
Urinary ACE Phenotyping as a Research and Diagnostic Tool: Identification of Sex-Dependent ACE Immunoreactivity
 ,
,  , , and
, , and 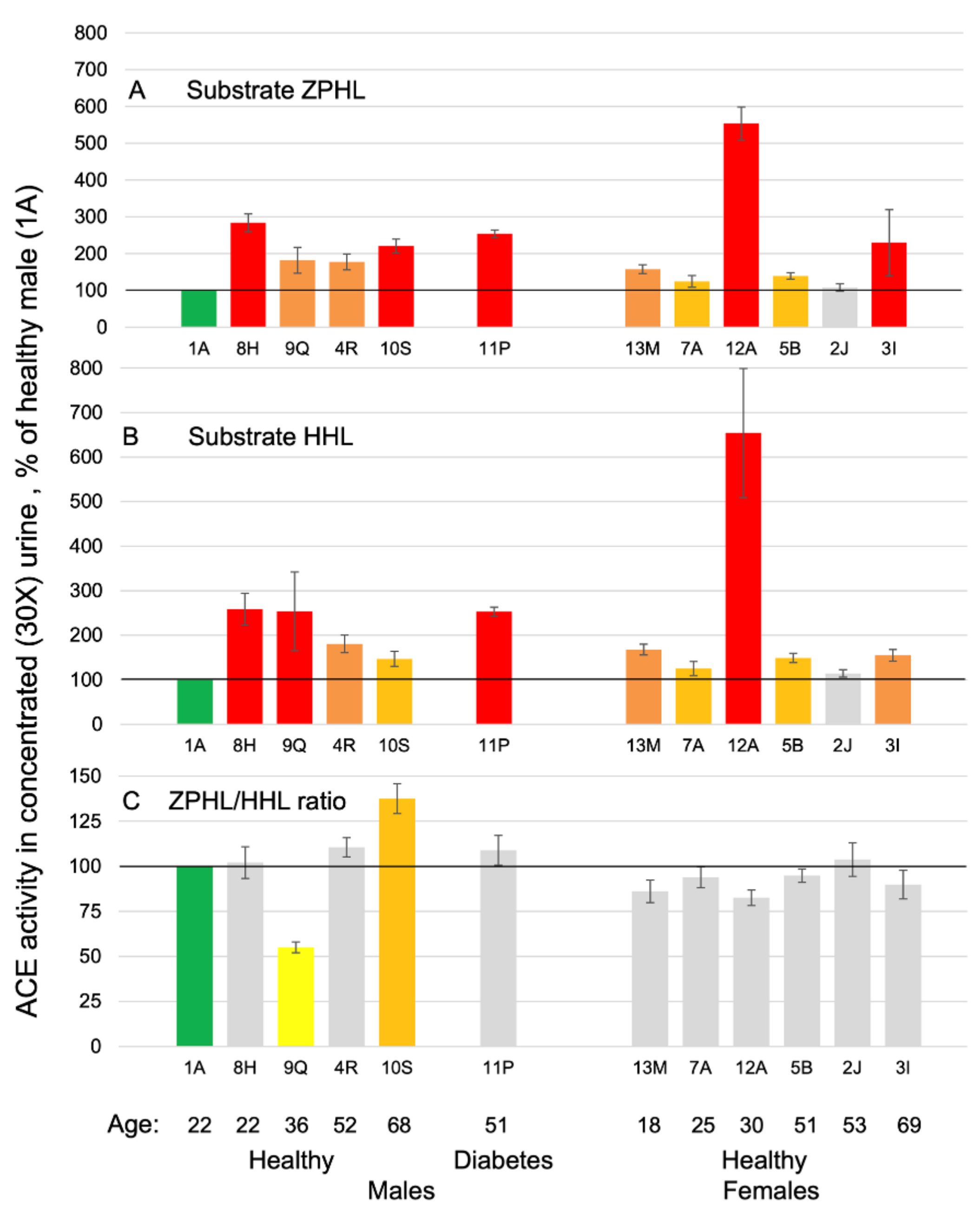{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Antibodies
2.3. Study Participants
2.4. Urines
2.5. ACE Activity Assay
2.6. Immunological Characterization of the Blood ACE
2.7. Whole Exome Sequencing
2.8. Computational Analysis of the Dimer Model with Human Serum Albumin (HSA)
2.9. Statistical Analysis
3. Results
3.1. ACE Activity in Human Urine Samples
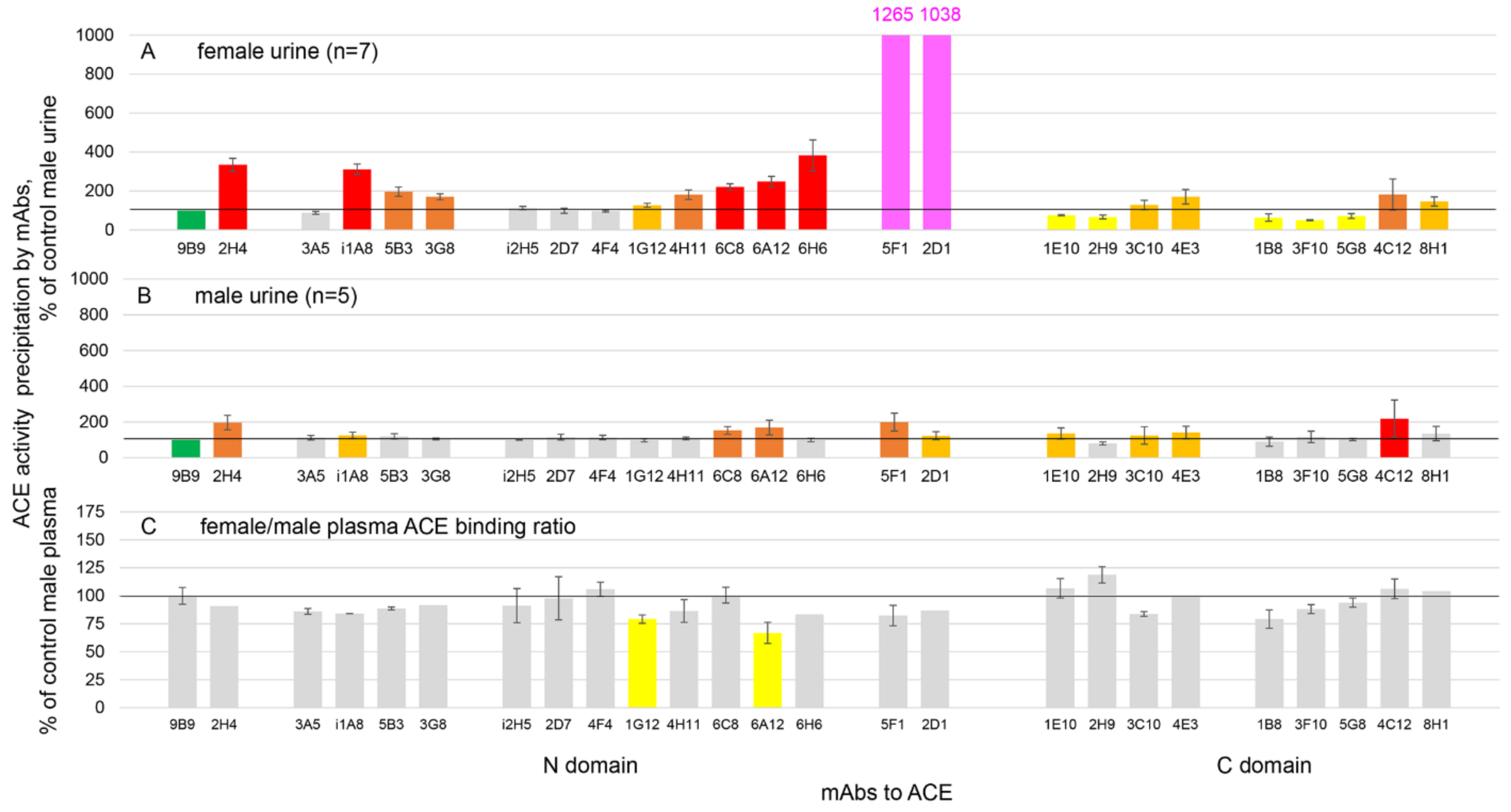
3.2. Immunoreactivity of ACE in Human Urine
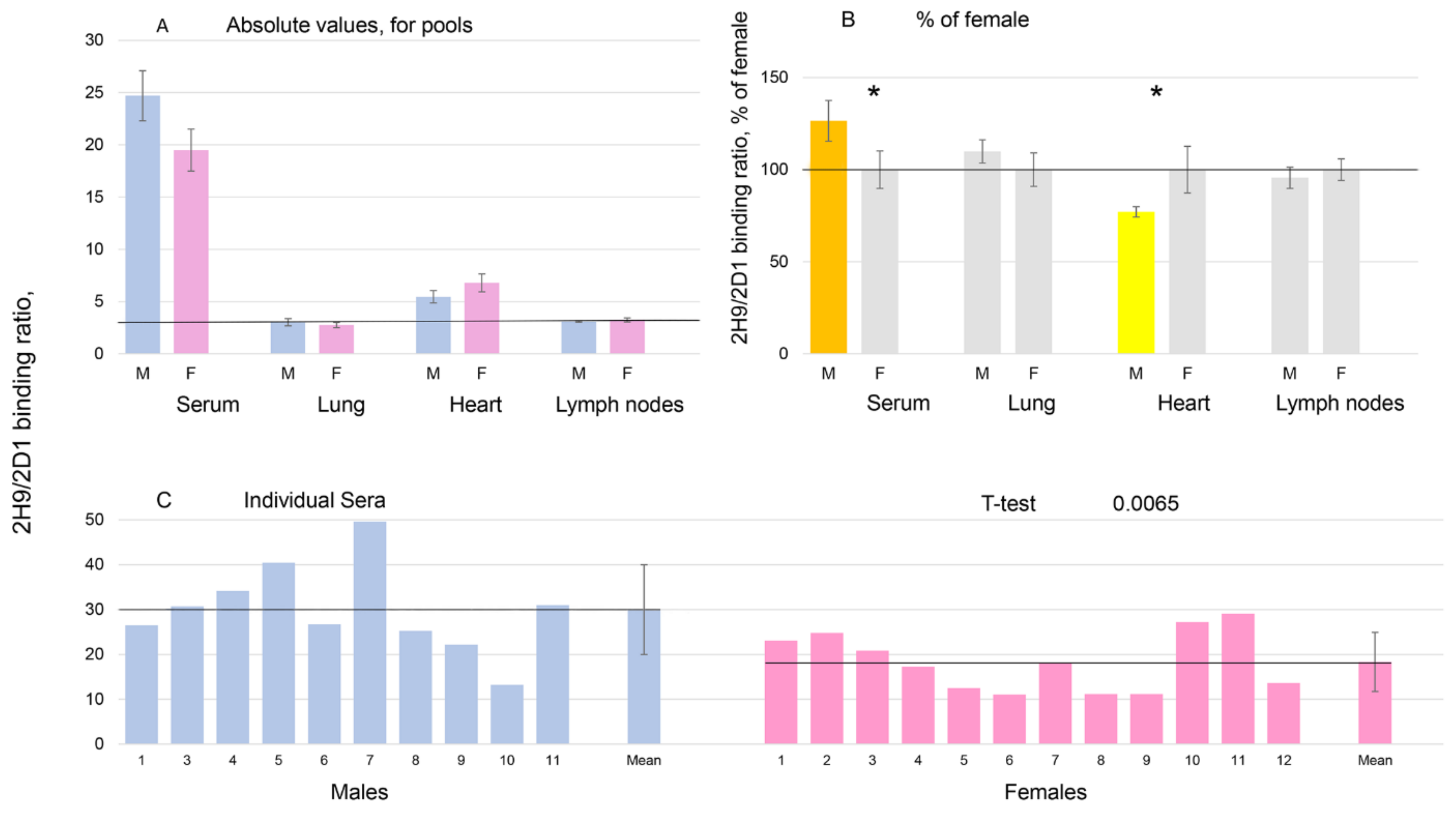
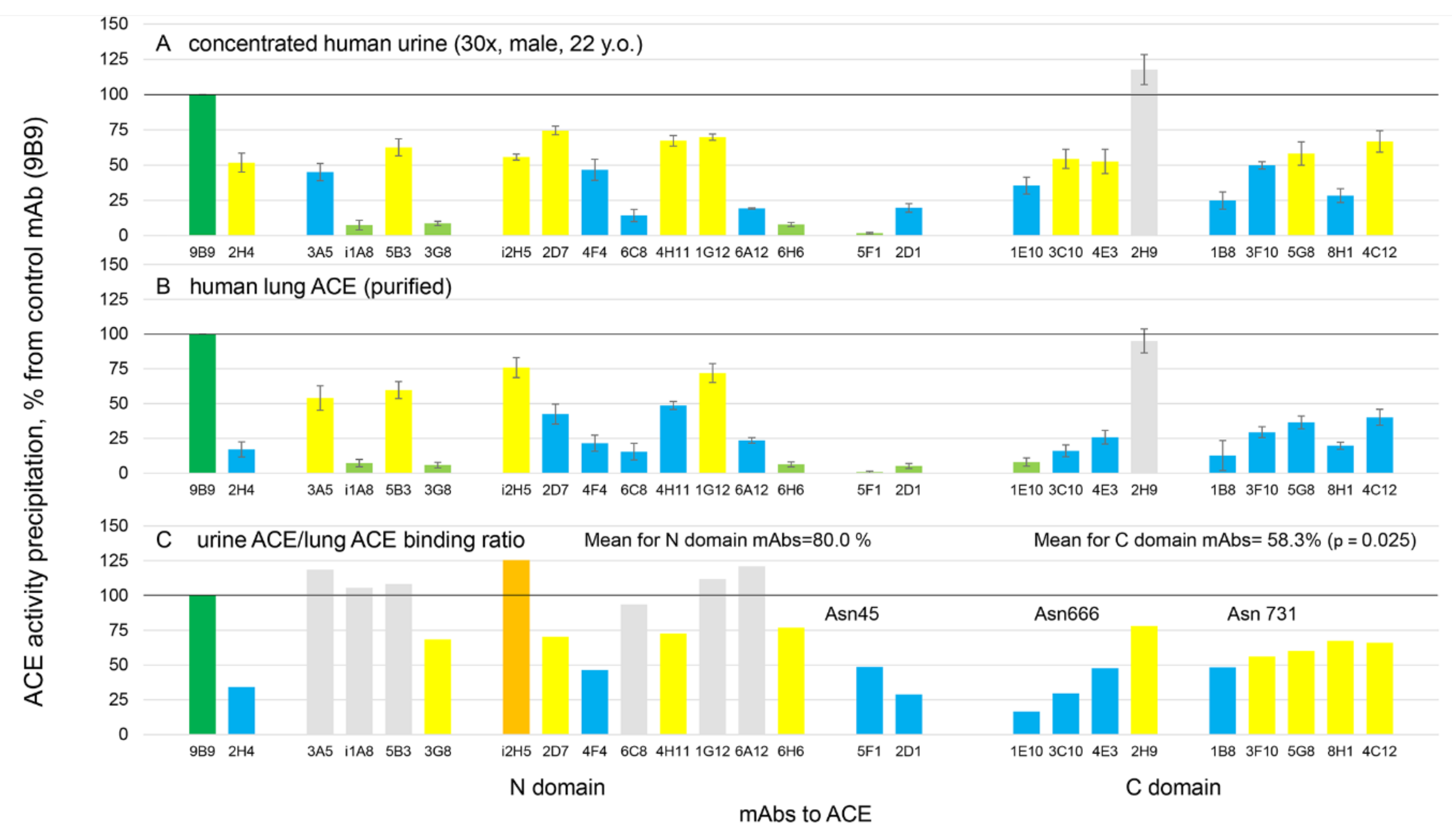
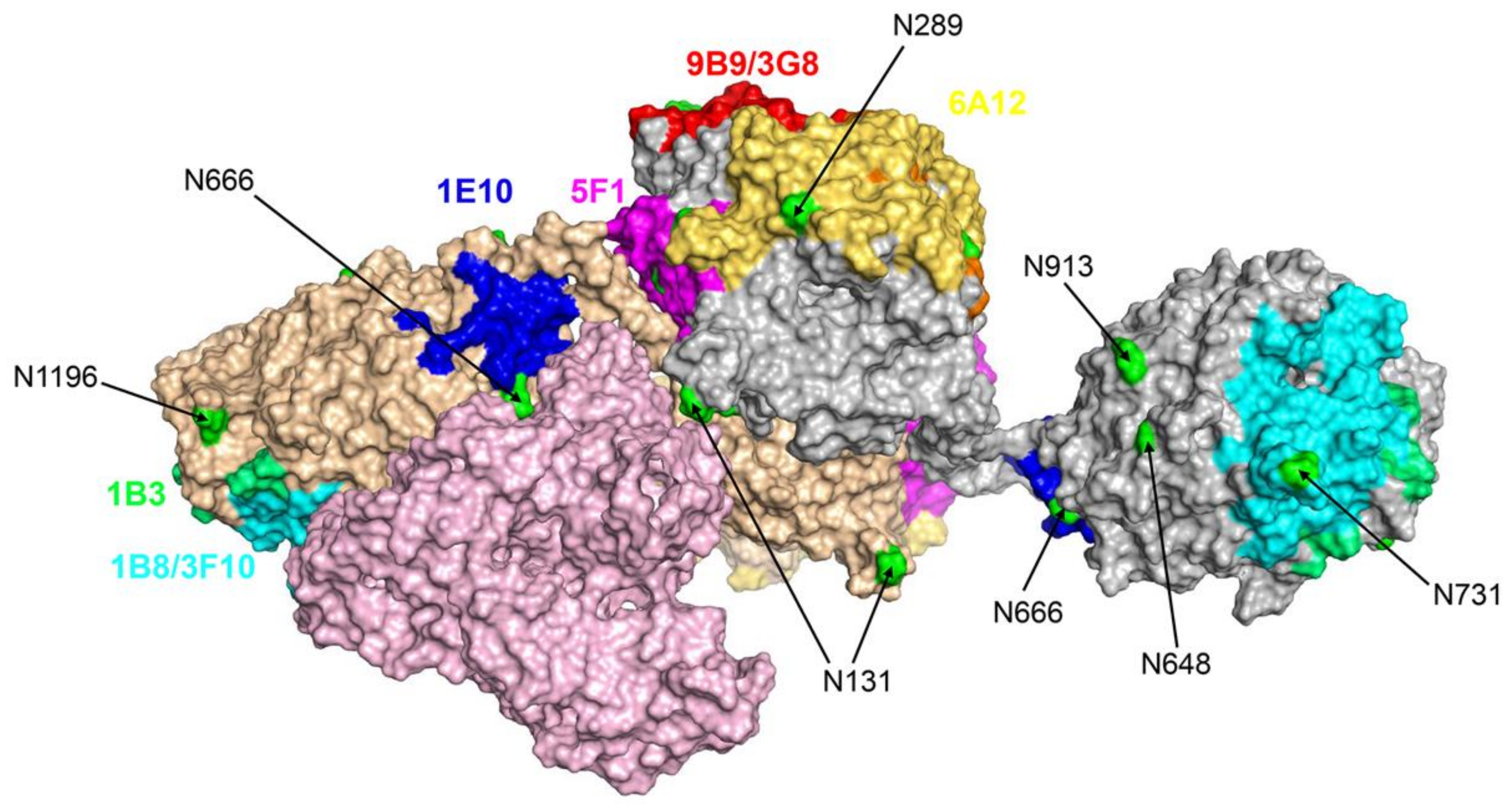
3.3. Characterization of Urinary ACE
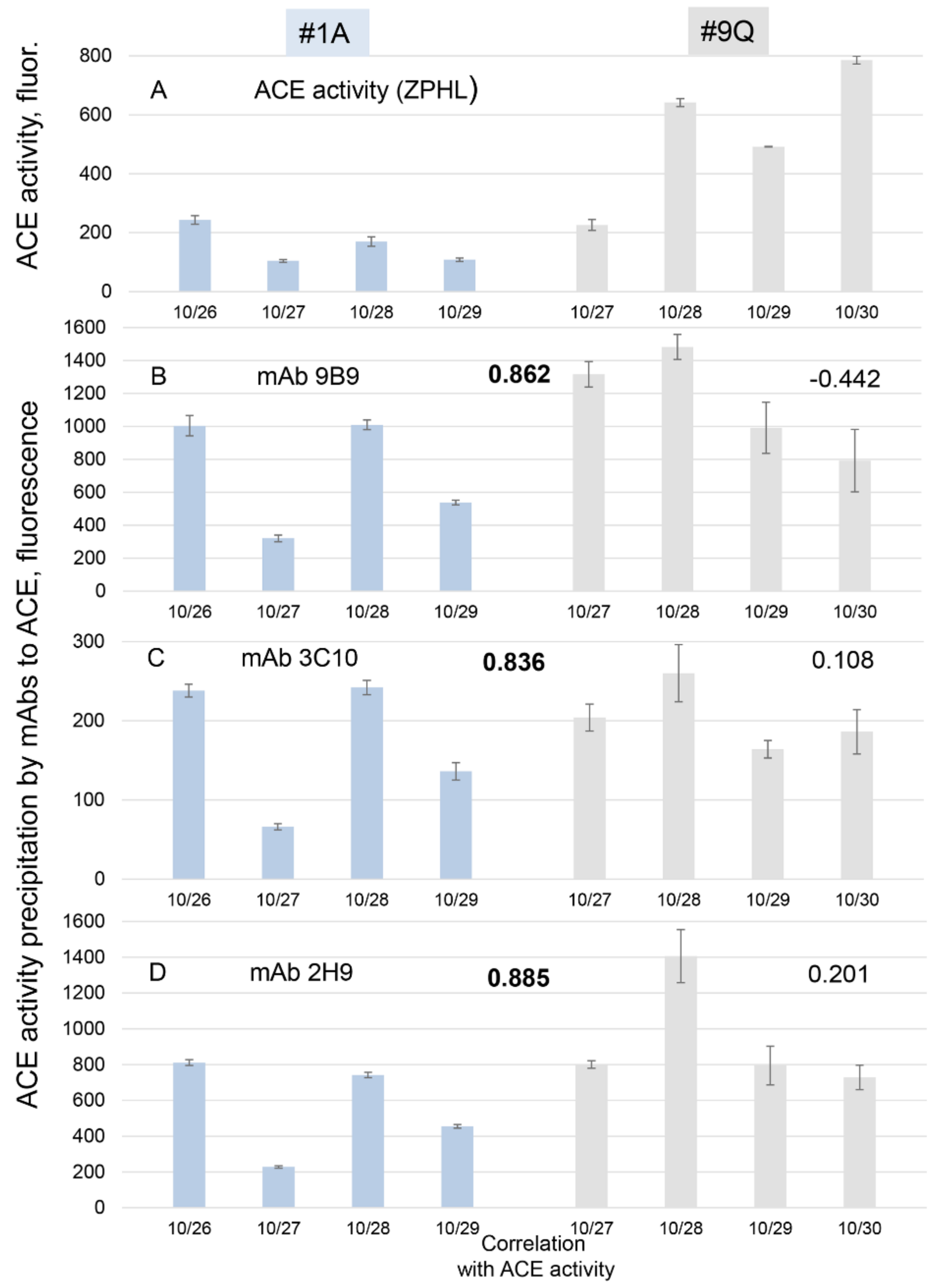
3.4. Urinary ACE as a Marker of Diet Efficiency?
3.5. Characterization of an ACE Phenotype Outlier (via Whole Exome Sequencing-WES)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sturrock, E.D.; Anthony, C.S.; Danilov, S.M. Peptidyl-dipeptidase A/Angiotensin I-converting enzyme. In Handbook of Proteolytic Enzymes, 3rd ed.; Rawlings, N., Salvesen, G., Eds.; Academic Press: Oxford, UK, 2012; Chapter 98; pp. 480–494. [Google Scholar]
- Bernstein, K.E.; Ong, F.S.; Blackwell, W.-L.B.; Shah, K.H.; Giani, J.F.; Gonzalez-Villalobos, R.A.; Shen, X.Z.; Fuchs, S. A Modern Understanding of the Traditional and Nontraditional Biological Functions of Angiotensin-Converting Enzyme. Pharmacol. Rev. 2012, 65, 1–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ching, S.F.; Hayes, L.W.; Slakey, L.L. Angiotensin-converting enzyme in cultured endothelial cells. Synthesis, degradation, and transfer to culture medium. Arteriosclerosis 1983, 3, 581–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metzger, R.; Franke, F.; Bohle, R.-M.; Alhenc-Gelas, F.; Danilov, S.M. Heterogeneous distribution of Angiotensin I-converting enzyme (CD143) in the human and rat vascular systems: Vessels, organs and species specificity. Microvasc. Res. 2011, 82, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Parkin, E.T.; Turner, A.J.; Hooper, N.M. Secretase-mediated cell surface shedding of the angiotensin-converting enzyme. Protein Pept. Lett. 2004, 11, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Alhenc-Gelas, F.; Richard, J.; Courbon, D.; Warnet, J.M.; Corvol, P. Distribution of plasma angiotensin I-converting enzyme levels in healthy men: Relationship to environmental and hormonal parameters. J. Lab. Clin. Med. 1991, 117, 33–39. [Google Scholar] [PubMed]
- Romer, F.K. Clinical and biochemical aspects of sarcoidosis. With special reference to angiotensin-converting enzyme (ACE). Acta Med. Scand. Suppl. 1984, 690, 3–96. [Google Scholar]
- Caldwell, P.R.; Seegal, B.C.; Hsu, K.C.; Das, H.; Soffer, R.L. Angiotensin-converting enzyme: Vascular endothelial localization. Science 1976, 191, 1050–1051. [Google Scholar] [CrossRef]
