
Melatonin Preserves Fluidity in Cell and Mitochondrial Membranes against Hepatic Ischemia–Reperfusion
 , , , ,
, , , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Model and Sample Collection
2.3. Membrane Isolation and Fluidity Measurements
2.4. Assay of Lipid Peroxidation
2.5. Protein Carbonylation
2.6. Immunohistochemical Studies
2.7. Statistical Analyses
3. Results
3.1. Cell and Mitochondrial Membrane Fluidity
3.2. Biochemical Indices of Oxidative Stress
3.2.1. Lipid Peroxidation
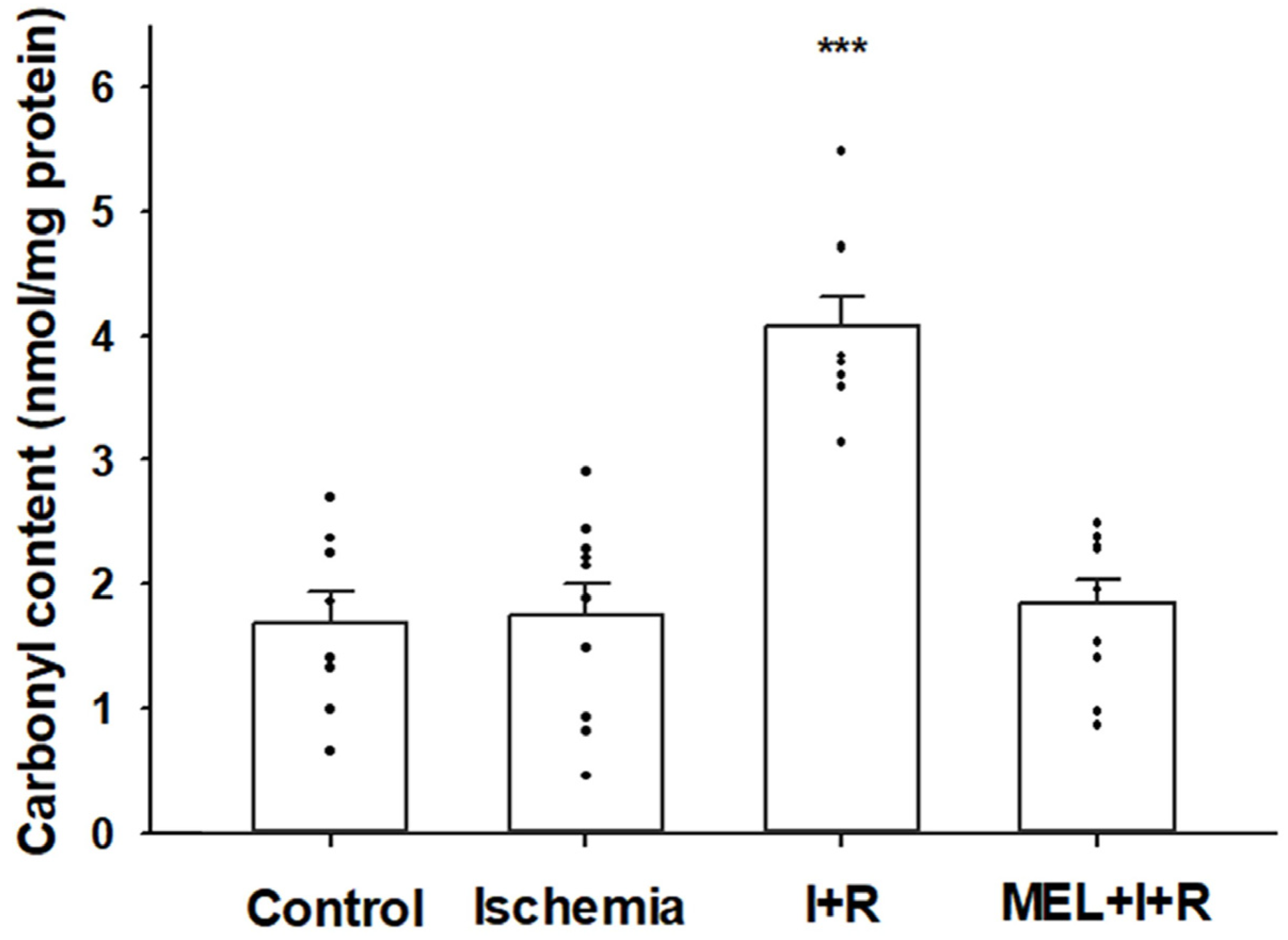
3.2.2. Protein Carbonylation
3.3. Immunohistochemical Features
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eltzschig, H.K.; Eckle, T. Ischemia and reperfusion—From mechanism to translation. Nat. Med. 2011, 17, 1391–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Ruiz, C.; Fernández-Checa, J.C. Mitochondrial glutathione: Hepatocellular survival-death switch. J. Gastroenterol. Hepatol. 2006, 21, S3–S6. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Xin, Z.; Di, W.; Yan, X.; Li, X.; Reiter, R.J.; Yang, Y. Melatonin and mitochondrial function during ischemia/reperfusion injury. Cell. Mol. Life Sci. 2017, 74, 3989–3998. [Google Scholar] [CrossRef] [PubMed]
- Zaouali, M.A.; Ben Abdennebi, H.; Padrissa-Altés, S.; Mahfoudh-Boussaid, A.; Roselló-Catafau, J. Pharmacological strategies against cold ischemia reperfusion injury. Expert Opin. Pharmacother. 2010, 11, 537–555. [Google Scholar] [CrossRef]
- Musatov, A.; Robinson, N.C. Susceptibility of mitochondrial electron-transport complexes to oxidative damage. Focus on cytochrome c oxidase. Free. Radic. Res. 2012, 46, 1313–1326. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [Green Version]
- Jaeschke, H.; Woolbright, B.L. Current strategies to minimize hepatic ischemia-reperfusion injury by targeting reactive oxygen species. Transpl. Rev. 2012, 26, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Karatzas, T.; Neri, A.A.; Baibaki, M.E.; Dontas, I.A. Rodent models of hepatic ischemia-reperfusion injury: Time and percentage-related pathophysiological mechanisms. J. Surg. Res. 2014, 191, 399–412. [Google Scholar] [CrossRef]
- Crompton, M. The mitochondrial permeability transition pore and its role in cell death. Biochem. J. 1999, 341, 233–249. [Google Scholar]
- Serracino-Inglott, F.; Habib, N.A.; Mathie, R.T. Hepatic ischemia-reperfusion injury. Am. J. Surg. 2001, 181, 160–166. [Google Scholar] [CrossRef]
- Perry, B.C.; Soltys, D.; Toledo, A.H.; Toledo-Pereyra, L.H. Tumor necrosis factor-α in liver ischemia/reperfusion injury. J. Investig. Surg. 2011, 24, 178–188. [Google Scholar] [CrossRef]
