
Clonal and Scalable Endothelial Progenitor Cell Lines from Human Pluripotent Stem Cells
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Derivation of Endothelial Progenitor Cell Lines
2.2. Cell Lines and Culture
2.3. Transcriptomic RNA-Sequencing Analysis
2.4. Whole Genome Microarray Analysis
2.5. Statistical Analyses
2.6. Flow Cytometry
2.7. Immunocytochemistry
2.8. Vascular Tube Forming Assay
3. Results
3.1. Derivation of Clonal Embryonic Endothelial Progenitor Cell (eEPC) Lines
3.2. Transcriptomic Analysis Indicates the Clonal eEPC Lines Are Endothelial Cells
3.3. eEPC Lines Express Surface Proteins and Cellular Markers of Endothelial Cells
3.4. eEPC Lines Enhance Tube Formation in Response to VEGF
3.5. eEPC Lines Retain Embryonic Phenotype
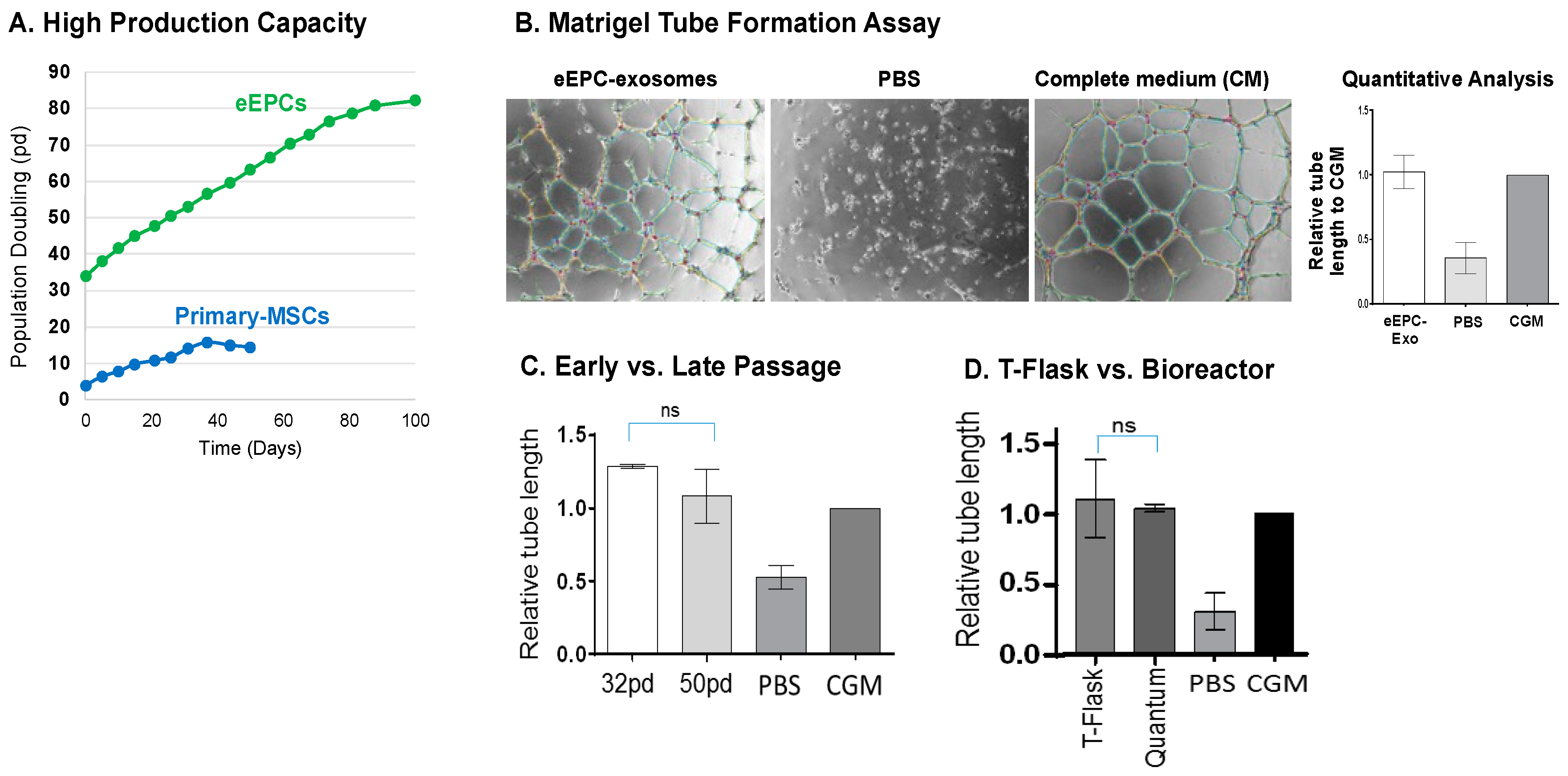
3.6. Stable Production of Angiogenic Embryonic Endothelial Progenitor Cells (eEPCs)
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Severino, P.; D’Amato, A.; Pucci, M.; Infusino, F.; Adamo, F.; Birtolo, L.I.; Netti, L.; Montefusco, G.; Chimenti, C.; Lavalle, C.; et al. Ischemic Heart Disease Pathophysiology Paradigms Overview: From Plaque Activation to Microvascular Dysfunction. Int. J. Mol. Sci. 2020, 21, 8118. [Google Scholar] [CrossRef] [PubMed]
- Ting, K.K.; Coleman, P.; Zhao, Y.; Vadas, M.A.; Gamble, J.R. The aging endothelium. Vasc. Biol. 2021, 3, R35–R47. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Kim, S.Y. Endothelial senescence in vascular diseases: Current understanding and future opportunities in senotherapeutics. Exp. Mol. Med. 2023, 55, 1–12. [Google Scholar] [CrossRef]
- Liew, A.; O’Brien, T. Therapeutic potential for mesenchymal stem cell transplantation in critical limb ischemia. Stem Cell Res. Ther. 2012, 3, 28. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.M.; Hur, S.M.; Park, K.Y.; Kim, C.K.; Kim, Y.M.; Kim, H.S.; Shin, H.C.; Won, M.H.; Ha, K.S.; Kwon, Y.G.; et al. Multiple paracrine factors secreted by mesenchymal stem cells contribute to angiogenesis. Vasc. Pharmacol. 2014, 63, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.C.; Shyu, W.C.; Lin, S.Z.; Li, H. The role of endothelial progenitor cells in ischemic cerebral and heart diseases. Cell Transplant. 2007, 16, 273–284. [Google Scholar] [CrossRef]
- Malliaras, K.; Li, T.S.; Luthringer, D.; Terrovitis, J.; Cheng, K.; Chakravarty, T.; Galang, G.; Zhang, Y.; Schoenhoff, F.; Van Eyk, J.; et al. Safety and efficacy of allogeneic cell therapy in infarcted rats transplanted with mismatched cardiosphere-derived cells. Circulation 2012, 125, 100–112. [Google Scholar] [CrossRef]
- Yu, H.; Lu, K.; Zhu, J.; Wang, J. Stem cell therapy for ischemic heart diseases. Br. Med. Bull. 2017, 121, 135–154. [Google Scholar] [CrossRef]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef]
