
Growth, Enzymatic, and Transcriptomic Analysis of xyr1 Deletion Reveals a Major Regulator of Plant Biomass-Degrading Enzymes in Trichoderma harzianum

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media, and General Growth Conditions
2.2. Construction of xyr1 Deletion T. harzianum Strains
2.3. Growth Profiling Analysis of T. harzianum Δxyr1 Mutants
2.4. Shake-Flask Cultures for Enzymatic and Transcriptomic Analyses
2.5. Enzyme Activity and PAGE Gel Analysis of the T. harzianum Δxyr1 Mutant
2.6. RNA Extraction, Sequencing, and Analysis of the T. harzianum Δxyr1 Mutant
3. Results
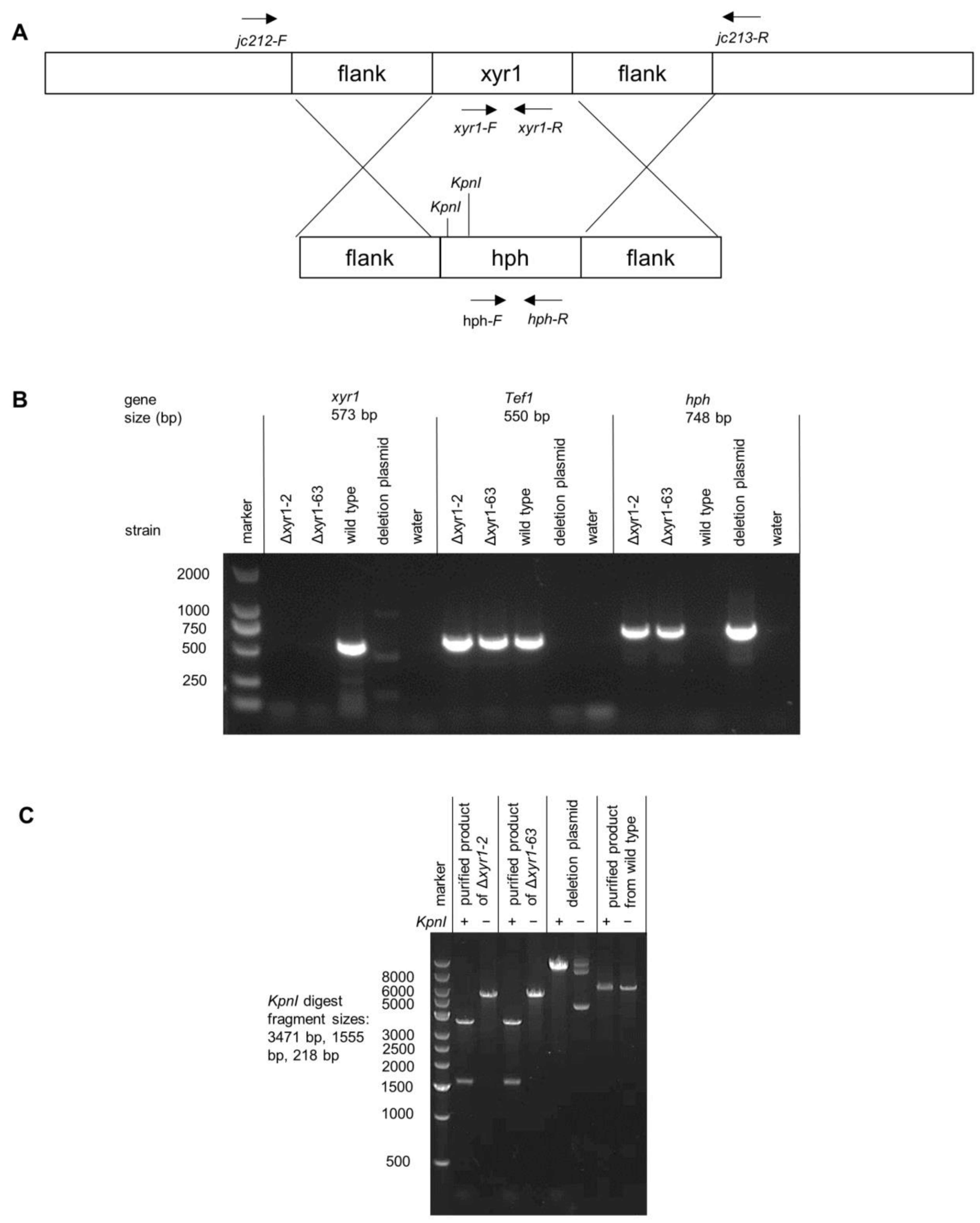
3.1. T. harzianum xyr1 Deletion Mutants Generated Using Hygromycin Resistance Cassette
3.2. The T. harzianum Δxyr1 Mutant Showed Reduced Growth on a Subset of Carbon Sources
3.3. Enzyme Activity and Protein Secretion Show Major Decreases in the T. harzianum Δxyr1 Mutant
3.4. Overview RNA-Seq Analysis of the T. harzianum Δxyr1 Mutant
3.5. RNA-Seq Analysis of the T. harzianum Δxyr1 Mutant Shows the Regulation of Xylanases as well as Cellulases
3.6. Expression Changes in the Δxyr1 Mutant beyond Plant Biomass-Degrading CAZymes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kubicek, C.P.; Steindorff, A.S.; Chenthamara, K.; Manganiello, G.; Henrissat, B.; Zhang, J.; Cai, F.; Kopchinskiy, A.G.; Kubicek, E.M.; Kuo, A.; et al. Evolution and comparative genomics of the most common Trichoderma species. BMC Genom. 2019, 20, 485. [Google Scholar] [CrossRef]
- Samuels, G.J. Trichoderma: Systematics, the sexual state, and ecology. Phytopathology 2006, 96, 195–206. [Google Scholar] [CrossRef]
- Benocci, T.; Aguilar-Pontes, M.V.; Zhou, M.; Seiboth, B.; de Vries, R.P. Regulators of plant biomass degradation in ascomycetous fungi. Biotechnol. Biofuels 2017, 10, 152. [Google Scholar] [CrossRef]
- Kowalczyk, J.E.; Daly, P. Transcriptional Regulation: How Saprobic Fungi Tune the Production of Plant Cell Wall Degrading Enzymes. In Encyclopedia of Mycology; Zaragoza, Ó., Casadevall, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 2, pp. 528–535. [Google Scholar]
- Xiao, Z.; Zhao, Q.; Li, W.; Gao, L.; Liu, G. Strain improvement of Trichoderma harzianum for enhanced biocontrol capacity: Strategies and prospects. Front. Microbiol. 2023, 14, 1146210. [Google Scholar] [CrossRef]
- Ferreira Filho, J.A.; Horta, M.A.C.; Beloti, L.L.; dos Santos, C.A.; de Souza, A.P. Carbohydrate-active enzymes in Trichoderma harzianum: A bioinformatic analysis bioprospecting for key enzymes for the biofuels industry. BMC Genom. 2017, 18, 779. [Google Scholar] [CrossRef] [PubMed]
- Amore, A.; Giacobbe, S.; Faraco, V. Regulation of Cellulase and Hemicellulase Gene Expression in Fungi. Curr. Genom. 2013, 14, 230–249. [Google Scholar] [CrossRef] [PubMed]
