
Photodynamic Effects with 5-Aminolevulinic Acid on Cytokines and Exosomes in Human Peripheral Blood Mononuclear Cells

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Isolation and Culture of PBMCs
2.3. Incubation with ALA and ALA Induced PpIX Production in PBMCs
2.4. Light Source
2.5. PDT Treatment of PBMCs
2.6. Flow Cytometry Analysis
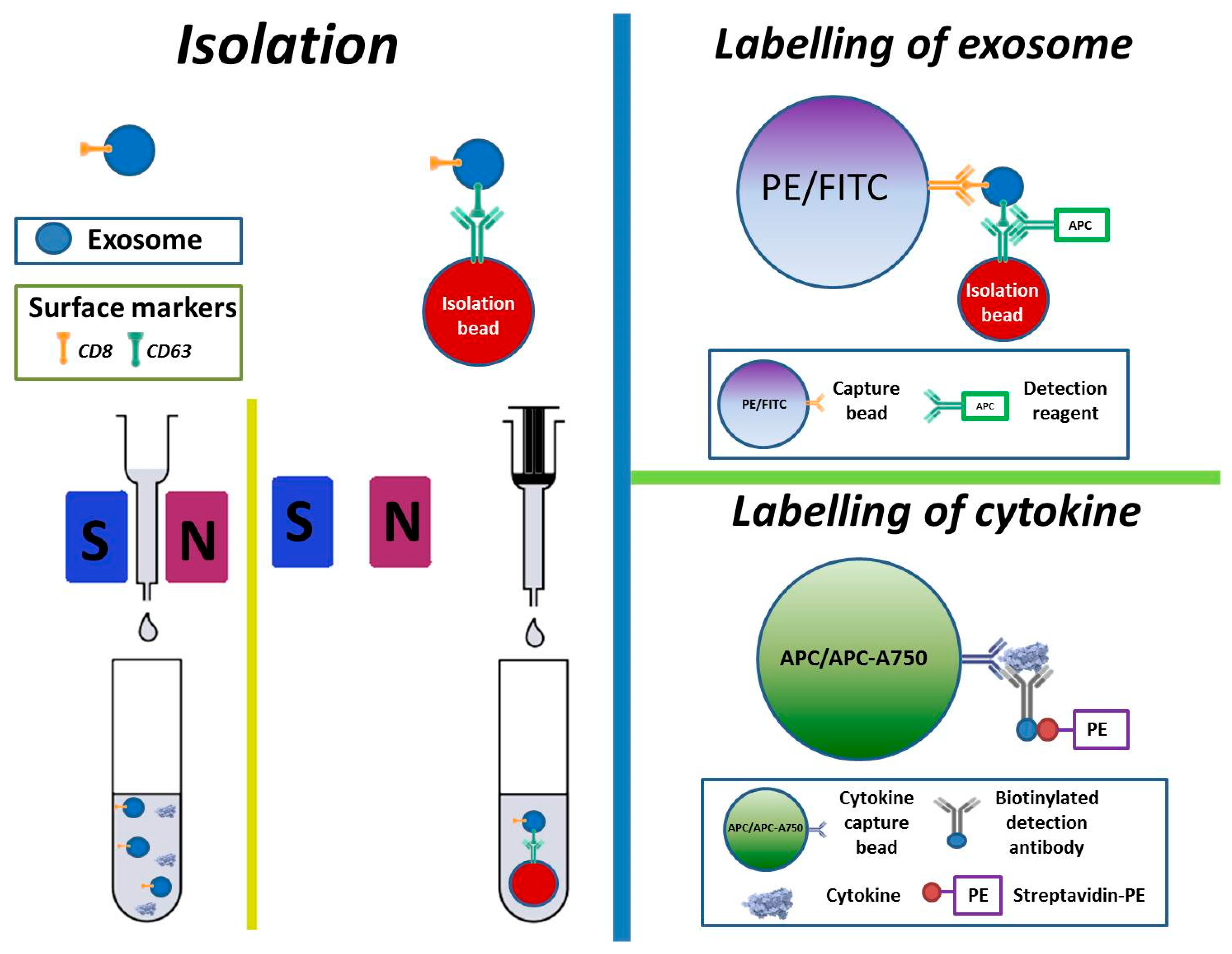
2.7. Isolation, Labelling, and Measurement of Cytokines and Exosomes
2.8. Electron Microscopy of Exosomes
2.9. Statistical Analyses
3. Results
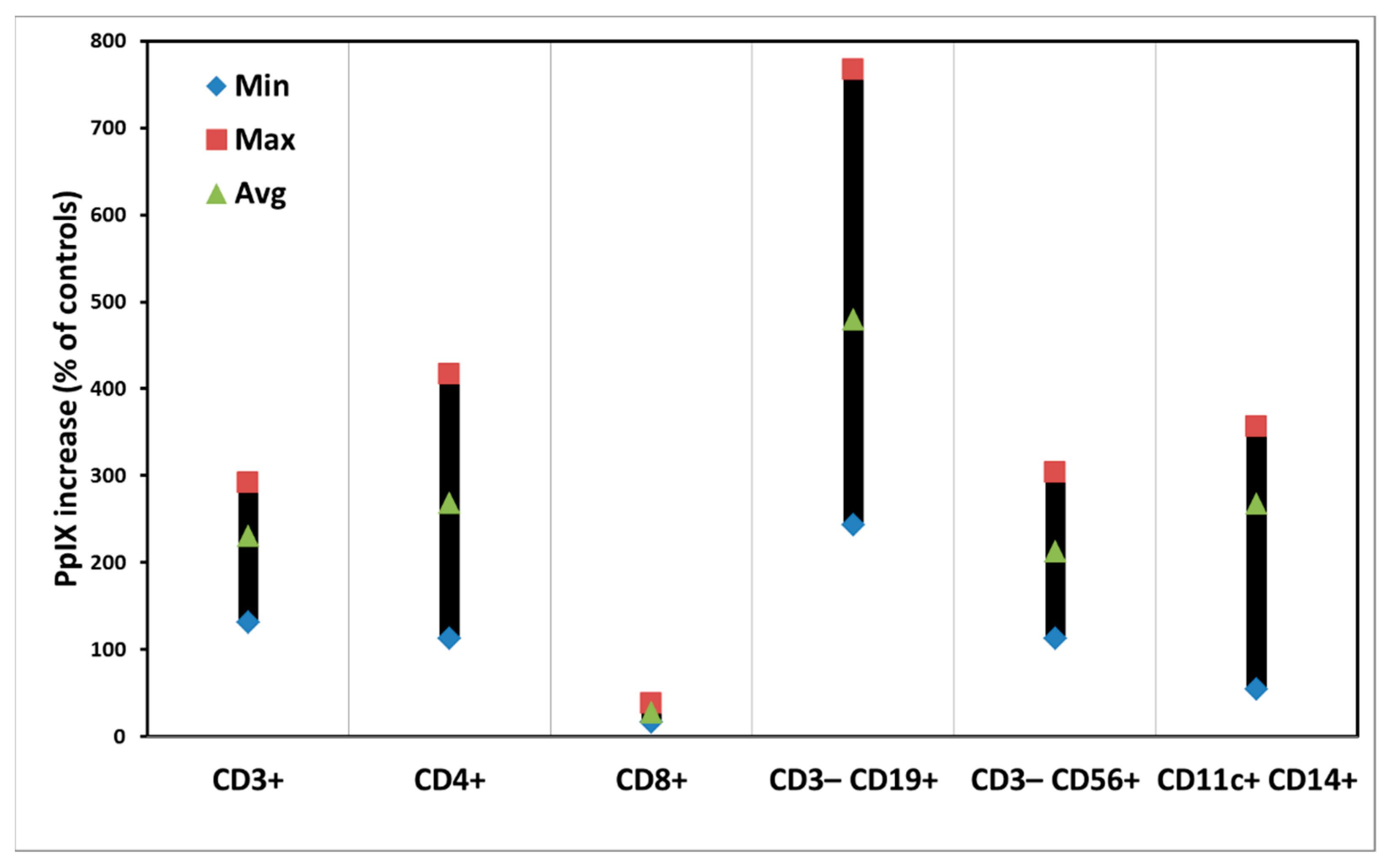
3.1. ALA-Induced PpIX Production in PBMCs
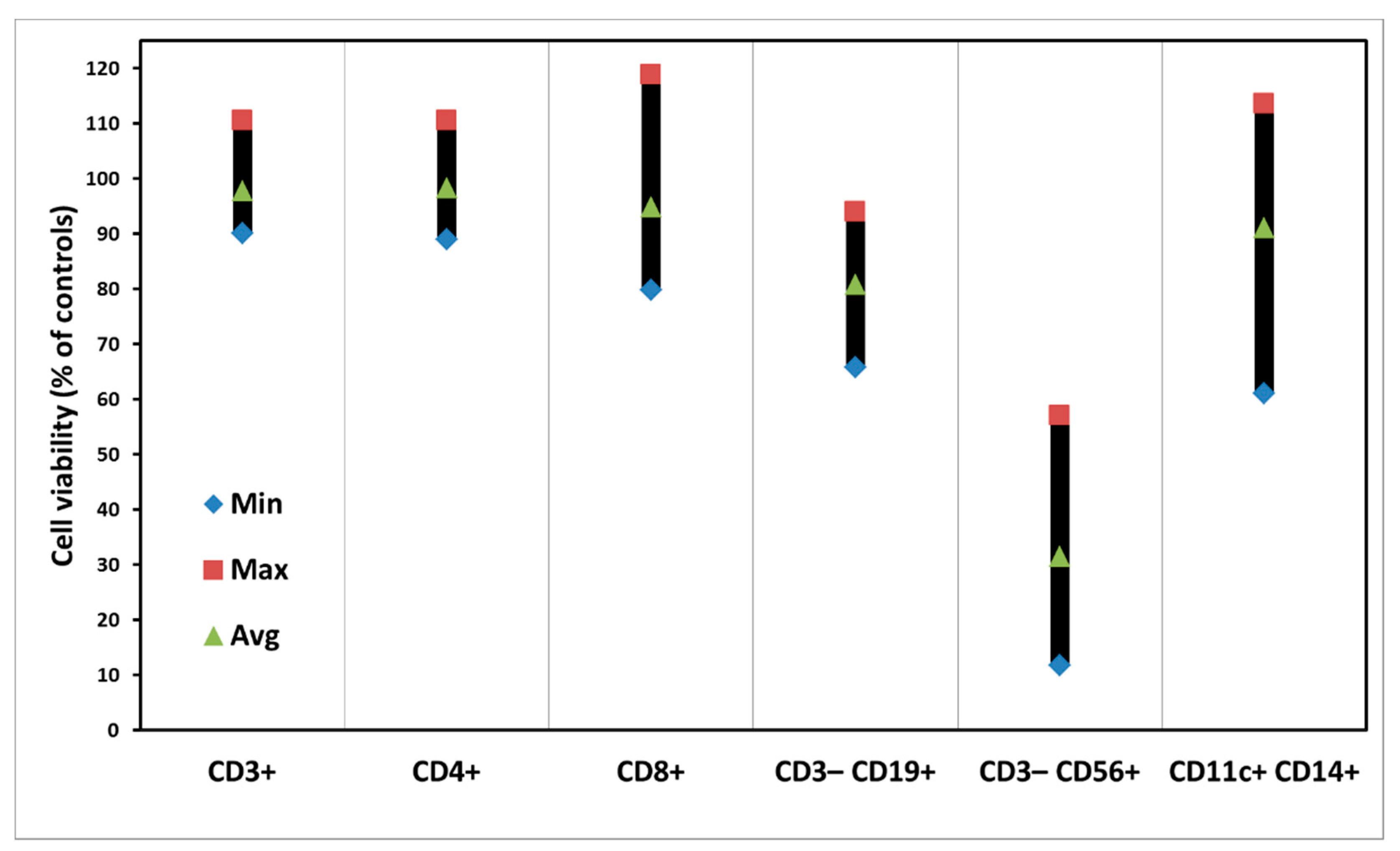
3.2. Dark Toxicity of PBMCs with ALA Alone
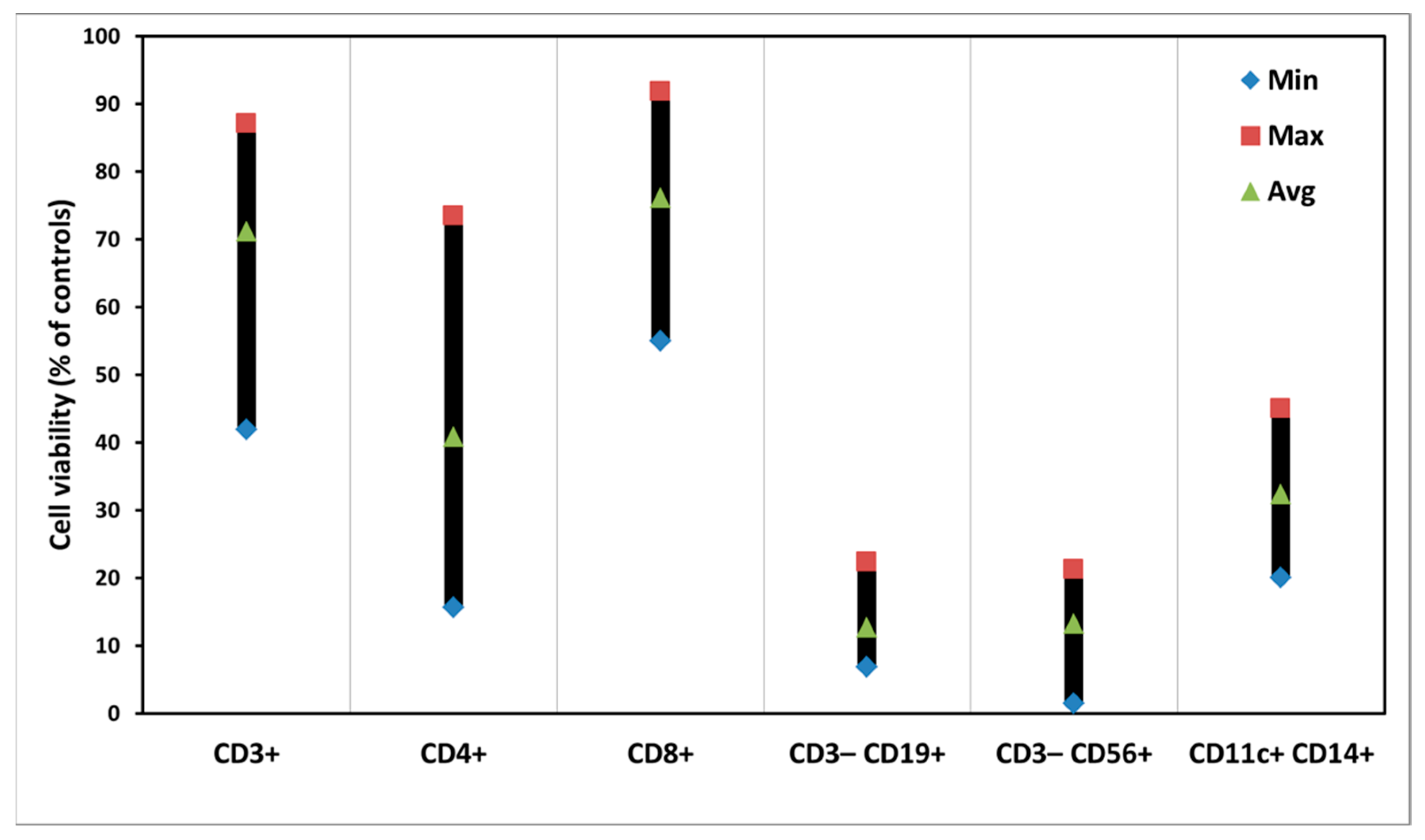
3.3. PDT of PBMCs with ALA
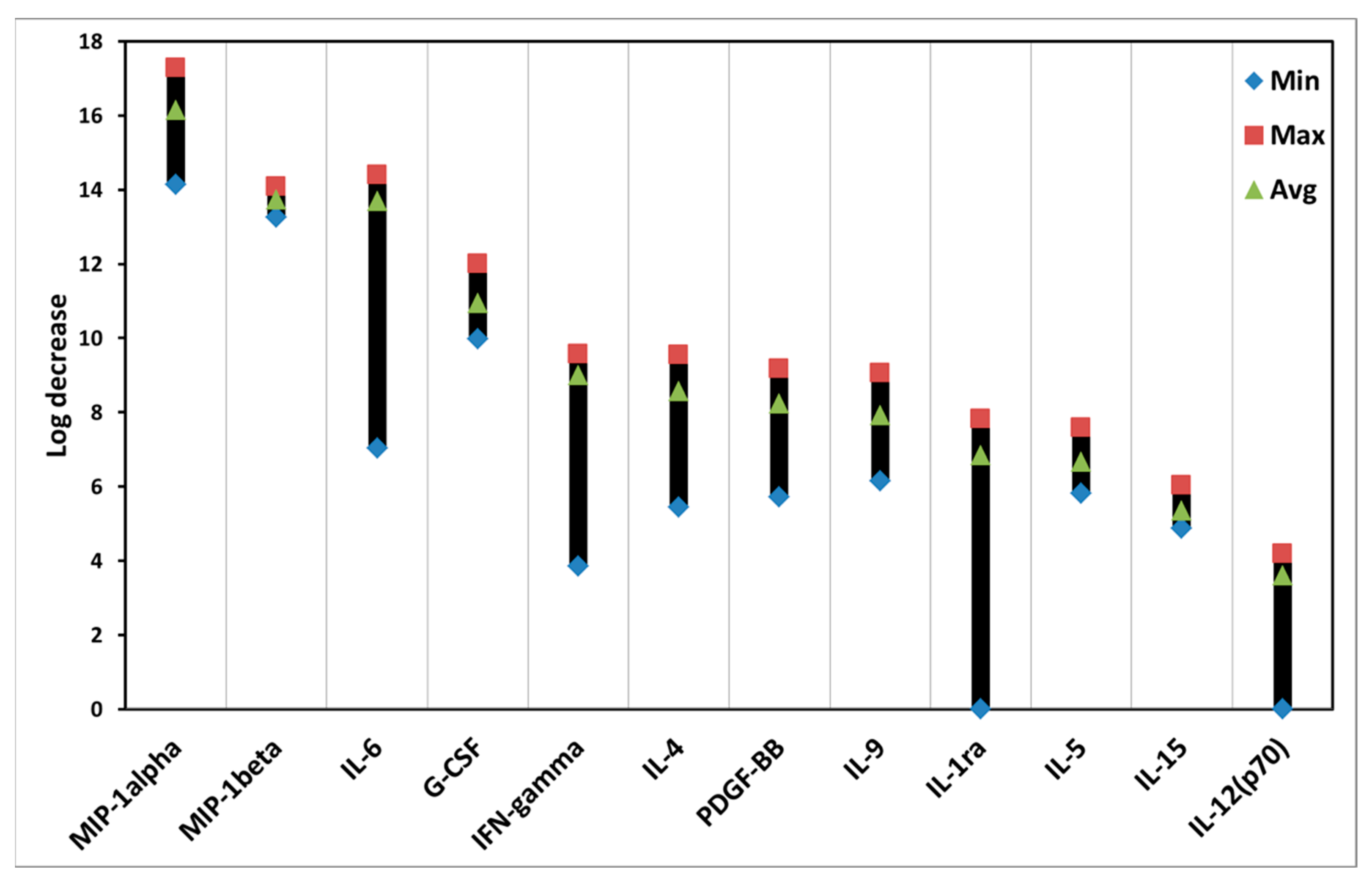
3.4. Effects of ALA-PDT on Cytokines of PBMCs
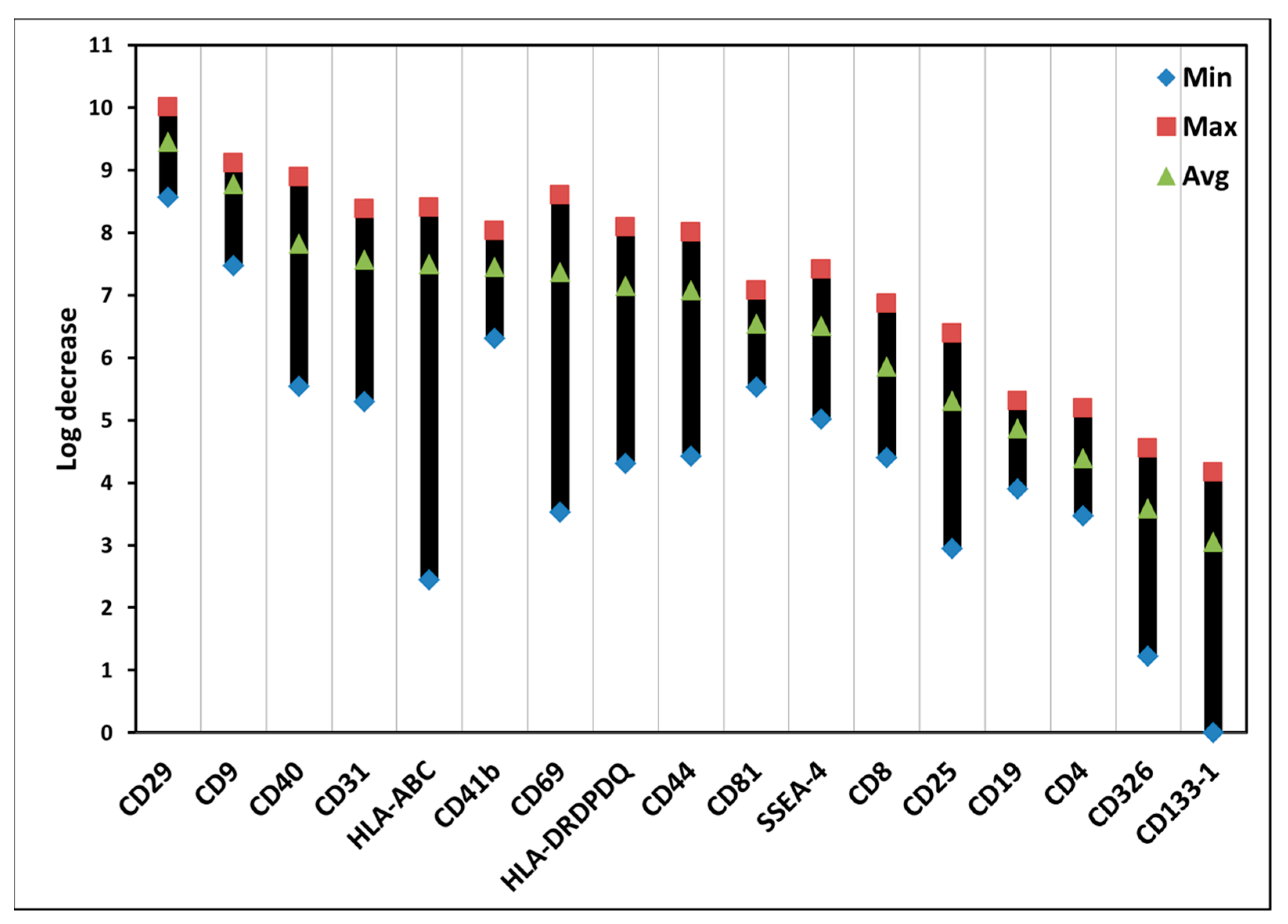
3.5. Effects of ALA-PDT on Exosomes of PBMCs
3.6. Effects of Light Alone on Subsets, Exosomes, and Cytokines of PBMCs
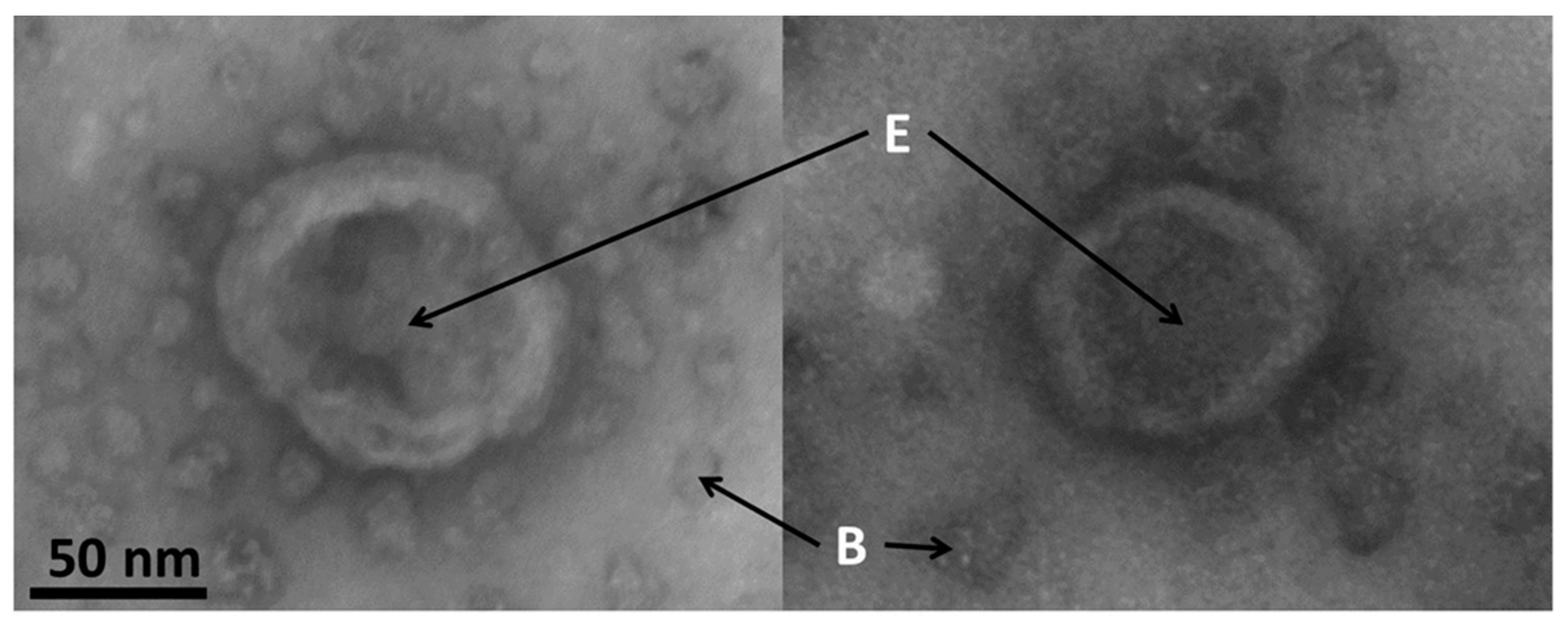
3.7. Electron Microscopy of Exosomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dougherty, T.J.; Gomer, C.J.; Henderson, B.W.; Jori, G.; Kessel, D.; Korbelik, M.; Moan, J.; Peng, Q. Photodynamic therapy. J. Nat. Cancer Inst. 1998, 90, 889–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Straten, D.; Mashayekhi, V.; de Bruijn, H.S.; Oliveira, S.; Robinson, D.J. Oncologic Photodynamic Therapy: Basic Principles, Current Clinical Status and Future Directions. Cancers 