
Bacteria-Mediated Modulatory Strategies for Colorectal Cancer Treatment




Abstract
:1. Introduction
2. Bacteriotherapy in CRC Treatment
2.1. Mechanisms Used in Bacteriotherapy in CRC
2.1.1. Bacteriotherapy for Modulating Innate Immunity
2.1.2. Bacterial Peptides

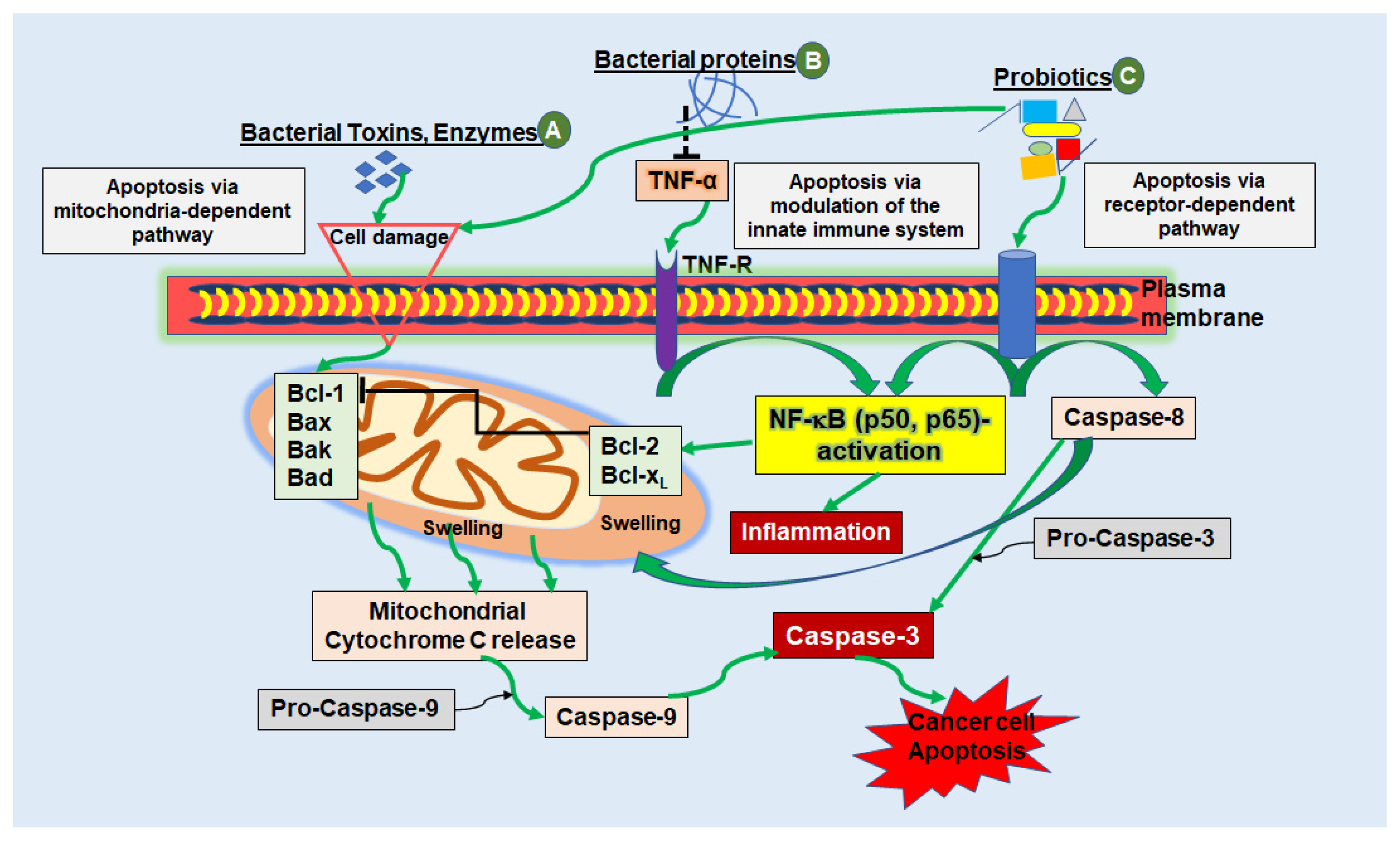{kind=link}
| Protein/Peptides | Bacterial Source | Mode of Action | Refs. |
|---|---|---|---|
| Enterotoxin (CPE) | Clostridiumperfringens | Binds to Claudin-3/-4 surface receptors and leads to CRC cell lysis in SW480, HCT-116, SW620, Caco-2, HT-29 and PDX CRC cells. | [62,63] |
| Induces TNF-α-upregulation leading to decreased Claudin-4 expression, disrupted tight junctions, reduced EMT-, adherence- and metastasis-capacity in HT-29 CRC cells. | [93] | ||
| Diphteria toxin (DT) | Corynebacterium diphteria | Subunit A blocks protein synthesis by ADP-ribosylation of EF-2 leading to cell death. | [65,66] |
| Non-toxic CRM197 suppresses angiogenesis and proliferation in SW480, SW620, HCT-116, Caco-2 and HT-29 CRC cells. | [67,68] | ||
| Nisin | Lactococcus lactis | Leads to apoptosis via promoting hole formation in the cell membrane of Caco-2 and HT-29 CRC cells. | [69] |
| Prevents expression of MMP-2, MMP-9, CDTs and Cif in SW480, HT-29, Caco-2 and LS-180 CRC cells. | [70,71] | ||
| Increases cytotoxicity of anti-cancer agents resulting in lower doses necessary for treatment. | [70] | ||
| Colicin | Escherichiacoli | Acts cytotoxically through membrane hole formation, and non-specified DN/RNase activity in HT-29 CRC cells. | [72,73] |
| Subunits A, E1, E3, U inhibit cell growth and promote apoptosis in HT-29 CRC cells. | [73,74] | ||
| Microcin | Klebsiella pneumonia | Pore forming into cell membranes, thus leading to apoptosis in CRC cells and other cancer cell lines such as HeLa. | [75,76] |
| Subunit E492 shows a noticeable cytotoxicity especially in HT-29 but also in SW620 CRC cells. | [94] | ||
| Subunit E492 reduces tumor proliferation in a xenograft model with SW620 CRC cells. | [94] | ||
| Pediocin | Pediococcus acidilactici | Shows lethal effects on HT-29 and DLD-1 CRC cells. | [78] |
| Subunit PA-1 interoperates with TLRs and initiates cell death. | [79] | ||
| Inhibits cancer cell proliferation as a carrier combination. | [95] | ||
| Phenazine | Pseudomonas aeruginosa | Subtype PDC weakens viability and DNA synthesis and initiates cell cycle arrest leading to apoptosis in HT-29 CRC cells. | [80,81] |
| Azurin | Pseudomonas aeruginosa | Influences p53/EphB2/VEGFR-2 signaling pathway and prevents angiogenesis in CRC cells. | [26,84] |
| Inhibits cancer cell mobility and shows strong anti-cancer effect in HCT-116 CRC cells. | [96] | ||
| Exotoxin A (PE) | Pseudomonas aeruginosa | Inhibits EF-2 and protein biosynthesis via ADP-ribosylation, induces apoptosis in HCT116, HT-29 and COLO320 CRC cells. | [86,87,89] |
| Subtype PE24-based amyloid injection leads to growth arrest and metastasis prevention in CRC-diseased mice. | [97] | ||
| Listeriolysin O (LLO) | Listeria monocytogenes | Acts as a membrane-damaging cytotoxin in Caco-2 CRC cells. | [98] |
| Shows hemolytic activity and anti-tumor properties in Colo205 CRC cells. | [90,92] |
| Protein/Peptides | Bacterial Source | Mode of Action | Refs. |
|---|---|---|---|
| Lucentamycin | Nocardiopsis lucentensis | Shows cytotoxicity and induces apoptosis against HCT-116 CRC cells. | [99] |
| Arenamides | Salinispora arenicola | Subtypes A and B block TNF, nitric oxide and prostaglandin E2 and act cytotoxic on HCT-116 CRC cells. Investigation for chemopreventive and anti-inflammatory properties in HCT-116 CRC cells is proposed. | [100] |
| Ohmyungsamycins | Streptomyces sp. | Subtype A modulates Skp-p27 axis leading to cell cycle arrest (G0/G1 phase), apoptosis and selectively targeted reduction of proliferation in HCT-116 CRC cells. | [99,102] |
| Subtype A shows stronger activity against human cancer cells compared to subtype B. | [106] | ||
| Mixirins | Marine Bacillus sp. | Subtypes A, B and C act cytotoxic against cancer cells and inhibit cell growth in HCT-116 CRC cells. | [103] |
| Urukthapelstatin | Mechercharimyces asporophorigenens | Subtype A inhibits DNA synthesis, growth and proliferation of HCT-116 CRC cells. | [104] |
2.1.3. Bacteria as Carriers for Therapeutic Agents
2.1.4. Bacteria-Mediated Anti-Angiogenesis Therapy
2.1.5. Bacterial Biofilms
2.1.6. Maintaining Microbial Equilibrium
3. Microbiota in CRC
3.1. Influence of Microbiota on Drug Metabolism
3.2. Influence of Microbiota on Conventional CRC Therapy
4. Discussion and Perspectives
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 5-FU | 5-fluorouracil |
| Bad | Bcl-2-Antagonist of Cell Death |
| Bak | BCL2 Antagonist/Killer 1 |
| Bax | Bcl-2-associated X protein |
| Bcl-1 | B-cell lymphoma 1 |
| CD | cluster of differentiation |
| CDI | Clostridium difficile infection |
| CDT | cytolethal distending toxin |
| Cif | cycle inhibiting factor |
| CPE | C. perfringens enterotoxin |
| CRC | colorectal cancer |
| DT | diphteria toxin |
| EF | elongation factor |
| FMT | fecal microbiota transplantation |
| GlcA | glucuronic acid |
| GUS | gut bacterial-glucuronidases |
| IFN | interferon |
| IL | interleukin |
| LLO | Listeriolysin O |
| MDM | microbiome-derived metabolism |
| MMP | matrix metalloproteinase |
| NF-κB | nuclear factor ‘kappa-light-chain-enhancer’ of activated B-cells |
| PCA | phenazine 1-carboxylic acid |
| PDC | phenazine 1,6-di-carboxylic acid |
| PE | Pseudomonas aeruginosa endotoxin |
| TNF | tumor necrosis factor |
| TNF-R | Tumor necrosis factor receptor |
| TLR | Toll-like receptor |
| VEGF | Rvesicular endothelial growth factor receptor |
References