- Defendini, R.; Zimmerman, E.A.; Weare, J.A.; Alhenc-Gelas, F.; Erdos, E.G. Angiotensin-converting enzyme in epithelial and neuroepithelial cells. Neuroendocrinology 1983, 37, 32–40. [Google Scholar] [CrossRef]
- Metzger, R.; Bohle, R.-M.; Kerkman, L.; Pauls, K.; Eichner, G.; Alhenc-Gelas, F.; Franke, F.E.; Danilov, S.M. Distribution of angiotensin I-converting enzyme (CD 143) in the normal human kidney and in non-neoplastic kidney diseases. Kidney Int. 1999, 56, 1442–1454. [Google Scholar] [CrossRef] [Green Version]
- Casarini, D.E.; Boim, M.A.; Stella, R.C.; Krieger-Azzolini, M.H.; Krieger, J.E.; Schor, N. Angiotensin I-converting enzyme activity in tubular fluid along the rat nephron. Am. J. Physiol. Ren. Physiol. 1997, 272, F405–F409. [Google Scholar] [CrossRef]
- Kryukova, O.V.; Tikhomirova, V.E.; Golukhova, E.Z.; Evdokimov, V.V.; Kalantarov, G.F.; Trakht, I.N.; Schwartz, D.E.; Dull, R.O.; Gusakov, A.V.; Uporov, I.V.; et al. Tissue Specificity of Human Angiotensin I-Converting Enzyme. PLoS ONE 2015, 10, e0143455. [Google Scholar] [CrossRef] [Green Version]
- Danilov, S.M. Conformational Fingerprinting Using Monoclonal Antibodies (on the Example of Angiotensin I-Converting Enzyme-ACE). Mol. Biol. 2017, 51, 1046–1061. [Google Scholar] [CrossRef]
- Kokubu, T.; Kato, I.; Nishimura, K.; Hiwada, K.; Ueda, E. Angiotensin I-converting enzyme in human urine. Clin. Chim. Acta 1978, 89, 375–379. [Google Scholar] [CrossRef]
- Ryan, J.W.; Martin, L.C.; Chung, A.; Pena, G.A. Mammalian inhibitors of angiotensin converting enzyme (kininase II). Adv. Exp. Med. Biol. 1979, 120, 599–606. [Google Scholar]
- Baggio, B.; Favaro, S.; Cantaro, S.; Bertazzo, L.; Frunzio, A.; Borsatti, A. Increased urine angiotensin I converting enzyme activity in patients with upper urinary tract infection. Clin. Chim. Acta 1981, 109, 211–218. [Google Scholar] [CrossRef]
- Gupta, A.; Madhavan, M.V.; Sehgal, K.; Nair, N.; Mahajan, S.; Sehrawat, T.S.; Bikdeli, B.; Ahluwalia, N.; Ausiello, J.C.; Wan, E.Y.; et al. Extrapulmonary manifestations of COVID-19. Nat. Med. 2020, 26, 1017–1032. [Google Scholar] [CrossRef]
- Diao, B.; Wang, C.; Wang, R.; Feng, Z.; Zhang, J.; Yang, H.; Tan, Y.; Wang, H.; Wang, C.; Liu, L.; et al. Human kidney is a target for novel severe acute respiratory syndrome coronavirus 2 infection. Nat. Commun. 2021, 12, 2506. [Google Scholar] [CrossRef]
- Baudin, B.; Bénéteau-Burnat, B.; Baumann, F.C.; Giboudeau, J. A reliable radiometric assay for the determination of angiotensin I-converting enzyme activity in urine. J. Clin. Chem. Clin. Biochem. 1990, 28, 857–861. [Google Scholar] [CrossRef]
- Kato, I.; Takada, Y.; Nishimura, K.; Hiwada, K.; Kokubu, T. Increased urinary excretion of angiotensin converting enzyme in patients with renal diseases. J. Clin. Chem. Clin. Biochem. 1982, 20, 473–476. [Google Scholar] [CrossRef] [Green Version]
- Hosojima, H.; Miyauchi, E.; Morimoto, S. Urinary excretion of angiotensin-converting enzyme in NIDDM. Diabetes Care 1989, 12, 580–582. [Google Scholar] [CrossRef]