2017, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Warloe, T.; Berg, K.; Moan, J.; Kongshaug, M.; Giercksky, K.E.; Nesland, J.M. 5-Aminolevulinic acid-based photodynamic therapy. Clinical research and future challenges. Cancer 1997, 79, 2282–2308. [Google Scholar] [CrossRef]
- Peng, Q.; Berg, K.; Moan, J.; Kongshaug, M.; Nesland, J.M. 5-Aminolevulinic acid-based photodynamic therapy: Principles and experimental research. Photochem. Photobiol. 1997, 65, 235–251. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Juzeniene, A.; Chen, J.; Svaasand, L.O.; Warloe, T.; Giercksky, K.E.; Moan, J. Lasers in medicine. Rep. Prog. Phys. 2008, 71, 056701. [Google Scholar] [CrossRef]
- Stummer, W.; Pichlmeier, U.; Meinel, T.; Wiestler, O.D.; Zanella, F.; Reulen, H.J. Fluorescence-guided surgery with 5-aminolevulinic acid for resection of malignant glioma: A randomised controlled multicentre phase III trial. Lancet Oncol. 2006, 7, 392–401. [Google Scholar] [CrossRef]
- Mosquera-Heredia, M.I.; Morales, L.C.; Vidal, O.M.; Barceló, E.; Silvera-Redondo, C.; Vélez, J.I.; Garavito-Galofre, P. Exosomes: Potential Disease Biomarkers and New Therapeutic Targets. Biomedicines 2021, 9, 1061. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, 6977. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, W.; Zhang, H.; Zhang, F.; Chen, L.; Ma, L.; Larcher, L.M.; Chen, S.; Liu, N.; Zhao, Q.; et al. Progress, opportunity, and perspective on exosome isolation-efforts for efficient exosome-based theranostics. Theranostics 2020, 10, 3684–3707. [Google Scholar] [CrossRef]