- Rufaihah, A.J.; Huang, N.F.; Jame, S.; Lee, J.C.; Nguyen, H.N.; Byers, B.; De, A.; Okogbaa, J.; Rollins, M.; Reijo-Pera, R.; et al. Endothelial cells derived from human iPSCS increase capillary density and improve perfusion in a mouse model of peripheral arterial disease. Arterioscler. Thromb. Vasc. Biol. 2011, 31, e72–e79. [Google Scholar] [CrossRef]
- Cho, S.W.; Moon, S.H.; Lee, S.H.; Kang, S.W.; Kim, J.; Lim, J.M.; Kim, H.S.; Kim, B.S.; Chung, H.M. Improvement of postnatal neovascularization by human embryonic stem cell derived endothelial-like cell transplantation in a mouse model of hindlimb ischemia. Circulation 2007, 116, 2409–2419. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.F.; Niiyama, H.; Peter, C.; De, A.; Natkunam, Y.; Fleissner, F.; Li, Z.; Rollins, M.D.; Wu, J.C.; Gambhir, S.S.; et al. Embryonic stem cell-derived endothelial cells engraft into the ischemic hindlimb and restore perfusion. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 984–991. [Google Scholar] [CrossRef] [PubMed]
- James, D.; Nam, H.S.; Seandel, M.; Nolan, D.; Janovitz, T.; Tomishima, M.; Studer, L.; Lee, G.; Lyden, D.; Benezra, R.; et al. Expansion and maintenance of human embryonic stem cell-derived endothelial cells by TGFbeta inhibition is Id1 dependent. Nat. Biotechnol. 2010, 28, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Suzuki, Y.; Huang, M.; Cao, F.; Xie, X.; Connolly, A.J.; Yang, P.C.; Wu, J.C. Comparison of reporter gene and iron particle labeling for tracking fate of human embryonic stem cells and differentiated endothelial cells in living subjects. Stem Cells 2008, 26, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Sumi, T.; Tsuneyoshi, N.; Nakatsuji, N.; Suemori, H. Defining early lineage specification of human embryonic stem cells by the orchestrated balance of canonical Wnt/beta-catenin, Activin/Nodal and BMP signaling. Development 2008, 135, 2969–2979. [Google Scholar] [CrossRef]
- Orlova, V.V.; Drabsch, Y.; Freund, C.; Petrus-Reurer, S.; van den Hil, F.E.; Muenthaisong, S.; Dijke, P.T.; Mummery, C.L. Functionality of endothelial cells and pericytes from human pluripotent stem cells demonstrated in cultured vascular plexus and zebrafish xenografts. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 177–186. [Google Scholar] [CrossRef]
- Alsanie, W.F.; Niclis, J.C.; Petratos, S. Human embryonic stem cell-derived oligodendrocytes: Protocols and perspectives. Stem Cells Dev. 2013, 22, 2459–2476. [Google Scholar] [CrossRef]
- Mehat, M.S.; Sundaram, V.; Ripamonti, C.; Robson, A.G.; Smith, A.J.; Borooah, S.; Robinson, M.; Rosenthal, A.N.; Innes, W.; Weleber, R.G.; et al. Transplantation of Human Embryonic Stem Cell-Derived Retinal Pigment Epithelial Cells in Macular Degeneration. Ophthalmology 2018, 125, 1765–1775. [Google Scholar] [CrossRef]
- Jang, S.; de l’Hortet, A.C.; Soto-Gutierrez, A. Induced Pluripotent Stem Cell-Derived Endothelial Cells: Overview, Current Advances, Applications, and Future Directions. Am. J. Pathol. 2019, 189, 502–512. [Google Scholar] [CrossRef]
