An Efficient Agrobacterium Mediated Transformation of Pineapple with GFP-Tagged Protein Allows Easy, Non-Destructive Screening of Transgenic Pineapple Plants

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
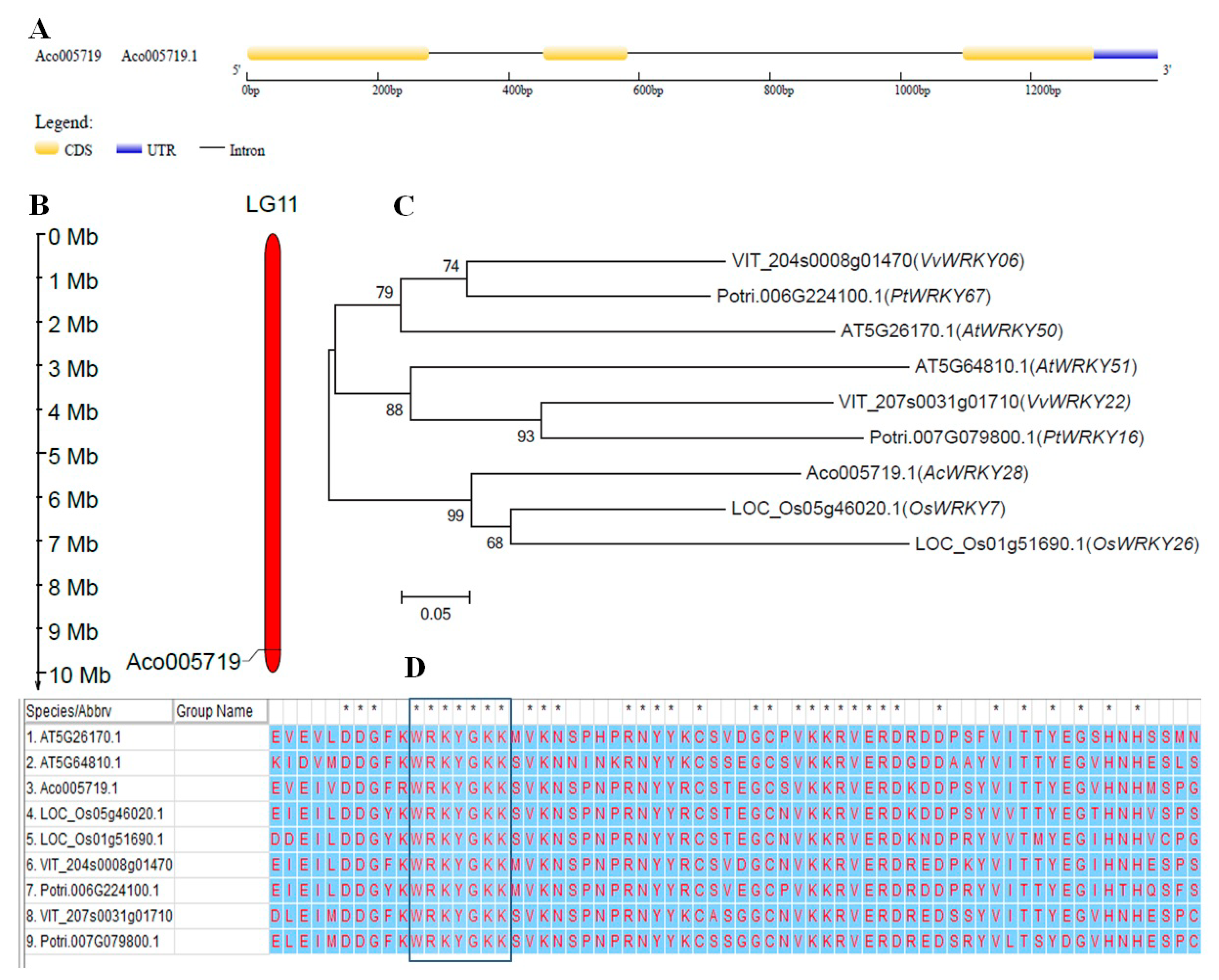
2.1. Bioinformatics Analysis of AcWRKY28
2.2. Plant Material Preparation
2.3. Vector Construction and Agrobacterium Culture Preparation for Transformation
2.4. Pineapple Transformation
2.5. Optimization of Factors Influencing Transformation Efficiency
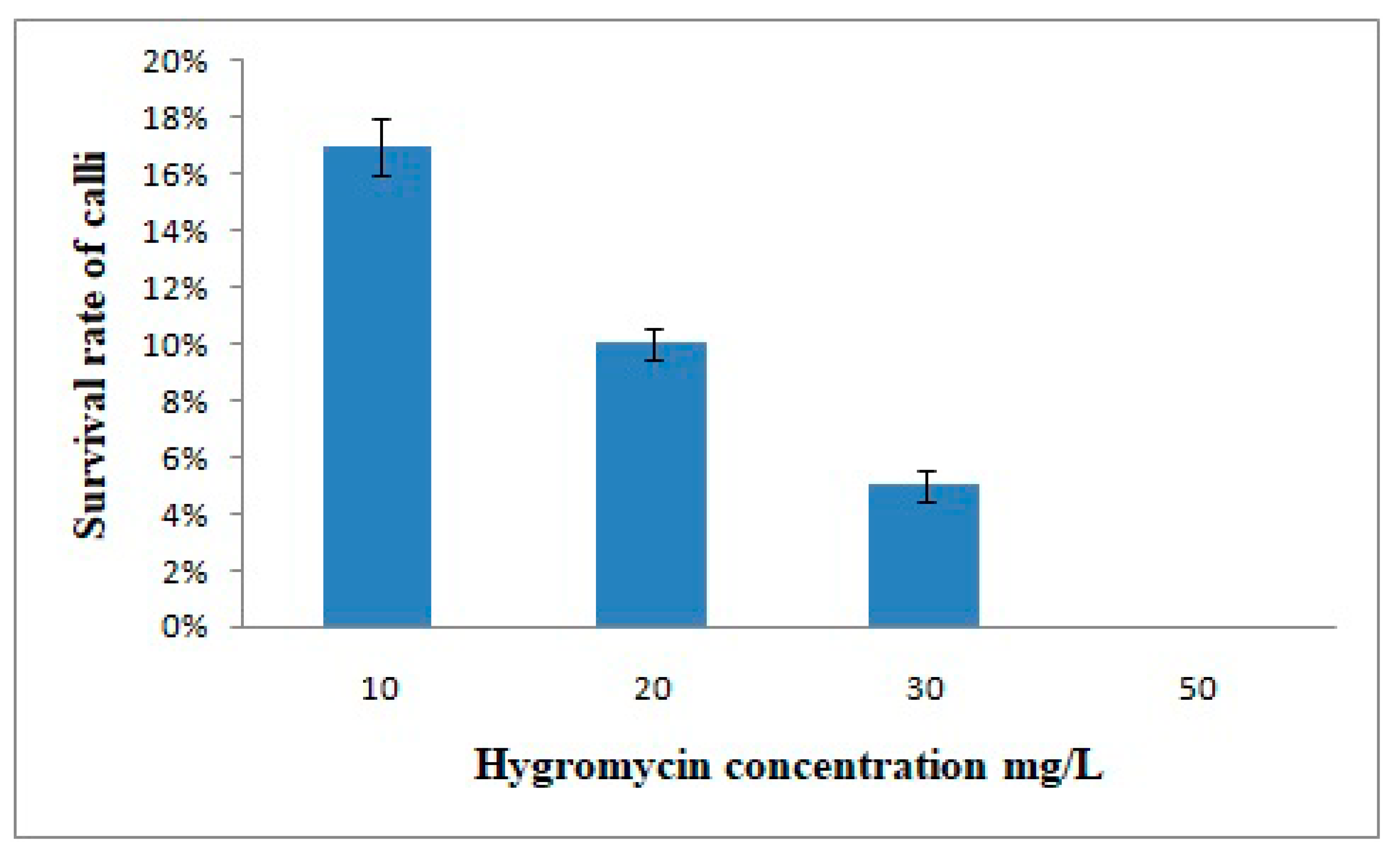
2.6. Optimization of Hygromycin Concentration for the Selection of Putative Transgenic Lines
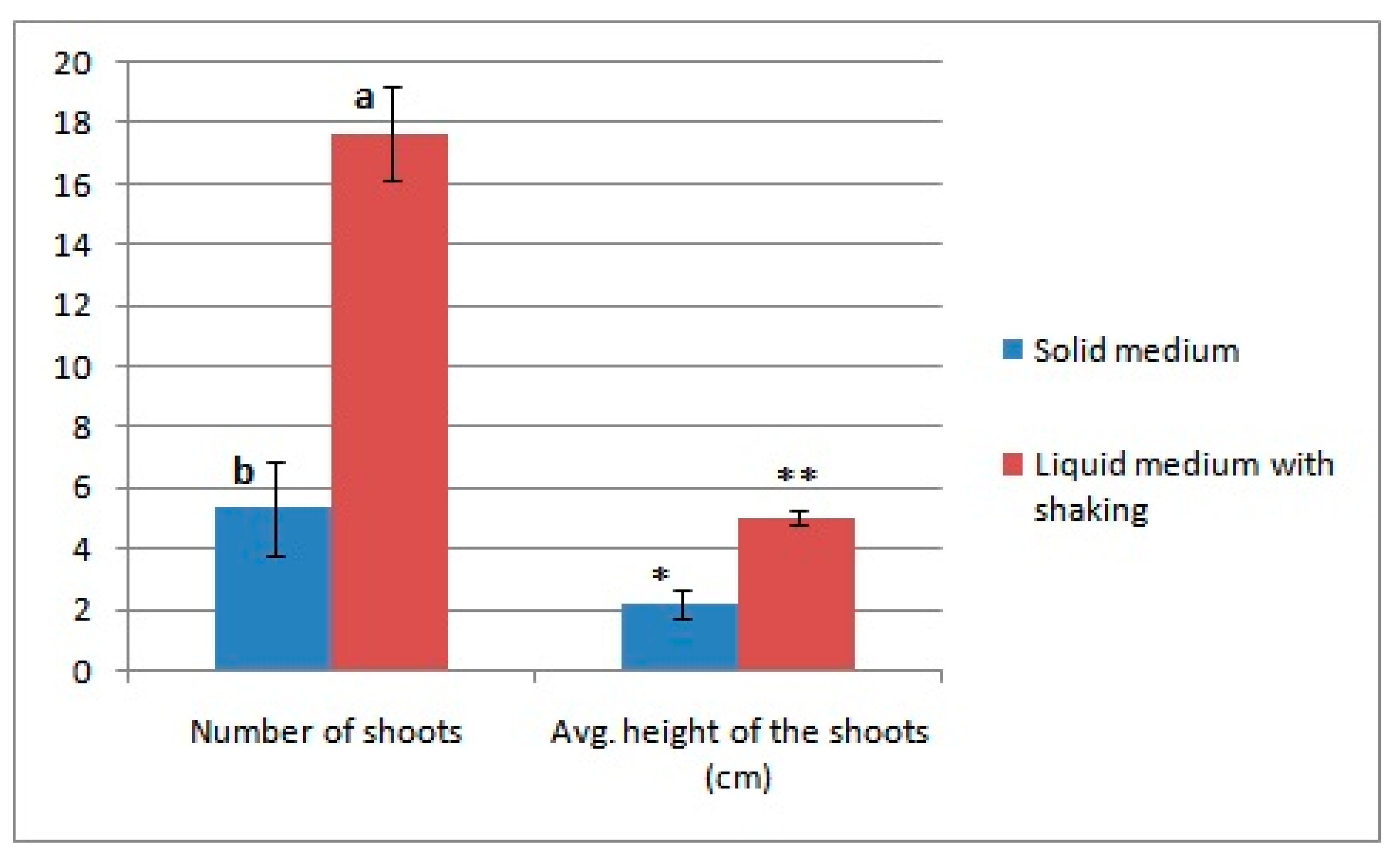
2.7. Effect of Different Growth Conditions on Callus Growth after Selection
2.8. Data Collection and Statistical Analysis
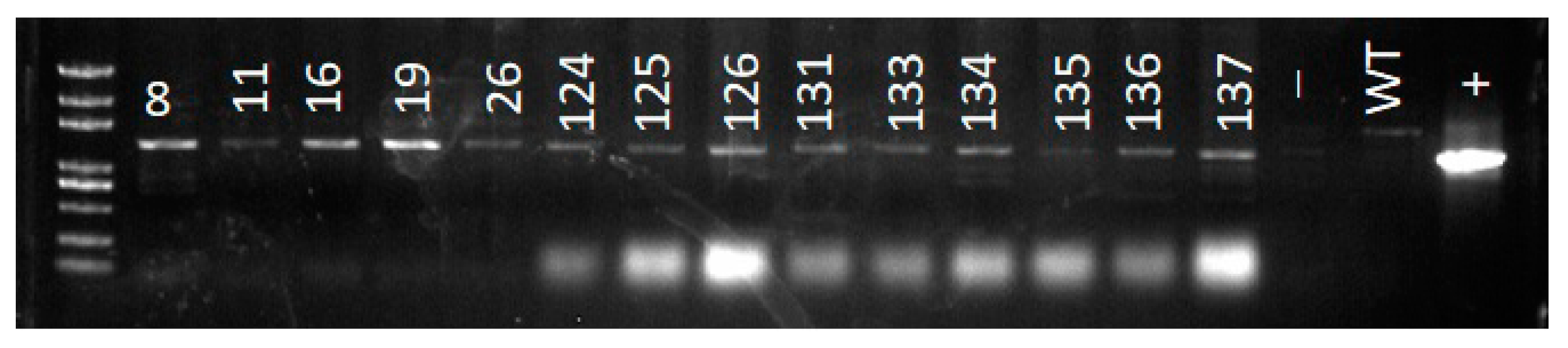
2.9. DNA Extraction and PCR Analysis
2.10. Microscopy
3. Results
3.1. Sequence Analysis
3.2. Optimization of Incubation Time for Infection, Co-Cultivation Duration, and Acetosyringone Concentration in the Co-Cultivation Medium
3.3. Optimization of Hygromycin Concentration for the Selection of Transgenic Lines
3.4. Effect of Different Growth Conditions on Callus Growth after Selection
3.5. PCR Analysis
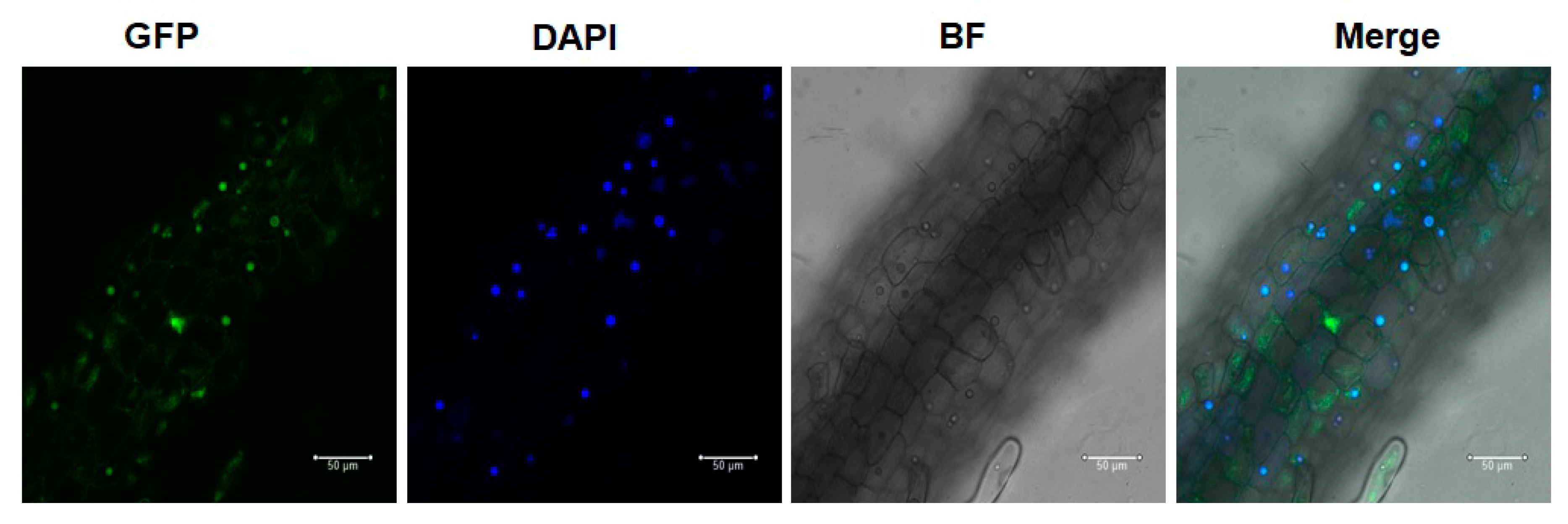
3.6. GFP-AcWRKY28 Is Localized in the Nucleus
4. Discussion
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TF | transcription factor |
| OD | Optical density |