- Mkhobongo, B.; Chandran, R.; Abrahamse, H. The Role of Melanoma Cell-Derived Exosomes (MTEX) and Photodynamic Therapy (PDT) within a Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 9726. [Google Scholar] [CrossRef]
- Jiang, Y.; Xu, C.; Leung, W.; Lin, M.; Cai, X.; Guo, H.; Zhang, J.; Yang, F. Role of Exosomes in Photodynamic Anticancer Therapy. Curr. Med. Chem. 2020, 27, 6815–6824. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhang, H.; Zeng, Q.; Wang, P.; Zhang, G.; Ji, J.; Li, M.; Shen, S.; Wang, X. Exosomes from 5-aminolevulinic acid photodynamic therapy-treated squamous carcinoma cells promote dendritic cell maturation. Photodiagn. Photodyn. Ther. 2020, 30, 101746. [Google Scholar] [CrossRef] [PubMed]
- Sando, Y.; Matsuoka, K.-I.; Sumii, Y.; Kondo, T.; Ikegawa, S.; Sugiura, H.; Nakamura, M.; Iwamoto, M.; Meguri, Y.; Asada, N.; et al. 5-aminolevulinic acid-mediated photodynamic therapy can target aggressive adult T cell leukemia/lymphoma resistant to conventional chemotherapy. Sci. Rep. 2020, 10, 17237. [Google Scholar] [CrossRef]
- Christensen, E.; Foss, O.A.; Quist-Paulsen, P.; Staur, I.; Pettersen, F.; Holien, T.; Juzenas, P.; Peng, Q. Application of Photodynamic Therapy with 5-Aminolevulinic Acid to Extracorporeal Photopheresis in the Treatment of Patients with Chronic Graft-versus-Host Disease: A First-in-Human Study. Pharmaceutics 2021, 13, 1558. [Google Scholar] [CrossRef] [PubMed]