- Casarini, D.E.; Plavinik, F.L.; Zanella, M.T.; Marson, O.; Krieger, J.E.; Hirata, I.Y.; Stella, R.C.R. Angiotensin converting enzymes from human urine of mild hypertensive untreated patients resemble the N-terminal fragment of human angiotensin I-converting enzyme. Int. J. Biochem. Cell Biol. 2001, 33, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Danilov, S.M.; Balyasnikova, I.V.; Albrecht, R.F., II; Kost, O.A. Simultaneous determination of ACE activity with two substrates provide an information on the nativity of somatic ACE and allow to detect ACE inhibitors in the human blood. J. Cardiovasc. Pharmacol. 2008, 52, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Samokhodskaya, L.M.; Jain, M.S.; Kurilova, O.V.; Bobkov, A.P.; Kamalov, A.A.; Dudek, S.M.; Danilov, S.M. Phenotyping of angiotensin-converting enzyme in blood: A necessary approach for precision medicine. J. Appl. Lab. Med. 2021, 6, 1179–1191. [Google Scholar] [CrossRef] [PubMed]
- Danilov, S.M.; Balyasnikova, I.V.; Danilova, A.S.; Naperova, I.A.; Arablinskaya, N.E.; Borisov, S.E.; Metzger, R.; Franke, F.E.; Schwartz, D.E.; Gachok, I.V.; et al. Conformational fingerprinting of the angiotensin-converting enzyme (ACE): Application in sarcoidosis. J. Proteome. Res. 2010, 9, 5782–5793. [Google Scholar] [CrossRef] [PubMed]
- Popova, I.A.; Lubbe, L.; Petukhov, P.A.; Kalantarov, G.F.; Trakht, I.N.; Chernykh., E.R.; Leplina, O.Y.; Lyubimov, A.V.; Garcia, J.G.N.; Dudek, S.M.; et al. Epitope mapping of novel mAbs to human ACE. Protein Sci. 2021, 30, 1577–1593. [Google Scholar] [CrossRef]
- Bánhegyi, V.; Enyedi, A.; Fülöp, G.Á.; Oláh, A.; Siket, I.M.; Váradi, C.; Bottyán, K.; Lódi, M.; Csongrádi, A.; Umar, A.J.; et al. Human Tissue Angiotensin Converting Enzyme (ACE) Activity Is Regulated by Genetic Polymorphisms, Posttranslational Modifications, Endogenous Inhibitors and Secretion in the Serum, Lungs and Heart. Cells 2021, 10, 1708. [Google Scholar] [CrossRef]
- Danilov, S.; Savoie, F.; Lenoir, B.; Jeunemaitre, X.; Azizi, M.; Tarnow, L.; Alhenc-Gelas, F. Development of enzyme-linked immunoassays for human angiotensin I-converting enzyme suitable for large-scale studies. J. Hypertens. 1996, 14, 719–727. [Google Scholar] [CrossRef]
- Danilov, S.M.; Gordon, K.; Nesterovitch, A.B.; Lünsdorf, H.; Chen, Z.; Castellon, M.; Popova, I.A.; Kalinin, S.; Mendonca, E.; Petukhov, P.A.; et al. Angiotensin I-converting enzyme mutation (Y465D) cause dramatic increase in blood ACE via accelerated ACE shedding due to changes of ACE dimerization. PLoS ONE 2011, 6, e25952. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, A.A.; Curry, S.; Franks, N.P. Binding of the general anesthetics propofol and halothane to human serum albumin. High resolution crystal structures. J. Biol. Chem. 2000, 275, 38731–38738. [Google Scholar] [CrossRef] [Green Version]
- Gerber, P.R.; Muller, K. MAB, a generally applicable molecular force field for structure modelling in medicinal chemistry. J. Comput. Aided Mol. Des. 1995, 9, 251–268. [Google Scholar] [CrossRef]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. Ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. J. Chem. Theor. Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [Green Version]
