
Regulation of Angiogenesis by Non-Coding RNAs in Cancer

1
Department of Pathology, The First Affiliated Hospital of Soochow University, Suzhou 215006, China
2
State Key Laboratory of Radiation Medicine and Protection, School of Radiation Medicine and Protection, School for Radiological and Interdisciplinary Sciences (RAD-X), Collaborative Innovation Center of Radiation Medicine of Jiangsu Higher Education Institutions, Soochow University, Suzhou 215123, China
3
Department of Basic Medicine, Kangda College, Nanjing Medical University, Lianyungang 222000, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Biomolecules 2024, 14(1), 60; https://doi.org/10.3390/biom14010060
Submission received: 31 October 2023
/
Revised: 19 December 2023
/
Accepted: 22 December 2023
/
Published: 2 January 2024
(This article belongs to the Special Issue Non-coding RNAs and Circular RNAs in Tumor Angiogenesis)
Abstract
:Non-coding RNAs, including microRNAs, long non-coding RNAs, and circular RNAs, have been identified as crucial regulators of various biological processes through epigenetic regulation, transcriptional regulation, and post-transcriptional regulation. Growing evidence suggests that dysregulation and activation of non-coding RNAs are closely associated with tumor angiogenesis, a process essential for tumor growth and metastasis and a major contributor to cancer-related mortality. Therefore, understanding the molecular mechanisms underlying tumor angiogenesis is of utmost importance. Numerous studies have documented the involvement of different types of non-coding RNAs in the regulation of angiogenesis. This review provides an overview of how non-coding RNAs regulate tumor angiogenesis. Additionally, we discuss emerging strategies that exploit non-coding RNAs for anti-angiogenic therapy in cancer treatment. Ultimately, this review underscores the crucial role played by non-coding RNAs in tumor angiogenesis and highlights their potential as therapeutic targets for anti-angiogenic interventions against cancer.
1. Introduction
Non-coding RNA (ncRNA) refers to a class of RNA molecules that are transcribed from the genome but do not have the ability to code for proteins [1,2]. They can be classified into two main types: housekeeping ncRNA and regulatory ncRNA [3]. Regulatory ncRNAs can be further categorized as microRNA (miRNA), small interfering RNA (siRNA), piwi-interacting RNA (piRNA), long non-coding RNA (lncRNA), and circular RNA (circRNA) [3]. In recent years, there has been extensive research on regulatory ncRNAs, especially in the field of cancer research. These regulatory ncRNAs play important biological functions through epigenetic regulation, transcriptional regulation, and post-transcriptional regulation.
Tumor angiogenesis refers to the process of new blood vessel formation during the development of malignant tumors, which provides nutrients and oxygen to tumor cells [4,5]. Moreover, tumor angiogenesis offers a pathway for tumor metastasis, which is the leading cause of death in cancer patients [6,7]. Sustaining angiogenesis is a significant hallmark of cancer [8]. When vascular support is lacking, tumors may become necrotic or even apoptotic [9]. Therefore, targeting angiogenesis is a promising strategy for cancer treatment [10].
In recent years, there has been growing evidence suggesting that miRNA, lncRNA, and circRNA play a significant role in the regulation of tumor angiogenesis. It has become necessary to categorize and summarize the molecular mechanisms of various ncRNAs in tumor angiogenesis. This article aims to summarize the functions of various ncRNAs during tumor angiogenesis and discuss their potential implications for tumor diagnosis and treatment.
2. The Characteristics of Tumor Angiogenesis
Angiogenesis is a sequential, multi-step process that includes the destruction of the extracellular matrix, the budding and elongation of endothelial cells, the migration and proliferation of endothelial cells, and the formation and maturation of tubes [11]. A variety of cell types, including tumor cells, endothelial cells, immune cells, and fibroblasts are involved in tumor angiogenesis, which correlates with the complexity of the tumor microenvironment (TME) [12]. There is a diverse group of mediators secreted from these cells including growth factors, matrix-degrading enzymes, cytokines, bioactive lipids, and a variety of small molecules in TEM [13]. Among these mediators, vascular endothelial growth (VEGF) factors are thought to play a crucial role in regulating tumor angiogenesis. It can activate intracellular signaling pathways by binding to the corresponding receptors on endothelial cell membranes, ultimately forming blood vessels [14]. Hypoxia is a major feature of the tumor microenvironment, leading to the activation of the hypoxia-inducible factor-1 (HIF-1) transcription factor in tumor cells, which promotes the expression of VEGF [15,16]. HIF-1 complex activity can also be influenced by inflammation and cellular stress among other factors [17]. Fibroblasts accumulate in the early stages of tumor tissue formation and participate in regulating angiogenesis by secreting plasminogen activators (PAs) [18]. Immune cells, such as tumor-associated macrophages, secrete various growth factors and chemical mediators like VEGF, fibroblast growth factor-2, and angiogenesis-modulating enzymes. These substances directly or indirectly affect the process of angiogenesis [19]. In addition, several signaling pathways, such as transforming growth factor-beta (TGF-β) and signal transducer and activator of transcription (STAT), are involved in regulating the expression of these mediators [20,21]. The above features contribute to the complex nature of tumor angiogenesis. Additionally, these newly formed blood vessels often exhibit irregular, incomplete, and highly permeable characteristics, which promote cancer cell growth and metastasis [5,22]. Taken together, tumor angiogenesis is a complex process characterized by multi-step composition, the involvement of multiple cell types, and multi-factorial regulation.