- de Souza, W.R.; Maitan-Alfenas, G.P.; de Gouvêa, P.F.; Brown, N.A.; Savoldi, M.; Battaglia, E.; Goldman, M.H.S.; de Vries, R.P.; Goldman, G.H. The influence of Aspergillus niger transcription factors AraR and XlnR in the gene expression during growth in D-xylose, L-arabinose and steam-exploded sugarcane bagasse. Fungal Genet. Biol. 2013, 60, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Stricker, A.R.; Grosstessner-Hain, K.; Würleitner, E.; Mach, R.L. Xyr1 (Xylanase Regulator 1) Regulates both the Hydrolytic Enzyme System and D-Xylose Metabolism in Hypocrea jecorina. Eukaryot. Cell 2006, 5, 2128–2137. [Google Scholar] [CrossRef]
- Sun, J.; Tian, C.; Diamond, S.; Glass, N.L. Deciphering Transcriptional Regulatory Mechanisms Associated with Hemicellulose Degradation in Neurospora crassa. Eukaryot. Cell 2012, 11, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Aro, N.; Saloheimo, A.; Ilmén, M.; Penttilä, M. ACEII, a Novel Transcriptional Activator Involved in Regulation of Cellulase and Xylanase Genes of Trichoderma reesei. J. Biol. Chem. 2001, 276, 24309–24314. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, Y.; Wu, C.; Liu, P.; Wang, W.; Wei, D. The transcription factor ACE3 controls cellulase activities and lactose metabolism via two additional regulators in the fungus Trichoderma reesei. J. Biol. Chem. 2019, 294, 18435–18450. [Google Scholar] [CrossRef]
- Coradetti, S.T.; Craig, J.P.; Xiong, Y.; Shock, T.; Tian, C.; Glass, N.L. Conserved and essential transcription factors for cellulase gene expression in ascomycete fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 7397–7402. [Google Scholar] [CrossRef]
- Derntl, C.; Rassinger, A.; Srebotnik, E.; Mach, R.L.; Mach-Aigner, A.R. Xpp1 regulates the expression of xylanases, but not of cellulases in Trichoderma reesei. Biotechnol. Biofuels 2015, 8, 112. [Google Scholar] [CrossRef]
- Benocci, T.; Aguilar-Pontes, M.V.; Kun, R.S.; Seiboth, B.; de Vries, R.P.; Daly, P. ARA1 regulates not only L-arabinose but also D-galactose catabolism in Trichoderma reesei. FEBS Lett. 2018, 592, 60–70. [Google Scholar] [CrossRef]
- Portnoy, T.; Margeot, A.; Linke, R.; Atanasova, L.; Fekete, E.; Sandor, E.; Hartl, L.; Karaffa, L.; Druzhinina, I.; Seiboth, B.; et al. The CRE1 carbon catabolite repressor of the fungus Trichoderma reesei: A master regulator of carbon assimilation. BMC Genom. 2011, 12, 269. [Google Scholar] [CrossRef]
- Aro, N.; Ilmén, M.; Saloheimo, A.; Penttilä, M. ACEI of Trichoderma reesei is a repressor of cellulase and xylanase expression. Appl Environ. Microbiol. 2003, 69, 56–65. [Google Scholar] [CrossRef]
- Cao, Y.; Zheng, F.; Wang, L.; Zhao, G.; Chen, G.; Zhang, W.; Liu, W. Rce1, a novel transcriptional repressor, regulates cellulase gene expression by antagonizing the transactivator Xyr1 in Trichoderma reesei. Mol. Microbiol. 2017, 105, 65–83. [Google Scholar] [CrossRef]
- Mello-de-Sousa, T.M.; Rassinger, A.; Pucher, M.E.; dos Santos Castro, L.; Persinoti, G.F.; Silva-Rocha, R.; Poças-Fonseca, M.J.; Mach, R.L.; Nascimento Silva, R.; Mach-Aigner, A.R. The impact of chromatin remodelling on cellulase expression in Trichoderma reesei. BMC Genom. 2015, 16, 588. [Google Scholar] [CrossRef]
