Phenolic Profiling and Biological Potential of Ficus curtipes Corner Leaves and Stem Bark: 5-Lipoxygenase Inhibition and Interference with NO Levels in LPS-Stimulated RAW 264.7 Macrophages

 ,
,  , , , , and
, , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. General Chemicals and Standards
2.2. Plant Material
2.3. Extraction
2.4. Phenolic Profile Characterization
2.4.1. HPLC-DAD-ESI (Ion Trap)/MSn Qualitative Analysis
2.4.2. UPLC-ESI-QTOF-MS Qualitative Analysis
2.4.3. HPLC-DAD Quantitative Analysis
2.5. 5-LOX Inhibition
2.6. RAW 264.7 Macrophages
2.6.1. Cell Culture
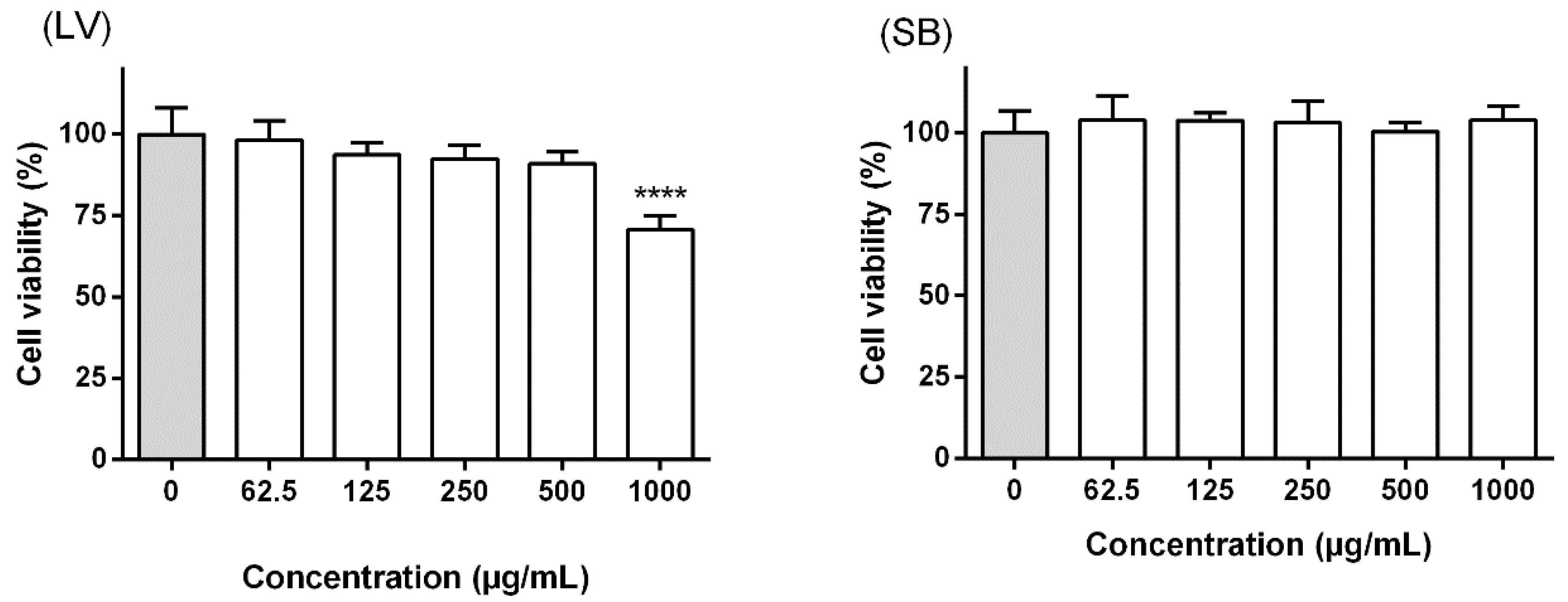
2.6.2. Cell Viability
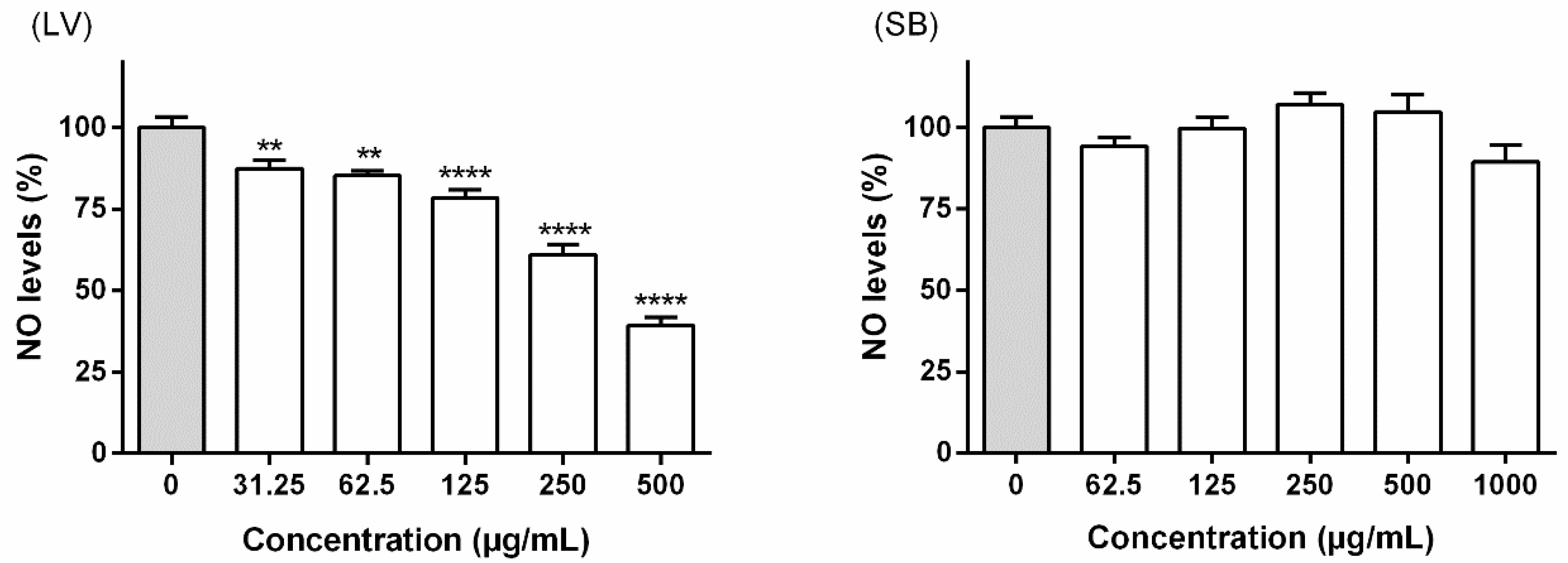
2.6.3. Determination of NO Levels in Culture Medium
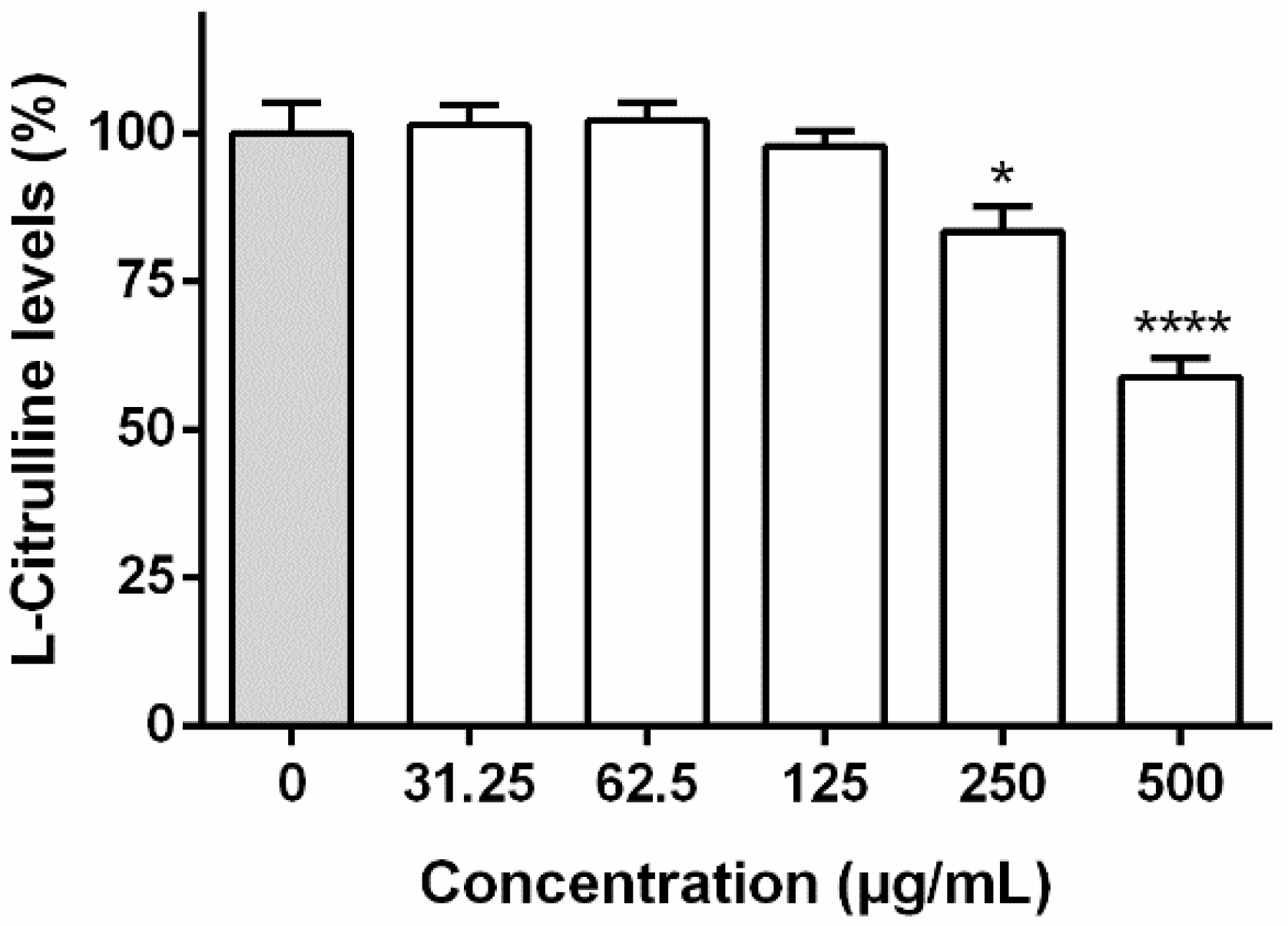
2.6.4. Determination of l-Citrulline Levels in Culture Medium
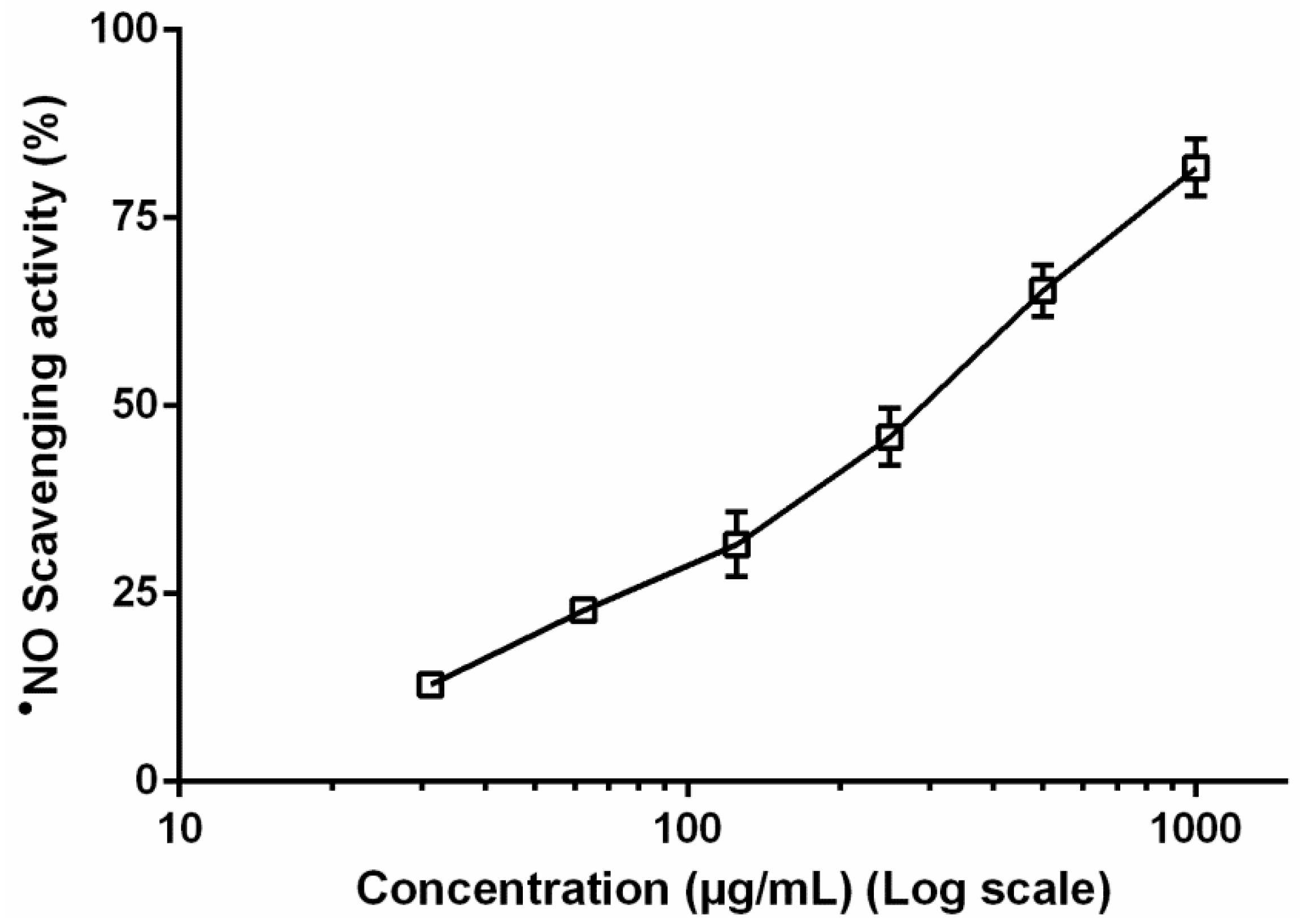
2.7. Determination of •NO Levels in Non-Cellular System
2.8. Statistical Analysis
3. Results
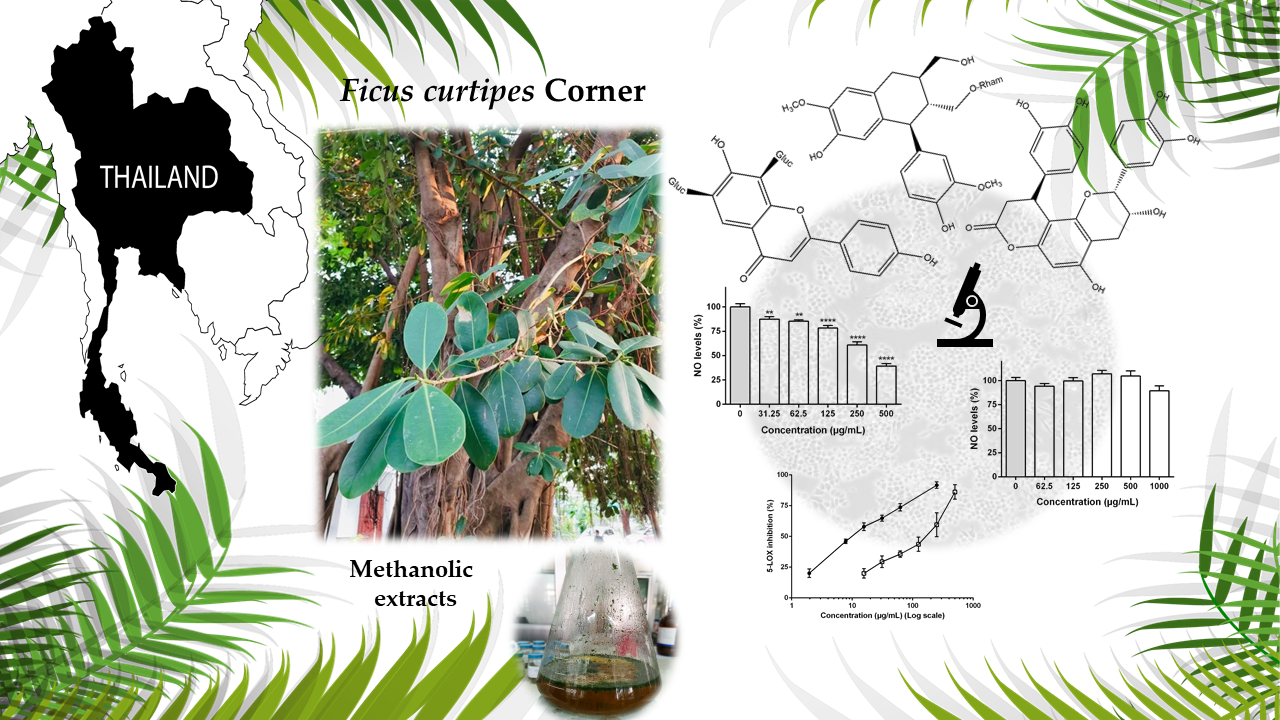
3.1. Characterization of the Phenolic Profile
3.1.1. HPLC-DAD-ESI (Ion Trap)/MSn Qualitative Analysis
3.1.2. HPLC-DAD Quantitative Analysis
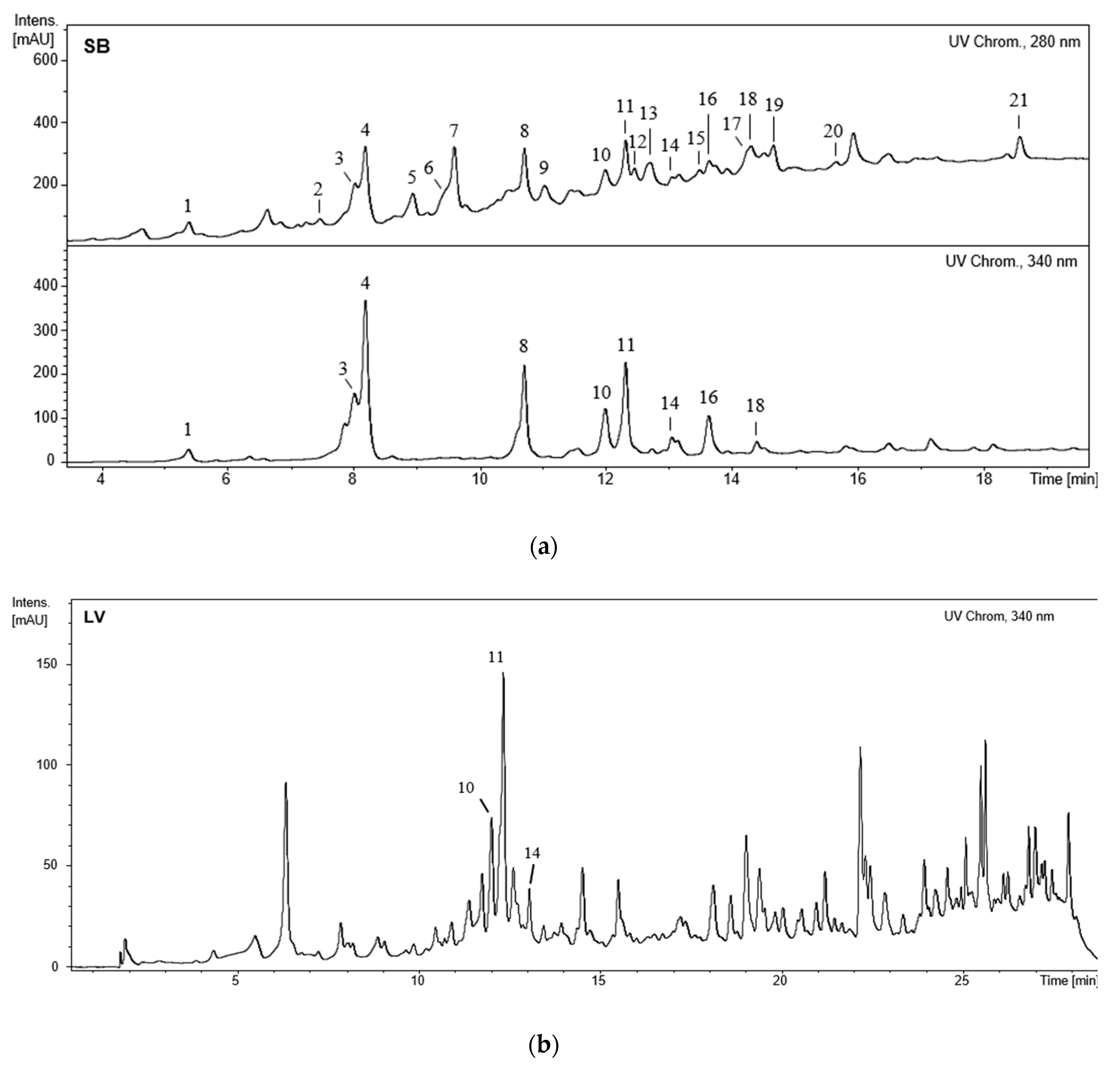
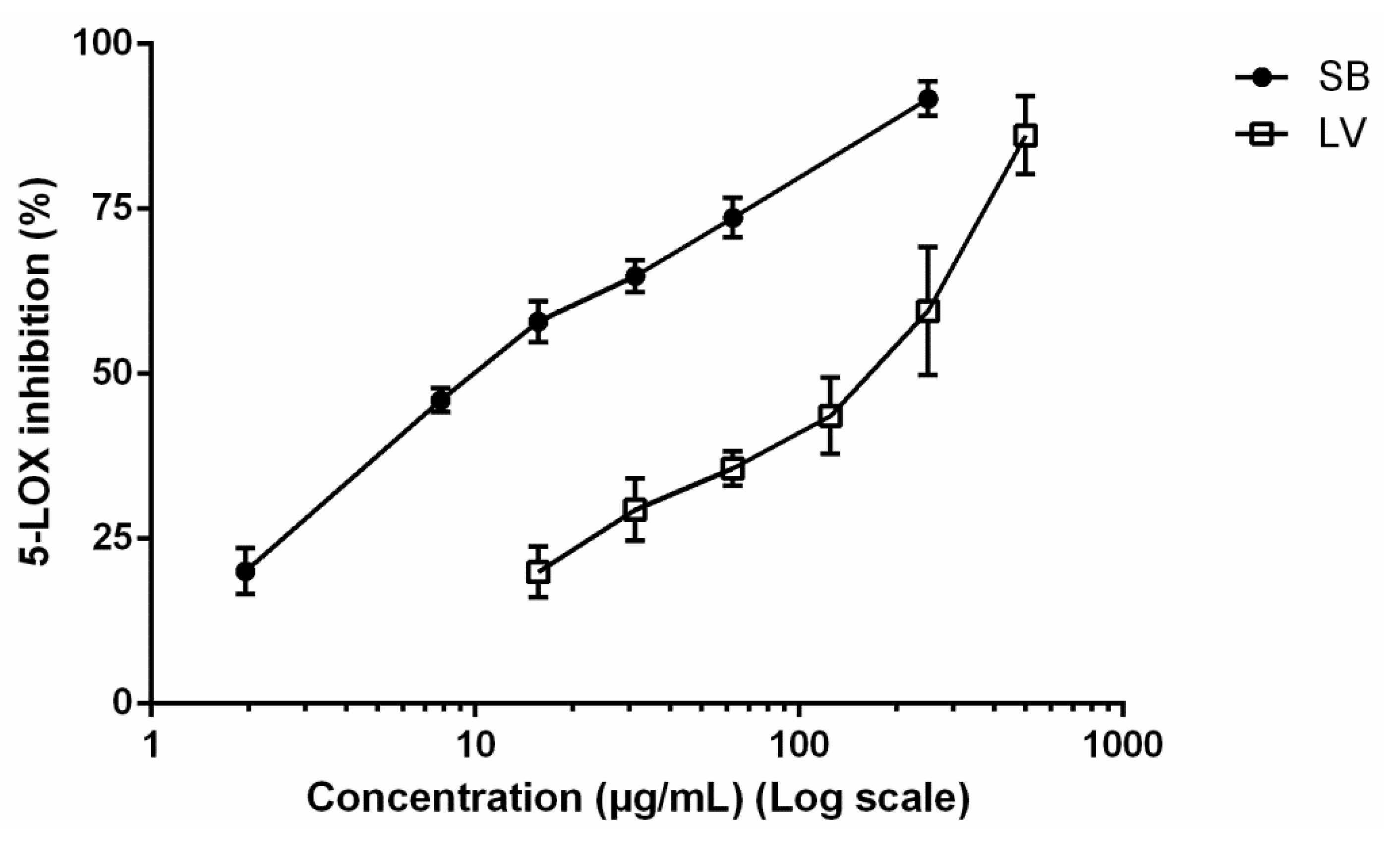
3.2. 5-LOX Inhibition
3.3. Effect on RAW 264.7 Macrophages
3.3.1. Interference with NO Levels
3.3.2. Interference with l-Citrulline Levels
3.4. Nitric Oxide Radical Levels in Non-Cellular System
4. Discussion
4.1. Phenolic Profile Characterization
4.2. 5-LOX Inhibition
4.3. Effect on RAW 264.7 Macrophages and ●NO Scavenging Capacity
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Atanasov, A.G.; Waltenberger, B.; Pferschy-wenzig, E.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veeresham, C. Natural products derived from plants as a source of drugs. J. Adv. Pharm. Technol. Res. 2012, 3, 200–201. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, M.; Heinrich, M.; Edwards, S.; Moerman, D.E.; Leonti, M. Ethnopharmacological field studies: A critical assessment of their conceptual basis and methods. J. Ethnopharmacol. 2009, 124, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lansky, E.P.; Paavilainen, H.M. Figs: The genus Ficus, 1st ed.; CRC Press: Boca Raton, FL, USA, 2010; p. 1. [Google Scholar]
