CRISPR/Cas9-Mediated Multiplex Genome Editing of JAGGED Gene in Brassica napus L.

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
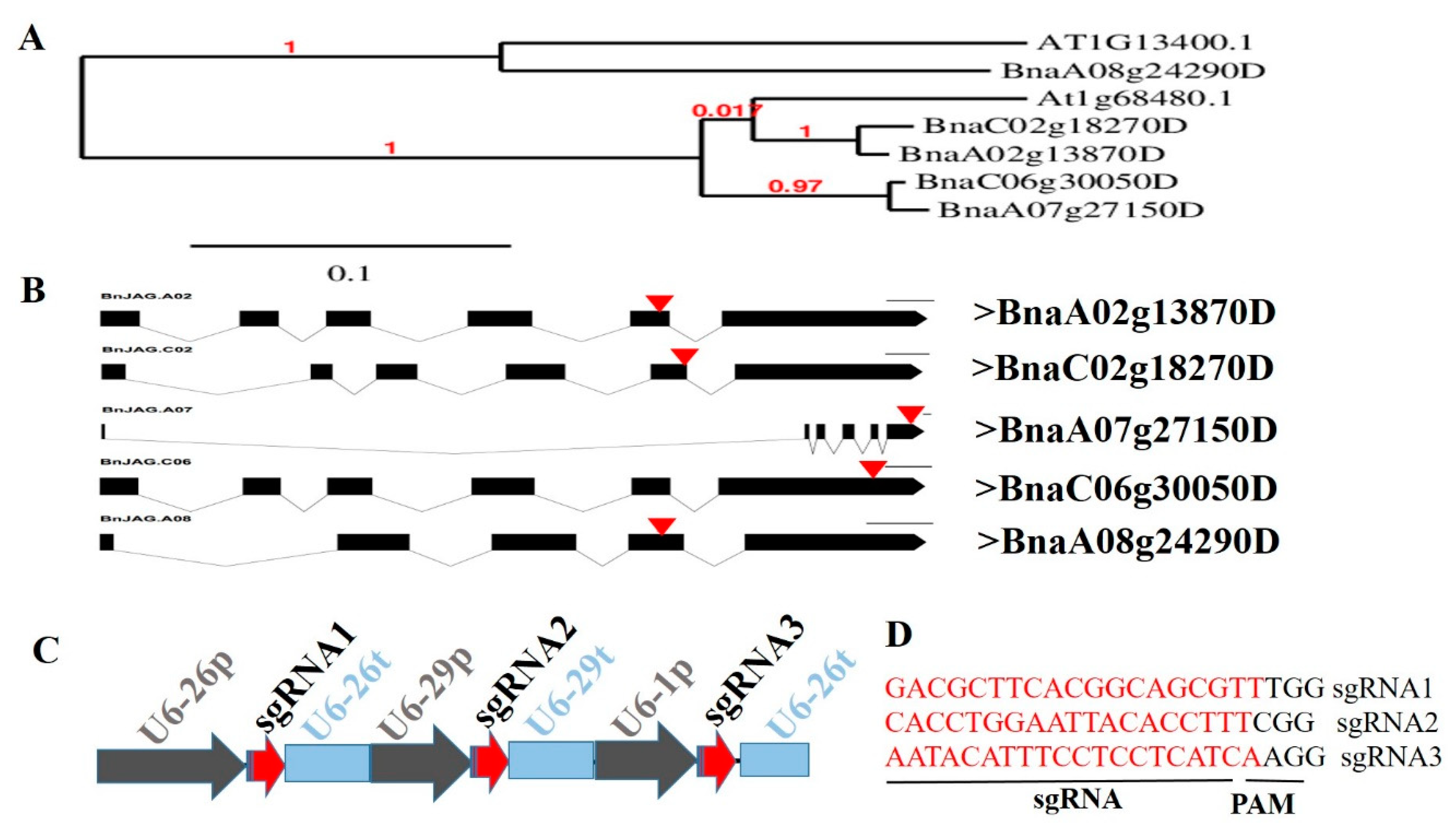
2.1. sgRNA Design and Vector Construction
2.2. Plant Materials and Vector Transformation
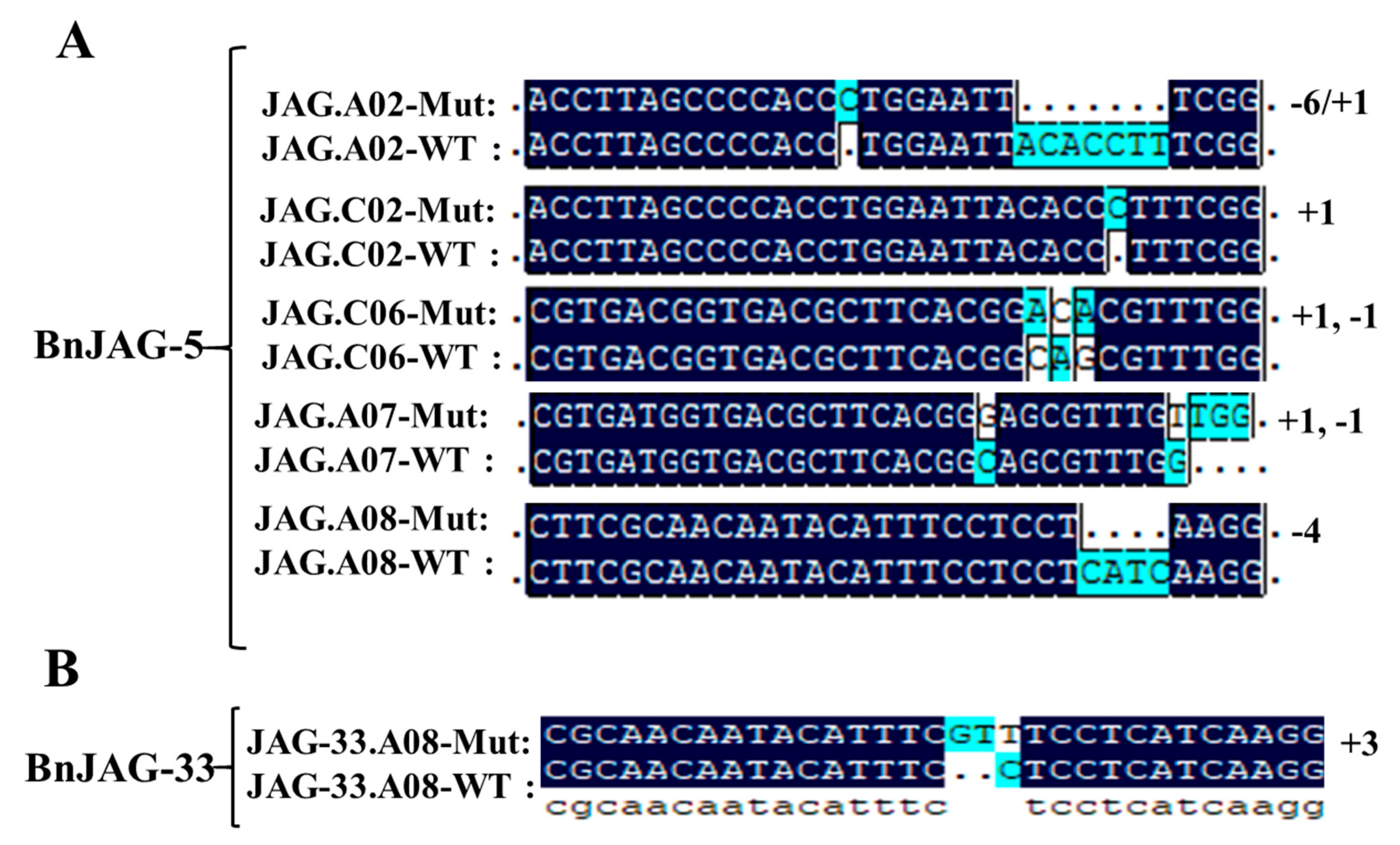
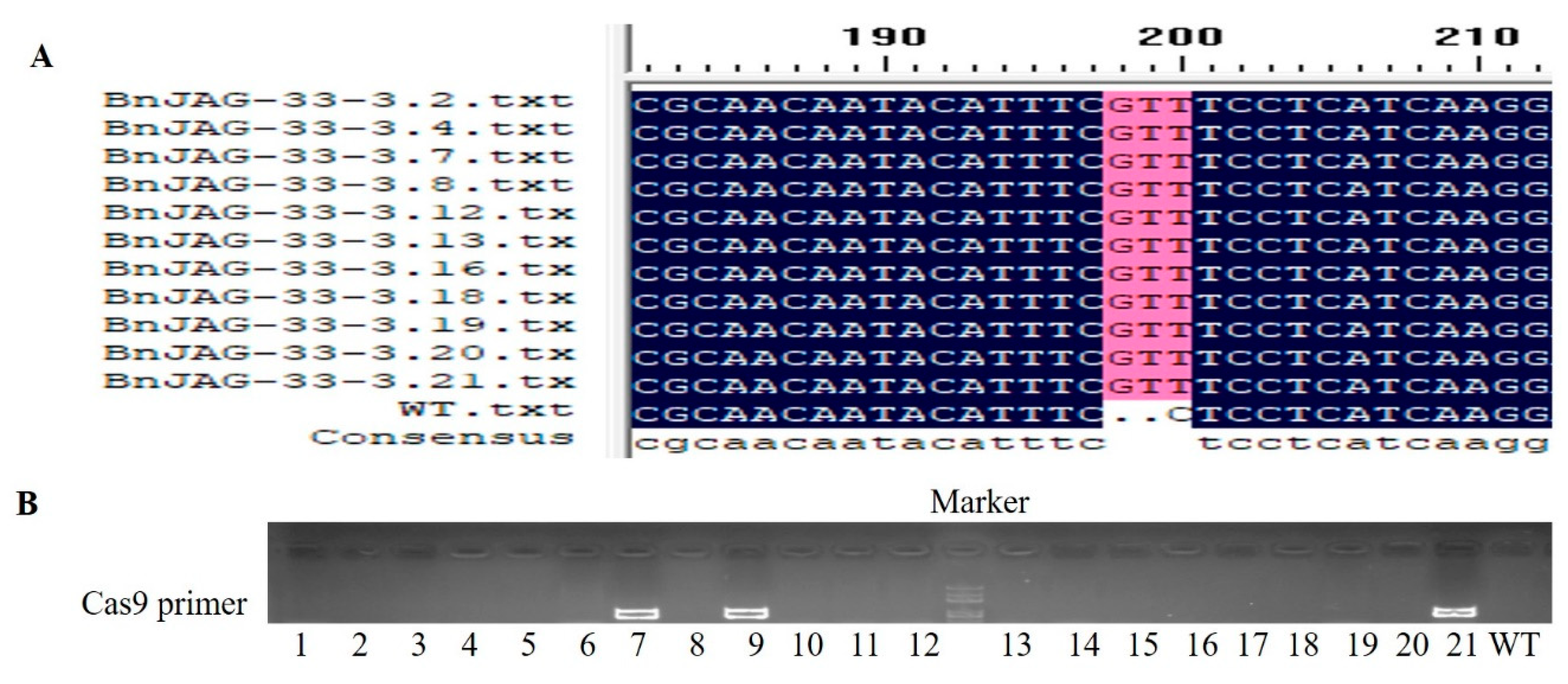
2.3. DNA Extraction and Identification of Positive Mutants
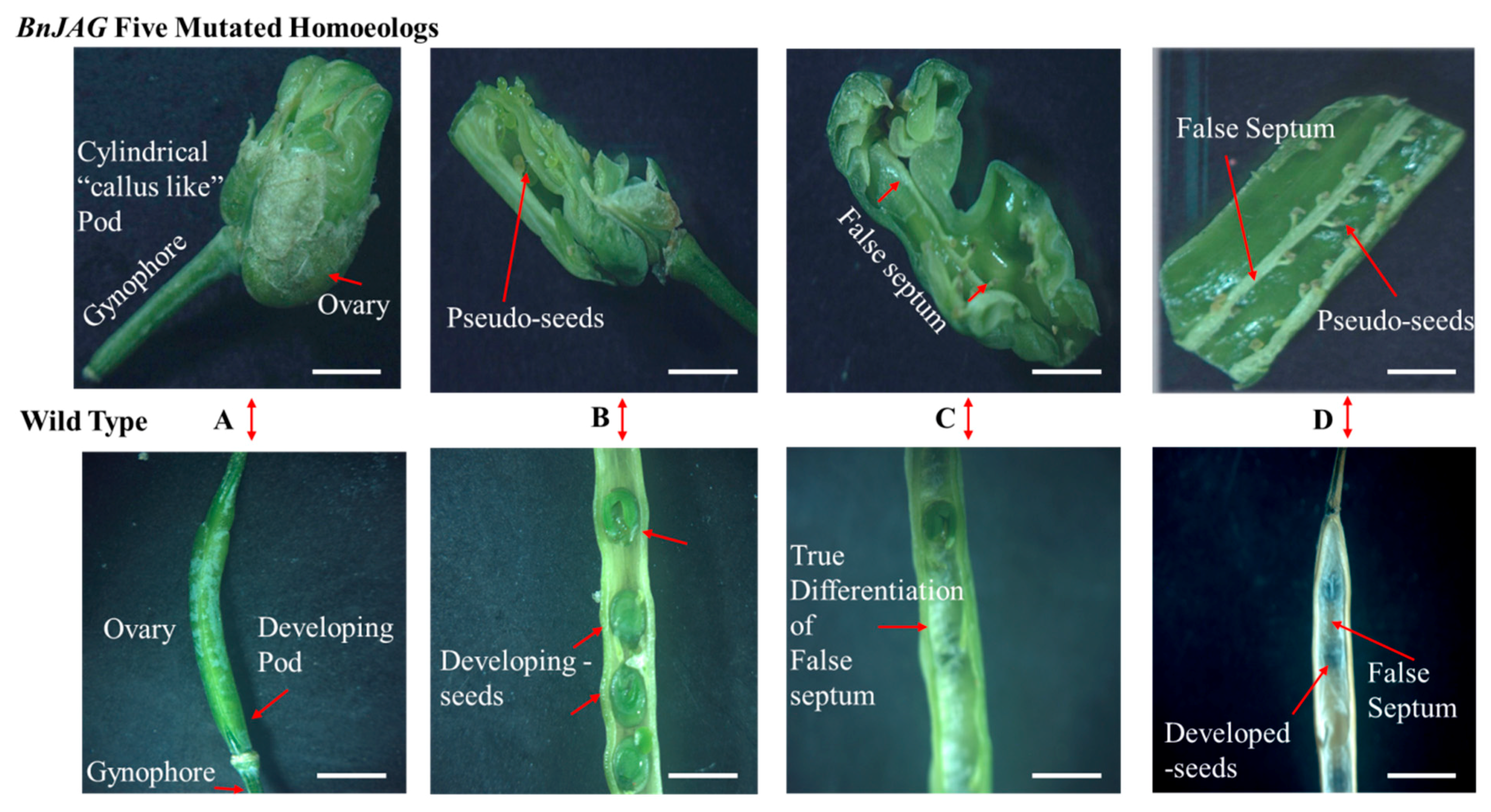
2.4. Phenotypic Characterization of Pods
2.5. RNA Isolation and Quantitative Real-Time PCR
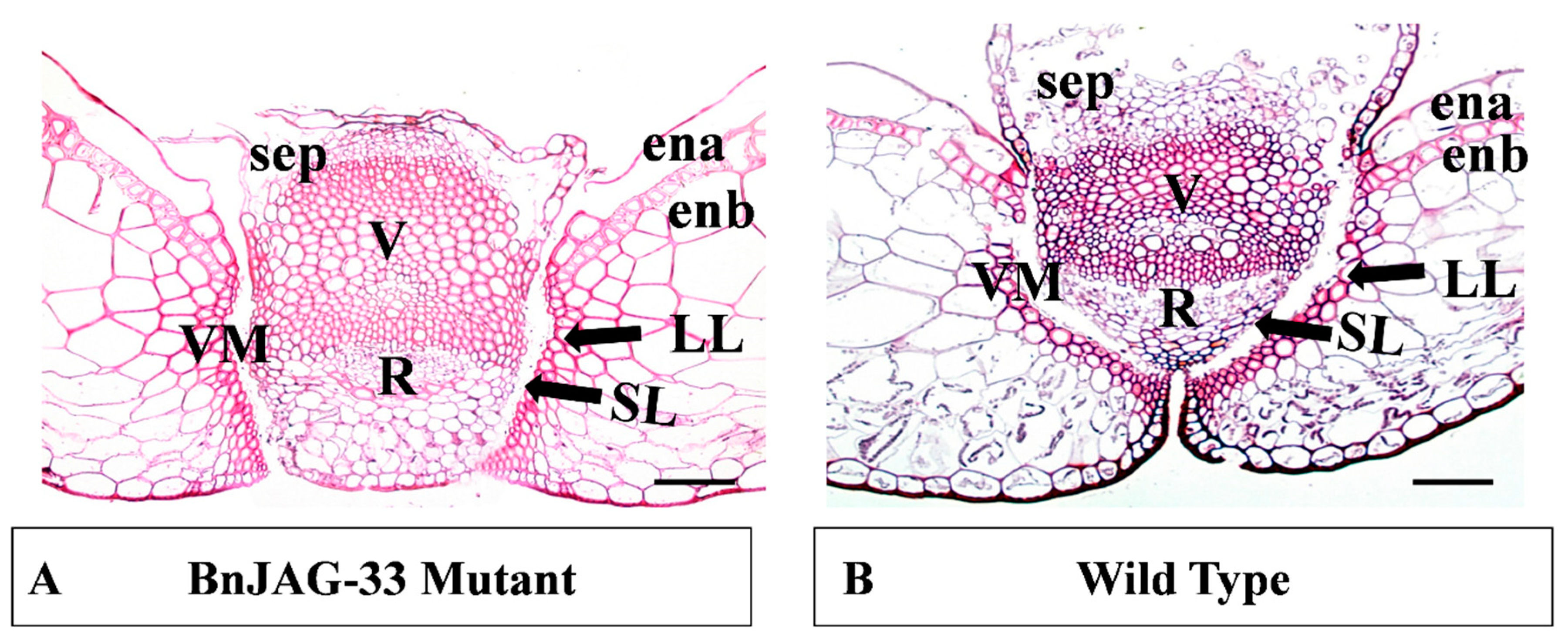
2.6. Staining of Pod Transverse Section of BnJAG.A08 Mutant
2.7. Statistical Analysis
3. Results
3.1. Sequence Analysis of BnJAG Gene
3.2. Knocking Out All Five JAG Homoeologs Dramatically Hampered Pod Development
3.3. Mutagenesis in BnJAG.A08 Enhanced Replum Width in the Dehiscence Zone
3.4. Inheritance Pattern of Mutagenesis at BnJAG.A08 Homoeolog
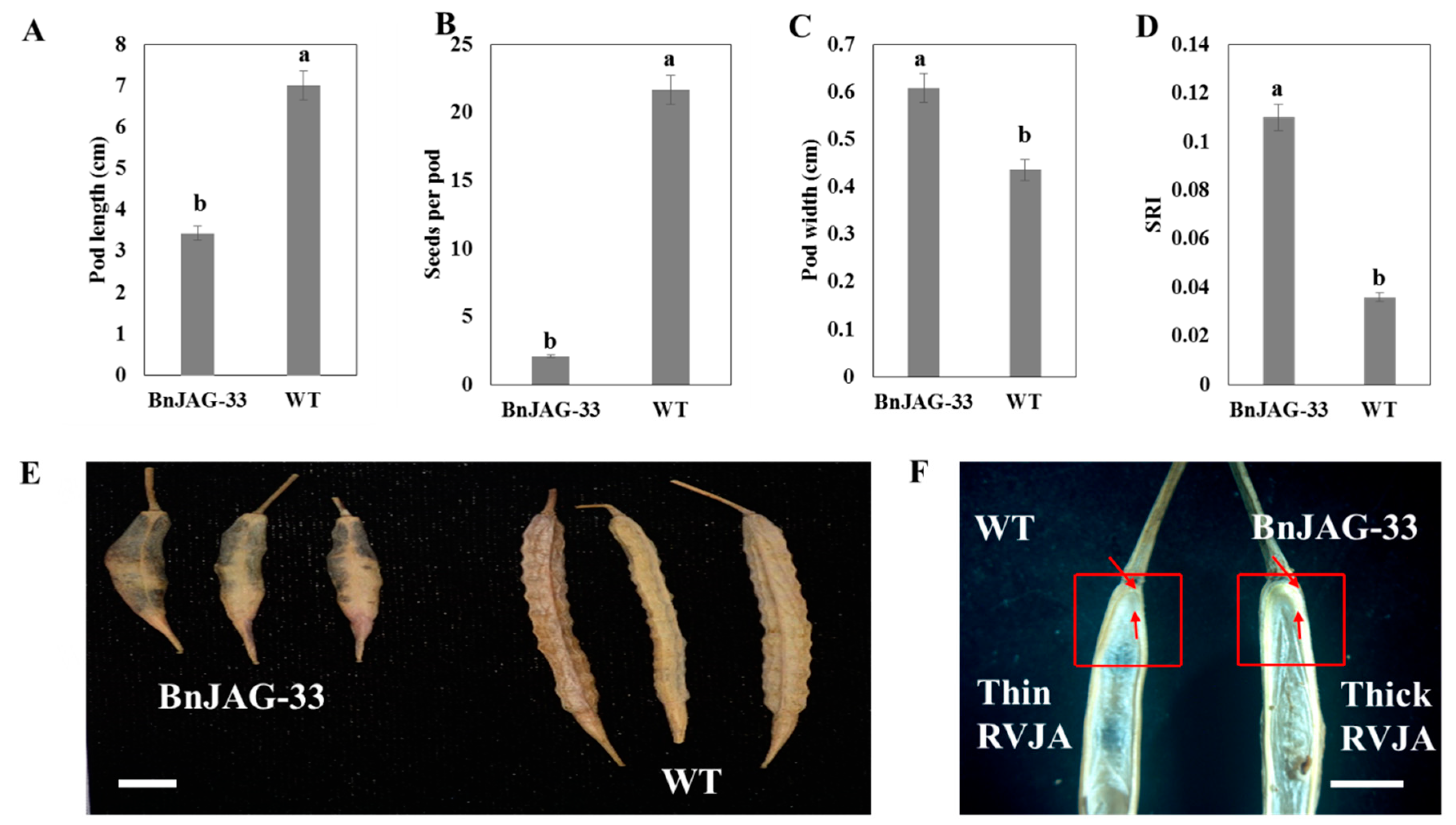
3.5. Variation of Pod Phenotype and SRI
3.6. Analysis of Transgene-Free Mutants
3.7. Off-Target Prediction
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nagaharu, U. Genome analysis in Brassica carinata with special reference to the experimental formation of Brassica napus, a peculiar mode of fertilization. Jpn. J. Bot. 1935, 7, 389–452. [Google Scholar]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.P.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Friedt, W.; Snowdon, R. Oilseed rape. In Handbook of Plant Breeding; Springer: New York, NY, USA, 2012; pp. 91–126. [Google Scholar]
- Prohens, J. Plant breeding: A success story to be continued thanks to the advances in genomics. Front. Plant. Sci. 2011, 2, 51. [Google Scholar] [CrossRef] [PubMed]