- Rufaihah, A.J.; Huang, N.F.; Kim, J.; Herold, J.; Volz, K.S.; Park, T.S.; Lee, J.C.; Zambidis, E.T.; Reijo-Pera, R.; Cooke, J.P. Human induced pluripotent stem cell-derived endothelial cells exhibit functional heterogeneity. Am. J. Transl. Res. 2013, 5, 21–35. [Google Scholar]
- West, M.D.; Sargent, R.G.; Long, J.; Brown, C.; Chu, J.S.; Kessler, S.; Derugin, N.; Sampathkumar, J.; Burrows, C.; Vaziri, H.; et al. The ACTCellerate initiative: Large-scale combinatorial cloning of novel human embryonic stem cell derivatives. Regen. Med. 2008, 3, 287–308. [Google Scholar] [CrossRef]
- West, M.D.; Labat, I.; Sternberg, H.; Larocca, D.; Nasonkin, I.; Chapman, K.B.; Singh, R.; Makarev, E.; Aliper, A.; Kazennov, A.; et al. Use of deep neural network ensembles to identify embryonic-fetal transition markers: Repression of COX7A1 in embryonic and cancer cells. Oncotarget 2018, 9, 7796–7811. [Google Scholar] [CrossRef]
- West, M.D.; Chang, C.F.; Larocca, D.; Li, J.; Jiang, J.; Sim, P.; Labat, I.; Chapman, K.B.; Wong, K.E.; Nicoll, J.; et al. Clonal derivation of white and brown adipocyte progenitor cell lines from human pluripotent stem cells. Stem Cell Res. Ther. 2019, 10, 7. [Google Scholar] [CrossRef]
- Sternberg, H.; Kidd, J.; Murai, J.T.; Jiang, J.; Rinon, A.; Erickson, I.E.; Funk, W.D.; Wang, Q.; Chapman, K.B.; Vangsness, C.T., Jr.; et al. Seven diverse human embryonic stem cell-derived chondrogenic clonal embryonic progenitor cell lines display site-specific cell fates. Regen. Med. 2013, 8, 125–144. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, H.; Murai, J.T.; Erickson, I.E.; Funk, W.D.; Das, S.; Wang, Q.; Snyder, E.; Chapman, K.B.; Vangsness, C.T., Jr.; West, M.D. A human embryonic stem cell-derived clonal progenitor cell line with chondrogenic potential and markers of craniofacial mesenchyme. Regen. Med. 2012, 7, 481–501. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, H.; Jiang, J.; Sim, P.; Kidd, J.; Janus, J.; Rinon, A.; Edgar, R.; Shitrit, A.; Larocca, D.; Chapman, K.B.; et al. Human embryonic stem cell-derived neural crest cells capable of expressing markers of osteochondral or meningeal-choroid plexus differentiation. Regen. Med. 2014, 9, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Dunning, M.J.; Smith, M.L.; Ritchie, M.E.; Tavare, S. beadarray: R classes and methods for Illumina bead-based data. Bioinformatics 2007, 23, 2183–2184. [Google Scholar] [CrossRef] [PubMed]
- Greenwood-Goodwin, M.; Yang, J.; Hassanipour, M.; Larocca, D. A novel lineage restricted, pericyte-like cell line isolated from human embryonic stem cells. Sci. Rep. 2016, 6, 24403. [Google Scholar] [CrossRef]
- Grant, D.; Wanner, N.; Frimel, M.; Erzurum, S.; Asosingh, K. Comprehensive phenotyping of endothelial cells using flow cytometry 2: Human. Cytometry A 2021, 99, 257–264. [Google Scholar] [CrossRef]