- Philip, E.; Paavilainen, H.M.; Pawlus, A.D.; Newman, R.A. Ficus spp. (fig): Ethnobotany and potential as anticancer and anti-inflammatory agents. J. Ethnopharmacol. 2008, 119, 195–213. [Google Scholar]
- Park, S.; Han, J.; Im, K.; Whang, W.K.; Min, H. Antioxidative and anti-inflammatory activities of an ethanol extract from fig (Ficus carica) branches. Food Technol. Biotechnol. 2013, 22, 1071–1075. [Google Scholar] [CrossRef]
- Ali, H.; Monga, J.; Gupta, L.; Singh, A.; Narayan, S.; Siddiqui, Z.A.; Trivedi, V.B.E.N.; Ahi, J. Anti-inflammatory effects of hydro-methanolic extract of Ficus carica. Biomed. Pharmacol. J. 2009, 2, 129–132. [Google Scholar]
- Jung, H.W.; Son, H.Y.; Minh, C.V.; Kim, Y.H.; Park, Y.K. Methanol extract of Ficus leaf inhibits the production of nitric oxide and proinflammatory cytokines in LPS-stimulated microglia via the MAPK pathway. Phytother. Res. 2008, 1069, 1064–1069. [Google Scholar] [CrossRef]
- Gulecha, V.; Sivakumar, T.; Upaganlawar, A.; Mahajan, M.; Upasani, C. Screening of Ficus religiosa leaves fractions for analgesic and anti-inflammatory activities. Indian J. Pharmacol. 2011, 43, 662–666. [Google Scholar]
- Mandal, S.C.; Maity, T.K.; Das, J.; Saba, B.P.; Pal, M. Anti-inflammatory evaluation of Ficus racemosa Linn. leaf extract. J. Ethnopharmacol. 2000, 72, 87–92. [Google Scholar] [CrossRef]
- Gu, D.; Yang, D. Utilisation of chemical signals by inquiline wasps in entering their host figs. J. Insect Physiol. 2013, 59, 1065–1068. [Google Scholar] [CrossRef]
- Ferreres, F.; Grosso, C.; Gil-Izquierdo, A.; Fernandes, A.; Valentão, P.; Andrade, P.B. Comparing the phenolic profile of Pilocarpus pennatifolius Lem. by HPLC-DAD-ESI/MSn with respect to authentication and enzyme inhibition potential. Ind. Crops Prod. 2015, 77, 391–401. [Google Scholar] [CrossRef]