- Darvekar, S.; Juzenas, P.; Oksvold, M.; Kleinauskas, A.; Holien, T.; Christensen, E.; Stokke, T.; Sioud, M.; Peng, Q. Selective Killing of Activated T Cells by 5-Aminolevulinic Acid Mediated Photodynamic Effect: Potential Improvement of Extracorporeal Photopheresis. Cancers 2020, 12, 377. [Google Scholar] [CrossRef] [Green Version]
- Casas, A. Clinical uses of 5-aminolaevulinic acid in photodynamic treatment and photodetection of cancer: A review. Cancer Lett. 2020, 490, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Furre, I.E.; Møller, M.T.; Shahzidi, S.; Nesland, J.M.; Peng, Q. Involvement of both caspase-dependent and -independent pathways in apoptotic induction by hexaminolevulinate-mediated photodynamic therapy in human lymphoma cells. Apoptosis 2006, 11, 2031–2042. [Google Scholar] [CrossRef] [PubMed]
- Furre, I.E.; Shahzidi, S.; Luksiene, Z.; Møller, M.T.N.; Borgen, E.; Morgan, J.; Tkacz-Stachowska, K.; Nesland, J.M.; Peng, Q. Targeting PBR by hexaminolevulinate-mediated photodynamic therapy induces apoptosis through translocation of apoptosis-inducing factor in human leukemia cells. Cancer Res. 2005, 65, 11051–11060. [Google Scholar] [CrossRef] [Green Version]
- Shahzidi, S.; Čunderlíková, B.; Więdłocha, A.; Zhen, Y.; Vasovič, V.; Nesland, J.M.; Peng, Q. Simultaneously targeting mitochondria and endoplasmic reticulum by photodynamic therapy induces apoptosis in human lymphoma cells. Photochem. Photobiol. Sci. 2011, 10, 1773–1782. [Google Scholar] [CrossRef]