- Dresselhaus, T.; Hückelhoven, R.J. Biotic and Abiotic Stress Responses in Crop Plants. Agronomy 2018, 8, 267. [Google Scholar] [CrossRef]
- Hu, Q.; Wei, H.; Yan, Y.; Zhang, X.; Liu, L.; Shi, J.; Zhao, Y.; Qin, L.; Chen, C.; Wang, H. Rapeseed research and production in China. Crop. J. 2017, 5, 127–135. [Google Scholar] [CrossRef]
- Raman, H.; Raman, R.; Kilian, A.; Detering, F.; Carling, J.; Coombes, N.; Diffey, S.; Kadkol, G.; Edwards, D.; Mccully, M.; et al. Genome-wide delineation of natural variation for pod shatter resistance in Brassica napus. PLoS ONE 2014, 9, e101673. [Google Scholar] [CrossRef] [PubMed]
- Jie, K.; Sun, Y.; Liu, T.; Zhang, P.; Zhou, M.; Wu, J.; Zhou, G. Physiological mechanisms behind differences in pod shattering resistance in rapeseed (Brassica napus L.) Varieties. PLoS ONE 2016, 11, e0157341. [Google Scholar]
- Child, R.D.; Summers, J.E.; Babij, J.; Farrent, J.W.; Bruce, D.M. Increased resistance to pod shatter is associated with changes in the vascular structure in pods of a resynthesized Brassica napus line. J. Exp. Bot. 2003, 54, 1919–1930. [Google Scholar] [CrossRef] [PubMed]
- Child, R.D.; Huttly, A.K. Anatomical variation in the dehiscence zone of oilseed rape pods and its relevance to pod shatter. In Proceedings of the 10th International Rapeseed Congress, Canberra, Australia, 26–29 September 1999. [Google Scholar]
- Meakin, J.P.; Roberts, J.A. Dehiscence of fruit in oilseed rape (Brassica napus L.). I. Anatomy of pod dehiscence. J. Exp. Bot. 1990, 41, 995–1002. [Google Scholar] [CrossRef]
- Dinneny, J.R.; Weigel, D.; Yanofsky, M.F. A genetic framework for fruit patterning in Arabidopsis thaliana. Development 2005, 132, 4687–4697. [Google Scholar] [CrossRef] [PubMed]
- Eshed, Y.; Baum, S.F.; Bowman, J.L. Distinct mechanisms promote polarity establishment in carpels of Arabidopsis. Cell 1999, 99, 199–209. [Google Scholar] [CrossRef]