3. miRNA and Tumor Angiogenesis
miRNAs are a type of small RNA molecules that exist naturally within organisms and are approximately 20–24 nucleotides long [23]. They are present in various organisms and play a significant role in regulating gene expression [24]. They achieve this regulation mainly by targeting specific recognition sites in the 3′ untranslated region (UTR), leading to mRNA degradation [25] (Figure 1). Dysregulation of miRNAs can lead to the development and progression of several diseases. In particular, aberrant miRNA expression has been linked to tumor angiogenesis [26,27]. Understanding the roles and behaviors of miRNAs offers valuable insights into the mechanisms of angiogenesis and potential targets for therapeutic interventions. The role of miRNAs in tumor angiogenesis is complex and multifaceted. We have classified the recent studies in Table 1 to highlight the functions of miRNAs in the following aspects.
miRNAs have a promoting effect on tumor angiogenesis. Since miRNAs are repressive for the regulation of target genes, angiogenesis inhibitors such as platelet response protein 1 (TSP-1) become prime targets when miRNAs promote tumor angiogenesis [50]. It has been demonstrated that miR-467 targets TSP-1, leading to increased inflammation resolution and angiogenesis [51]. Moreover, overexpression of miR-194 has been observed in advanced colorectal cancer, which binds to the 3′-UTR region of the TSP1 gene [52]. On the other hand, when miRNAs directly regulate angiogenic factors, they inhibit tumor angiogenesis. One crucial group of angiogenic factors is the VEGF family, and overactivation of these factors can result in abnormal angiogenesis [53]. For example, studies have demonstrated that miR-126 downregulates VEGF-A expression, inducing apoptosis and impeding tumor angiogenesis across various cancer types including breast cancer, lung cancer, oral cancer, and esophageal cancer [54,55,56,57].
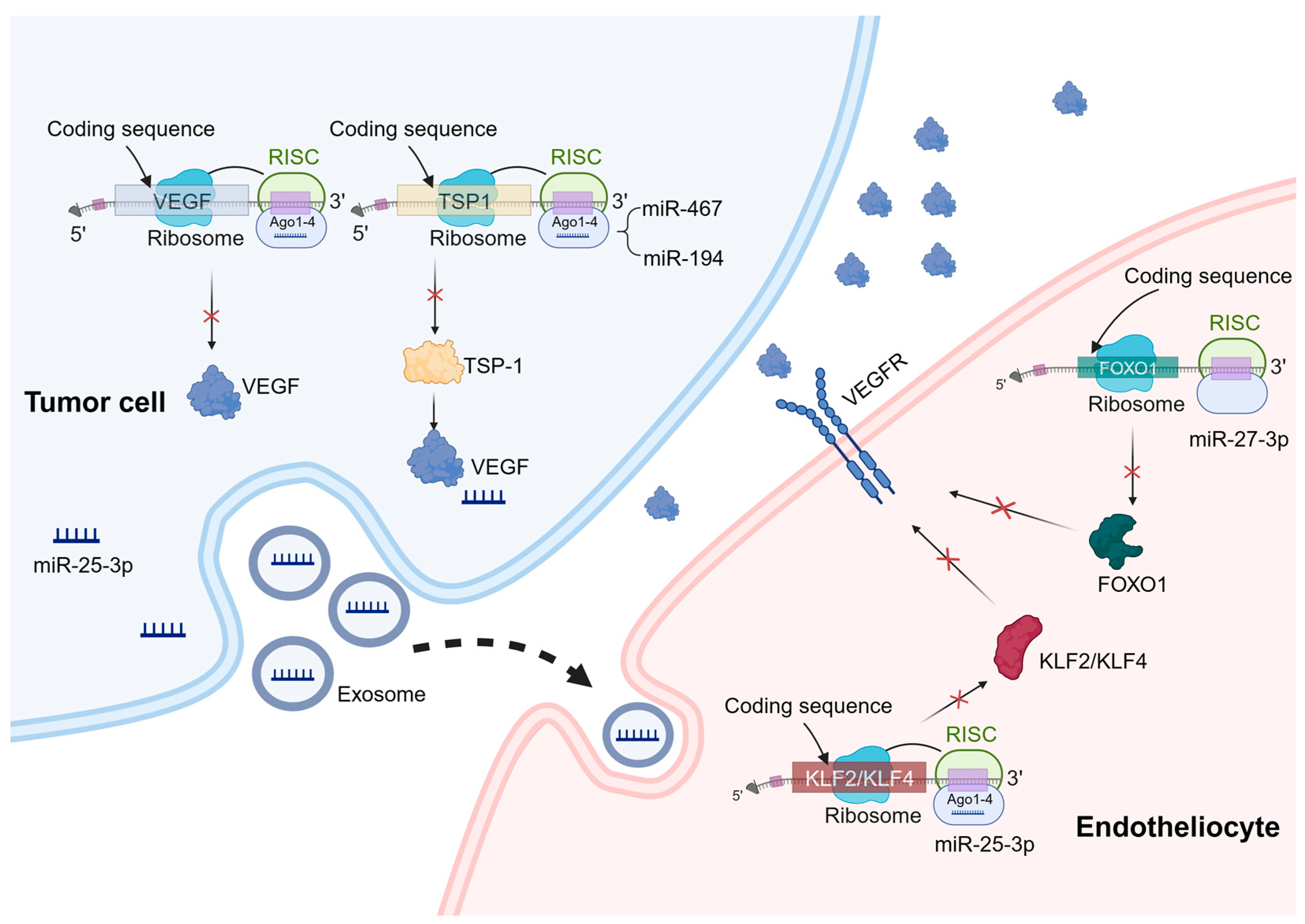
miRNAs also play important roles in vascular endothelial cells to regulate tumor angiogenesis. Specifically, miR-27b can suppress the activation of inflammatory pathways, consequently inhibiting intrinsic apoptosis. Mechanistically, miR-27b-3p targets the 3′-UTR region of FOXO1 mRNA, thereby downregulating its expression and subsequently attenuating the activation of the AKT/FOXO1 pathway [58].
