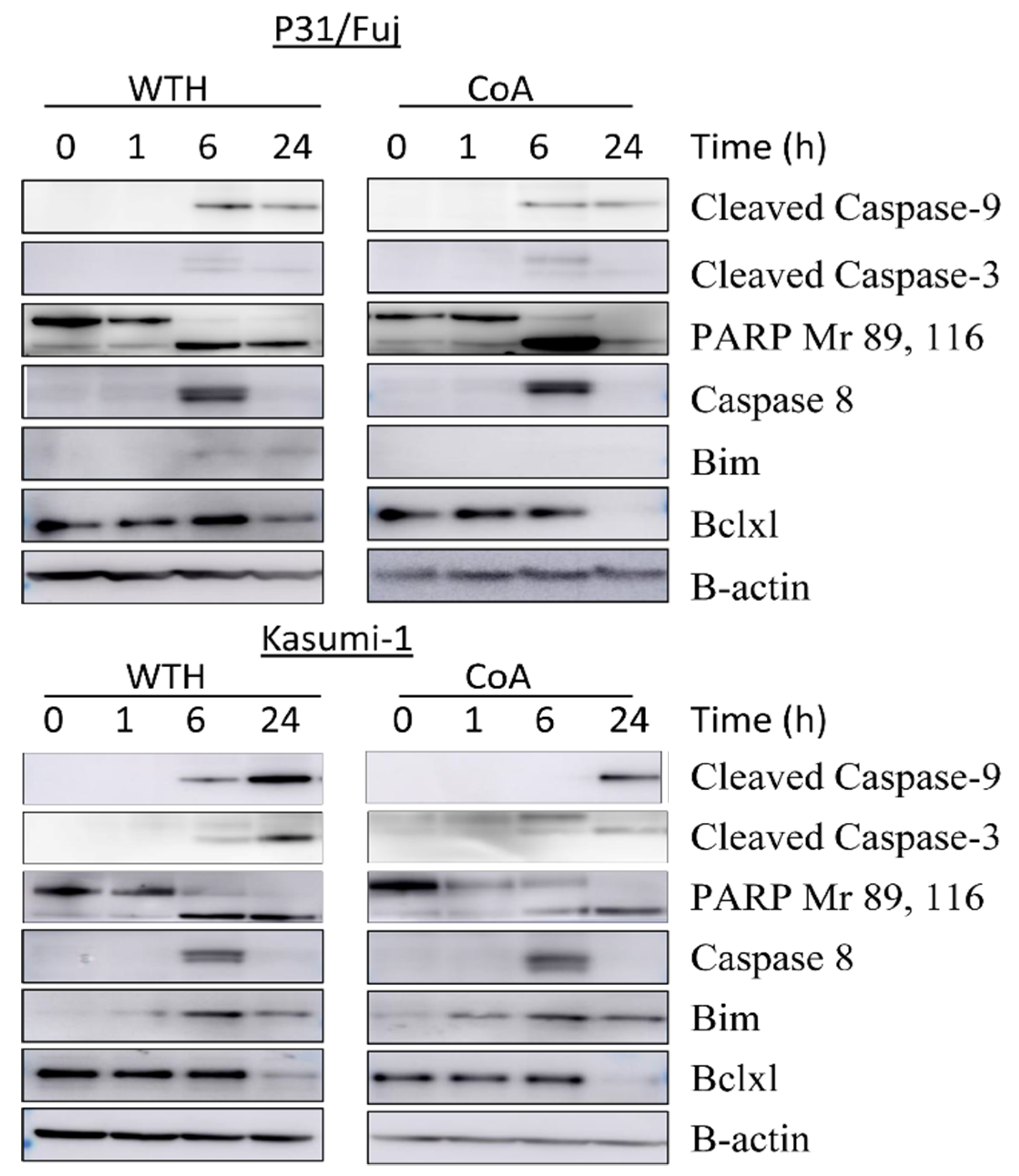
3.4. Withametelin and Cagulansin A Induce Apoptosis in AML Cells
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Tawana, K.; Bödör, C.; Cavenagh, J.; Jenner, M.; Fitzgibbon, J. The molecular pathogenesis of acute myeloid leukemia. Curr. Med. Lit. 2013, 21, 67. [Google Scholar]
- Howlader, N.; Noone, A.; Krapcho, M.; Miller, D.; Brest, A.; Yu, M.; Ruhl, J.; Tatalovich, Z.; Mariotto, A.; Lewis, D. SEER Cancer Statistics Review, 1975–2016, 2019. Available online: https://seer.cancer.gov/archive/csr/1975_2016/ (accessed on 6 September 2020).