- García, C.J.; García-Villalba, R.; Gil, M.I.; Tomas-Barberan, F.A. LC-MS untargeted metabolomics to explain the signal metabolites inducing browning in fresh-cut lettuce. J. Agric. Food Chem. 2017, 65, 4526–4535. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Gomes, N.G.M.; Valentão, P.; Pereira, D.M.; Gil-izquierdo, A.; Araújo, L.; Silva, T.C.; Andrade, P.B. Leaves and stem bark from Allophylus africanus P. Beauv.: An approach to anti-inflammatory properties and characterization of their flavonoid profile. Food Chem. Toxicol. 2018, 118, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Duangsrisai, S.; Gomes, N.G.M.; Suksungworn, R.; Pereira, D.M.; Gil-Izquierdo, A.; Valentão, P.; Choowongkomon, K.; Andrade, P.B. Anti-inflammatory properties of the stem bark from the herbal drug Vitex peduncularis Wall. Ex Schauer and characterization of its polyphenolic profile. Food Chem. Toxicol. 2017, 106, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Moita, E.; Gil-Izquierdo, A.; Sousa, C.; Ferreres, F.; Silva, L.R.; Valentão, P.; Domínguez-Perles, R.; Baenas, N.; Andrade, P.B. Integrated analysis of COX-2 and iNOS derived inflammatory mediators in LPS-stimulated RAW macrophages pre-exposed to Echium plantagineum L. bee pollen extract. PLoS ONE 2013, 8, e59131. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Silva, M.B.; Andrade, P.B.; Seabra, R.M.; Ferreira, M.A. Approach to the study of C-glycosyl flavones by ion trap HPLC-PAD-ESI/MS/MS: Application to seeds of quince (Cydonia oblonga). Phytochem. Anal. 2003, 14, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Ferreres, F.; Gil-Izquierdo, A.; Andrade, P.B.; Valentão, P.; Tomás-Barberán, F.A. Characterization of C-glycosyl flavones O-glycosylated by liquid chromatography—Tandem mass spectrometry. J. Chromatogr. A 2007, 1161, 214–223. [Google Scholar] [CrossRef]
- Kim, H.J.; Woo, E.-R.; Park, H. A novel lignan and flavonoids from Polygonum aviculare. J. Nat. Prod. 1994, 57, 581–586. [Google Scholar] [CrossRef]
- Konai, N.; Raidandi, D.; Pizzi, A.; Meva, L. Characterization of Ficus sycomorus tannin using ATR-FT MIR, MALDI-TOF MS and 13 C NMR Methods. Eur. J. Wood Wood Prod. 2017, 75, 807–815. [Google Scholar] [CrossRef]