In addition to miRNAs present within endothelial cells, extracellular vesicle miRNAs (exosomal miRNAs) have a notable impact on angiogenesis. For instance, recent studies have revealed that exosomal transfer of miR-25-3p from colorectal cancer (CRC) cells to endothelial cells promotes CRC metastasis. Mechanistically, this exosomal miRNA targets and silences KLF2 and KLF4, ultimately regulating gene expression associated with VEGFR2 and ZO-1 within endothelial cells. Consequently, this process promotes vascular permeability and neovascularization [59]. Importantly, extracellular vesicle miRNAs can also exert their influence on tumor angiogenesis by modulating immune cells. Research conducted by Zhao et al. demonstrates that exosomal miR-934 derived from CRC cells induces M2 macrophage polarization through downregulation of PTEN expression and activation of the PI3K/AKT signaling pathway. As a result, M2 macrophages produce various growth factors and cytokines like CXCL13 and CXCR5 that play crucial roles in regulating tumor growth, facilitating migration, and promoting angiogenesis [60]. In vivo, exosomes derived from stem cells of human deciduous exfoliated teeth significantly reduce the micro-vascular formation of tumors generated from xenografted oral squamous cell carcinoma cells via the transfer of miR-100-5p and miR-1246 [61].
4. lncRNA and Tumor Angiogenesis
lncRNAs, a group of RNA molecules longer than 200 nucleotides with a structure similar to mRNA, have been shown to possess more versatile mechanisms for regulating gene expression compared to miRNAs [62,63,64]. In recent years, lncRNAs have been implicated in various biological processes, including tumor proliferation, migration, invasion, and angiogenesis [65]. Here, we aimed to present an overview of the molecular mechanisms by which lncRNAs function as miRNA sponges, protein scaffolds, and coding peptides in the context of tumor angiogenesis (Figure 2). Table 2 categorizes recent studies on lncRNAs’ involvement in tumor angiogenesis.