- Schrodinger, Inc. The PyMOL Molecular Graphics System, Version 2.5.2; Schrodinger, Inc.: New York, NY, USA, 2021. [Google Scholar]
- Mizuno, K.; Toki, T.; Gotoh, M.; Matsui, J.; Yaginuma, K.; Fukuchi, K. A clinical study on the angiotensin I-converting enzyme in human urine. (II) Excretion in hypertensive patients with special reference to the renin-angiotensin-aldosterone system. Nihon Naibunpi Gakkai Zasschi 1983, 59, 45–52. [Google Scholar]
- Cidl, K.; Strelcová, L.; Znojil, V.; Váchi, J. Angiotensin I-converting enzyme (ACE) poly-mor-phism and ABO blood groups as factors codetermining plasma ACE activity. Exp. Hem. 1996, 24, 790–794. [Google Scholar]
- Zhang, Y.F.; Cheng, Q.; Tang, N.L.; Chu, T.T.W.; Tomlinson, B.; Liu, F.; Kwok, T.C.Y. Gender difference of serum angiotensin-converting enzyme (ACE) activity in DD genotype of ACE insertion/ deletion polymorphism in elderly Chinese. J. Renin Angiotensin Aldosterone Syst. 2014, 15, 547–552. [Google Scholar] [CrossRef] [Green Version]
- Oshima, G.; Gecse, A.; Erdös, E.G. Angiotensin I-converting enzyme of the kidney cortex. Biochim. Biophys. Acta 1974, 350, 26–37. [Google Scholar] [CrossRef]
- Maack, T.; Johnson, V.; Kau, S.T.; Figueiredo, J.; Sigulem, D. Renal filtration, transport, and metabolism of low-molecular-weight proteins: A review. Kidney Int. 1979, 16, 251–270. [Google Scholar] [CrossRef] [Green Version]
- Elias, M.M.; Comin, E.J.; Garay, E.A. Renal clearance of unbound bilirubin in the rat. Clin. Sci. Mol. Med. 1977, 53, 193–196. [Google Scholar] [CrossRef]
- Danilov, S.M.; Lunsdorf, H.; Akinbi, H.T.; Nesterovitch, A.B.; Epshtein, Y.; Letsiou, E.; Kryukova, O.V.; Piegeler, T.; Golukhova, E.Z.; Schwartz, D.E.; et al. Lysozyme and bilirubin bind to ACE and regulate its conformation and shedding. Sci. Rep. 2016, 6, 34913. [Google Scholar] [CrossRef] [Green Version]
- Danilov, S.M.; Wade, M.S.; Schwager, S.L.; Douglas, R.G.; Nesterovitch, A.B.; Popova, I.A.; Hogarth, K.D.; Bhardwaj, N.; Schwartz, D.E.; Sturrock, E.D.; et al. A novel angiotensin I-converting enzyme mutation (S333W) impairs N-domain enzymatic cleavage of the anti-fibrotic peptide, AcSDKP. PLoS ONE 2014, 9, e88001. [Google Scholar] [CrossRef] [Green Version]
- Deddish, P.A.; Wang, J.; Michel, B.; Morris, P.W.; Davidson, N.O.; Skidgel, R.A.; Erdös, E.G. Naturally occurring active N-domain of human angiotensin I-converting enzyme. Proc. Natl. Acad. Sci. USA 1994, 91, 7807–7811. [Google Scholar] [CrossRef] [Green Version]
- Sturrock, E.D.; Danilov, S.M.; Riordan, J.F. Limited proteolysis of human kidney angiotensin-converting enzyme and generation of catalytically active N- and C-terminal domains. Biochem. Biophys. Res. Commun. 1997, 236, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Danilov, S.; Jaspard, E.; Churakova, T.; Towbin, H.; Savoie, F.; Wei, L.; Alhenc-Gelas, F. Structure-function analysis of angiotensin I-converting enzyme using monoclonal antibodies. J. Biol. Chem. 1994, 269, 26806–26814. [Google Scholar] [CrossRef] [PubMed]