- Lu, L.; Zhou, H.; Ni, M.; Wang, X.; Busuttil, R.; Kupiec-Weglinski, J.; Zhai, Y. Innate immune regulations and liver ischemia-reperfusion injury. Transplantation 2016, 100, 2601–2610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kvetnoy, I.M. Extrapineal melatonin: Location and role within diffuse neuroendocrine system. Histochem. J. 1999, 31, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Rodríguez, A.; Abreu-González, P.; Sánchez-Sánchez, J.J.; Kaski, J.C.; Reiter, R.J. Melatonin and circadian biology in human cardiovascular disease. J. Pineal Res. 2010, 49, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Espino, J.; Pariente, J.A.; Rodríguez, A.B. Oxidative stress and immunosenescence: Therapeutic effects of melatonin. Oxid. Med. Cell. Longev. 2012, 2012, 670294. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.X.; Chen, L.D.; Poeggeler, B.; Manchester, L.C.; Reiter, R.J. Melatonin: A potent, endogenous hydroxyl radical scavenger. Endocr. J. 1993, 1, 57–60. [Google Scholar]
- Tan, D.X.; Manchester, L.C.; Reiter, R.J.; Plummer, B.F.; Limson, J.; Wintraub, S.T.; Qi, W. Melatonin directly scavenges hydrogen peroxide: A potentially new metabolic pathway of melatonin biotransformation. Free. Radic. Biol. Med. 2000, 29, 1177–1185. [Google Scholar] [CrossRef]
- Ximenez, V.F.; Silva, S.O.; Rodrigues, M.R.; Catalani, L.H.; Maghzal, G.J.; Kettle, A.J.; Campa, A. Superoxide dependent oxidation of melatonin by myeloperoxidase. J. Biol. Chem. 2005, 280, 38160–38169. [Google Scholar] [CrossRef] [Green Version]
- Tomás-Zapico, C.; Coto-Montes, A. A proposed mechanism to explain the stimulatory effect of melatonin on antioxidative enzymes. J. Pineal Res. 2005, 39, 99–104. [Google Scholar] [CrossRef]
- Galano, A.; Tan, D.X.; Reiter, R.J. On the free radical scavenging activities of melatonin’s metabolites, AFMK and AMK. J. Pineal Res. 2013, 54, 245–257. [Google Scholar] [CrossRef]
- Bermudez-Gonzalez, J.L.; Sanchez-Quintero, D.; Proaño-Bernal, L.; Santana-Apreza, R.; Jimenez-Chavarria, M.A.; Luna-Alvarez-Amezquita, J.A.; Straface, J.I.; Perez-Partida, A.M.; Berarducci, J.; Armenta-Moreno, J.I.; et al. Role of the Antioxidant Activity of Melatonin in Myocardial Ischemia-Reperfusion Injury. Antioxidants 2022, 11, 627. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Duan, W.; Jin, Z.; Yi, W.; Yan, J.; Zhang, S.; Wang, N.; Liang, Z.; Li, Y.; Chen, W.; et al. JAK2/STAT3 activation by melatonin attenuates the mitochondrial oxidative damage induced by myocardial ischemia/reperfusion injury. J. Pineal Res. 2013, 55, 275–286. [Google Scholar] [CrossRef]
- Mansouri, A.; Gaou, I.; de Kerguenec, C.; Amsellem, S.; Haouzi, D.; Berson, A.; Moreau, A.; Feldmann, G.; Letteron, P.; Pessayre, D.; et al. An alcoholic binge causes massive degradation of hepatic mitochondrial DNA in mice. Gastroenterology 1999, 117, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Martín, M.; Macías, M.; Escames, G.; Reiter, R.J.; Agapito, M.T.; Ortiz, G.G.; Acuña-Castroviejo, D. Melatonin-induced increased activity of the respiratory chain complexes I and IV can prevent mitochondrial damage induced by ruthenium red in vivo. J. Pineal Res. 2000, 28, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escames, G.; León, J.; Macías, M.; Khaldy, H.; Acuña-Castroviejo, D. Melatonin counteracts lipopolysaccharide induced expression and activity of mitochondrial nitric oxide synthase in rats. FASEB J. 2003, 17, 932–934. [Google Scholar] [CrossRef]