- Deschler, B.; Lübbert, M. Acute myeloid leukemia: Epidemiology and etiology. Cancer Interdiscip. Int. J. Am. Cancer Soc. 2006, 107, 2099–2107. [Google Scholar] [CrossRef] [PubMed]
- Chamani, E.; Ebrahimi, R.; Khorsandi, K.; Meshkini, A.; Zarban, A.; Sharifzadeh, G. In vitro cytotoxicity of polyphenols from Datura innoxia aqueous leaf-extract on human leukemia K562 cells: DNA and nuclear proteins as targets. Drug Chem. Toxicol. 2020, 43, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Stone, R.M.; Mandrekar, S.J.; Sanford, B.L.; Laumann, K.; Geyer, S.; Bloomfield, C.D.; Thiede, C.; Prior, T.W.; Dohner, K.; Marcucci, G.; et al. Midostaurin plus Chemotherapy for Acute Myeloid Leukemia with a FLT3 Mutation. N. Engl. J. Med. 2017, 377, 454–464. [Google Scholar] [CrossRef]
- Quek, L.; David, M.D.; Kennedy, A.; Metzner, M.; Amatangelo, M.; Shih, A.; Stoilova, B.; Quivoron, C.; Heiblig, M.; Willekens, C.; et al. Clonal heterogeneity of acute myeloid leukemia treated with the IDH2 inhibitor enasidenib. Nat. Med. 2018, 24, 1167–1177. [Google Scholar] [CrossRef]
- Roboz, G.J.; DiNardo, C.D.; Stein, E.M.; de Botton, S.; Mims, A.S.; Prince, G.T.; Altman, J.K.; Arellano, M.L.; Donnellan, W.; Erba, H.P.; et al. Ivosidenib induces deep durable remissions in patients with newly diagnosed IDH1-mutant acute myeloid leukemia. Blood 2020, 135, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Bower, H.; Andersson, T.M.; Björkholm, M.; Dickman, P.; Lambert, P.; Derolf, Å.R. Continued improvement in survival of acute myeloid leukemia patients: An application of the loss in expectation of life. Blood Cancer J. 2016, 6, e390. [Google Scholar] [CrossRef] [Green Version]
- DiNardo, C.D.; Wei, A.H. How I treat acute myeloid leukemia in the era of new drugs. Blood 2020, 135, 85–96. [Google Scholar] [CrossRef]
- Intlekofer, A.M.; Shih, A.H.; Wang, B.; Nazir, A.; Rustenburg, A.S.; Albanese, S.K.; Patel, M.; Famulare, C.; Correa, F.M.; Takemoto, N.; et al. Acquired resistance to IDH inhibition through trans or cis dimer-interface mutations. Nature 2018, 559, 125–129. [Google Scholar] [CrossRef]
- Cragg, G.M.; Pezzuto, J.M. Natural products as a vital source for the discovery of cancer chemotherapeutic and chemopreventive agents. Med. Princ. Pract. 2016, 25, 41–59. [Google Scholar] [CrossRef]
- Hwang, D.; Kim, M.; Park, H.; Jeong, M.I.; Jung, W.; Kim, B. Natural products and acute myeloid leukemia: A review highlighting mechanisms of action. Nutrients 2019, 11, 1010. [Google Scholar] [CrossRef] [Green Version]
- Roomi, M.; Kalinovsky, T.; Roomi, N.; Niedzwiecki, A.; Rath, M. In vitro and in vivo inhibition of human Fanconi anemia head and neck squamous carcinoma by a phytonutrient combination. Int. J. Oncol. 2015, 46, 2261–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, K.H.; Barve, A.; Yu, S.; Huang, M.T.; Kong, A.N.T. Cancer chemoprevention by phytochemicals: Potential molecular targets, biomarkers and animal models 1. Acta Pharmacol. Sin. 2007, 28, 1409–1421. [Google Scholar] [CrossRef] [PubMed]
- Nazeema, B.; Julie, J.; Abirami, J.; Kumareasan, R.; Muthukumaran, T.; Rajasree, S.; Jeya, J.; Kumaran, S. Anti-cancer activity of Datura metel on MCF-7 cell line. Asian J. Pharm. Clin. Res. 2014, 7, 181–183. [Google Scholar]
- Arulvasu, C.; Babu, G.; Manikandan, R.; Srinivasan, P.; Sellamuthu, S.; Prabhu, D.; Dinesh, D. Anti-cancer effect of Datura innoxia P. Mill. Leaf extract in vitro through induction of apoptosis in human Colon Adenocarcinoma and larynx cancer cell lines. J. Pharm. Res. 2010, 3, 1485–1488. [Google Scholar]