- Gordon, K.; Balyasnikova, I.V.; Nesterovitch, A.B.; Schwartz, D.E.; Sturrock, E.D.; Danilov, S.M. Fine epitope mapping of monoclonal antibodies 9B9 and 3G8, to the N domain of human angiotensin I-converting enzyme (ACE) defines a region involved in regulating ACE dimerization and shedding. Tissue Antigens 2010, 75, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Danilov, S.M.; Tikhomirova, V.E.; Kryukova, O.V.; Balatsky, A.V.; Bulaeva, N.I.; Golukhova, E.Z.; Bokeria, L.A.; Samokhodskaya, L.M.; Kost, O.A. Conformational fingerprint of blood and tissue ACEs: Personalized approach. PLoS ONE 2018, 13, e0209861. [Google Scholar] [CrossRef] [Green Version]
- Arcolino, F.O.; Tort Piella, A.; Papadimitriou, E.; Bussolati, B.; Antonie, D.J.; Murray, P.; van den Heuvel, L.; Levtchenko, E. Human Urine as a Noninvasive Source of Kidney Cells. Stem Cells Int. 2015, 2015, 362562. [Google Scholar]
- Goerlich, N.; Brand, H.A.; Langhans, V.; Tesch, S.; Schachtner, T.; Koch, B.; Paliege, A.; Schneider, W.; Grützkau, A.; Reinke, P.; et al. Kidney transplant monitoring by urinary flow cytometry: Biomarker combination of T cells, renal tubular epithelial cells, and podocalyxin-positive cells detects rejection. Sci. Rep. 2020, 10, 796. [Google Scholar] [CrossRef] [Green Version]
- Kost, O.A.; Tikhomirova, V.E.; Kryukova, O.V.; Gusakov, A.V.; Bulaeva, N.I.; Evdokimov, V.V.; Golukhova, E.Z.; Danilov, S.M. Conformational fingerprint of angiotensin-converting enzyme. Russ. J. Bioorg. Chem. 2018, 44, 48–60. [Google Scholar] [CrossRef]
- Rubtsova, K.; Marrack, P.; Rubtsov, A.V. Sexual dimorphism in autoimmunity. J. Clin. Investig. 2015, 125, 2187–2193. [Google Scholar] [CrossRef] [Green Version]
- Ercan, A.; Kohrt, W.M.; Cui, J.; Deane, K.D.; Pezer, M.; Yu, E.W.; Hausmann, J.S.; Campbell, H.; Kaiser, U.B.; Rudd, P.M.; et al. Estrogens regulate glycosylation of IgG in women and men. JCI Insight 2017, 2, e89703. [Google Scholar] [CrossRef]
- Qin, Y.; Zhong, Y.; Zhu, M.; Dang, L.; Yu, H.; Chen, Z.; Chen, W.; Wang, X.; Zhang, H.; Li, Z. Age- and sex-associated differences in the glycopatterns of human salivary glycoproteins and their role against influenza A virus. J. Proteome Res. 2013, 12, 2742–2754. [Google Scholar] [CrossRef]
- Reiding, K.R.; Hipgrave-Ederveen, A.L.; Rombouts, Y.; Wuhrer, M. Murine Plasma N-Glycosylation Traits Associated with Sex and Strain. J. Proteome Res. 2016, 15, 3489–3499. [Google Scholar] [CrossRef]
- Dotz, V.; Wuhrer, M. N-glycome signatures in human plasma: Associations with physiology and major diseases. FEBS Lett. 2019, 593, 2966–2976. [Google Scholar] [CrossRef] [Green Version]
- Zelanis, A.; Menezes, M.C.; Kitano, E.S.; Liberato, T.; Tashima, A.K.; Pinto, A.F.M.; Sherman, N.E.; Ho, P.L.; Fox, J.W.; Serrano, S.M.T. Proteomic identification of gender molecular markers in Bothrops jararaca venom. J. Proteomics 2016, 139, 26–37. [Google Scholar] [CrossRef]