| DNA | Deoxyribonucleic acid |
| GFP | Green flourascent protein |
References
- Azam, S.M.; Liu, Y.; Rahman, Z.U.; Ali, H.; Yan, C.; Wang, L.; Priyadarshani, S.V.G.N.; Hu, B.; Huang, X.; Xiong, J.; et al. Identification, Characterization and Expression Profiles of Dof Transcription Factors in Pineapple (Ananas comosus L.). Trop. Plant Biol. 2018, 11, 49–64. [Google Scholar] [CrossRef]
- Rahman, Z.U.; Azam, S.M.; Liu, Y.; Yan, C.; Ali, H.; Zhao, L.; Chen, P.; Yi, L.; Priyadarshani, S.V.G.N.; Yuan, Q. Expression Profiles of Wuschel-Related Homeobox Gene Family in Pineapple (Ananas comosus L.). Trop. Plant Biol. 2017, 10, 204–215. [Google Scholar] [CrossRef]
- Kato, C.Y.; Nagai, C.; Moore, P.H.; Zee, F.; Kim, M.S.; Steiger, D.L.; Ming, R. Intra-specific DNA polymorphism in pineapple (Ananas comosus (L.) Merr.) assessed by AFLP markers. Genet. Resour. Crop Evol. 2004, 51, 815–825. [Google Scholar] [CrossRef]
- Ko, H.; Campbell, P.; Jobin-Décor, M.; Eccleston, K.; Graham, M.; Smith, M. The introduction of transgenes to control blackheart in pineapple (Ananas comosus L.) cv. Smooth cayenne by micro projectile bombardment. Euphytica 2006, 150, 387–395. [Google Scholar] [CrossRef]
- Sripaoraya, S. Herbicide-tolerant Transgenic Pineapple (Ananas comosus) Produced by Microprojectile Bombardment. Ann. Bot. 2001, 88, 597–603. [Google Scholar] [CrossRef]