- Dinneny, J.R.; Yanofsky, M.F. Drawing lines and borders: How the dehiscent fruit of Arabidopsis is patterned. Bioessays 2005, 27, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Huang, Y.; Zhang, L.; Wang, X.; Liu, G.; Wang, H. BnLATE, a Cys2/His2-type zinc-finger protein, enhances silique shattering resistance by negatively regulating lignin accumulation in the silique walls of Brassica napus. PLoS ONE 2017, 12, e0168046. [Google Scholar] [CrossRef] [PubMed]
- Kord, H.; Shakib, A.M.; Daneshvar, M.H.; Azadi, P.; Bayat, V.; Mashayekhi, M.; Zarea, M.; Seifi, A.; Ahmadraji, M. RNAi-mediated down-regulation of SHATTERPROOF gene in transgenic oilseed rape. 3 Biotech 2015, 5, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, J.; Wang, H.; Wang, W.; Zhou, R.; Mei, D.; Cheng, H.; Yang, J.; Raman, H.; Hu, Q. Multigenic control of pod shattering resistance in chinese rapeseed germplasm revealed by genome-wide association and linkage analyses. Front. Plant Sci. 2016, 7, 476. [Google Scholar] [CrossRef] [PubMed]
- Peng, P.F.; Li, Y.C.; Mei, D.S.; Colasanti, J.; Fu, L.; Liu, J.; Chen, Y.F.; Hu, Q. Expression divergence of FRUITFULL homeologs enhanced pod shatter resistance in Brassica napus. Genet. Mol. Res. 2014, 14, 871–885. [Google Scholar] [CrossRef] [PubMed]
- Ogutcen, E.; Pandey, A.; Khan, M.K.; Marques, E.; Penmetsa, R.V.; Kahraman, A.; Von-Wettberg, E.J. Pod Shattering: A homologous series of variation underlying domestication and an avenue for crop improvement. Agronomy 2018, 8, 137. [Google Scholar] [CrossRef]