- Teng, Y.C.; Tai, Y.I.; Huang, H.J.; Lin, A.M. Melatonin ameliorates arsenite-induced neurotoxicity: Involvement of autophagy and mitochondria. Mol. Neurobiol. 2015, 52, 1015–1022. [Google Scholar] [CrossRef]
- Nair, S.M.; Rahman, R.M.; Clarkson, A.N.; Sutherland, B.A.; Taurin, S.; Sammut, I.A.; Appleton, I. Melatonin treatment following stroke induction modulates l-arginine metabolism. J. Pineal Res. 2011, 51, 313–323. [Google Scholar] [CrossRef]
- Yu, L.; Gong, B.; Duan, W.; Fan, C.; Zhang, J.; Li, Z.; Xue, X.; Xu, Y.; Meng, D.; Li, B.; et al. Melatonin ameliorates myocardial ischemia/reperfusion injury in type 1 diabetic rats by preserving mitochondrial function: Role of AMPK-PGC-1alpha-SIRT3 signaling. Sci. Rep. 2017, 7, 41337. [Google Scholar] [CrossRef] [Green Version]
- Waseem, M.; Tabassum, H.; Parvez, S. Melatonin modulates permeability transition pore and 5-hydroxydecanoate induced KATP channel inhibition in isolated brain mitochondria. Mitochondrion 2016, 31, 1–8. [Google Scholar] [CrossRef]
- Paradies, G.; Petrosillo, G.; Paradies, V.; Reiter, R.J.; Ruggiero, F.M. Melatonin, cardiolipin and mitochondrial bioenergetics in health and disease. J. Pineal Res. 2010, 48, 297–310. [Google Scholar] [CrossRef]
- Kato, H.; Tanaka, G.; Masuda, S.; Ogasawara, J.; Sakurai, T.; Kizaki, T.; Ohno, H.; Izawa, T. Melatonin promotes adipogenesis and mitochondrial biogenesis in 3T3-L1 preadipocytes. J. Pineal Res. 2015, 59, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Vitale, S.G.; Rossetti, P.; Corrado, F.; Rapisarda, A.M.; La Vignera, S.; Condorelli, R.A.; Valenti, G.; Sapia, F.; Laganà, A.S.; Buscema, M. How to achieve high-quality oocytes? The key role of myo-Inositol and melatonin. Int. J. Endocrinol. 2016, 2016, 4987436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, D.X.; Manchester, L.C.; Qin, L.; Reiter, R.J. Melatonin: A mitochondrial targeting molecule involving mitochondrial protection and dynamics. Int. J. Mol. Sci. 2016, 17, 2124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mignotte, B.; Vayssiere, J.L. Mitochondria and apoptosis. Eur. J. Biochem. 1988, 252, 1–15. [Google Scholar] [CrossRef]
- Agil, A.; Navarro-Alarcon, M.; Ali, F.A.Z.; Albrakati, A.; Salagre, D.; Campoy, C.; Elmahallawy, E.K. Melatonin Enhances the Mitochondrial Functionality of Brown Adipose Tissue in Obese-Diabetic Rats. Antioxidants 2021, 10, 1482. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Rosales-Corral, S.; Galano, A.; Zhou, X.J. Mitochondria: Central Organelles for Melatonin Antioxidant and Anti-Aging Actions. Molecules 2018, 23, 509. [Google Scholar] [CrossRef] [Green Version]