- Holien, T.; Gederaas, O.A.; Darvekar, S.R.; Christensen, E.; Peng, Q. Comparison between 8-methoxypsoralen and 5-aminolevulinic acid in killing T cells of photopheresis patients ex vivo. Lasers Surg. Med. 2018, 50, 469–475. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Bi, J.; Huang, J.; Tang, Y.; Du, S.; Li, P. Exosome: A Review of Its Classification, Isolation Techniques, Storage, Diagnostic and Targeted Therapy Applications. Int. J. Nanomed. 2020, 15, 6917–6934. [Google Scholar] [CrossRef] [PubMed]
- Ragab, D.; Salah Eldin, H.; Taeimah, M.; Khattab, R.; Salem, R. The COVID-19 Cytokine Storm; What We Know so Far. Front. Immunol. 2020, 11, 1446. [Google Scholar] [CrossRef] [PubMed]
- Gorabi, A.M.; Hajighasemi, S.; Kiaie, N.; Gheibi-Hayat, S.M.; Jamialahmadi, T.; Johnston, T.P.; Sahebkar, A. The pivotal role of CD69 in autoimmunity. J. Autoimmun. 2020, 111, 102453. [Google Scholar] [CrossRef] [PubMed]
- Marino, J.; Paster, J.; Benichou, G. Allorecognition by T Lymphocytes and Allograft Rejection. Front. Immunol. 2016, 7, 582. [Google Scholar] [CrossRef] [Green Version]
- Timrott, K.; Beetz, O.; Oldhafer, F.; Klempnauer, J.; Vondran, F.W.R.; Jäger, M.D. The importance of MHC class II in allogeneic bone marrow transplantation and chimerism-based solid organ tolerance in a rat model. PLoS ONE 2020, 15, e0233497. [Google Scholar] [CrossRef]
- Khaddour, K.; Hana, C.K.; Mewawalla, P. Hematopoietic Stem Cell Transplantation; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Howell, W.M.; Carter, V.; Clark, B. The HLA system: Immunobiology, HLA typing, antibody screening and crossmatching techniques. J. Clin. Pathol. 2010, 63, 387. [Google Scholar] [CrossRef]