- Jemal, A.; Siegel, R.; Ward, E.; Hao, Y.; Xu, J.; Murray, T.; Thun, M.J. Cancer statistics, 2008. CA 2008, 58, 71–96. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, R.M.; Rothenberg, M.L.; Van Cutsem, E.; Benson, A.B., 3rd; Blanke, C.D.; Diasio, R.B.; Grothey, A.; Lenz, H.J.; Meropol, N.J.; Ramanathan, R.K.; et al. The continuum of care: A paradigm for the management of metastatic colorectal cancer. Oncology 2007, 12, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Field, K.; Lipton, L. Metastatic colorectal cancer-past, progress and future. World J. Gastroenterol. 2007, 13, 3806–3815. [Google Scholar] [CrossRef]
- Zacharakis, M.; Xynos, I.D.; Lazaris, A.; Smaro, T.; Kosmas, C.; Dokou, A.; Felekouras, E.; Antoniou, E.; Polyzos, A.; Sarantonis, J.; et al. Predictors of survival in stage IV metastatic colorectal cancer. Anticancer Res. 2010, 30, 653–660. [Google Scholar] [PubMed]
- Ambalam, P.; Raman, M.; Purama, R.K.; Doble, M. Probiotics, prebiotics and colorectal cancer prevention. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef]
- Buhrmann, C.; Shayan, P.; Goel, A.; Shakibaei, M. Resveratrol Regulates Colorectal Cancer Cell Invasion by Modulation of Focal Adhesion Molecules. Nutrients 2017, 9, 1073. [Google Scholar] [CrossRef]
- Azadi, A.; Golchini, A.; Delazar, S.; Abarghooi Kahaki, F.; Dehnavi, S.M.; Payandeh, Z.; Eyvazi, S. Recent Advances on Immune Targeted Therapy of Colorectal Cancer Using bi-Specific Antibodies and Therapeutic Vaccines. Biol. Proced. Online 2021, 23, 13. [Google Scholar] [CrossRef] [PubMed]
- Sambi, M.; Haq, S.; Samuel, V.; Qorri, B.; Haxho, F.; Hill, K.; Harless, W.; Szewczuk, M.R. Alternative therapies for metastatic breast cancer: Multimodal approach targeting tumor cell heterogeneity. Breast Cancer 2017, 9, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, B.B.; Shishodia, S.; Sandur, S.K.; Pandey, M.K.; Sethi, G. Inflammation and cancer: How hot is the link? Biochem. Pharmacol. 2006, 72, 1605–1621. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A. Molecular pathways linking inflammation and cancer. Curr. Mol. Med. 2010, 10, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Payandeh, Z.; Khalili, S.; Somi, M.H.; Mard-Soltani, M.; Baghbanzadeh, A.; Hajiasgharzadeh, K.; Samadi, N.; Baradaran, B. PD-1/PD-L1-dependent immune response in colorectal cancer. J. Cell. Physiol. 2020, 235, 5461–5475. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Shayan, P.; Banik, K.; Kunnumakkara, A.B.; Kubatka, P.; Koklesova, L.; Shakibaei, M. Targeting NF-κB Signaling by Calebin A, a Compound of Turmeric, in Multicellular Tumor Microenvironment: Potential Role of Apoptosis Induction in CRC Cells. Biomedicines 2020, 8, 236. [Google Scholar] [CrossRef] [PubMed]
- Brenner, H.; Stock, C.; Hoffmeister, M. Effect of screening sigmoidoscopy and screening colonoscopy on colorectal cancer incidence and mortality: Systematic review and meta-analysis of randomised controlled trials and observational studies. BMJ 2014, 348, g2467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chassaing, B.; Kumar, M.; Baker, M.T.; Singh, V.; Vijay-Kumar, M. Mammalian gut immunity. Biomed. J. 2014, 37, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Duong, M.T.; Qin, Y.; You, S.H.; Min, J.J. Bacteria-cancer interactions: Bacteria-based cancer therapy. Exp. Mol. Med. 2019, 51, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzutsev, A.; Goldszmid, R.S.; Viaud, S.; Zitvogel, L.; Trinchieri, G. The role of the microbiota in inflammation, carcinogenesis, and cancer therapy. Eur. J. Immunol. 2015, 45, 17–31. [Google Scholar] [CrossRef]
- Laliani, G.; Ghasemian Sorboni, S.; Lari, R.; Yaghoubi, A.; Soleimanpour, S.; Khazaei, M.; Hasanian, S.M.; Avan, A. Bacteria and cancer: Different sides of the same coin. Life Sci. 2020, 246, 117398. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahimzadeh, S.; Ahangari, H.; Soleimanian, A.; Hosseini, K.; Ebrahimi, V.; Ghasemnejad, T.; Soofiyani, S.R.; Tarhriz, V.; Eyvazi, S. Colorectal cancer treatment using bacteria: Focus on molecular mechanisms. BMC Microbiol. 2021, 21, 218. [Google Scholar] [CrossRef] [PubMed]
- Elyasifar, B.; Jafari, S.; Hallaj-Nezhadi, S.; Chapeland-leclerc, F.; Ruprich-Robert, G.; Dilmaghani, A. Isolation and identification of antibiotic-producing halophilic bacteria from dagh biarjmand and haj aligholi salt deserts, iran. Pharm. Sci. 2019, 25, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Seyfi, R.; Kahaki, F.A.; Ebrahimi, T.; Montazersaheb, S.; Eyvazi, S.; Babaeipour, V.; Tarhriz, V. Antimicrobial peptides (AMPs): Roles, functions and mechanism of action. Int. J. Pept. Res. Ther. 2020, 26, 1451–1463. [Google Scholar] [CrossRef]
- Tarhriz, V.; Eyvazi, S.; Shakeri, E.; Hejazi, M.S.; Dilmaghani, A. Antibacterial and antifungal activity of novel freshwater bacterium Tabrizicola aquatica as a prominent natural antibiotic available in Qurugol Lake. Pharm. Sci. 2020, 26, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Arruebo, M.; Vilaboa, N.; Sáez-Gutierrez, B.; Lambea, J.; Tres, A.; Valladares, M.; González-Fernández, Á. Assessment of the evolution of cancer treatment therapies. Cancers 2011, 3, 3279–3330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gang, W.; Wang, J.J.; Guan, R.; Yan, S.; Shi, F.; Zhang, J.Y.; Li, Z.M.; Gao, J.; Fu, X.L. Strategy to targeting the immune resistance and novel therapy in colorectal cancer. Cancer Med. 2018, 7, 1578–1603. [Google Scholar] [CrossRef] [PubMed]
- Karpiński, T.M.; Adamczak, A. Anticancer activity of bacterial proteins and peptides. Pharmaceutics 2018, 10, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeffler, D.I.; Schoen, C.U.; Goebel, W.; Pilgrim, S. Comparison of different live vaccine strategies in vivo for delivery of protein antigen or antigen-encoding DNA and mRNA by virulence-attenuated Listeria monocytogenes. Infect. Immun. 2006, 74, 3946–3957. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, K.; Jain, A.; Allen, H.E.; Laird, L.S.; Chia, C.Y.; Ravi, S.; Brockstedt, D.G.; Giedlin, M.A.; Bahjat, K.S.; Leong, M.L.; et al. Selective targeting of antitumor immune responses with engineered live-attenuated Listeria monocytogenes. Cancer Res. 2006, 66, 1096–1104. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.A.; Shabahang, S.; Timiryasova, T.M.; Zhang, Q.; Beltz, R.; Gentschev, I.; Goebel, W.; Szalay, A.A. Visualization of tumors and metastases in live animals with bacteria and vaccinia virus encoding light-emitting proteins. Nat. Biotechnol. 2004, 22, 313–320. [Google Scholar] [CrossRef]