- Frich, L.; Mala, T.; Gladhaug, I.P. Hepatic radiofrequency ablation using perfusion electrodes in a pig model: Effect of the Pringle manoeuvre. Eur. J. Surg. Oncol. 2006, 32, 527–532. [Google Scholar] [CrossRef]
- Aranda, M.; Albendea, C.D.; Lostalé, F.; López-Pingarrón, L.; Fuentes-Broto, L.; Martínez-Ballarín, E.; Reiter, R.J.; Pérez-Castejón, M.C.; García, J.J. In vivo hepatic oxidative stress because of carbon tetrachloride toxicity: Protection by melatonin and pinoline. J. Pineal Res. 2010, 49, 78–85. [Google Scholar] [CrossRef]
- Yu, B.P.; Suescun, E.A.; Yang, S.Y. Effect of age-related lipid peroxidation on membrane fluidity and phospholipase A2: Modulation by dietary restriction. Mech. Ageing Dev. 1992, 65, 17–33. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for quantification of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–253. [Google Scholar] [CrossRef]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chio, K.S.; Tappel, A.L. Synthesis and characterization of the fluorescent products derived from malonaldehyde and amino acids. Biochemistry 1969, 8, 2821–2826. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Wu, X.; Naito, M.; Nomura, H.; Kitamoto, N.; Osawa, T. Immunochemical detection of protein dityrosine in atherosclerotic lesion of apo-E-deficient mice using a novel monoclonal antibody. Biochem. Biophys. Res. Commun. 2000, 275, 11–15. [Google Scholar] [CrossRef]
- Casillas-Ramírez, A.; Mosbah, I.B.; Ramalho, F.; Roselló-Catafau, J.; Peralta, C. Past and future approaches to ischemia-reperfusion lesion associated with liver transplantation. Life Sci. 2006, 79, 1881–1894. [Google Scholar] [CrossRef]
- Zhou, X.M.; Cao, Y.L.; Dou, D.Q. Protective effect of ginsenoside-Re against cerebral ischemia/reperfusion damage in rats. Biol. Pharm. Bull. 2006, 29, 2502–2505. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Rao, S.; Yang, M.; Ma, C.; Hong, F.; Yang, S. Role of Mitochondrial Pathways in Cell Apoptosis during Hepatic Ischemia/Reperfusion Injury. Int. J. Mol. Sci. 2022, 23, 2357. [Google Scholar] [CrossRef]
- Okatani, Y.; Wakatsuki, A.; Reiter, R.J.; Miyahara, Y. Acutely administered melatonin restores hepatic mitochondrial physiology in old mice. Int. J. Biochem. Cell Biol. 2003, 35, 367–375. [Google Scholar] [CrossRef]
- Kang, J.W.; Cho, H.I.; Lee, S.M. Melatonin inhibits mTOR-dependent autophagy during liver ischemia/reperfusion. Cell Physiol. Biochem. 2014, 33, 23–36. [Google Scholar] [CrossRef] [Green Version]
- Teoh, N.C.; Farrell, G.C. Hepatic ischemia reperfusion injury: Pathogenic mechanisms and basis for hepatoprotection. J. Gastroenterol. Hepatol. 2003, 18, 891–902. [Google Scholar] [CrossRef]