- Ao, C.; Higa, T.; Ming, H.; Ding, Y.; Tawata, S. Isolation and identification of antioxidant and hyaluronidase inhibitory compounds from Ficus microcarpa L. Fil. Bark. J. Enzym. Inhib. Med. Chem. 2010, 25, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Khattab, A.; Elhawary, S.Y.; Younis, I.; Elbishbishy, M. LC–MS/MS-Based chemometric analysis of phytochemical diversity in 13 Ficus spp. (Moraceae): Correlation to their in vitro antimicrobial and in silico quorum sensing inhibitory activities. Ind. Crops Prod. 2018, 126, 261–271. [Google Scholar]
- Oliveira, A.P.; Valentão, P.; Pereira, J.A.; Silva, B.M.; Tavares, F.; Andrade, P.B. Ficus carica L.: Metabolic and biological screening. Food Chem. Toxicol. 2009, 47, 2841–2846. [Google Scholar] [CrossRef] [PubMed]
- Mouho, D.G.; Oliveira, A.P.; Kodjo, C.G.; Valentão, P.; Gil-Izquierdo, A.; Andrade, P.B.; Ouattara, Z.A.; Bekro, Y.-A.; Ferreres, F. Chemical findings and in vitro biological studies to uphold the use of Ficus exasperata Vahl leaf and stem bark. Food Chem. Toxicol. 2018, 112, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Rasool, N.; Rizwan, K.; Zubair, M.; Riaz, M.; Zia-ul-haq, M.; Rana, U.A.; Nafady, A.; Jaafar, H.Z.E. Chemical composition and biological studies of Ficus benjamina. Chem. Cent. J. 2014, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Choo, C.Y.; Sulong, N.Y.; Man, F.; Wong, T.W. Vitexin and isovitexin from the leaves of Ficus deltoidea with in vivo α-glucosidase inhibition. J. Ethnopharmacol. 2012, 142, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Werz, O. Inhibition of 5-lipoxygenase product synthesis by natural compounds of plant origin. Planta Med. 2007, 73, 1331–1357. [Google Scholar] [CrossRef]
- Mashima, R.; Okuyama, T. The role of lipoxygenases in pathophysiology; new insights and future perspectives. Redox. Biol. 2015, 6, 297–310. [Google Scholar] [CrossRef] [Green Version]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The immunomodulatory and anti-inflammatory role of polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef]
- Zheng, M.; Yang, J.-H.; Li, Y.; Li, X.; Chang, H.; Son, J. Anti-Inflammatory activity of constituents isolated from Ulmus davidiana Var. Japonica. Biomol. Ther. 2010, 18, 321–328. [Google Scholar] [CrossRef]