- Mengesha, A.; Dubois, L.; Chiu, R.K.; Paesmans, K.; Wouters, B.G.; Lambin, P.; Theys, J. Potential and limitations of bacterial-mediated cancer therapy. Front. Biosci. 2007, 12, 3880–3891. [Google Scholar] [CrossRef] [Green Version]
- Min, J.J.; Kim, H.J.; Park, J.H.; Moon, S.; Jeong, J.H.; Hong, Y.J.; Cho, K.O.; Nam, J.H.; Kim, N.; Park, Y.K.; et al. Noninvasive real-time imaging of tumors and metastases using tumor-targeting light-emitting Escherichia coli. Mol. Imaging Biol. 2008, 10, 54–61. [Google Scholar] [CrossRef]
- De Almeida, C.V.; de Camargo, M.R.; Russo, E.; Amedei, A. Role of diet and gut microbiota on colorectal cancer immunomodulation. World J. Gastroenterol. 2019, 25, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-Y.; Yan, Z.-B.; Meng, Y.-M.; Hong, X.-Y.; Shao, G.; Ma, J.-J.; Cheng, X.-R.; Liu, J.; Kang, J.; Fu, C.-Y. Antimicrobial peptides: Mechanism of action, activity and clinical potential. Mil. Med. Res. 2021, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Fong, W.; Li, Q.; Yu, J. Gut microbiota modulation: A novel strategy for prevention and treatment of colorectal cancer. Oncogene 2020, 39, 4925–4943. [Google Scholar] [CrossRef] [PubMed]
- Raman, M.; Ambalam, P.; Kondepudi, K.K.; Pithva, S.; Kothari, C.; Patel, A.T.; Purama, R.K.; Dave, J.; Vyas, B. Potential of probiotics, prebiotics and synbiotics for management of colorectal cancer. Gut Microbes 2013, 4, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Kucerova, P.; Cervinkova, M. Spontaneous regression of tumour and the role of microbial infection–possibilities for cancer treatment. Anti-Cancer Drugs 2016, 27, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasai, C.; Sugimoto, K.; Moritani, I.; Tanaka, J.; Oya, Y.; Inoue, H.; Tameda, M.; Shiraki, K.; Ito, M.; Takei, Y.; et al. Comparison of human gut microbiota in control subjects and patients with colorectal carcinoma in adenoma: Terminal restriction fragment length polymorphism and next-generation sequencing analyses. Oncol. Rep. 2016, 35, 325–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, C.; Kasnitz, N.; Kocijancic, D.; Trittel, S.; Riese, P.; Guzman, C.A.; Leschner, S.; Weiss, S. Induction of CD4+ and CD8+ anti-tumor effector T cell responses by bacteria mediated tumor therapy. Int. J. Cancer 2015, 137, 2019–2028. [Google Scholar] [CrossRef]
- Soleimanpour, S.; Hasanian, S.M.; Avan, A.; Yaghoubi, A.; Khazaei, M. Bacteriotherapy in gastrointestinal cancer. Life Sci. 2020, 254, 117754. [Google Scholar] [CrossRef]
- Song, S.; Vuai, M.S.; Zhong, M. The role of bacteria in cancer therapy–enemies in the past, but allies at present. Infect. Agents Cancer 2018, 13, 9. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, B.A.; El-Deiry, W.S. Targeting apoptosis in cancer therapy. Nat. Rev. Clin. Oncol. 2020, 17, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Shakibaei, M.; Schulze-Tanzil, G.; Takada, Y.; Aggarwal, B.B. Redox regulation of apoptosis by members of the TNF superfamily. Antioxid. Redox Signal. 2005, 7, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Igney, F.H.; Krammer, P.H. Death and anti-death: Tumour resistance to apoptosis. Nat. Rev. Cancer 2002, 2, 277–288. [Google Scholar] [CrossRef]
- Candeias, S.M.; Gaipl, U.S. The Immune System in Cancer Prevention, Development and Therapy. Anti-Cancer Agents Med. Chem. 2016, 16, 101–107. [Google Scholar] [CrossRef]
- Leone, R.D.; Powell, J.D. Metabolism of immune cells in cancer. Nat. Rev. Cancer 2020, 20, 516–531. [Google Scholar] [CrossRef]
- Clara, J.A.; Monge, C.; Yang, Y.; Takebe, N. Targeting signalling pathways and the immune microenvironment of cancer stem cells—A clinical update. Nat. Rev. Clin. Oncol. 2020, 17, 204–232. [Google Scholar] [CrossRef]
- Netea, M.G.; Simon, A.; van de Veerdonk, F.; Kullberg, B.-J.; Van der Meer, J.W.; Joosten, L.A. IL-1β processing in host defense: Beyond the inflammasomes. PLoS Pathog. 2010, 6, e1000661. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.H.; Kim, H.S.; Ha, J.M.; Hong, Y.; Choy, H.E.; Min, J.J. Genetically engineered Salmonella typhimurium as an imageable therapeutic probe for cancer. Cancer Res. 2010, 70, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Erdman, S.E.; Rao, V.P.; Poutahidis, T.; Ihrig, M.M.; Ge, Z.; Feng, Y.; Tomczak, M.; Rogers, A.B.; Horwitz, B.H.; Fox, J.G. CD4+ CD25+ regulatory lymphocytes require interleukin 10 to interrupt colon carcinogenesis in mice. Cancer Res. 2003, 63, 6042–6050. [Google Scholar]
- Agrawal, S.; Acharya, D.; Adholeya, A.; Barrow, C.J.; Deshmukh, S.K. Nonribosomal peptides from marine microbes and their antimicrobial and anticancer potential. Front. Pharmacol. 2017, 8, 828. [Google Scholar] [CrossRef]
- Zhao, M.; Yang, M.; Li, X.M.; Jiang, P.; Baranov, E.; Li, S.; Xu, M.; Penman, S.; Hoffman, R.M. Tumor-targeting bacterial therapy with amino acid auxotrophs of GFP-expressing Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 2005, 102, 755–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, M.E.; Lemmon, M.J.; Mauchline, M.L.; Davis, T.O.; Giaccia, A.J.; Minton, N.P.; Brown, J.M. Anaerobic bacteria as a delivery system for cancer gene therapy: In Vitro activation of 5-fluorocytosine by genetically engineered clostridia. Gene Ther. 1996, 3, 173–178. [Google Scholar]
- Kasper, S.H.; Morell-Perez, C.; Wyche, T.P.; Sana, T.R.; Lieberman, L.A.; Hett, E.C. Colorectal cancer-associated anaerobic bacteria proliferate in tumor spheroids and alter the microenvironment. Sci. Rep. 2020, 10, 5321. [Google Scholar] [CrossRef] [Green Version]