- Leeaphorn, N.; Pena, J.; Thamcharoen, N.; Khankin, E.; Pavlakis, M.; Cardarelli, F. HLA-DQ Mismatching and Kidney Transplant Outcomes. Clin. J. Am. Soc. Nephrol. 2018, 13, 763–771. [Google Scholar] [CrossRef] [Green Version]
- Barzegar Behrooz, A.; Syahir, A.; Ahmad, S. CD133: Beyond a cancer stem cell biomarker. J. Drug Target. 2019, 27, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Trusler, O.; Huang, Z.; Goodwin, J.; Laslett, A.L. Cell surface markers for the identification and study of human naive pluripotent stem cells. Stem Cell Res. 2018, 26, 36–43. [Google Scholar] [CrossRef]
- Castellani, C.; Burrello, J.; Fedrigo, M.; Burrello, A.; Bolis, S.; Di Silvestre, D.; Tona, F.; Bottio, T.; Biemmi, V.; Toscano, G.; et al. Circulating extracellular vesicles as non-invasive biomarker of rejection in heart transplant. J. Heart Lung Transplant. 2020, 39, 1136–1148. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokine | PDT Effect (Avg.) | Cell Type | Possible Biological Function |
|---|---|---|---|
| MIP-1alpha (macrophage inflammatory protein-1 alpha) | 16.1 | Macrophage | Pro-inflammation |
| MIP-1beta (macrophage inflammatory protein-1beta) | 13.7 | Macrophage | Pro-inflammation |
| IL-6 (interleukin-6) | 13.7 | Macrophage | Pro-inflammation |
| G-CSF (granulocyte colony stimulating factor) | 10.9 | Macrophage and other cells | Growth stimulation of white blood cells |
| IFN-gamma (interferon gamma) | 9 | T-cell and NK cell | Pro-inflammation and worsen autoimmune diseases |