- Burnett, B.P.; Jia, Q.; Zhao, Y.; Levy, R.M. A medicinal extract of Scutellaria baicalensis and Acacia catechu acts as a dual inhibitor of cyclooxygenase and 5-lipoxygenase to reduce inflammation. J. Med. Food 2007, 10, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Levy, R.M.; Khokhlov, A.; Kopenkin, S.; Bart, B.; Ermolova, T.; Kantemirova, R.; Mazurov, V.; Bell, M.; Caldron, P.; Pillai, L.; et al. Efficacy and safety of flavocoxid, a novel therapeutic, compared with naproxen: A randomized multicenter controlled trial in subjects with osteoarthritis of the knee. Adv. Ther. 2010, 27, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Schewe, T.; Ku, H. Biochemical and molecular actions of nutrients flavonoids of cocoa inhibit recombinant human 5-lipoxygenase. J. Nutr. 2002, 132, 1825–1829. [Google Scholar] [CrossRef] [PubMed]
- Schramm, D.D.; Wang, J.F.; Holt, R.R.; Ensunsa, J.L.; Gonsalves, J.L.; Lazarus, S.A.; Schmitz, H.H.; German, J.B.; Keen, C.L. Chocolate procyanidins decrease the leukotriene-prostacyclin ratio in humans and human aortic endothelial cells. Am. J. Clin. Nutr. 2001, 73, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Gawlik-dziki, U.; Dziki, D.; Habza-kowalska, E. Impact of interactions between ferulic and chlorogenic acids on enzymatic and non-enzymatic lipids oxidation: An example of bread enriched with green coffee flour. Appl. Sci. 2019, 9, 568. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Zayed, M.F.; Arabia, S. Lipoxygenase inhibitors flavonoids from Cyperus rotundus aerial parts. Rev. Bras. Farmacogn. 2018, 28, 320–324. [Google Scholar] [CrossRef]
- Pham, A.T.; Nguyen, C.; Malterud, K.E.; Diallo, D.; Wangensteen, H. Bioactive flavone-C-glycosides of the African medicinal plant Biophytum umbraculum. Molecules 2013, 18, 10312–10319. [Google Scholar] [CrossRef] [PubMed]
- Bosc, L.; Zeini, M.; Trav, P.G. Nitric oxide and cell viability in inflammatory cells: A role for NO in macrophage function and fate. Toxicology 2005, 208, 249–258. [Google Scholar] [CrossRef]