- Leschner, S.; Westphal, K.; Dietrich, N.; Viegas, N.; Jablonska, J.; Lyszkiewicz, M.; Lienenklaus, S.; Falk, W.; Gekara, N.; Loessner, H. Tumor invasion of Salmonella enterica serovar Typhimurium is accompanied by strong hemorrhage promoted by TNF-α. PLoS ONE 2009, 4, e6692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westphal, K.; Leschner, S.; Jablonska, J.; Loessner, H.; Weiss, S. Containment of tumor-colonizing bacteria by host neutrophils. Cancer Res. 2008, 68, 2952–2960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masso-Welch, P.; Girald Berlingeri, S.; King-Lyons, N.D.; Mandell, L.; Hu, J.; Greene, C.J.; Federowicz, M.; Cao, P.; Connell, T.D.; Heakal, Y. LT-IIc, a bacterial Type II heat-labile enterotoxin, induces specific lethality in triple negative breast cancer cells by modulation of autophagy and induction of apoptosis and necroptosis. Int. J. Mol. Sci. 2019, 20, 85. [Google Scholar] [CrossRef] [Green Version]
- Nes, I.F.; Holo, H. Class II antimicrobial peptides from lactic acid bacteria. Biopolymers 2000, 55, 50–61. [Google Scholar] [CrossRef]
- Weerakkody, L.R.; Witharana, C. The role of bacterial toxins and spores in cancer therapy. Life Sci. 2019, 235, 116839. [Google Scholar] [CrossRef]
- Nougayrède, J.-P.; Taieb, F.; De Rycke, J.; Oswald, E. Cyclomodulins: Bacterial effectors that modulate the eukaryotic cell cycle. Trends Microbiol. 2005, 13, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Pahle, J.; Menzel, L.; Niesler, N.; Kobelt, D.; Aumann, J.; Rivera, M.; Walther, W. Rapid eradication of colon carcinoma by Clostridium perfringens Enterotoxin suicidal gene therapy. BMC Cancer 2017, 17, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, T.; Mori, S.; Kishi, S.; Fujiwara-Tani, R.; Ohmori, H.; Nishiguchi, Y.; Hojo, Y.; Kawahara, I.; Nakashima, C.; Fujii, K. Effect of proton pump inhibitors on colorectal cancer. Int. J. Mol. Sci. 2020, 21, 3877. [Google Scholar] [CrossRef] [PubMed]
- Drider, D.; Bendali, F.; Naghmouchi, K.; Chikindas, M.L. Bacteriocins: Not only antibacterial agents. Probiotics Antimicrob. Proteins 2016, 8, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Martarelli, D.; Pompei, P.; Mazzoni, G. Inhibition of adrenocortical carcinoma by diphtheria toxin mutant CRM197. Chemotherapy 2009, 55, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Vallera, D.A.; Li, C.; Jin, N.; Panoskaltsis-Mortari, A.; Hall, W.A. Targeting urokinase-type plasminogen activator receptor on human glioblastoma tumors with diphtheria toxin fusion protein DTAT. J. Natl. Cancer Inst. 2002, 94, 597–606. [Google Scholar] [CrossRef] [Green Version]
- Yaghoubi, A.; Khazaei, M.; Avan, A.; Hasanian, S.M.; Soleimanpour, S. The bacterial instrument as a promising therapy for colon cancer. Int. J. Colorectal Dis. 2020, 35, 595–606. [Google Scholar] [CrossRef]
- Lutz, M.B.; Baur, A.S.; Schuler-Thurner, B.; Schuler, G. Immunogenic and tolerogenic effects of the chimeric IL-2-diphtheria toxin cytocidal agent Ontak(®) on CD25(+) cells. Oncoimmunology 2014, 3, e28223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadi, S.; Ghollasi, M.; Hosseini, H.M. The apoptotic impact of nisin as a potent bacteriocin on the colon cancer cells. Microb. Pathog. 2017, 111, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Norouzi, Z.; Salimi, A.; Halabian, R.; Fahimi, H. Nisin, a potent bacteriocin and anti-bacterial peptide, attenuates expression of metastatic genes in colorectal cancer cell lines. Microb. Pathog. 2018, 123, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Lewies, A.; Du Plessis, L.H.; Wentzel, J.F. The Cytotoxic, Antimicrobial and Anticancer Properties of the Antimicrobial Peptide Nisin Z Alone and in Combination with Conventional Treatments; Cytotoxicity; IntechOpen Limited: London, UK, 2018; pp. 21–42. [Google Scholar]
- Kohoutova, D.; Forstlova, M.; Moravkova, P.; Cyrany, J.; Bosak, J.; Smajs, D.; Rejchrt, S.; Bures, J. Bacteriocin production by mucosal bacteria in current and previous colorectal neoplasia. BMC Cancer 2020, 20, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chumchalova, J.; Šmarda, J. Human tumor cells are selectively inhibited by colicins. Folia Microbiol. 2003, 48, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, L.E.; Wintermeyer, W.; Rodnina, M.V. Colicins and their potential in cancer treatment. Blood Cells Mol. Dis. 2007, 38, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C.; Bono, M.R.; Barros, L.F.; Lagos, R. Microcin E492, a channel-forming bacteriocin from Klebsiella pneumoniae, induces apoptosis in some human cell lines. Proc. Natl. Acad. Sci. USA 2002, 99, 2696–2701. [Google Scholar] [CrossRef] [Green Version]
- Lagos, R.; Tello, M.; Mercado, G.; García, V.; Monasterio, O. Antibacterial and antitumorigenic properties of microcin E492, a pore-forming bacteriocin. Curr. Pharm. Biotechnol. 2009, 10, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Baindara, P.; Korpole, S.; Grover, V. Bacteriocins: Perspective for the development of novel anticancer drugs. Appl. Microbiol. Biotechnol. 2018, 102, 10393–10408. [Google Scholar] [CrossRef] [PubMed]
- Villarante, K.I.; Elegado, F.B.; Iwatani, S.; Zendo, T.; Sonomoto, K.; de Guzman, E.E. Purification, characterization and in vitro cytotoxicity of the bacteriocin from Pediococcus acidilactici K2a2-3 against human colon adenocarcinoma (HT29) and human cervical carcinoma (HeLa) cells. World J. Microbiol. Biotechnol. 2011, 27, 975–980. [Google Scholar] [CrossRef]