| IL-4 (interleukin-4) | 8.6 | Mast cell, T-cell, granulocyte | Pro-inflammation |
| PDGF-BB (platelet-derived growth factor-BB) | 8.2 | Platelet, macrophage, and other cells | Wound healing and repair blood vessel |
| IL-9 (interleukin-9) | 7.9 | CD4+ T cell | Pro-inflammation |
| IL-1ra (interleukin-1 receptor antagonist) | 6.8 | Macrophage and other cells | Anti-inflammation |
| IL-5 (interleukin-5) | 6.7 | T cell, granulocyte, and other cells | Pro-inflammation |
| IL-15 (interleukin-15) | 5.4 | Macrophage | Pro-inflammation |
| IL-12(p70) (interleukin 12p70) | 3.6 | Macrophage | Pro-inflammation |
| Surface Marker | PDT Effect (Avg.) | Cell Type | Possible Biological Function |
|---|---|---|---|
| CD29 | 9.4 | White blood cells | Cell adhesion |
| CD9 | 8.8 | Lymphocyte, macrophage | Platelet activation and aggregation and cell adhesion and migration |
| CD40 | 7.8 | B-cell, macrophage | Cell proliferation and signal transduction |
| CD31 | 7.6 | White blood cells | Cell adhesion, activation, and migration |
| HLA-ABC | 7.5 | Nucleated cells | MHC class I molecules presented to CD8+ T cells |
| CD41b | 7.4 | Stem cell, platelet | Cell adhesion and platelet aggregation |
| CD69 | 7.4 | White blood cells | Lymphocyte activation and proliferation |
| HLA-DRDPDQ | 7.1 | Antigen-presenting cell | MHC class II molecules presented to CD4+ T cells |
| CD44 | 7.1 | White blood cells | Cell adhesion |
| CD81 | 6.5 | White blood cells but granulocyte | Cell adhesion |