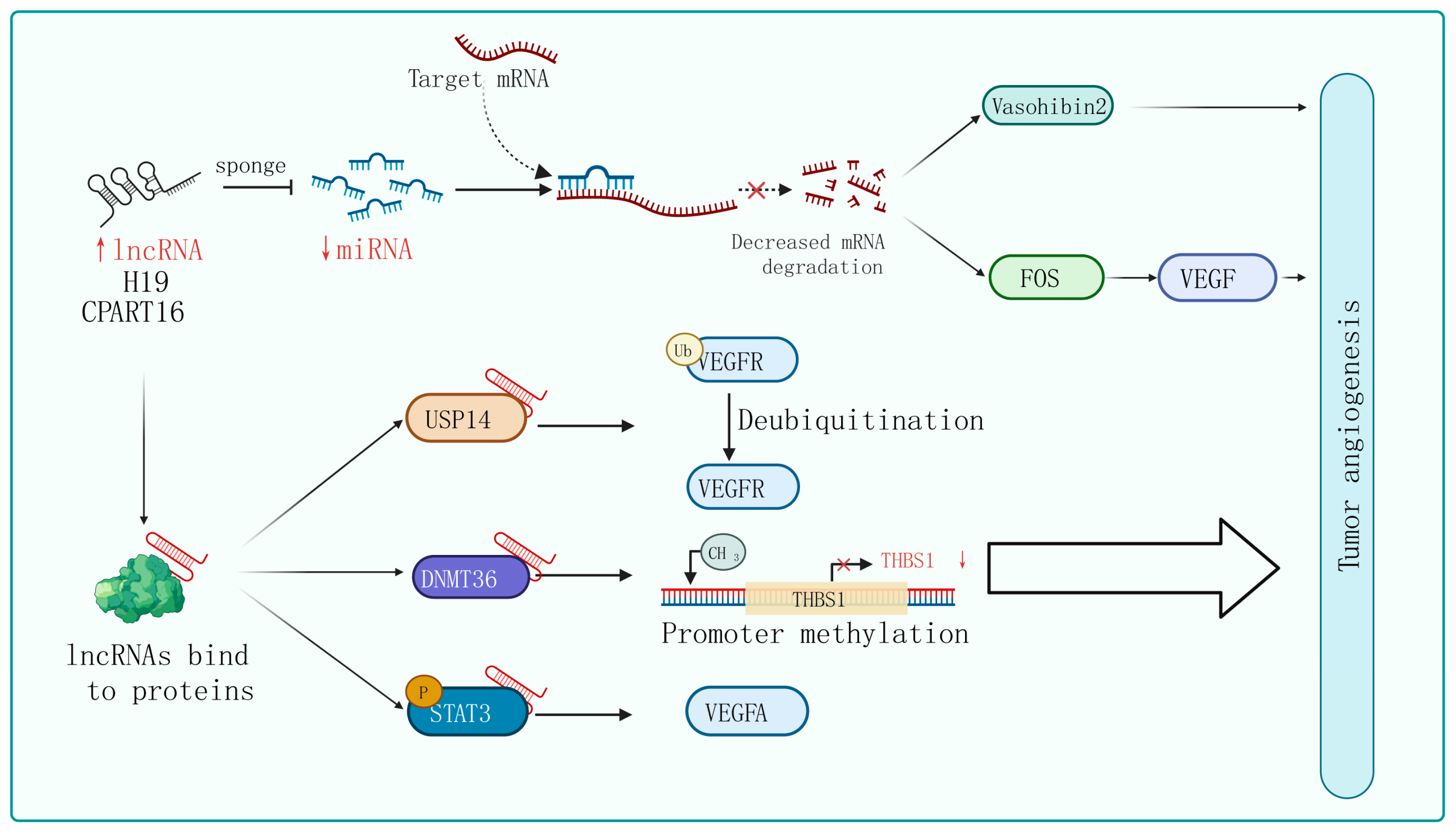
The phenomenon of lncRNAs acting as sponges that absorb miRNAs to promote target gene expression is universal in tumor angiogenesis. For instance, lncRNA H19 is upregulated in glioblastoma and plays an important role in driving angiogenesis by sequestering miR-29a and increasing the expression of vasohibin 2, an angiogenesis factor [87]. Similarly, Zhang et al. identified an lncRNA called CRART16 that is significantly overexpressed in gastric cancer tissue. They found that CRART16 acts as a sponge for miR-122-5p and upregulates the expression of the oncogene FOS. The overexpression of FOS leads to an increase in VEGF levels, promoting cancer cell growth and angiogenesis [73]. In addition, using the xenograft animal model, lncRNA ZNRD1-AS1 has been proven to inhibit the development of lung cancer by attenuating tumor angiogenesis [66].