- Buss, G.P.; Wilson, C.M. Exploring the cytotoxic mechanisms of Pediocin PA-1 towards HeLa and HT29 cells by comparison to known bacteriocins: Microcin E492, enterocin heterodimer and Divercin V41. PLoS ONE 2021, 16, e0251951. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, D.; Kumar, A.; Mukhopadhyay, B.; Sengupta, T.K. Isolation of phenazine 1,6-di-carboxylic acid from Pseudomonas aeruginosa strain HRW.1-S3 and its role in biofilm-mediated crude oil degradation and cytotoxicity against bacterial and cancer cells. Appl. Microbiol. Biotechnol. 2015, 99, 8653–8665. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, R.K.; Naik, P.R.; Veena, V.; Lakshmi, B.S.; Lakshmi, P.; Krishna, R.; Sakthivel, N. 5-Methyl phenazine-1-carboxylic acid: A novel bioactive metabolite by a rhizosphere soil bacterium that exhibits potent antimicrobial and anticancer activities. Chem.-Biol. Interact. 2015, 231, 71–82. [Google Scholar] [CrossRef]
- Warso, M.A.; Richards, J.M.; Mehta, D.; Christov, K.; Schaeffer, C.; Rae Bressler, L.; Yamada, T.; Majumdar, D.; Kennedy, S.A.; Beattie, C.W.; et al. A first-in-class, first-in-human, phase I trial of p28, a non-HDM2-mediated peptide inhibitor of p53 ubiquitination in patients with advanced solid tumours. Br. J. Cancer 2013, 108, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Lulla, R.R.; Goldman, S.; Yamada, T.; Beattie, C.W.; Bressler, L.; Pacini, M.; Pollack, I.F.; Fisher, P.G.; Packer, R.J.; Dunkel, I.J.; et al. Phase I trial of p28 (NSC745104), a non-HDM2-mediated peptide inhibitor of p53 ubiquitination in pediatric patients with recurrent or progressive central nervous system tumors: A Pediatric Brain Tumor Consortium Study. Neuro-Oncology 2016, 18, 1319–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, M.; Zhou, J.; Su, Z.; Huang, Y. Bacterial cupredoxin azurin hijacks cellular signaling networks: Protein-protein interactions and cancer therapy. Protein Sci. 2017, 26, 2334–2341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garizo, A.R.; Coelho, L.F.; Pinto, S.; Dias, T.P.; Fernandes, F.; Bernardes, N.; Fialho, A.M. The Azurin-Derived Peptide CT-p19LC Exhibits Membrane-Active Properties and Induces Cancer Cell Death. Biomedicines 2021, 9, 1194. [Google Scholar] [CrossRef] [PubMed]
- Wolf, P.; Elsässer-Beile, U. Pseudomonas exotoxin A: From virulence factor to anti-cancer agent. Int. J. Med. Microbiol. 2009, 299, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Iglewski, B.H.; Kabat, D. NAD-dependent inhibition of protein synthesis by Pseudomonas aeruginosa toxin. Proc. Natl. Acad. Sci. USA 1975, 72, 2284–2288. [Google Scholar] [CrossRef] [Green Version]
- Shinohara, H.; Fan, D.; Ozawa, S.; Yano, S.; Van Arsdell, M.; Viner, J.L.; Beers, R.; Pastan, I.; Fidler, I.J. Site-specific expression of transferrin receptor by human colon cancer cells directly correlates with eradication by antitransferrin recombinant immunotoxin. Int. J. Oncol. 2000, 17, 643–651. [Google Scholar] [CrossRef]
- Shapira, S.; Shapira, A.; Starr, A.; Kazanov, D.; Kraus, S.; Benhar, I.; Arber, N. An immunoconjugate of anti-CD24 and Pseudomonas exotoxin selectively kills human colorectal tumors in mice. Gastroenterology 2011, 140, 935–946. [Google Scholar] [CrossRef]
- Gedde, M.M.; Higgins, D.E.; Tilney, L.G.; Portnoy, D.A. Role of listeriolysin O in cell-to-cell spread of Listeria monocytogenes. Infect. Immun. 2000, 68, 999–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunn, G.R.; Zubair, A.; Peters, C.; Pan, Z.-K.; Wu, T.-C.; Paterson, Y. Two Listeria monocytogenes vaccine vectors that express different molecular forms of human papilloma virus-16 (HPV-16) E7 induce qualitatively different T cell immunity that correlates with their ability to induce regression of established tumors immortalized by HPV-16. J. Immunol. 2001, 167, 6471–6479. [Google Scholar]
- Shahabi, V.; Seavey, M.; Maciag, P.; Rivera, S.; Wallecha, A. Development of a live and highly attenuated Listeria monocytogenes-based vaccine for the treatment of Her2/neu-overexpressing cancers in human. Cancer Gene Ther. 2011, 18, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara-Tani, R.; Sasaki, T.; Luo, Y.; Goto, K.; Kawahara, I.; Nishiguchi, Y.; Kishi, S.; Mori, S.; Ohmori, H.; Kondoh, M.; et al. Anti-claudin-4 extracellular domain antibody enhances the antitumoral effects of chemotherapeutic and antibody drugs in colorectal cancer. Oncotarget 2018, 9, 37367–37378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varas, M.A.; Muñoz-Montecinos, C.; Kallens, V.; Simon, V.; Allende, M.L.; Marcoleta, A.E.; Lagos, R. Exploiting Zebrafish Xenografts for Testing the in vivo Antitumorigenic Activity of Microcin E492 Against Human Colorectal Cancer Cells. Front. Microbiol. 2020, 11, 405. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.; Arya, R.; Goyal, K.; Singh, M.; Sharma, A.K. Bio-preservative and therapeutic potential of pediocin: Recent trends and future perspectives. Recent Pat. Biotechnol. 2013, 7, 172–178. [Google Scholar] [CrossRef]
- Al-Hazmi, N.E.; Naguib, D.M. Microbial Azurin Immobilized on Nano-Chitosan as Anticancer and Antibacterial Agent Against Gastrointestinal Cancers and Related Bacteria. J. Gastrointest. Cancer 2021. [Google Scholar] [CrossRef]
- Céspedes, M.V.; Cano-Garrido, O.; Álamo, P.; Sala, R.; Gallardo, A.; Serna, N.; Falgàs, A.; Voltà-Durán, E.; Casanova, I.; Sánchez-Chardi, A.; et al. Engineering Secretory Amyloids for Remote and Highly Selective Destruction of Metastatic Foci. Adv. Mater. 2020, 32, e1907348. [Google Scholar] [CrossRef] [PubMed]
- Mounier, J.; Ryter, A.; Coquis-Rondon, M.; Sansonetti, P.J. Intracellular and cell-to-cell spread of Listeria monocytogenes involves interaction with F-actin in the enterocytelike cell line Caco-2. Infect. Immun. 1990, 58, 1048–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.Y.; Williams, P.G.; Kwon, H.C.; Jensen, P.R.; Fenical, W. Lucentamycins A − D, cytotoxic peptides from the marine-derived actinomycete Nocardiopsis lucentensis. J. Nat. Prod. 2007, 70, 1321–1328. [Google Scholar] [CrossRef] [PubMed]
- Asolkar, R.N.; Freel, K.C.; Jensen, P.R.; Fenical, W.; Kondratyuk, T.P.; Park, E.-J.; Pezzuto, J.M. Arenamides A− C, Cytotoxic NFκB Inhibitors from the Marine Actinomycete Salinispora arenicola. J. Nat. Prod. 2010, 73, 796. [Google Scholar] [CrossRef]