- Scheys, F.; Van Damme, E.J.M.; Pauwels, J.; Staes, A.; Gevaert, K.; Smagghe, G. N-glycosylation Site Analysis Reveals Sex-related Differences in Protein N-glycosylation in the Rice Brown Planthopper (Nilaparvata lugens). Mol. Cell. Proteomics 2020, 19, 529–539. [Google Scholar] [CrossRef]
- Herak-Kramberger, C.M.; Breljak, D.; Ljubojević, M.; Matokanović, M.; Lovrić, M.; Rogić, D.; Brzica, H.; Vrhovac, I.; Karaica, D.; Micek, V.; et al. Sex-dependent expression of water channel AQP1 along the rat nephron. Am. J. Physiol. Renal Physiol. 2015, 308, F809–F821. [Google Scholar] [CrossRef] [Green Version]
- Morris, H.R.; Dell, A.; Easton, R.L.; Panico, M.P.; Koistinen, H.; Koistinen, R.; Oehninger, S.; Patankar, M.S.; Seppala, M.; Clark, G.F. Gender-specific glycosylation of human glycodelin affects its contraceptive activity. J. Biol. Chem. 1996, 271, 32159–32167. [Google Scholar] [CrossRef] [Green Version]
- Shang, S.; Dong, H.; Li, Y.; Zhang, W.; Li, H.; Qin, W.; Qian, X. Large-scale enrichment and identification of human urinary N-glycoproteins/N-glycopeptides. Se Pu Chin. J. Chromatogr. 2021, 39, 686–694. [Google Scholar] [CrossRef]
- Das, M.; Hartley, J.L.; Soffer, R.L. Serum angiotensin-converting enzyme. Isolation and relationship to the pulmonary enzyme. J. Biol. Chem. 1997, 252, 1316–1319. [Google Scholar] [CrossRef]
- Ashwell, G.; Harford, J. Carbohydrate-specific receptors of the liver. Annu. Rev. Biochem. 1982, 51, 531–554. [Google Scholar] [CrossRef]
- Kobori, H.; Nangaku, M.; Navar, L.G.; Nishiyama, A. The intrarenal renin–angiotensin system: From physiology to the pathobiology of hypertension and kidney disease. Pharmacol. Rev. 2007, 59, 251–287. [Google Scholar] [CrossRef]
- Giani, J.F.; Veiras, L.C.; Shen, J.Z.Y.; Bernstein, E.A.; Cao, D.; Okwan-Duodu, D.; Khan, Z.; Gonzalez-Villalobos, R.A.; Bernstein, K.E. Novel roles of the renal angiotensin-converting enzyme. Mol. Cell. Endocrinol. 2021, 529, 111257. [Google Scholar] [CrossRef] [PubMed]
- Soubrier, F.; Alhenc-Gelas, F.; Hubert, C.; Allegrini, J.; John, M.; Tregear, G.; Corvol, P. Two putative active centers in human angiotensin I-converting enzyme revealed by molecular cloning. Proc. Natl. Acad. Sci. USA 1988, 85, 9386–9390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watermeyer, J.M.; Sewell, B.T.; Schwager, S.L.; Natesh, R.; Corradi, H.R.; Archarya, K.R.; Sturrock, E.D. Structure of testis ACE glycosylation mutants and evidence for conserved domain movement. Biochemistry 2006, 45, 12654–12663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, H.G.; Redelinghuys, P.; Schwager, S.L.; Sturrock, E.D. The role of glycosylation and domain interactions in the thermal stability of human angiotensin-converting enzyme. Biol. Chem. 2008, 389, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Anthony, C.S.; Corradi, H.R.; Schwager, S.L.; Redelinghuys, P.; Georgiadis, D.; Dive, V.; Acharya, K.R.; Sturrock, E.D. The N domain of human angiotensin-I-converting enzyme: The role of N-glycosylation and the crystal structure in complex with an N domain-specific phosphinic inhibitor, RXP407. J. Biol. Chem. 2010, 285, 35685–35693. [Google Scholar] [CrossRef] [Green Version]