- Pereira, R.; Taveira, M.; Valentão, P.; Sousa, C.; Andrade, P. Fatty acids from edible sea hares: Anti-inflammatory capacity in LPS-stimulated RAW 264.7 cells involves iNOS modulation. RSC Adv. 2014, 5, 8981–8987. [Google Scholar] [CrossRef]
- Raso, G.M.; Meli, R.; Di Carlo, G.; Pacilio, M.; Di Carlo, R. Inhibition of inducible nitric oxide synthase and cyclooxygenase-2 expression by flavonoids in macrophage J774A.1. Life Sci. 2001, 68, 921–931. [Google Scholar] [CrossRef]
- Liang, Y.; Huang, Y.; Tsai, S.; Chen, C.; Lin, J. Suppression of inducible cyclooxygenase and inducible nitric oxide synthase by apigenin and related flavonoids in mouse macrophages. Carcinogenesis 1999, 20, 1945–1952. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Park, H.; Kim, H.P. Inhibition of nitric oxide production from lipopolysaccharide-treated RAW 264.7 cells by synthetic flavones: Structure-activity relationship and action mechanism. Arch. Pharm. Res. 2004, 27, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Morikawa, T.; Ando, S.; Toguchida, I.; Yoshikawa, M. Structural requirements of flavonoids for nitric oxide production inhibitory activity and mechanism of action. Bioorg. Med. Chem. 2003, 11, 1995–2000. [Google Scholar] [CrossRef]
- Gomes, A.; Fernandes, E.; Silva, A.M.S.; Santos, C.M.M.; Cavaleiro, A.S.; Pinto, D.C.G.A. 2- 2-Styrylchromones: Novel strong scavengers of reactive oxygen and nitrogen species. Bioorg. Med. Chem. 2007, 15, 6027–6036. [Google Scholar] [CrossRef] [PubMed]
- Awad, H.M.; Abd-alla, H.I.; Mahmoud, K.H.; El-toumy, S.A. In vitro anti-nitrosative, antioxidant, and cytotoxicity activities of plant flavonoids: A comparative study. Med. Chem. Res. 2014, 23, 3298–3307. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Rt (min) | Formula (M) | [M − H]− m/z | MS3[M − H]−, m/z (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| −60 | −90 | −120 | −162 | Agl2 + 113 | Agl2 + 83 | Agl2 + 71 | Agl2 + 41 | ||||
| 8 | 10.7 | C27H30O15 | 593.1508 | 533 (2) | 503 (30) | 473 (100) | 383 (45) | 353 (80) | |||
| 10 | 12.0 | C27H30O15 | 593.1506 | 503 (2) | 473 (30) | 431 (60) | 341 (15) | 311 (100) | |||