The interactions between lncRNAs and various types of proteins confer a diversity of modalities for regulating tumor angiogenesis. Firstly, they can influence post-translational modifications of proteins. For instance, lncRNA PCAT6 binds to USP14 (a deubiquitinase) to induce the deubiquitination of VEGFR2, thereby increasing VEGFR2 expression levels and promoting angiogenesis in triple-negative breast cancer (TNBC) [68]. Secondly, lncRNAs can regulate gene transcription and translation. For example, in breast cancer cells, lncRNA RAB11B-AS1 promotes VEGFA and ANGPTL4 gene transcription by recruiting RNA Pol II to their promoter, enhancing tumor angiogenesis [82]. Additionally, lncRNA BZRAP1-AS1 indirectly enhances the methylation of the THBS1 promoter by increasing the stability of the DNMT3b protein, which inhibits the transcription of the anti-angiogenic gene THBS1 and promotes the angiogenesis process in tumors [88]. Furthermore, lncRNA HITT weakens the binding between YB-1 proteins and the 5′-UTR of HIF1α mRNA, impairing HIF1α translation. Finally, lncRNA can regulate signaling pathway activity. lncPVT1 interacts with phosphorylated STAT3 directly in the cell nucleus and activates the STAT3 signaling pathway, leading to increased expression of VEGFA [89]. Similarly, in non-small-cell lung cancer, researchers have found that lncRNA EPIC1 promotes tumor angiogenesis by activating the Ang2/Tie2 axis [90].
Although lncRNAs do not have typical protein-coding ability, some specific lncRNAs are capable of encoding small peptides due to their open reading frames (ORFs). Researchers found that LINC00908 can encode a 60-amino acid peptide in the TNBC. Interestingly, the peptide can directly interact with STAT3 and reduce the phosphorylation level of STAT3, thereby decreasing the expression of VEGF [91].
5. cirRNA and Tumor Angiogenesis
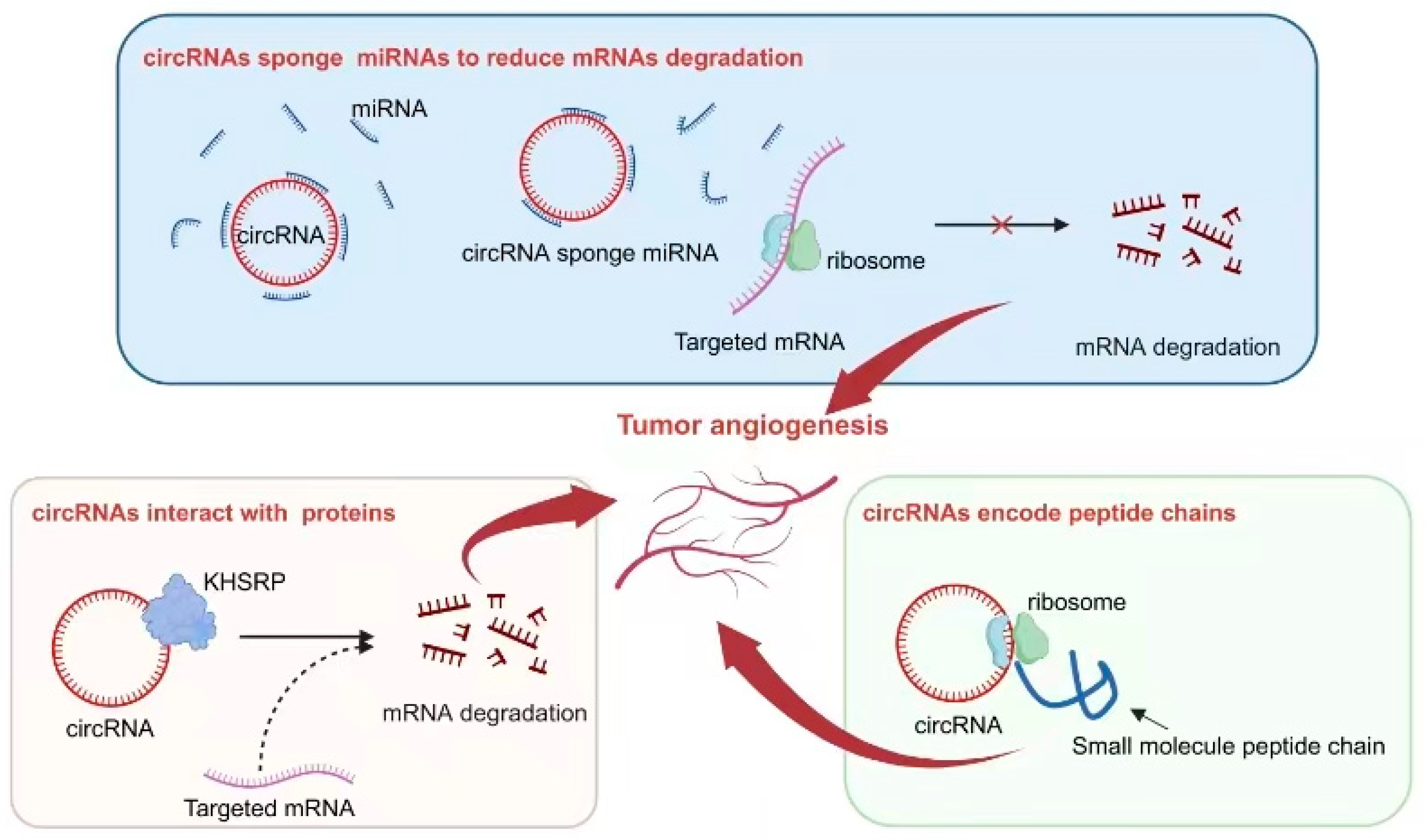
circRNAs are a unique form of non-coding RNA that possesses a closed circular structure, providing it with exceptional stability. Consequently, circRNAs are abundantly found in various cells and tissues and play significant roles in diverse biological processes, including tumor angiogenesis. Previous studies have elucidated several mechanisms by which circRNAs function: (1) They act as sponges, sequestering miRNAs and thereby inhibiting their regulatory effects on target genes. (2) They interact with proteins to modulate their localization, post-translational modifications, and stability. (3) Notably, circRNAs themselves can serve as potential sources for encoding small functional peptides that exhibit specific biological activities (Figure 3).