- Benocci, T.; Aguilar Pontes, M.V.; Kun, R.S.; Lubbers, R.J.M.; Lail, K.; Wang, M.; Lipzen, A.; Ng, V.; Grigoriev, I.V.; Seiboth, B.; et al. Deletion of either the regulatory gene ara1 or metabolic gene xki1 in Trichoderma reesei leads to increased CAZyme gene expression on crude plant biomass. Biotechnol. Biofuels 2019, 12, 81. [Google Scholar] [CrossRef]
- Dos Santos, C.L.; Paula, R.G.; Antonieto, A.C.; Persinoti, G.F.; Silva-Rocha, R.; Silva, R.N. Understanding the Role of the Master Regulator XYR1 in Trichoderma reesei by Global Transcriptional Analysis. Front. Microbiol. 2016, 7, 175. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Chen, L.; Zhang, L.; Zou, G.; Liu, R.; Jiang, Y.; Zhou, Z. RNA Sequencing Reveals Xyr1 as a Transcription Factor Regulating Gene Expression beyond Carbohydrate Metabolism. BioMed Res. Int. 2016, 2016, 4841756. [Google Scholar] [CrossRef]
- Xia, Y.; Wang, J.; Guo, C.; Xu, H.; Wang, W.; Yang, M.; Shen, Q.; Zhang, R.; Miao, Y. Exploring the multi-level regulation of lignocellulases in the filamentous fungus Trichoderma guizhouense NJAU4742 from an omics perspective. Microb. Cell Fact. 2022, 21, 144. [Google Scholar] [CrossRef]
- Reithner, B.; Mach-Aigner, A.R.; Herrera-Estrella, A.; Mach, R.L. Trichoderma atroviride Transcriptional Regulator Xyr1 Supports the Induction of Systemic Resistance in Plants. Appl. Environ. Microbiol. 2014, 80, 5274–5281. [Google Scholar] [CrossRef]
- da Silva Delabona, P.; Rodrigues, G.N.; Zubieta, M.P.; Ramoni, J.; Codima, C.A.; Lima, D.J.; Farinas, C.S.; da Cruz Pradella, J.G.; Seiboth, B. The relation between xyr1 overexpression in Trichoderma harzianum and sugarcane bagasse saccharification performance. J. Biotechnol. 2017, 246, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Rosolen, R.R.; Aono, A.H.; Almeida, D.A.; Ferreira Filho, J.A.; Horta, M.A.C.; De Souza, A.P. Network Analysis Reveals Different Cellulose Degradation Strategies Across Trichoderma harzianum Strains Associated with XYR1 and CRE1. Front. Genet. 2022, 13, 807243. [Google Scholar] [CrossRef] [PubMed]
- Penttilä, M.; Nevalainen, H.; Rättö, M.; Salminen, E.; Knowles, J. A versatile transformation system for the cellulolytic filamentous fungus Trichoderma reesei. Gene 1987, 61, 155–164. [Google Scholar] [CrossRef]
- Cai, F.; Kubicek, C.P.; Druzhinina, I.S. Genetic Transformation of Trichoderma spp. In Biofuels and Biodiesel; Basu, C., Ed.; Springer: New York, NY, USA, 2021; pp. 171–185. [Google Scholar]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Chenthamara, K.; Zhang, J.; Atanasova, L.; Yang, D.; Miao, Y.; Rahimi, M.J.; Grujic, M.; Cai, F.; Pourmehdi, S.; et al. Massive lateral transfer of genes encoding plant cell wall-degrading enzymes to the mycoparasitic fungus Trichoderma from its plant-associated hosts. PLoS Genet. 2018, 14, e1007322. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-Seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 18. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, H.; Liu, Y.; Liu, Y.X.; Huang, L. EVenn: Easy to create repeatable and editable Venn diagrams and Venn networks online. J. Genet. Genom. 2021, 48, 863–866. [Google Scholar] [CrossRef]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Cock, P.J.; Chilton, J.M.; Grüning, B.; Johnson, J.E.; Soranzo, N. NCBI BLAST+ integrated into Galaxy. Gigascience 2015, 4, 39. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef]