- Trusov, Y.; Botella, J.R. Silencing of the ACC synthase gene ACACS2 causes delayed flowering in pineapple [Ananas comosus (L.) Merr.]. J. Exp. Bot. 2006, 57, 3953–3960. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Uruu, G.; Xiong, L.; He, X.; Nagai, C.; Cheah, K.T.; Hu, J.S.; Nan, G.L.; Sipes, B.S.; Atkinson, H.J.; et al. Production of transgenic pineapple (Ananas cosmos (L.) Merr.) plants via adventitious bud regeneration. In Vitro Cell. Dev. Biol. Plant 2009, 45, 112–121. [Google Scholar] [CrossRef]
- Thomas Martin, T.; Wöhner, R.-V.; Hummel, S.; Willmitzer, L.; Frommer, W.B. The GUS Reporter System as a Tool to Study Plant Gene Expression. In Gus Protocols; Gallagher, S.R., Ed.; Academic Press: Cambridge, MA, USA, 1992; pp. 23–43. [Google Scholar]
- Guivarc’H, A.; Caissard, J.C.; Azmi, A.; Elmayan, T.; Chriqui, D.; Tepfer, M. In situ detection of expression of the gus reporter gene in transgenic plants: Ten years of blue genes. Transgenic Res. 1996, 5, 281–288. [Google Scholar] [CrossRef]
- Santi, C.; Svistoonoff, S.; Constans, L.; Auguy, F.; Duhoux, E.; Bogusz, D.; Claudine Franche, C. Choosing a reporter for gene expression studies in transgenic actinorhizal plants of the Casuarinaceae family. Plant Soil 2003, 254, 229–237. [Google Scholar] [CrossRef]
- Jian, L.; Weijie, J.; Ying, Z.; Hongjun, Y.; Zhenchuan, M.; Xingfang, G.; Sanwen, H.; Bingyan, X. Genome-wide analysis of WRKY gene family in Cucumis sativus. BMC Genom. 2011, 12. [Google Scholar] [CrossRef]