As a crucial regulator of tumor angiogenesis, VEGF serves as an excellent indicator for investigating the role of circRNAs in regulating this process. The abnormal activation of circRNAs often directly or indirectly influences the expression of VEGF, thereby exerting an impact on tumor angiogenesis. For instance, circ-29 enhances gastric cancer invasion and angiogenesis by sequestering miR-29a and consequently boosting the VEGF signaling pathway. The underlying mechanism involves the interaction between circ-29 and miR-29a, which reduces the latter’s ability to target the 3′-UTR region of VEGF mRNA and ultimately increases VEGF expression. Elevated levels of circ-29 contribute to the enhancement of invasive capabilities in gastric cancer cells and the promotion of neovascularization within the tumor microenvironment [92]. Furthermore, we have compiled and summarized the recent studies exploring how circRNA–miRNA interactions regulate various types of tumor angiogenesis in Table 3.
Interactions between circRNAs and various proteins play a crucial role in the process of tumor angiogenesis. One example is circLMP2A, which forms a complex with KHSRP, an RNA-splicing regulatory protein. This interaction inhibits the stability of VHL mRNA and relieves its inhibitory effect on HIF1α-induced VEGFA expression [93]. Another notable interaction involves circSHKBP1 and HSP90. In this case, their interaction inhibits the degradation of HSP90 by STUB1, the E3 ubiquitin ligase, resulting in enhanced VEGF expression in tumor tissue. As a result, progression and angiogenesis are enhanced in gastric cancer [94]. Table 3 provides further classification and documentation for similar mechanisms that have been reported.
Interestingly, some specific circRNAs have been found to possess the capability of encoding small peptides. This ability is attributed to the presence of an ORF within their sequence. In the context of tumor angiogenesis, this phenomenon has been extensively studied and reported. One such example is circ-0000437, which has been identified as encoding a small peptide comprising 47 amino acids. Through research efforts, it has been discovered that this particular peptide plays a significant role in inhibiting the interaction between ARNT and TACC3 proteins. Consequently, this inhibition leads to a reduction in VEGF expression, ultimately resulting in the suppression of tumor angiogenesis [95].
In addition to its effects on tumor cells, circRNA can regulate other cells in the tumor microenvironment through the secretion of exosomes by tumor cells. Exosomes are nanoscale lipid-enclosed structures with a diameter of 30–100 nm that are released by cells and contain proteins, nucleic acids, and other substances. This regulation contributes to the promotion of tumor angiogenesis. For instance, circHIPK3, which is highly expressed in breast cancer, can be carried by exosomes and enter human endothelial cells after being released into the extracellular space. circHIPK3 relieves the inhibitory effect of miR-124-3p on MTDH gene expression, ultimately promoting angiogenesis [96]. Furthermore, other cells from the tumor microenvironment can impact angiogenesis. Studies have demonstrated that M2 macrophages promote angiogenesis in cutaneous squamous cell carcinoma (cSCC). One potential mechanism involves the interaction between circ_TNFRSF21 and miR-3619-5p, leading to the increased expression of the ROCK2 gene and subsequent promotion of angiogenesis [97].