- Um, S.; Choi, T.J.; Kim, H.; Kim, B.Y.; Kim, S.H.; Lee, S.K.; Oh, K.B.; Shin, J.; Oh, D.C. Ohmyungsamycins A and B: Cytotoxic and antimicrobial cyclic peptides produced by Streptomyces sp. from a volcanic island. J. Org. Chem. 2013, 78, 12321–12329. [Google Scholar] [CrossRef] [PubMed]
- Byun, W.S.; Kim, S.; Shin, Y.-H.; Kim, W.; Oh, D.-C.; Lee, S. Antitumor Activity of Ohmyungsamycin A through the Regulation of the Skp2-p27 Axis and MCM4 in Human Colorectal Cancer Cells. J. Nat. Prod. 2020, 83, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Hua, H.M.; Pei, Y.H.; Yao, X.S. Three new cytotoxic cyclic acylpeptides from marine Bacillus sp. Chem. Pharm. Bull. 2004, 52, 1029–1030. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, Y.; Kanoh, K.; Yamori, T.; Kasai, H.; Katsuta, A.; Adachi, K.; Shin-Ya, K.; Shizuri, Y. Urukthapelstatin A, a novel cytotoxic substance from marine-derived Mechercharimyces asporophorigenens YM11-542. J. Antibiot. 2007, 60, 251–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanoh, K.; Matsuo, Y.; Adachi, K.; Imagawa, H.; Nishizawa, M.; Shizuri, Y. Mechercharmycins A and B, cytotoxic substances from marine-derived Thermoactinomyces sp. YM3-251. J. Antibiot. 2005, 58, 289–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Du, Y.E.; Ban, Y.H.; Shin, Y.H.; Oh, D.C.; Yoon, Y.J. Enhanced Ohmyungsamycin A Production via Adenylation Domain Engineering and Optimization of Culture Conditions. Front. Microbiol. 2021, 12, 626881. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, S.; Fujimori, M.; Sasaki, T.; Tsutsui, H.; Shimatani, Y.; Seki, K.; Amano, J. Targeting solid tumors with non-pathogenic obligate anaerobic bacteria. Cancer Sci. 2010, 101, 1925–1932. [Google Scholar] [CrossRef]
- Dang, L.H.; Bettegowda, C.; Huso, D.L.; Kinzler, K.W.; Vogelstein, B. Combination bacteriolytic therapy for the treatment of experimental tumors. Proc. Natl. Acad. Sci. USA 2001, 98, 15155–15160. [Google Scholar] [CrossRef] [Green Version]
- Mody, K.; Baldeo, C.; Bekaii-Saab, T. Antiangiogenic Therapy in Colorectal Cancer. Cancer J. 2018, 24, 165–170. [Google Scholar] [CrossRef]
- Jia, L.J.; Xu, H.M.; Ma, D.Y.; Hu, Q.G.; Huang, X.F.; Jiang, W.H.; Li, S.F.; Jia, K.Z.; Huang, Q.L.; Hua, Z.C. Enhanced therapeutic effect by combination of tumor-targeting Salmonella and endostatin in murine melanoma model. Cancer Biol. Ther. 2005, 4, 840–845. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Fu, G.F.; Fan, Y.R.; Liu, W.H.; Liu, X.J.; Wang, J.J.; Xu, G.X. Bifidobacterium adolescentis as a delivery system of endostatin for cancer gene therapy: Selective inhibitor of angiogenesis and hypoxic tumor growth. Cancer Gene 2003, 10, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.F.; Zhu, L.P.; Hu, B.; Fu, G.F.; Zhang, H.Y.; Wang, J.J.; Xu, G.X. A new expression plasmid in Bifidobacterium longum as a delivery system of endostatin for cancer gene therapy. Cancer Gene 2007, 14, 151–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niethammer, A.G.; Xiang, R.; Becker, J.C.; Wodrich, H.; Pertl, U.; Karsten, G.; Eliceiri, B.P.; Reisfeld, R.A. A DNA vaccine against VEGF receptor 2 prevents effective angiogenesis and inhibits tumor growth. Nat. Med. 2002, 8, 1369–1375. [Google Scholar] [CrossRef] [PubMed]
- Kinnari, T.J. The role of biofilm in chronic laryngitis and in head and neck cancer. Curr. Opin. Otolaryngol. Head Neck Surg. 2015, 23, 448–453. [Google Scholar] [CrossRef]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef]
- Komor, U.; Bielecki, P.; Loessner, H.; Rohde, M.; Wolf, K.; Westphal, K.; Weiss, S.; Häussler, S. Biofilm formation by Pseudomonas aeruginosa in solid murine tumors–a novel model system. Microbes Infect. 2012, 14, 951–958. [Google Scholar] [CrossRef]
- Venkatesan, N.; Perumal, G.; Doble, M. Bacterial resistance in biofilm-associated bacteria. Future Microbiol. 2015, 10, 1743–1750. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Dejea, C.M.; Edler, D.; Hoang, L.T.; Santidrian, A.F.; Felding, B.H.; Ivanisevic, J.; Cho, K.; Wick, E.C.; Hechenbleikner, E.M. Metabolism links bacterial biofilms and colon carcinogenesis. Cell Metab. 2015, 21, 891–897. [Google Scholar] [CrossRef] [Green Version]
- Weitao, T. Bacteria form biofilms against cancer metastasis. Med. Hypotheses 2008, 72, 477–478. [Google Scholar] [CrossRef]
- Miyake, K.; Yamamoto, S.; Iijima, S. Blocking adhesion of cancer cells to endothelial cell types by S. agalactiae type-specific polysaccharides. Cytotechnology 1996, 22, 205–209. [Google Scholar] [CrossRef]
- Cheng, Y.; Ling, Z.; Li, L. The Intestinal Microbiota and Colorectal Cancer. Front. Immunol. 2020, 11, 615056. [Google Scholar] [CrossRef]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Gagnière, J.; Raisch, J.; Veziant, J.; Barnich, N.; Bonnet, R.; Buc, E.; Bringer, M.A.; Pezet, D.; Bonnet, M. Gut microbiota imbalance and colorectal cancer. World J. Gastroenterol. 2016, 22, 501–518. [Google Scholar] [CrossRef]
- Okumura, S.; Konishi, Y.; Narukawa, M.; Sugiura, Y.; Yoshimoto, S.; Arai, Y.; Sato, S.; Yoshida, Y.; Tsuji, S.; Uemura, K.; et al. Gut bacteria identified in colorectal cancer patients promote tumourigenesis via butyrate secretion. Nat. Commun. 2021, 12, 5674. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Gao, Z.; Huang, L.; Qin, H. Gut microbiota and colorectal cancer. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 757–769. [Google Scholar] [CrossRef]
- Lucas, C.; Barnich, N.; Nguyen, H.T.T. Microbiota, Inflammation and Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 1310. [Google Scholar] [CrossRef] [Green Version]
- Rubinstein, M.R.; Baik, J.E.; Lagana, S.M.; Han, R.P.; Raab, W.J.; Sahoo, D.; Dalerba, P.; Wang, T.C.; Han, Y.W. Fusobacterium nucleatum promotes colorectal cancer by inducing Wnt/β-catenin modulator Annexin A1. EMBO Rep. 2019, 20, e47638. [Google Scholar] [CrossRef] [PubMed]