- Hassanpour, M.; Salybekov, A.A.; Kobayashi, S.; Asahara, T. CD34 positive cells as endothelial progenitor cells in biology and medicine. Front. Cell Dev. Biol. 2023, 11, 1128134. [Google Scholar] [CrossRef]
- Santhanam, L.; Liu, G.; Jandu, S.; Su, W.; Wodu, B.P.; Savage, W.; Poe, A.; Liu, X.; Alexander, L.M.; Cao, X.; et al. Skeleton-secreted PDGF-BB mediates arterial stiffening. J. Clin. Investig. 2021, 131, e147116. [Google Scholar] [CrossRef] [PubMed]
- Winnicki, A.; Gadd, J.; Ohanyan, V.; Hernandez, G.; Wang, Y.; Enrick, M.; McKillen, H.; Kiedrowski, M.; Kundu, D.; Kegecik, K.; et al. Role of endothelial CXCR4 in the development of aortic valve stenosis. Front. Cardiovasc. Med. 2022, 9, 971321. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, A.; O’Clair, B.; Groppi, V.E.; McEwen, D.P. Pharmacologic characterization of a kinetic in vitro human co-culture angiogenesis model using clinically relevant compounds. J. Biomol. Screen. 2013, 18, 1234–1245. [Google Scholar] [CrossRef]
- Jackson, M.; Fidanza, A.; Taylor, A.H.; Rybtsov, S.; Axton, R.; Kydonaki, M.; Meek, S.; Burdon, T.; Medvinsky, A.; Forrester, L.M. Modulation of APLNR Signaling Is Required during the Development and Maintenance of the Hematopoietic System. Stem Cell Rep. 2021, 16, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Maeng, Y.S.; Choi, H.J.; Kwon, J.Y.; Park, Y.W.; Choi, K.S.; Min, J.K.; Kim, Y.H.; Suh, P.G.; Kang, K.S.; Won, M.H.; et al. Endothelial progenitor cell homing: Prominent role of the IGF2-IGF2R-PLCbeta2 axis. Blood 2009, 113, 233–243. [Google Scholar] [CrossRef]
- Jeong, H.W.; Hernandez-Rodriguez, B.; Kim, J.; Kim, K.P.; Enriquez-Gasca, R.; Yoon, J.; Adams, S.; Scholer, H.R.; Vaquerizas, J.M.; Adams, R.H. Transcriptional regulation of endothelial cell behavior during sprouting angiogenesis. Nat. Commun. 2017, 8, 726. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Diao, S.; Yang, H.; Cao, Y.; Du, J.; Yang, D. IGFBP5 promotes angiogenic and neurogenic differentiation potential of dental pulp stem cells. Dev. Growth Differ. 2019, 61, 457–465. [Google Scholar] [CrossRef]
- Vander Roest, M.J.; Merryman, W.D. Cyclic Strain Promotes H19 Expression and Vascular Tube Formation in iPSC-Derived Endothelial Cells. Cell Mol. Bioeng. 2020, 13, 369–377. [Google Scholar] [CrossRef]
- Roa-Mansergas, X.; Fado, R.; Atari, M.; Mir, J.F.; Muley, H.; Serra, D.; Casals, N. CPT1C promotes human mesenchymal stem cells survival under glucose deprivation through the modulation of autophagy. Sci. Rep. 2018, 8, 6997. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, Q.; Yu, J.; Zhou, Q.; Deng, Y.; Liu, J.; Zhang, L.; Xu, Y.; Xiong, W.; Wang, Y. Tartrate-resistant acid phosphatase 5 promotes pulmonary fibrosis by modulating beta-catenin signaling. Nat. Commun. 2022, 13, 114. [Google Scholar] [CrossRef]