{kind=link}
{kind=link}
{kind=link}
Table 3.
Targets and dysregulation of circRNA associated with tumor angiogenesis.
| Basic Mechanisms | CircRNA | Molecular Target | Cancer Types | The Role in Tumor Angiogenesis | References |
|---|---|---|---|---|---|
| Sponging miRNAs (circRNA-miRNA-targeted gene) | CircRNA ARF1 | miR-342–3p/ISL2 | glioma cancer | promoter | [98] |
| Circ_0008344 | miR-638/SZRD1 | glioma cancer | promoter | [99] | |
| Circ-ATXN1 | miR-526b-3p/VEGFA | glioma cancer | promoter | [100] | |
| Circ3823 | miR-30c-5p/TCF7 | colorectal cancer | promoter | [101] | |
| CircTUBGCP4 | miR-146b-3p/PDK2 | colorectal cancer | promoter | [102] | |
| Circ_0030998 | miR-567/VEGFA | colorectal cancer | promoter | [103] | |
| Circ-ZNF609 | miR-145/STMN1 | nasopharyngeal carcinoma | promoter | [104] | |
| CircFIRRE | miR-486-3p and miR-1225-5p/LUZP1 | osteosarcoma | promoter | [105] | |
| CircFOXP1 | microRNA -127-5p/CDKN2AIP | osteosarcoma | promoter | [106] | |
| Circ_001587 | microRNA-223 | pancreatic cancer | inhibitor | [107] | |
| Circ_000684 | miR-145/KLF5 | pancreatic ductal adenocarcinoma cells | inhibitor | [108] | |
| CircRNF13 | miR-654-3p/PDK3 | pancreatic cancer | promoter | [109] | |
| CircKDM4B | miR-675/NEDD4L | breast cancer | inhibitor | [110] | |
| CircHIPK3 | miR-124-3p/MTDH | Breast cancer | promoter | [96] | |
| Circ_0001667 | miR-6838-5p/CXCL10 | breast cancer | promoter | [111] | |
| Circ29 | miR-29a/VEGF | Gastric cancer | promoter | [92] | |
| Circ_0001190 | miR-586/SOSTDC1 | Gastric Cancer | inhibitor | [112] | |
| Circ_0025033 | miR-370-3p/SLC1A5 | ovarian cancer | promoter | [113] | |
| CircNFIX | miR-518a-3p/TRIM44 | ovarian cancer | promoter | [114] | |
| CircATRNL1 | miR-378/SMAD4 | ovarian cancer | inhibitor | [115] | |
| Circ_0111738 | miR-1233-3p/HIF-1 | Multiple Myeloma | inhibitor | [116] | |
| Circ_0058058 | miR-338-3p/ATG14 | multiple myeloma | promoter | [117] | |
| Circ_TNFRSF21 | miR-3619-5p/ROCK | cutaneous squamous cell carcinoma | promoter | [97] | |
| Circfip1L1 | miR-125a-5p/VEGFA | Nasopharyngeal Carcinoma Cells | inhibitor | [118] | |
| CircHIPK2 | miR-1249-3p/VEGFA | non-small cell lung cancer | promoter | [119] | |
| Circ_0006988 | miR-491-5p/MAP3K3 | non-small cell lung cancer | promoter | [120] | |
| Circ-RAD23B | miR-142-3p/MAP4K3 | non-small cell lung cancer | promoter | [121] | |
| CircPRRC2A | miR-514a-5p and miR-6776-5p/TRPM3 | renal cell carcinoma | promoter | [122] | |
| CircAFAP1 | miR-374b-3p/VEGFA | renal cell carcinoma | promoter | [123] | |
| Circ_0015004 | miR-130a-3p/CEP55 | renal cell carcinoma | promoter | [117] | |
| Circ_0001955 | miR-646/FZD4 | Hepatocellular Carcinoma | promoter | [124] | |
| Circ_0000519 | miR-1296/E2F7 | hepatocellular carcinoma | promoter | [125] | |
| CircHDAC1_004 | miR-361-3p/NACC1 | Hepatocellular Carcinoma | promoter | [126] | |
| Circ_0000144 | miR-1178-3p/YWHAH | papillary thyroid cancer | promoter | [127] | |
| Circ_0011058 | miR-335-5p/YAP1 | papillary thyroid cancer | promoter | [128] | |
| CircSHKBP1 | miR-766-5p/HMGA2 | laryngeal squamous cell carcinoma | promoter | [129] | |
| Circ_0062019 | miR-1253/NRBP1 | prostate cancer | promoter | [130] | |
| CircSLC8A1 | miR-21 | prostate cancer | inhibitor | [131] | |
| Circ_0008726 | miR-206/HOXA13 | Esophageal squamous cell carcinoma | promoter | [132] | |
| Interacting (circRNA-Proteins) | circLMP2A | KHSRP | gastric carcinoma | promoter | [93] |
| circKIF18A | FOXC2 | Glioblastoma | promoter | [133] | |
| circSHKBP1 | HSP90 | gastric cancer | promoter | [94] | |
| circPOLR2A | UBE3C and PEBP1 | clear cell renal cell carcinoma | promoter | [134] | |
| circFNDC3B | FUS | Oral Squamous Cell Carcinoma | promoter | [135] | |