- Cheshomi, H.; Aldaghi, L.S.; Seresht, H.R. Cytotoxicity of the methanol extract of Datura innoxia petals on MCF-7 and HEK-293 cell lines. J. Biomed. 2016, 1, e6623. [Google Scholar] [CrossRef] [Green Version]
- Samadi, A.K. Potential anticancer properties and mechanisms of action of withanolides. In The Enzymes; Elsevier: Amsterdam, The Netherlands, 2015; Volume 37, pp. 73–94. [Google Scholar]
- Wang, H.-C.; Tsai, Y.-L.; Wu, Y.-C.; Chang, F.-R.; Liu, M.-H.; Chen, W.-Y.; Wu, C.-C. Withanolides-induced breast cancer cell death is correlated with their ability to inhibit heat protein 90. PLoS ONE 2012, 7, e37764. [Google Scholar] [CrossRef]
- Doepfner, K.T.; Boller, D.; Arcaro, A. Targeting receptor tyrosine kinase signaling in acute myeloid leukemia. Crit. Rev. Oncol. Hematol. 2007, 63, 215–230. [Google Scholar] [CrossRef] [Green Version]
- Hainaut, P.; Plymoth, A. Targeting the hallmarks of cancer: Towards a rational approach to next-generation cancer therapy. Curr. Opin. Oncol. 2013, 25, 50–51. [Google Scholar] [CrossRef]
- Borthakur, G.; Popplewell, L.; Boyiadzis, M.; Foran, J.; Platzbecker, U.; Vey, N.; Walter, R.B.; Olin, R.; Raza, A.; Giagounidis, A.; et al. Activity of the oral mitogen-activated protein kinase kinase inhibitor trametinib in RAS-mutant relapsed or refractory myeloid malignancies. Cancer 2016, 122, 1871–1879. [Google Scholar] [CrossRef] [Green Version]
- Ragon, B.K.; Odenike, O.; Baer, M.R.; Stock, W.; Borthakur, G.; Patel, K.; Han, L.; Chen, H.; Ma, H.; Joseph, L.; et al. Oral MEK 1/2 Inhibitor Trametinib in Combination With AKT Inhibitor GSK2141795 in Patients With Acute Myeloid Leukemia With RAS Mutations: A Phase II Study. Clin. Lymphoma Myeloma Leuk. 2019, 19, 431–440.e13. [Google Scholar] [CrossRef] [PubMed]
- Sandhofer, N.; Metzeler, K.H.; Rothenberg, M.; Herold, T.; Tiedt, S.; Groiss, V.; Carlet, M.; Walter, G.; Hinrichsen, T.; Wachter, O.; et al. Dual PI3K/mTOR inhibition shows antileukemic activity in MLL-rearranged acute myeloid leukemia. Leukemia 2015, 29, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Hijazi, M.; Smith, R.; Rajeeve, V.; Bessant, C.; Cutillas, P.R. Reconstructing kinase network topologies from phosphoproteomics data reveals cancer-associated rewiring. Nat. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Kong, Y.; Kejun, N.; Yin, Y. Identification and characterization of CAC1 as a novel CDK2-associated cullin. Cell Cycle 2009, 8, 3552–3561. [Google Scholar] [CrossRef] [PubMed]
- Loughran, G.; Healy, N.C.; Kiely, P.A.; Huigsloot, M.; Kedersha, N.L.; O’Connor, R. Mystique is a new insulin-like growth factor-I-regulated PDZ-LIM domain protein that promotes cell attachment and migration and suppresses Anchorage-independent growth. Mol. Biol. Cell 2005, 16, 1811–1822. [Google Scholar] [CrossRef] [Green Version]
- Fatima, H.; Khan, K.; Zia, M.; Ur-Rehman, T.; Mirza, B.; Haq, I.-u. Extraction optimization of medicinally important metabolites from Datura innoxia Mill: An in vitro biological and phytochemical investigation. BMC Complement. Altern. Med. 2015, 15, 376. [Google Scholar] [CrossRef] [Green Version]
- Youn, U.J.; Chai, X.; Park, E.-J.; Kondratyuk, T.P.; Simmons, C.J.; Borris, R.P.; Mirza, B.; Pezzuto, J.M.; Chang, L.C. Biologically active withanolides from Withania coagulans. J. Nat. Prod. 2013, 76, 22–28. [Google Scholar]