- Lin, P.; Yan, P.; Zhu, J.; Huang, S.; Wang, Z.; Hu, O.; Jin, H.; Li, Y.; Zhang, L.; Zhao, J.; et al. Spatially multicellular variability of intervertebral disc degeneration by comparative single-cell analysis. Cell Prolif. 2023, 56, e13464. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Kuang, S.; Zhang, M.; Huang, X.; Chen, J.; Guan, M.; Qin, W.; Xu, H.H.K.; Lin, Z. Inhibition of CCL2 by bindarit alleviates diabetes-associated periodontitis by suppressing inflammatory monocyte infiltration and altering macrophage properties. Cell Mol. Immunol. 2021, 18, 2224–2235. [Google Scholar] [CrossRef]
- Qiu, K.; Liang, W.; Wang, S.; Kong, T.; Wang, X.; Li, C.; Wang, Z.; Wu, Y. BACE2 degradation is mediated by both the proteasome and lysosome pathways. BMC Mol. Cell Biol. 2020, 21, 13. [Google Scholar] [CrossRef] [PubMed]
- Nivet-Antoine, V.; Labat, C.; El Shamieh, S.; Dulcire, X.; Cottart, C.H.; Beaudeux, J.L.; Zannad, F.; Visvikis-Siest, S.; Benetos, A. Relationship between catalase haplotype and arterial aging. Atherosclerosis 2013, 227, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Metzemaekers, M.; Gouwy, M.; Proost, P. Neutrophil chemoattractant receptors in health and disease: Double-edged swords. Cell Mol. Immunol. 2020, 17, 433–450. [Google Scholar] [CrossRef]
- Chiang, K.C.; Huang, S.T.; Wu, R.C.; Huang, S.C.; Yeh, T.S.; Chen, M.H.; Hsu, J.T.; Chen, L.W.; Kuo, S.F.; Chueh, H.Y.; et al. Interferon alpha-inducible protein 27 is an oncogene and highly expressed in cholangiocarcinoma patients with poor survival. Cancer Manag. Res. 2019, 11, 1893–1905. [Google Scholar] [CrossRef]
- Yao, J.; Guihard, P.J.; Blazquez-Medela, A.M.; Guo, Y.; Liu, T.; Bostrom, K.I.; Yao, Y. Matrix Gla protein regulates differentiation of endothelial cells derived from mouse embryonic stem cells. Angiogenesis 2016, 19, 1–7. [Google Scholar] [CrossRef]
- He, C.; Yang, W.; Yang, J.; Ding, J.; Li, S.; Wu, H.; Zhou, F.; Jiang, Y.; Teng, L.; Yang, J. Long Noncoding RNA MEG3 Negatively Regulates Proliferation and Angiogenesis in Vascular Endothelial Cells. DNA Cell Biol. 2017, 36, 475–481. [Google Scholar] [CrossRef]
- Zhao, Y.H.; Yuan, B.; Chen, J.; Feng, D.H.; Zhao, B.; Qin, C.; Chen, Y.F. Endothelial progenitor cells: Therapeutic perspective for ischemic stroke. CNS Neurosci. Ther. 2013, 19, 67–75. [Google Scholar] [CrossRef]
- Cantaluppi, V.; Biancone, L.; Figliolini, F.; Beltramo, S.; Medica, D.; Deregibus, M.C.; Galimi, F.; Romagnoli, R.; Salizzoni, M.; Tetta, C.; et al. Microvesicles derived from endothelial progenitor cells enhance neoangiogenesis of human pancreatic islets. Cell Transplant. 2012, 21, 1305–1320. [Google Scholar] [CrossRef]
- Yan, F.; Li, J.; Zhang, W. Transplantation of Endothelial Progenitor Cells: Summary and prospect. Acta Histochem. 2023, 125, 151990. [Google Scholar] [CrossRef]