| SSEA-4 | 6.5 | Embryonic stem cell | Pluripotent stem cell marker |
| CD8 | 5.9 | T cell | Cytotoxic T cell marker |
| CD25 | 5.3 | Lymphocyte and macrophage | Lymphocyte activation |
| CD19 | 4.9 | B cell | B cell marker |
| CD4 | 4.4 | T cell | Helper T cell marker |
| CD326 | 3.6 | T cell, dendritic cell, epithelial cell | Epithelial cell marker unknown functions on immune cells |
| CD133-1 | 3.1 | Stem cell and endothelial cell | Stem cell marker |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espeland, K.; Kleinauskas, A.; Juzenas, P.; Brech, A.; Darvekar, S.; Vasovic, V.; Warloe, T.; Christensen, E.; Jahnsen, J.; Peng, Q. Photodynamic Effects with 5-Aminolevulinic Acid on Cytokines and Exosomes in Human Peripheral Blood Mononuclear Cells. Biomedicines 2022, 10, 232. https://doi.org/10.3390/biomedicines10020232
Espeland K, Kleinauskas A, Juzenas P, Brech A, Darvekar S, Vasovic V, Warloe T, Christensen E, Jahnsen J, Peng Q. Photodynamic Effects with 5-Aminolevulinic Acid on Cytokines and Exosomes in Human Peripheral Blood Mononuclear Cells. Biomedicines. 2022; 10(2):232. https://doi.org/10.3390/biomedicines10020232
Chicago/Turabian StyleEspeland, Kristian, Andrius Kleinauskas, Petras Juzenas, Andreas Brech, Sagar Darvekar, Vlada Vasovic, Trond Warloe, Eidi Christensen, Jørgen Jahnsen, and Qian Peng. 2022. "Photodynamic Effects with 5-Aminolevulinic Acid on Cytokines and Exosomes in Human Peripheral Blood Mononuclear Cells" Biomedicines 10, no. 2: 232. https://doi.org/10.3390/biomedicines10020232