- Mondal, S.; Mandal, C.; Sangwan, R.; Chandra, S.; Mandal, C. Withanolide D induces apoptosis in leukemia by targeting the activation of neutral sphingomyelinase-ceramide cascade mediated by synergistic activation of c-Jun N-terminal kinase and p38 mitogen-activated protein kinase. Mol. Cancer 2010, 9, 239. [Google Scholar] [CrossRef] [Green Version]
- Malik, F.; Kumar, A.; Bhushan, S.; Khan, S.; Bhatia, A.; Suri, K.A.; Qazi, G.N.; Singh, J. Reactive oxygen species generation and mitochondrial dysfunction in the apoptotic cell death of human myeloid leukemia HL-60 cells by a dietary compound withaferin A with concomitant protection by N-acetyl cysteine. Apoptosis 2007, 12, 2115–2133. [Google Scholar] [CrossRef]
- Kumar, R.; Sanawar, R.; Li, X.; Li, F. Structure, biochemistry, and biology of PAK kinases. Gene 2017, 605, 20–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forde, J.A.; Dale, T.C. Glycogen synthase kinase 3: A key regulator of cellular fate. Cell. Mol. Life Sci. 2007, 64, 1930–1944. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682. [Google Scholar] [CrossRef]
- Casado, P.; Rodriguez-Prados, J.-C.; Cosulich, S.C.; Guichard, S.; Vanhaesebroeck, B.; Joel, S.; Cutillas, P.R. Kinase-substrate enrichment analysis provides insights into the heterogeneity of signaling pathway activation in leukemia cells. Sci. Signal. 2013, 6, rs6. [Google Scholar] [CrossRef] [PubMed]
- Billottet, C.; Grandage, V.; Gale, R.; Quattropani, A.; Rommel, C.; Vanhaesebroeck, B.; Khwaja, A. A selective inhibitor of the p110 δ isoform of PI 3-kinase inhibits AML cell proliferation and survival and increases the cytotoxic effects of VP16. Oncogene 2006, 25, 6648–6659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.; Sesay, A.; Mitchell, B. Effect of CAL-101, a PI3Kδ inhibitor, on ribosomal rna synthesis and cell proliferation in acute myeloid leukemia cells. Blood Cancer J. 2014, 4, e228. [Google Scholar] [CrossRef] [Green Version]
- Senthil, V.; Ramadevi, S.; Venkatakrishnan, V.; Giridharan, P.; Lakshmi, B.; Vishwakarma, R.; Balakrishnan, A. Withanolide induces apoptosis in HL-60 leukemia cells via mitochondria mediated cytochrome c release and caspase activation. Chem. Biol. Interact. 2007, 167, 19–30. [Google Scholar] [CrossRef]
- Dutta, R.; Khalil, R.; Green, R.; Mohapatra, S.S.; Mohapatra, S. Withania somnifera (Ashwagandha) and withaferin A: Potential in integrative oncology. Int. J. Mol. Sci. 2019, 20, 5310. [Google Scholar] [CrossRef] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akhtar, N.; Baig, M.W.; Haq, I.-u.; Rajeeve, V.; Cutillas, P.R. Withanolide Metabolites Inhibit PI3K/AKT and MAPK Pro-Survival Pathways and Induce Apoptosis in Acute Myeloid Leukemia Cells. Biomedicines 2020, 8, 333. https://doi.org/10.3390/biomedicines8090333
Akhtar N, Baig MW, Haq I-u, Rajeeve V, Cutillas PR. Withanolide Metabolites Inhibit PI3K/AKT and MAPK Pro-Survival Pathways and Induce Apoptosis in Acute Myeloid Leukemia Cells. Biomedicines. 2020; 8(9):333. https://doi.org/10.3390/biomedicines8090333
Chicago/Turabian StyleAkhtar, Nosheen, Muhammad Waleed Baig, Ihsan-ul Haq, Vinothini Rajeeve, and Pedro Rodriguez Cutillas. 2020. "Withanolide Metabolites Inhibit PI3K/AKT and MAPK Pro-Survival Pathways and Induce Apoptosis in Acute Myeloid Leukemia Cells" Biomedicines 8, no. 9: 333. https://doi.org/10.3390/biomedicines8090333