| 11 | 12.3 | C26H28O14 | 563.1396 | 503 (15) | 473 (80) | 443 (100) | 383 (65) | 353 (90) | |||
| 14 | 13.0 | C26H28O14 | 563.1402 | 503 (7) | 473 (85) | 443 (50) | 383 (75) | 353 (100) | |||
| 16 | 13.6 | C21H20O10 | 431.0980 | 341 (3) | 311 (100) | ||||||
| 18 | 14.3 | C21H20O10 | 431.0974 | 341 (30) | 311 (100) | ||||||
| Compounds | Rt (min) | Formula (M) | [M − H]− m/z | MS3[M − H]− m/z (%) |
|---|---|---|---|---|
| 2 | 7.4 | C15H14O6 | 289.0717 | 245(100), 205(50) |
| 5 | 8.9 | C30H26O12 | 577.1345 | 425(100), 407(80), 289(25) |
| 6 | 9.4 | C15H14O6 | 289.0714 | 245(100), 205(35) |
| 7 | 9.6 | C15H14O6 | 289.0714 | 245(100), 205(30) |
| 9 | 11.0 | C45H38O18 | 865.1988 | 695(100), 577(90), 425(60), 407(60), 287(30) |
| 12 | 12.4 | C39H32O15 | 739.1651 | 587(100), 569(25), 435(65), 417(40), 339(30), 289(20) |
| 13 | 12.6 | C39H32O15 | 739.1678 | 587(100), 569(30), 435(50), 417(10), 339(30), 289(15) |
| 15 | 13.4 | C24H20O9 | 451.1020 | 341(10) |
| 17 | 14.3 | C30H26O12 | 577.1347 | 425(100), 407(70), 289(30) |
| 20 | 15.6 | C24H20O9 | 451.1026 | 341(100) |
| 21 | 18.5 | C24H20O9 | 451.1024 | 341(10) |
| Standard | Regression Equation | Linearity Range (µg/mL) | LOD (µg/mL) | LOQ (µg/mL) | ||
|---|---|---|---|---|---|---|
| Slope (σ) | Intercept (b) | R2 (n = 3) | ||||
| 3-O-Caffeoylquinic acid | 44.661 | −17.577 | 0.999 | 2–32 | 0.439 | 1.330 |
| Catechin | 16.114 | 22.846 | 0.998 | 141–4.4 | 1.040 | 3.152 |
| 5-O-Caffeoylquinic acid | 157.450 | 100.020 | 0,999 | 7.5–120 | 2.125 | 6.439 |
| Epicatechin | 35.932 | 44.752 | 0.998 | 145–4.5 | 0.691 | 2.094 |
| Vicenin-2 | 55.527 | −0.958 | 0.999 | 3–48 | 0.606 | 1.837 |
| Epigallocatechin | 3.291 | −5.585 | 0.999 | 250–3.9 | 1.125 | 3.408 |
| Vitexin | 80.631 | 73.566 | 0.997 | 3–48 | 0.256 | 0.777 |
| Isovitexin | 98.771 | 27.029 | 0.998 | 1–16 | 0.013 | 0.040 |
| Compound | SB | LV | |
|---|---|---|---|
| 1 | 3-O-Caffeoylquinic acid | 33.79 ± 2.45 | Nd |
| 2 | Catechin | 10.17 ± 2.64 | Nd |
| 3 | Chlorogenic acid isomer | 33.78 ± 2.29 | Nd |
| 4 | 5-O-Caffeoylquinic acid | 201.23 ± 6.88 | Nd |
| 5 | Procyanidin type B | 52.17 ± 1.10 | Nd |