- Walia, S.; Kamal, R.; Dhawan, D.; Kanwar, S. Chemoprevention by probiotics during 1,2-dimethylhydrazine-induced colon carcinogenesis in rats. Dig. Dis. Sci. 2018, 63, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Jenab, A.; Roghanian, R.; Emtiazi, G. Bacterial Natural Compounds with Anti-Inflammatory and Immunomodulatory Properties (Mini Review). Drug Des. Dev. Ther. 2020, 14, 3787–3801. [Google Scholar] [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.-L. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti–PD-L1 efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakatani, A.; Fujiya, M.; Ueno, N.; Kashima, S.; Sasajima, J.; Moriichi, K.; Ikuta, K.; Tanabe, H.; Kohgo, Y. Polyphosphate derived from Lactobacillus brevis inhibits colon cancer progression through induction of cell apoptosis. Anticancer Res. 2016, 36, 591–598. [Google Scholar] [PubMed]
- Jacouton, E.; Chain, F.; Sokol, H.; Langella, P.; Bermudez-Humaran, L.G. Probiotic strain Lactobacillus casei BL23 prevents colitis-associated colorectal cancer. Front. Immunol. 2017, 8, 1553. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.S.; Wynder, E.L. Metabolic epidemiology of colon cancer: Fecal bile acids and neutral sterols in colon cancer patients and patients with adenomatous polyps. Cancer 1977, 39, 2533–2539. [Google Scholar] [CrossRef]
- Choi, S.S.; Kang, B.Y.; Chung, M.J.; Kim, S.D.; Park, S.H.; Kim, J.S.; Kang, C.Y.; Ha, N.J. Safety assessment of potential lactic acid bacteria Bifidobacterium longum SPM1205 isolated from healthy Koreans. J. Microbiol. 2005, 43, 493–498. [Google Scholar] [PubMed]
- Lee, D.K.; Jang, S.; Kim, M.J.; Kim, J.H.; Chung, M.J.; Kim, K.J.; Ha, N.J. Anti-proliferative effects of Bifidobacterium adolescentis SPM0212 extract on human colon cancer cell lines. BMC Cancer 2008, 8, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.; Van Treuren, W.; Fischer, C.R.; Merrill, B.D.; DeFelice, B.C.; Sanchez, J.M.; Higginbottom, S.K.; Guthrie, L.; Fall, L.A.; Dodd, D. A metabolomics pipeline for the mechanistic interrogation of the gut microbiome. Nature 2021, 595, 415–420. [Google Scholar]
- Foti, R.S.; Tyndale, R.F.; Garcia, K.L.; Sweet, D.H.; Nagar, S.; Sharan, S.; Rock, D.A. “Target-Site” drug metabolism and transport. Drug Metab. Dispos. 2015, 43, 1156–1168. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.R.; Mazmanian, S.K. Control of brain development, function, and behavior by the microbiome. Cell Host Microbe 2015, 17, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Sivashanmugam, M.; Jaidev, J.; Umashankar, V.; Sulochana, K.N. Ornithine and its role in metabolic diseases: An appraisal. Biomed. Pharmacother. 2017, 86, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Maier, L.; Pruteanu, M.; Kuhn, M.; Zeller, G.; Telzerow, A.; Anderson, E.E.; Brochado, A.R.; Fernandez, K.C.; Dose, H.; Mori, H. Extensive impact of non-antibiotic drugs on human gut bacteria. Nature 2018, 555, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.; Zimmermann-Kogadeeva, M.; Wegmann, R.; Goodman, A.L. Separating host and microbiome contributions to drug pharmacokinetics and toxicity. Science 2019, 363, eaat9931. [Google Scholar] [CrossRef]
- Kong, J.; Lee, H.; Kim, D.; Han, S.K.; Ha, D.; Shin, K.; Kim, S. Network-based machine learning in colorectal and bladder organoid models predicts anti-cancer drug efficacy in patients. Nat. Commun. 2020, 11, 5485. [Google Scholar] [CrossRef] [PubMed]
- Mitsala, A.; Tsalikidis, C.; Pitiakoudis, M.; Simopoulos, C.; Tsaroucha, A.K. Artificial Intelligence in Colorectal Cancer Screening, Diagnosis and Treatment. A New Era. Curr. Oncol. 2021, 28, 1581–1607. [Google Scholar] [CrossRef] [PubMed]
- Sousa, T.; Yadav, V.; Zann, V.; Borde, A.; Abrahamsson, B.; Basit, A.W. On the colonic bacterial metabolism of azo-bonded prodrugsof 5-aminosalicylic acid. J. Pharm. Sci. 2014, 103, 3171–3175. [Google Scholar] [CrossRef]
- Haiser, H.J.; Gootenberg, D.B.; Chatman, K.; Sirasani, G.; Balskus, E.P.; Turnbaugh, P.J. Predicting and manipulating cardiac drug inactivation by the human gut bacterium Eggerthella lenta. Science 2013, 341, 295–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iida, N.; Dzutsev, A.; Stewart, C.A.; Smith, L.; Bouladoux, N.; Weingarten, R.A.; Molina, D.A.; Salcedo, R.; Back, T.; Cramer, S. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science 2013, 342, 967–970. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Jia, W. Cometabolism of microbes and host: Implications for drug metabolism and drug-induced toxicity. Clin. Pharmacol. Ther. 2013, 94, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Jariwala, P.B.; Pellock, S.J.; Goldfarb, D.; Cloer, E.W.; Artola, M.; Simpson, J.B.; Bhatt, A.P.; Walton, W.G.; Roberts, L.R.; Major, M.B. Discovering the microbial enzymes driving drug toxicity with activity-based protein profiling. ACS Chem. Biol. 2019, 15, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Javdan, B.; Lopez, J.G.; Chankhamjon, P.; Lee, Y.-C.J.; Hull, R.; Wu, Q.; Wang, X.; Chatterjee, S.; Donia, M.S. Personalized mapping of drug metabolism by the human gut microbiome. Cell 2020, 181, 1661–1679.e22. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.; Zimmermann-Kogadeeva, M.; Wegmann, R.; Goodman, A.L. Mapping human microbiome drug metabolism by gut bacteria and their genes. Nature 2019, 570, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, L.; Wolfson, S.; Kelly, L. The human gut chemical landscape predicts microbe-mediated biotransformation of foods and drugs. Elife 2019, 8, e42866. [Google Scholar] [CrossRef] [PubMed]
- Van Leerdam, M.E.; Roos, V.H.; van Hooft, J.E.; Balaguer, F.; Dekker, E.; Kaminski, M.F.; Latchford, A.; Neumann, H.; Ricciardiello, L.; Rupińska, M. Endoscopic management of Lynch syndrome and of familial risk of colorectal cancer: European Society of Gastrointestinal Endoscopy (ESGE) Guideline. Endoscopy 2019, 51, 1082–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillère, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [Green Version]