- Ishii, M.; Koike, C.; Igarashi, A.; Yamanaka, K.; Pan, H.; Higashi, Y.; Kawaguchi, H.; Sugiyama, M.; Kamata, N.; Iwata, T.; et al. Molecular markers distinguish bone marrow mesenchymal stem cells from fibroblasts. Biochem. Biophys. Res. Commun. 2005, 332, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.S.; Arslan, F.; Yin, Y.; Tan, S.S.; Lai, R.C.; Choo, A.B.; Padmanabhan, J.; Lee, C.N.; de Kleijn, D.P.; Lim, S.K. Enabling a robust scalable manufacturing process for therapeutic exosomes through oncogenic immortalization of human ESC-derived MSCs. J. Transl. Med. 2011, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Jakob, P.; Landmesser, U. Role of microRNAs in stem/progenitor cells and cardiovascular repair. Cardiovasc. Res. 2012, 93, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. The role of miR-126 in embryonic angiogenesis, adult vascular homeostasis, and vascular repair and its alterations in atherosclerotic disease. J. Mol. Cell Cardiol. 2016, 97, 47–55. [Google Scholar] [CrossRef]
- Venkat, P.; Cui, C.; Chopp, M.; Zacharek, A.; Wang, F.; Landschoot-Ward, J.; Shen, Y.; Chen, J. MiR-126 Mediates Brain Endothelial Cell Exosome Treatment-Induced Neurorestorative Effects After Stroke in Type 2 Diabetes Mellitus Mice. Stroke 2019, 50, 2865–2874. [Google Scholar] [CrossRef]
- Bezenah, J.R.; Kong, Y.P.; Putnam, A.J. Evaluating the potential of endothelial cells derived from human induced pluripotent stem cells to form microvascular networks in 3D cultures. Sci. Rep. 2018, 8, 2671. [Google Scholar] [CrossRef]
- Hur, J.; Yoon, C.H.; Kim, H.S.; Choi, J.H.; Kang, H.J.; Hwang, K.K.; Oh, B.H.; Lee, M.M.; Park, Y.B. Characterization of two types of endothelial progenitor cells and their different contributions to neovasculogenesis. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 288–293. [Google Scholar] [CrossRef]
- Fadini, G.P.; Losordo, D.; Dimmeler, S. Critical reevaluation of endothelial progenitor cell phenotypes for therapeutic and diagnostic use. Circ. Res. 2012, 110, 624–637. [Google Scholar] [CrossRef]
- Liu, J.W.; Dunoyer-Geindre, S.; Serre-Beinier, V.; Mai, G.; Lambert, J.F.; Fish, R.J.; Pernod, G.; Buehler, L.; Bounameaux, H.; Kruithof, E.K. Characterization of endothelial-like cells derived from human mesenchymal stem cells. J. Thromb. Haemost. 2007, 5, 826–834. [Google Scholar] [CrossRef]
- Fan, Y.; Shen, F.; Frenzel, T.; Zhu, W.; Ye, J.; Liu, J.; Chen, Y.; Su, H.; Young, W.L.; Yang, G.Y. Endothelial progenitor cell transplantation improves long-term stroke outcome in mice. Ann. Neurol. 2010, 67, 488–497. [Google Scholar] [CrossRef]
- Koltowska, K.; Paterson, S.; Bower, N.I.; Baillie, G.J.; Lagendijk, A.K.; Astin, J.W.; Chen, H.; Francois, M.; Crosier, P.S.; Taft, R.J.; et al. mafba is a downstream transcriptional effector of Vegfc signaling essential for embryonic lymphangiogenesis in zebrafish. Genes. Dev. 2015, 29, 1618–1630. [Google Scholar] [CrossRef] [PubMed]