- Peralta, C.; Jiménez-Castro, M.B.; Gracia-Sancho, J. Hepatic ischemia and reperfusion injury: Effects on the liver sinusoidal milieu. J. Hepatol. 2013, 59, 1094–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Bae, E.H.; Ma, S.K.; Kim, S.W. Altered nitric oxide system in cardiovascular and renal diseases. Chonnam Med. J. 2016, 52, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, I.; Bertone, V.; Guarnaschelli, C.; Ferrigno, A.; Boncompagni, E.; Rizzo, V.; Reiter, R.J.; Barni, S.; Vairetti, M. In situ demonstration of improvement of liver mitochondria function by melatonin after cold ischemia. In Vivo 2006, 20, 229–237. [Google Scholar]
- Park, S.W.; Choi, S.M.; Lee, S.M. Effect of melatonin on altered expression of vasoregulatory genes during hepatic ischemia/reperfusion. Arch. Pharm. Res. 2007, 30, 1619–1624. [Google Scholar] [CrossRef]
- Kim, S.H.; Lee, S.M. Cytoprotective effects of melatonin against necrosis and apoptosis induced by ischemia/reperfusion injury in rat liver. J. Pineal Res. 2008, 44, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Kireev, R.A.; Cuesta, S.; Ibarrola, C.; Bela, T.; Moreno Gonzalez, E.; Vara, E.; Tresguerres, J.A. Age-related differences in hepatic ischemia/reperfusion: Gene activation, liver injury, and protective effect of melatonin. J. Surg. Res. 2012, 178, 922–934. [Google Scholar] [CrossRef] [PubMed]
- Kireev, R.; Bitoun, S.; Cuesta, S.; Tejerina, A.; Ibarrola, C.; Moreno, E.; Vara, E.; Tresguerres, J.A. Melatonin treatment protects liver of Zucker rats after ischemia/reperfusion by diminishing oxidative stress and apoptosis. Eur. J. Pharmacol. 2013, 701, 185–193. [Google Scholar] [CrossRef]
- Lehotský, J.; Murín, R.; Strapková, A.; Uríková, A.; Tatarková, Z.; Kaplán, P. Time course of ischemia/reperfusion-induced oxidative modification of neural proteins in rat forebrain. Gen. Physiol. Biophys. 2004, 23, 401–415. [Google Scholar]
- Kurcer, Z.; Oguz, E.; Ozbilge, H.; Baba, F.; Aksoy, N.; Celik, N. Effect of melatonin on testicular ischemia/reperfusion injury in rats: Is this effect related to the proinflammatory cytokines? Fertil. Steril. 2008, 89, 1468–1473. [Google Scholar] [CrossRef]
- Catalá, A. Lipid peroxidation of membrane phospholipids generates hydroxy-alkenals and oxidized phospholipids active in physiological and/or pathological conditions. Chem. Phys. Lipids 2009, 157, 1–11. [Google Scholar] [CrossRef]
- García, J.J.; López-Pingarrón, L.; Almeida-Souza, P.; Tres, A.; Escudero, P.; García-Gil, F.A.; Tan, D.X.; Reiter, R.J.; Ramírez, J.M.; Bernal-Pérez, M. Protective effects of melatonin in reducing oxidative stress and in preserving the fluidity of biological membranes: A review. J. Pineal Res. 2014, 56, 225–237. [Google Scholar] [CrossRef] [PubMed]
- García-Gil, F.A.; Albendea, C.D.; López-Pingarrón, L.; Royo-Dachary, P.; Martínez-Guillén, J.; Piedrafita, E.; Martínez-Díez, M.; Soria, J.; García, J.J. Altered cellular membrane fluidity levels and lipid peroxidation during experimental pancreas transplantation. J. Bioenerg. Biomembr. 2012, 44, 571–577. [Google Scholar] [CrossRef] [PubMed]