- Cuddy, L.K.; Prokopenko, D.; Cunningham, E.P.; Brimberry, R.; Song, P.; Kirchner, R.; Chapman, B.A.; Hofmann, O.; Hide, W.; Procissi, D.; et al. Aβ-accelerated neurodegeneration caused by Alzheimer’s-associated ACE variant R1279Q is rescued by angiotensin system inhibition in mice. Sci. Transl. Med. 2020, 12, eaaz2541. [Google Scholar] [CrossRef]
- Navratilova, E.; Fillingim, R.B.; Porreca, F. Sexual dimorphism in functional pain syndromes. Sci. Transl. Med. 2021, 13, eabj7180. [Google Scholar] [CrossRef]
- Fusayasu, E.; Kowa, H.; Takeshima, T.; Nakaso, K.; Nakashima, K. Increased plasma substance P and CGRP levels, and high ACE activity in migraineurs during headache-free periods. Pain 2007, 128, 209–214. [Google Scholar] [CrossRef] [Green Version]
- König, S.; Engl, C.; Bayer, M.; Escolano-Lozano, F.; Rittner, H.; Rebhorn, C.; Birklein, F. Substance P serum degradation in complex regional pain syndrome-another piece of the puzzle? J. Pain 2022, 23, 501–507. [Google Scholar] [CrossRef]
- Tikhomirova, V.E.; Kost, O.A.; Kryukova, O.V.; Golukhova, E.Z.; Bulaeva, N.I.; Zholbaeva, A.Z.; Bokeria, L.A.; Garcia, J.G.; Danilov, S.M. ACE phenotyping in human heart. PLoS ONE 2017, 12, e0181976. [Google Scholar] [CrossRef] [Green Version]
- Danilov, S.M.; Tikhomirova, V.E.; Metzger, R.; Naperova, I.A.; Bukina, T.M.; Goker-Alpan, O.; Tayebi, N.; Gayfullin, N.M.; Schwartz, D.E.; Samokhodskaya, L.M.; et al. ACE phenotyping in Gaucher disease. Mol. Genet. Metab. 2018, 123, 501–510. [Google Scholar] [CrossRef]
- Poortmans, J.; Jeanloz, R.W. Quantitative immunological determination of 12 plasma proteins excreted in human urine collected before and after exercise. J. Clin. Investig. 1968, 47, 386–393. [Google Scholar] [CrossRef] [Green Version]
- Klauser, R.J.; Robinson, C.J.; Marinkovic, D.V.; Erdös, E.G. Inhibition of human peptidyl dipeptidase (angiotensin I converting enzyme: Kininase II) by human serum albumin and its fragments. Hypertension 1979, 1, 281–286. [Google Scholar] [CrossRef] [Green Version]
- Fagyas, M.; Úri, K.; Siket, I.M.; Fülöp, G.Á.; Csató, V.; Daragó, A.; Boczán, J.; Bányai, E.; Szentkirályi, I.E.; Maros, T.M.; et al. New perspectives in the renin-angiotensin-aldosterone system (RAAS) II: Albumin suppresses angiotensin converting enzyme (ACE) activity in human. PLoS ONE 2014, 9, e87844. [Google Scholar] [CrossRef] [Green Version]
- Peterson, P.A.; Evrin, P.E.; Berggård, I. Differentiation of glomerular, tubular, and normal proteinuria: Determinations of urinary excretion of beta-2-macroglobulin, albumin, and total protein. J. Clin. Investig. 1969, 48, 1189–1198. [Google Scholar] [CrossRef] [Green Version]
- Ching, S.-F.; Hayes, L.W.; Slakey, L.L. Angiotensin converting enzyme in cultured endothelial cells and growth medium. Relationships to enzyme from kidney and plasma. Biochim. Biophys. Acta 1981, 651, 222–231. [Google Scholar] [CrossRef]
- Nakamura, Y.; Takeda, T.; Ishii, M.; Nishiyama, K.; Yamakada, M.; Hirata, Y.; Kimura, K.; Murao, S. Elevation of serum angiotensin-converting enzyme activity in patients with hyperthyroidism. J. Clin. Endocrinol. Metab. 1982, 55, 931–934. [Google Scholar] [CrossRef]