- Dieterich, L.C.; Tacconi, C.; Menzi, F.; Proulx, S.T.; Kapaklikaya, K.; Hamada, M.; Takahashi, S.; Detmar, M. Lymphatic MAFB regulates vascular patterning during developmental and pathological lymphangiogenesis. Angiogenesis 2020, 23, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Forbes, M.E.; Fuller, G.N.; Li, J.; Yang, X.; Zhang, W. IGFBP2: Integrative hub of developmental and oncogenic signaling network. Oncogene 2020, 39, 2243–2257. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Menzies, K.; Freeman, R.S.; Taubman, M.B. EGLN3 prolyl hydroxylase regulates skeletal muscle differentiation and myogenin protein stability. J. Biol. Chem. 2007, 282, 12410–12418. [Google Scholar] [CrossRef]
- Lisignoli, G.; Piacentini, A.; Toneguzzi, S.; Grassi, F.; Tschon, M.; Cristino, S.; Facchini, A.; Mariani, E. Age-associated changes in functional response to CXCR3 and CXCR5 chemokine receptors in human osteoblasts. Biogerontology 2003, 4, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Sweatt, A.; Sane, D.C.; Hutson, S.M.; Wallin, R. Matrix Gla protein (MGP) and bone morphogenetic protein-2 in aortic calcified lesions of aging rats. J. Thromb. Haemost. 2003, 1, 178–185. [Google Scholar] [CrossRef]
- Chapman, K.B.; Prendes, M.J.; Sternberg, H.; Kidd, J.L.; Funk, W.D.; Wagner, J.; West, M.D. COL10A1 expression is elevated in diverse solid tumor types and is associated with tumor vasculature. Future Oncol. 2012, 8, 1031–1040. [Google Scholar] [CrossRef]
- Nakajima, H.; Ishikawa, H.; Yamamoto, T.; Chiba, A.; Fukui, H.; Sako, K.; Fukumoto, M.; Mattonet, K.; Kwon, H.B.; Hui, S.P.; et al. Endoderm-derived islet1-expressing cells differentiate into endothelial cells to function as the vascular HSPC niche in zebrafish. Dev. Cell 2023, 58, 224–238.e7. [Google Scholar] [CrossRef]
- Goldman, O.; Han, S.; Hamou, W.; de Villeroche, V.J.; Uzan, G.; Lickert, H.; Gouon-Evans, V. Endoderm generates endothelial cells during liver development. Stem Cell Rep. 2014, 3, 556–565. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Sternberg, H.; Bignone, P.A.; Murai, J.; Malik, N.N.; West, M.D.; Larocca, D. Clonal and Scalable Endothelial Progenitor Cell Lines from Human Pluripotent Stem Cells. Biomedicines 2023, 11, 2777. https://doi.org/10.3390/biomedicines11102777
Lee J, Sternberg H, Bignone PA, Murai J, Malik NN, West MD, Larocca D. Clonal and Scalable Endothelial Progenitor Cell Lines from Human Pluripotent Stem Cells. Biomedicines. 2023; 11(10):2777. https://doi.org/10.3390/biomedicines11102777
Chicago/Turabian StyleLee, Jieun, Hal Sternberg, Paola A. Bignone, James Murai, Nafees N. Malik, Michael D. West, and Dana Larocca. 2023. "Clonal and Scalable Endothelial Progenitor Cell Lines from Human Pluripotent Stem Cells" Biomedicines 11, no. 10: 2777. https://doi.org/10.3390/biomedicines11102777