- Phukan, U.J.; Jeena, G.S.; Shukla, R.K. WRKY Transcription Factors: Molecular Regulation and Stress Responses in Plants. Front. Plant Sci. 2016, 7, 760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, T.; Chen, C.; Li, C.; Liu, J.; Liu, C.; He, Y. Genome-wide investigation of WRKY gene family in pineapple: Evolution and expression profiles during development and stress. BMC Genom. 2018, 19, 490. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.M.; Venugopal, S.; Navarre, D.; Kachroo, A. Low oleic acid-derived repression of jasmonic acid-inducible defense responses requires the WRKY50 and WRKY51 proteins. Plant Physiol. 2011, 155, 464–476. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Cai, C.; Ren, H.; Wang, W.; Xiang, M.; Tang, X.; Zhu, C.; Yin, T.; Zhang, L.; Zhu, Q. An Efficient Plant Regeneration and Transformation System of Ma Bamboo (Dendrocalamus latiflorus Munro) Started from Young Shoot as Explant. Front. Plant Sci. 2017, 8, 1298. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef]
- Ma, J.; He, Y.H.; Wu, C.H.; Liu, H.P.; Hu, Z.Y.; Sun, G.M. Effective Agrobacteriummediated transformation of pineapple with CYP1A1 by kanamycin selection technique. Afr. J. Biotechnol. 2012, 11, 2555–2562. [Google Scholar] [CrossRef] [Green Version]
- Priyadarshani, S.; Hu, B.; Li, W.; Ali, H.; Jia, H.; Zhao, L.; Ojolo, S.P.; Azam, S.M.; Xiong, J.; Yan, M.; et al. Simple protoplast isolation system for gene expression and protein interaction studies in pineapple (Ananas comosus L.). Plant Methods 2018, 14, 95. [Google Scholar] [CrossRef]