- Brandi, G.; Dabard, J.; Raibaud, P.; Di Battista, M.; Bridonneau, C.; Pisi, A.M.; Labate, A.M.M.; Pantaleo, M.A.; De Vivo, A.; Biasco, G. Intestinal microflora and digestive toxicity of irinotecan in mice. Clin. Cancer Res. 2006, 12, 1299–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuckerman, D.S.; Clark, J.W. Systemic therapy for metastatic colorectal cancer: Current questions. Cancer 2008, 112, 1879–1891. [Google Scholar] [CrossRef] [PubMed]
- Wallace, B.D.; Roberts, A.B.; Pollet, R.M.; Ingle, J.D.; Biernat, K.A.; Pellock, S.J.; Venkatesh, M.K.; Guthrie, L.; O’Neal, S.K.; Robinson, S.J. Structure and inhibition of microbiome β-glucuronidases essential to the alleviation of cancer drug toxicity. Chem. Biol. 2015, 22, 1238–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taper, H.S.; Roberfroid, M.B. Possible adjuvant cancer therapy by two prebiotics-inulin or oligofructose. In Vivo 2005, 19, 201–204. [Google Scholar] [PubMed]
- Brockmueller, A.; Sameri, S.; Liskova, A.; Zhai, K.; Varghese, E.; Samuel, S.M.; Büsselberg, D.; Kubatka, P.; Shakibaei, M. Resveratrol’s Anti-Cancer Effects through the Modulation of Tumor Glucose Metabolism. Cancers 2021, 13, 188. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Shayan, P.; Brockmueller, A.; Shakibaei, M. Resveratrol Suppresses Cross-Talk between Colorectal Cancer Cells and Stromal Cells in Multicellular Tumor Microenvironment: A Bridge between In Vitro and In Vivo Tumor Microenvironment Study. Molecules 2020, 25, 4292. [Google Scholar] [CrossRef]
- Buhrmann, C.; Kraehe, P.; Lueders, C.; Shayan, P.; Goel, A.; Shakibaei, M. Curcumin suppresses crosstalk between colon cancer stem cells and stromal fibroblasts in the tumor microenvironment: Potential role of EMT. PLoS ONE 2014, 9, e107514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhrmann, C.; Brockmueller, A.; Harsha, C.; Kunnumakkara, A.B.; Kubatka, P.; Aggarwal, B.B.; Shakibaei, M. Evidence That Tumor Microenvironment Initiates Epithelial-To-Mesenchymal Transition and Calebin A can Suppress it in Colorectal Cancer Cells. Front. Pharm. 2021, 12, 699842. [Google Scholar] [CrossRef] [PubMed]
- Buhrmann, C.; Kunnumakkara, A.B.; Kumar, A.; Samec, M.; Kubatka, P.; Aggarwal, B.B.; Shakibaei, M. Multitargeting Effects of Calebin A on Malignancy of CRC Cells in Multicellular Tumor Microenvironment. Front. Oncol. 2021, 11, 650603. [Google Scholar] [CrossRef]
- Buhrmann, C.; Kunnumakkara, A.B.; Popper, B.; Majeed, M.; Aggarwal, B.B.; Shakibaei, M. Calebin A Potentiates the Effect of 5-FU and TNF-β (Lymphotoxin α) against Human Colorectal Cancer Cells: Potential Role of NF-κB. Int. J. Mol. Sci. 2020, 21, 2393. [Google Scholar] [CrossRef] [Green Version]
- Eiseman, B.; Silen, W.; Bascom, G.S.; Kauvar, A.J. Fecal enema as an adjunct in the treatment of pseudomembranous enterocolitis. Surgery 1958, 44, 854–859. [Google Scholar]
- Gupta, S.; Allen-Vercoe, E.; Petrof, E.O. Fecal microbiota transplantation: In perspective. Ther. Adv. Gastroenterol. 2016, 9, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiao, S.L.; Kershaw, K.M.; Limon, J.J.; You, S.; Yoon, J.; Ko, E.Y.; Guarnerio, J.; Potdar, A.A.; McGovern, D.P.; Bose, S. Commensal bacteria and fungi differentially regulate tumor responses to radiation therapy. Cancer Cell 2021, 39, 1202–1213.e6. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Xiao, H.; Li, Y.; Zhou, L.; Zhao, S.; Luo, D.; Zheng, Q.; Dong, J.; Zhao, Y.; Zhang, X. Faecal microbiota transplantation protects against radiation-induced toxicity. EMBO Mol. Med. 2017, 9, 448–461. [Google Scholar] [CrossRef] [PubMed]
- Ciorba, M.A.; Riehl, T.E.; Rao, M.S.; Moon, C.; Ee, X.; Nava, G.M.; Walker, M.R.; Marinshaw, J.M.; Stappenbeck, T.S.; Stenson, W.F. Lactobacillus probiotic protects intestinal epithelium from radiation injury in a TLR-2/cyclo-oxygenase-2-dependent manner. Gut 2012, 61, 829–838. [Google Scholar] [CrossRef]
- Liscano, Y.; Oñate-Garzón, J.; Delgado, J.P. Peptides with dual antimicrobial–anticancer activity: Strategies to overcome peptide limitations and rational design of anticancer peptides. Molecules 2020, 25, 4245. [Google Scholar] [CrossRef]
- Ludgate, C.M. Optimizing cancer treatments to induce an acute immune response: Radiation Abscopal effects, PAMPs, and DAMPs. Clin. Cancer Res. 2012, 18, 4522–4525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedl, S.; Zweytick, D.; Lohner, K. Membrane-active host defense peptides—Challenges and perspectives for the development of novel anticancer drugs. Chem. Phys. Lipids 2011, 164, 766–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tørfoss, V.; Isaksson, J.; Ausbacher, D.; Brandsdal, B.O.; Flaten, G.E.; Anderssen, T.; Cavalcanti-Jacobsen Cde, A.; Havelkova, M.; Nguyen, L.T.; Vogel, H.J.; et al. Improved anticancer potency by head-to-tail cyclization of short cationic anticancer peptides containing a lipophilic β(2,2)-amino acid. J. Pept. Sci. 2012, 18, 609–619. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mueller, A.-L.; Brockmueller, A.; Fahimi, N.; Ghotbi, T.; Hashemi, S.; Sadri, S.; Khorshidi, N.; Kunnumakkara, A.B.; Shakibaei, M. Bacteria-Mediated Modulatory Strategies for Colorectal Cancer Treatment. Biomedicines 2022, 10, 832. https://doi.org/10.3390/biomedicines10040832
Mueller A-L, Brockmueller A, Fahimi N, Ghotbi T, Hashemi S, Sadri S, Khorshidi N, Kunnumakkara AB, Shakibaei M. Bacteria-Mediated Modulatory Strategies for Colorectal Cancer Treatment. Biomedicines. 2022; 10(4):832. https://doi.org/10.3390/biomedicines10040832
Chicago/Turabian StyleMueller, Anna-Lena, Aranka Brockmueller, Niusha Fahimi, Tahere Ghotbi, Sara Hashemi, Sadaf Sadri, Negar Khorshidi, Ajaikumar B. Kunnumakkara, and Mehdi Shakibaei. 2022. "Bacteria-Mediated Modulatory Strategies for Colorectal Cancer Treatment" Biomedicines 10, no. 4: 832. https://doi.org/10.3390/biomedicines10040832