- Zhai, Y.; Cai, S.; Hu, L.; Yang, Y.; Amoo, O.; Fan, C.; Zhou, Y. CRISPR/Cas9-mediated genome editing reveals differences in the contribution of INDEHISCENT homologues to pod shatter resistance in Brassica napus L. Theor. Appl. Genet. 2019, 132, 2111–2123. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, P.; Stacey, N.; Brüser, M.; Pullen, N.; Ilyas, M.; O’Neill, C.; Wells, R.; Østergaard, L. The power of model-to-crop translation illustrated by reducing seed loss from pod shatter in oilseed rape. Plant Reprod. 2019, 32, 331–340. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Bai, M.; Tong, P.; Hu, Y.; Wu, H. CELLULASE 6 and MANNANASE 7 affect cell differentiation and silique dehiscence. Plant Physiol. 2018, 176, 2186–2201. [Google Scholar] [CrossRef] [PubMed]
- Groszmann, M.; Paicu, T.; Alvarez, J.P.; Swain, S.M.; Smyth, D.R. SPATULA and ALCATRAZ, are partially redundant, functionally diverging bHLH genes required for Arabidopsis gynoecium and fruit development. Plant J. 2011, 68, 816–829. [Google Scholar] [CrossRef] [PubMed]
- Kerstetter, R.A.; Bollman, K.; Taylor, R.A.; Bomblies, K.; Poethig, R.S. KANADI regulates organ polarity in Arabidopsis. Nature 2001, 411, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.; Smyth, D.R. CRABS CLAW and SPATULA genes regulate growth and pattern formation during gynoecium development in Arabidopsis thaliana. Int. J. Plant Sci. 2002, 163, 17–41. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Zhang, W.; Yu, F.; Tian, J.; Li, D.; Guo, A. Functional analysis of the two brassica ap3 genes involved in apetalous and stamen carpelloid phenotypes. PLoS ONE 2011, 6, e20930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinneny, J.R.; Weigel, D.; Yanofsky, M.F. NUBBIN and JAGGED define stamen and carpel shape in Arabidopsis. Development 2006, 133, 1645–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinneny, J.R.; Yadegari, R.; Fischer, R.L.; Yanofsky, M.F.; Weigel, D. The