- Firoozabady, E.; Heckert, M.; Gutterson, N. Transformation and regeneration of pineapple. Plant Cell Tissue Organ Cult. 2005, 84, 1–16. [Google Scholar] [CrossRef]
- Nonaka, S.; Ezura, H. Plant-Agrobacterium interaction mediated by ethylene and super-Agrobacterium conferring efficient gene transfer. Front. Plant Sci. 2014, 5, 681. [Google Scholar] [CrossRef]
- Davis, M.E.; Miller, A.R.; Lineberger, R.D. Studies on the Effects of Ethylene on Transformation of Tomato Cotyledons (Lycopersicon esculentum Mill.) by Agrobacterium tumefaciens. J. Plant Physiol. 1992, 139, 309–312. [Google Scholar] [CrossRef]
- Ezura, H.; Yuhashi, K.; Yasuta, T.; Minamisawa, K. Effect of ethylene on Agrobacterium tumefaciens-mediated gene transfer to melon. Plant Breed. 2000, 75–79. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Media | Composition |
|---|---|
| Luria broth | Tryptophan 10 g, yeast extract 5 g, NaCl 10 g |
| AAM-AS (1 L) pH = 5.2 | AA macro nutrient 10× (100 mL)–(KH2PO4-170 mg/L, MgSO4.7H2O-370 mg/L, KCl-2940 mg/L, CaCl2-440 mg/L) AA micro nutrient 100× (10 mL)–(MnSO4.4H2O-10 mg/L, H3BO4-3 mg/L, ZnSO4.7H2O-2 mg/L, KI-0.75 mg/L) AA micro nutrient 1000× (1 mL)–(NaMoO4.2H2O-0.25 mg/L, CuSO4.5H2O-0.0387 mg/L, CoCl2.6H2O-0.025 mg/L) Ferrous 100× (10 mL) AAM-AS organic 100× (10 mL)–(glycine-7.5 mg/L, L-arginine-174 mg/L, L-glutamine-876 mg/L, inositol-100 mg/L, nicotinic acid-0.5 mg/L, vitamin B1-0.5 mg/L, vitamin B6-0.5 mg/L, casamino acid-500 mg/L) Sugar 30 g, glucose 68.5 g, 200 µM acetosyringone |