- Jalil, J.E.; Ocaranza, M.P.; Piddo, A.M.; Jalil, R. Reproducibility of plasma angiotensin-converting enzyme activity in human subjects determined by fluorimetry with Z-phenylalanine-histidyl-leucine as substrate. J. Lab. Clin. Med. 1999, 133, 501–506. [Google Scholar] [CrossRef]
- Wang, P.; Holst, C.; Wodzig, W.K.; Andersen, M.R.; Astrup, A.; van Baak, M.A.; Larsen, T.M.; Jebb, S.B.; Kafatos, A.; Pfeiffer, A.F.; et al. Circulating ACE is a predictor of weight loss maintenance not only in overweight and obese women, but also in men. Int. J. Obes. 2012, 36, 1545–1551. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Holst, C.; Andersen, M.R.; Astrup, A.; Bouwman, F.G.; van Otterdijk, S.; Wodzig, W.K.; van Baak, M.A.; Larsen, T.M.; Jebb, S.A.; et al. Blood profile of proteins and steroid hormones predicts weight change after weight loss with interactions of dietary protein level and glycemic index. PLoS ONE 2011, 6, e16773. [Google Scholar]
- Tejpal, S.; Sanghera, N.; Manoharan, V.; Planas-Iglesias, J.; Bastie, C.C.; Klein-Seetharaman, J. Angiotensin Converting Enzyme (ACE): A Marker for Personalized Feedback on Dieting. Nutrients 2020, 12, 660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiser, J. The Hunt for Missing Genes. Science 2014, 344, 687–689. [Google Scholar] [CrossRef] [PubMed]
- Santhamma, K.R.; Sen, I. Specific cellular proteins associated with angiotensin-converting enzyme and regulate its intracellular transport and cleavage-secretion. J. Biol. Chem. 2000, 275, 23253–23258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danilov, S.M.; Jain, M.S.; Petukhov, P.A.; Goldman, C.; DiSanto-Rose, M.; Vancavage, R.; Francuzevitch, L.Y.; Samokhodskaya, L.M.; Kamalov, A.A.; Arbieva, Z.H.; et al. Novel ACE mutations mimicking sarcoidosis by increasing blood ACE levels. Transl. Res. 2021, 230, 5–20. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozuch, A.J.; Petukhov, P.A.; Fagyas, M.; Popova, I.A.; Lindeblad, M.O.; Bobkov, A.P.; Kamalov, A.A.; Toth, A.; Dudek, S.M.; Danilov, S.M. Urinary ACE Phenotyping as a Research and Diagnostic Tool: Identification of Sex-Dependent ACE Immunoreactivity. Biomedicines 2023, 11, 953. https://doi.org/10.3390/biomedicines11030953
Kozuch AJ, Petukhov PA, Fagyas M, Popova IA, Lindeblad MO, Bobkov AP, Kamalov AA, Toth A, Dudek SM, Danilov SM. Urinary ACE Phenotyping as a Research and Diagnostic Tool: Identification of Sex-Dependent ACE Immunoreactivity. Biomedicines. 2023; 11(3):953. https://doi.org/10.3390/biomedicines11030953
Chicago/Turabian StyleKozuch, Alexander J., Pavel A. Petukhov, Miklos Fagyas, Isolda A. Popova, Matthew O. Lindeblad, Alexander P. Bobkov, Armais A. Kamalov, Attila Toth, Steven M. Dudek, and Sergei M. Danilov. 2023. "Urinary ACE Phenotyping as a Research and Diagnostic Tool: Identification of Sex-Dependent ACE Immunoreactivity" Biomedicines 11, no. 3: 953. https://doi.org/10.3390/biomedicines11030953