role of JAGGED in shaping lateral organs. Development 2004, 131, 1101–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hepworth, S.R.; Zhang, Y.; McKim, S.; Li, X.; Haughn, G.W. BLADE-ON-PETIOLE-dependent signaling controls leaf and floral patterning in arabidopsis. Plant Cell 2005, 17, 1434–1448. [Google Scholar] [CrossRef] [PubMed]
- Ha, C.M.; Kim, G.T.; Jun, J.H.; Soh, M.S.; Ueno, Y.; Machida, Y.; Tsukaya, H.; Nam, H.G. The BLADE-ON-PETIOLE 1 gene controls leaf pattern formation through the modulation of meristematic activity in Arabidopsis. Development 2003, 130, 161–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nath, U.; Brian, C.W.C.; Rosemary, C.; Enrico, C. Genetic control of surface curvature. Science 2003, 299, 1404–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palatnik, J.F.; Allen, E.; Wu, X.; Schommer, C.; Schwab, R.; Carrington, J.C.; Weigel, D. Control of leaf morphogenesis by microRNAs. Nature 2003, 425, 257–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, C.K.; Reddy, G.V.; Heisler, M.G.; Meyerowitz, E.M. The Arabidopsis JAGGED gene encodes a zinc finger protein that promotes leaf tissue development. Development 2004, 131, 1111–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Yan, S.; Jiang, L.; Zhao, W.; Ning, K.; Zhao, J.; Liu, X.; Zhang, J.; Wang, Q.; Zhang, X. HANABA TARANU (HAN) bridges meristem and organ primordia boundaries through PINHEAD, JAGGED, BLADE-ON-PETIOLE2 and CYTOKININ OXIDASE 3 during flower development in arabidopsis. PLoS Genet. 2015, 11, e1005479. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hao, M.; Wang, W.; Wang, H.; Chen, F.; Chu, W.; Zhang, B.; Mei, D.; Cheng, H.; Hu, Q. An efficient CRISPR/Cas9 platform for rapidly generating simultaneous mutagenesis of multiple gene homoeologs in allotetraploid oilseed rape. Front. Plant Sci. 2018, 9, 442. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Ding, Y.; Zhou, Y.; Jin, W.; Xie, K.; Cheng, L. CRISPR-P 2.0: An improved CRISPR/Cas9 tool for genome editing in plants. Mol. Plant 2017, 10, 530–532. [Google Scholar]
- Balanzã, V.; Roig-Villanova, I.; Di, M.M.; Masiero, S.; Colombo, L. Seed abscission and fruit dehiscence required for seed dispersal rely on similar genetic networks. Development 2016, 143, 3372–3381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, D.B. Multiple Range and Multiple F Tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Sauret-Güeto, S.; Schiessl, K.; Bangham, A.; Sablowski, R.; Coen, E. JAGGED controls arabidopsis petal growth and shape by interacting with a divergent polarity field. PLoS Biol. 2013, 11, e1001550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegfried, K.R.; Eshed, Y.; Baum, S.F.; Otsuga, D.; Drews, G.N.; Bowman, J.L. Members of the YABBY gene family specify abaxial cell fate in Arabidopsis. Development 1999, 126, 4117. [Google Scholar] [PubMed]
- Lampugnani, E.R.; Kilinc, A.; Smyth, D.R. Auxin controls petal initiation in Arabidopsis. Development 2013, 140, 185–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borghi, L.; Bureau, M.; Simon, R. Arabidopsis JAGGED LATERAL ORGANS is expressed in boundaries and coordinates knox and pin activity. Plant Cell 2007, 19, 1795–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolland-Lagan, A.-G.; Prusinkiewicz, P. Reviewing models of auxin canalization in the context of leaf vein pattern formation in Arabidopsis. Plant J. 2010, 44, 854–865. [Google Scholar] [CrossRef] [PubMed]