| Co-cultivation medium | 4.43 g/L MS powder, 30 g/L sugar, 3 g/L phytagel, 1 mg/L BAP, 0.2 mg/L NAA, 200 µmol/L acetosyringone pH = 5.8 |
| Proliferation medium | 4.43 g/L MS powder, 30 g/L sugar, 3 g/L phytagel, 1 mg/L BAP, 0.2 mg/L NAA, 200 mg/L carbenicillin pH = 5.8 |
| Selection medium | 4.43 g/L MS powder, 30 g/L sugar, 3 g/L phytagel, 1 mg/L BAP, 0.2 mg/L NAA, 200 mg/L carbenicillin, 20 mg/L hygromycin pH = 5.8 |
| Rooting medium | 4.43 g/L MS powder, 30 g/L sugar, 3 g/L phytagel, 0.2 mg/L NAA |
| Reaction Component | Volume |
|---|---|
| Taq buffer | 10 µL |
| Forward primer | 0.5 µL |
| Reverse primer | 0.5 µL |
| Sterilized distilled water | 8 µL |
| Template DNA | 1 µL |
| Total | 20 µL |
| Incubation Duration | Contaminated % | Death % before Selection | Growth % | Death % at 1st Selection |
|---|---|---|---|---|
| ≤1 h | 5 ± 0.0050 c | 1 ± 0.000 C | 84 ± 0.011 * | 95 ± 0.000 d |
| 1 h ≥ t ≤ 6 h | 7 ± 0.0100 c | 1 ± 0.000 C | 84 ± 0.010 * | 79 ± 0.000 e |
| 8 h | 24 ± 0.0115 b | 20 ± 0.020 B | 10 ± 0.000 ** | 79 ± 0.040 e |
| 12 h | 71 ± 0.0153 a | 84 ± 0.040 A | 5 ± 0.000 *** | 81 ± 0.026 e |
| Transformation Step | Time (Days) |
|---|---|
| Transformation with Agrobacterium | 1 |
| Co-cultivation | 3 |
| Non-selection medium | 7 |