- Klaubauf, S.; Narang, H.M.; Post, H.; Zhou, M.; Brunner, K.; Mach-Aigner, A.R.; Mach, R.L.; Heck, A.J.R.; Altelaar, A.F.M.; de Vries, R.P. Similar is not the same: Differences in the function of the (hemi-)cellulolytic regulator XlnR (Xlr1/Xyr1) in filamentous fungi. Fungal Genet. Biol. 2014, 72, 73–81. [Google Scholar] [CrossRef]
- Melo-Silveira, R.F.; Fidelis, G.P.; Costa, M.S.; Telles, C.B.; Dantas-Santos, N.; de Oliveira Elias, S.; Ribeiro, V.B.; Barth, A.L.; Macedo, A.J.; Leite, E.L.; et al. In vitro antioxidant, anticoagulant and antimicrobial activity and in inhibition of cancer cell proliferation by xylan extracted from corn cobs. Int. J. Mol. Sci. 2012, 13, 409–426. [Google Scholar] [CrossRef]
- Kim, K.C.; Yoo, S.S.; Oh, Y.A.; Kim, S.J. Isolation and characteristics of Trichoderma harzianum FJ1 producing cellulases and xylanase. J. Microbiol. Biotechnol. 2003, 13, 1–8. [Google Scholar]
- Lee, H.; Lee, Y.M.; Heo, Y.M.; Lee, A.; Hong, J.-H.; Jang, S.; Min, M.; Lee, J.; Kim, J.; Kim, G.-H.; et al. Optimization of Endoglucanase Production by Trichoderma harzianum KUC1716 and Enzymatic Hydrolysis of Lignocellulosic Biomass. BioResources 2015, 10, 7466–7476. [Google Scholar] [CrossRef]
- dos Santos Gomes, A.C.; Falkoski, D.; Battaglia, E.; Peng, M.; Nicolau de Almeida, M.; Coconi Linares, N.; Meijnen, J.-P.; Visser, J.; de Vries, R.P. Myceliophthora thermophila Xyr1 is predominantly involved in xylan degradation and xylose catabolism. Biotechnol. Biofuels 2019, 12, 220. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Zhao, Y.; Chen, S.; Wen, X.; Anjago, W.M.; Tian, T.; Chen, Y.; Zhang, J.; Deng, S.; Jiu, M.; et al. Growth, Enzymatic, and Transcriptomic Analysis of xyr1 Deletion Reveals a Major Regulator of Plant Biomass-Degrading Enzymes in Trichoderma harzianum. Biomolecules 2024, 14, 148. https://doi.org/10.3390/biom14020148
Wang L, Zhao Y, Chen S, Wen X, Anjago WM, Tian T, Chen Y, Zhang J, Deng S, Jiu M, et al. Growth, Enzymatic, and Transcriptomic Analysis of xyr1 Deletion Reveals a Major Regulator of Plant Biomass-Degrading Enzymes in Trichoderma harzianum. Biomolecules. 2024; 14(2):148. https://doi.org/10.3390/biom14020148
Chicago/Turabian StyleWang, Lunji, Yishen Zhao, Siqiao Chen, Xian Wen, Wilfred Mabeche Anjago, Tianchi Tian, Yajuan Chen, Jinfeng Zhang, Sheng Deng, Min Jiu, and et al. 2024. "Growth, Enzymatic, and Transcriptomic Analysis of xyr1 Deletion Reveals a Major Regulator of Plant Biomass-Degrading Enzymes in Trichoderma harzianum" Biomolecules 14, no. 2: 148. https://doi.org/10.3390/biom14020148