- Schiessl, K.; Muino, J.M.; Sablowski, R. Arabidopsis JAGGED links floral organ patterning to tissue growth by repressing Kip-related cell cycle inhibitors. Proc. Natl. Acad. Sci. USA 2014, 111, 2830–2835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnelly, P.M.; Bonetta, D.; Tsukaya, H.; Dengler, R.E.; Dengler, N.G. Cell cycling and cell enlargement in developing leaves of arabidopsis. Dev. Biol. 1999, 215, 407–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Reig, S.; Ripoll, J.J.; Vera, A.; Yanofsky, M.F.; Martínez-Laborda, A. Antagonistic gene activities determine the formation of pattern elements along the mediolateral axis of the arabidopsis fruit. PLoS Genet. 2012, 8, e1003020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, Y.; Kramer, E.M. The Aquilegia JAGGED homolog promotes proliferation of adaxial cell types in both leaves and stems. New Phytol. 2017, 216, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Horigome, A.; Nagasawa, N.; Ikeda, K.; Ito, M.; Itoh, J.I.; Nagato, Y. Rice OPEN BEAK is a negative regulator of class 1 knox genes and a positive regulator of class B floral homeotic gene. Plant J. 2010, 58, 724–736. [Google Scholar] [CrossRef] [PubMed]
- David-Schwartz, R.; Koenig, D.; Sinha, N.R. LYRATE is a key regulator of leaflet initiation and lamina outgrowth in tomato. Plant Cell 2009, 21, 3093–3104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, B.; Li, Z.; Zhu, Y.; Wang, H.; Ma, H.; Dong, A.; Huang, H. Arabidopsis genes AS1, AS2, and JAG negatively regulate boundary-specifying genes to promote sepal and petal development. Plant Physiol. 2008, 146, 566–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Yang, H.; Zhang, L.; Wang, X.; Liu, G.; Wang, H.; Hua, W. A large replum-valve joint area is associated with increased resistance to pod shattering in rapeseed. J. Plant Res. 2015, 128, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.S.; Lee, J.H.; Lee, J.S.; Ahn, J.H. Fruit indehiscence caused by enhanced expression of NO TRANSMITTING TRACT in Arabidopsis thaliana. Mol. Cells 2013, 35, 519–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avino, M.; Kramer, E.M.; Donohue, K.; Hammel, A.J.; Hall, J.C. Understanding the basis of a novel fruit type in Brassicaceae: Conservation and deviation in expression patterns of six genes. EvoDevo 2012, 3, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | BnJAG-33 (M) | Wild Type (WT) | Variance | Homogeneity | |||
|---|---|---|---|---|---|---|---|
| Range | Mean | Range | Mean | Treatment a | M b | WT c | |
| Pod length (cm) | 3.18–3.71 | 3.432 ± 0.17 | 6.42–7.5 | 7.008 ± 0.36 | *** | b | a |
| Seeds per pod | 1.1–3.1 | 2.11 ± 0.64 | 20.1–23.7 | 21.67 ± 1.21 | *** | b | a |
| Pod width (cm) | 0.58–0.63 | 0.608 ± 0.01 | 0.41–0.46 | 0.436 ± 0.01 | *** | a | b |
| SRI | 0.11 | 0.036 | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaman, Q.U.; Chu, W.; Hao, M.; Shi, Y.; Sun, M.; Sang, S.-F.; Mei, D.; Cheng, H.; Liu, J.; Li, C.; et al. CRISPR/Cas9-Mediated Multiplex Genome Editing of JAGGED Gene in Brassica napus L. Biomolecules 2019, 9, 725. https://doi.org/10.3390/biom9110725
Zaman QU, Chu W, Hao M, Shi Y, Sun M, Sang S-F, Mei D, Cheng H, Liu J, Li C, et al. CRISPR/Cas9-Mediated Multiplex Genome Editing of JAGGED Gene in Brassica napus L. Biomolecules. 2019; 9(11):725. https://doi.org/10.3390/biom9110725
Chicago/Turabian StyleZaman, Qamar U, Wen Chu, Mengyu Hao, Yuqin Shi, Mengdan Sun, Shi-Fei Sang, Desheng Mei, Hongtao Cheng, Jia Liu, Chao Li, and et al. 2019. "CRISPR/Cas9-Mediated Multiplex Genome Editing of JAGGED Gene in Brassica napus L." Biomolecules 9, no. 11: 725. https://doi.org/10.3390/biom9110725