| First selection medium | 28 |
| Non-selection medium | 28 |
| Second selection medium | 14 |
| Non-selection medium | 28 |
| Third selection medium | 14 |
| Non-selection medium | 56 |
| Plant selection medium | 7 |
| Selected plants grown in rooting medium before transfer to soil | 28 |
| Total time | 214 ≈ 7.6 months |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Priyadarshani, S.V.G.N.; Cai, H.; Zhou, Q.; Liu, Y.; Cheng, Y.; Xiong, J.; Patson, D.L.; Cao, S.; Zhao, H.; Qin, Y. An Efficient Agrobacterium Mediated Transformation of Pineapple with GFP-Tagged Protein Allows Easy, Non-Destructive Screening of Transgenic Pineapple Plants. Biomolecules 2019, 9, 617. https://doi.org/10.3390/biom9100617
Priyadarshani SVGN, Cai H, Zhou Q, Liu Y, Cheng Y, Xiong J, Patson DL, Cao S, Zhao H, Qin Y. An Efficient Agrobacterium Mediated Transformation of Pineapple with GFP-Tagged Protein Allows Easy, Non-Destructive Screening of Transgenic Pineapple Plants. Biomolecules. 2019; 9(10):617. https://doi.org/10.3390/biom9100617
Chicago/Turabian StylePriyadarshani, S. V. G. N., Hanyang Cai, Qiao Zhou, Yanhui Liu, Yan Cheng, Junjie Xiong, Dikoko Lesego Patson, Shijiang Cao, Heming Zhao, and Yuan Qin. 2019. "An Efficient Agrobacterium Mediated Transformation of Pineapple with GFP-Tagged Protein Allows Easy, Non-Destructive Screening of Transgenic Pineapple Plants" Biomolecules 9, no. 10: 617. https://doi.org/10.3390/biom9100617