- García-Gil, F.A.; Fuentes-Broto, L.; Albendea, C.D.; Serrano, M.T.; Roselló-Catafau, J.; Lampreave, F.; López-Pingarrón, L.; Escartín, J.; Soria, J.; García, J.J.; et al. Evaluation of Institut Georges Lopez-1 preservation solution in pig pancreas transplantation: A pilot study. Transplantation 2014, 97, 901–907. [Google Scholar] [CrossRef] [Green Version]
- Dutheil, D.; Underhaug Gjerde, A.; Petit-Paris, I.; Mauco, G.; Holmsen, H. Polyethylene glycols interact with membrane glycerophospholipids: Is this part of their mechanism for hypothermic graft protection? J. Chem. Biol. 2009, 2, 39–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venegas, C.; García, J.A.; Escames, G.; Ortiz, F.; López, A.; Doerrier, C.; García-Corzo, L.; López, L.C.; Reiter, R.J.; Acuña-Castroviejo, D. Extrapineal Melatonin: Analysis of Its Subcellular Distribution and Daily Fluctuations. J. Pineal Res. 2012, 52, 217–227. [Google Scholar] [CrossRef]
- He, C.; Wang, J.; Zhang, Z.; Yang, M.; Li, Y.; Tian, X.; Ma, T.; Tao, J.; Zhu, K.; Song, Y.; et al. Mitochondria synthesize melatonin to ameliorate its function and improve mice oocyte’s quality under in vitro conditions. Int. J. Mol. Sci. 2016, 17, 939. [Google Scholar] [CrossRef] [Green Version]
- García, J.J.; Reiter, R.J.; Guerrero, J.M.; Escames, G.; Yu, B.P.; Oh, C.S.; Muñoz-Hoyos, A. Melatonin prevents changes in microsomal membrane fluidity during induced lipid peroxidation. FEBS Lett. 1997, 408, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Ochoa, J.J.; Vílchez, M.J.; Palacios, M.A.; García, J.J.; Reiter, R.J.; Muñoz-Hoyos, A. Melatonin protects against lipid peroxidation and membrane rigidity in erythrocytes from patients undergoing cardiopulmonary bypass surgery. J. Pineal Res. 2003, 35, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Tajeddine, N. How do reactive oxygen species and calcium trigger mitochondrial membrane permeabilisation? Biochim. Biophys. Acta 2016, 1860, 1079–1088. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esteban-Zubero, E.; López-Pingarrón, L.; Ramírez, J.M.; Reyes-Gonzales, M.C.; Azúa-Romeo, F.J.; Soria-Aznar, M.; Agil, A.; García, J.J. Melatonin Preserves Fluidity in Cell and Mitochondrial Membranes against Hepatic Ischemia–Reperfusion. Biomedicines 2023, 11, 1940. https://doi.org/10.3390/biomedicines11071940
Esteban-Zubero E, López-Pingarrón L, Ramírez JM, Reyes-Gonzales MC, Azúa-Romeo FJ, Soria-Aznar M, Agil A, García JJ. Melatonin Preserves Fluidity in Cell and Mitochondrial Membranes against Hepatic Ischemia–Reperfusion. Biomedicines. 2023; 11(7):1940. https://doi.org/10.3390/biomedicines11071940
Chicago/Turabian StyleEsteban-Zubero, Eduardo, Laura López-Pingarrón, José Manuel Ramírez, Marcos César Reyes-Gonzales, Francisco Javier Azúa-Romeo, Marisol Soria-Aznar, Ahmad Agil, and José Joaquín García. 2023. "Melatonin Preserves Fluidity in Cell and Mitochondrial Membranes against Hepatic Ischemia–Reperfusion" Biomedicines 11, no. 7: 1940. https://doi.org/10.3390/biomedicines11071940