| 6 | Catechin/Epicatechin derivative | 129.38 ± 19.16 | Nd |
| 7 | Epicatechin | 377.51 ± 21.29 | Nd |
| 8 | Vicenin-2 | 218.23 ± 11.41 | Nd |
| 9 | Procyanidin type C | 27.44 ± 0.82 | Nd |
| 10 | Apigenin-7-O-Hex-6/8-C-Hex | 96.99 ± 6.59 | 152.17 ± 2.83 |
| 11 | Apigenin-6-C-Pt-8-C-Hex | 204.19 ± 9.89 | 381.20 ± 10.58 |
| 12 | Cinchonain type II | 280.94 ± 56.52 | Nd |
| 13 | Cinchonain type II | 727.65 ± 67.62 | Nd |
| 14 | Apigenin-6-C-Hex-8-C-Pent | 8.46 ± 1.13 | 78.27 ± 16.40 |
| 15 | Cinchonain type I | 293.45 ± 113.90 | Nd |
| 16 | Vitexin | 72.73 ± 4.29 | Nd |
| 17 | Procyanidin type B | 8.81 ± 1.69 | Nd |
| 18 | Isovitexin | 18.08 ± 2.96 | Nd |
| 19 | Aviculin | 1024.17 ± 81.73 | Nd |
| 20 | Cinchonain type I | 77.30 ± 11.35 | Nd |
| 21 | Cinchonain type I | 1478.00 ± 18.67 | Nd |
| Total | 5374.15 ± 436.61 | 611.5 ± 29.81 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrade, C.; Ferreres, F.; Gomes, N.G.M.; Duangsrisai, S.; Srisombat, N.; Vajrodaya, S.; Pereira, D.M.; Gil-Izquierdo, A.; Andrade, P.B.; Valentão, P. Phenolic Profiling and Biological Potential of Ficus curtipes Corner Leaves and Stem Bark: 5-Lipoxygenase Inhibition and Interference with NO Levels in LPS-Stimulated RAW 264.7 Macrophages. Biomolecules 2019, 9, 400. https://doi.org/10.3390/biom9090400
Andrade C, Ferreres F, Gomes NGM, Duangsrisai S, Srisombat N, Vajrodaya S, Pereira DM, Gil-Izquierdo A, Andrade PB, Valentão P. Phenolic Profiling and Biological Potential of Ficus curtipes Corner Leaves and Stem Bark: 5-Lipoxygenase Inhibition and Interference with NO Levels in LPS-Stimulated RAW 264.7 Macrophages. Biomolecules. 2019; 9(9):400. https://doi.org/10.3390/biom9090400
Chicago/Turabian StyleAndrade, Catarina, Federico Ferreres, Nelson G. M. Gomes, Sutsawat Duangsrisai, Nattawut Srisombat, Srunya Vajrodaya, David M. Pereira, Angel Gil-Izquierdo, Paula B. Andrade, and Patrícia Valentão. 2019. "Phenolic Profiling and Biological Potential of Ficus curtipes Corner Leaves and Stem Bark: 5-Lipoxygenase Inhibition and Interference with NO Levels in LPS-Stimulated RAW 264.7 Macrophages" Biomolecules 9, no. 9: 400. https://doi.org/10.3390/biom9090400