| circ_0004018 | FUS | hepatocellular carcinoma | inhibitor | [136] | |
| CircSMARCA5 | SRSF1 | Glioblastoma | promoter | [137] |
6. Other Types of ncRNA and Tumor Angiogenesis
Compared to miRNA, lncRNA, and circRNA, there have been fewer research reports on tsRNA, snoRNA, and piRNA in tumor angiogenesis in recent years. Here, we present a classification summary of these molecules. tRNA-derived small RNAs (tsRNAs) are a class of small non-coding RNAs derived from different regions of tRNA, including the 5′ end, 3′ end, and internal regions. tsRNAs can regulate gene expression and thereby influence important biological processes such as cell proliferation, differentiation, and apoptosis [138]. One study has demonstrated that tsRNA-26576 is highly expressed in breast tissues and functions in promoting tumor cell proliferation and inhibiting tumor cell apoptosis [139]. Additionally, 5′tiRNA-His-GTG has been found to be highly expressed in colorectal cancer tissues and plays a role in response to hypoxic stress by activating the HIF1α/vascular axis and promoting the Hippo pathway [140].
Small nucleolar RNAs (snoRNAs) are a diverse class of non-coding RNAs (ncRNAs) that range in length from approximately 60 to 300 nucleotides. These specific ncRNAs are primarily localized in the nucleolus and play important roles in cleaving and chemically modifying ribosomal RNAs (rRNAs) [141,142]. In recent years, there have been limited studies investigating the association between snoRNA and tumor angiogenesis. However, reports have suggested abnormal expression of snoRNAs in various types of tumors, indicating their potential correlation with tumorigenesis. For instance, SNORA42 has been demonstrated to act as an oncogene in lung cancer, hepatocellular carcinoma, and colorectal cancer [143,144,145]. Similarly, upregulation of SNORD17 has been observed in hepatocellular carcinoma tissues compared to normal liver tissues, implicated in driving cancer progression. Mechanistically, SNORD17 anchors nucleophosmin 1 (NPM1) and MYB binding protein 1a (MYBBP1A) simultaneously within the nucleolus, leading to decreased activation of p53 [146].
piRNA is a recently discovered class of small non-coding RNA that is found in both germline cells and somatic cells. These RNA molecules can form complexes with PIWI proteins and typically comprise 24-31 nucleotides in length [147]. In contrast to other RNA sequences, piRNAs deviate from canonical sequences by having a uridine at the 5′ end or an adenine at position ten. Additionally, they lack clear secondary structure motifs [148]. The function of piRNAs encompasses various biological processes such as transposon silencing, spermatogenesis, genome rearrangements, epigenetic regulation, and protein regulation [147]. However, recent reports suggest that abnormal expression of piRNAs may be associated with tumor development and angiogenesis [149]. For instance, studies have observed the upregulation of piRNA-823 in patients with multiple myeloma (MM) as well as MM cell lines. This upregulation positively correlates with clinical staging. In MM cells specifically, downregulating piRNA-823 has been shown to inhibit the secretion of VEGF, subsequently reducing angiogenic activity [150]. Furthermore, low expression levels of piRNA-2158 have been detected in breast cancer tumors. It functions by suppressing transcription through binding to the IL-11 promoter. Interestingly enough, it has been demonstrated that this specific piRNA inhibits angiogenesis in breast cancer, as well [151].
7. ncRNA-Targeting Therapeutics in Tumor Angiogenesis
Targeted ncRNA therapy holds great promise for the treatment of tumors. In recent decades, significant clinical investment has been made in RNA-based therapeutic modalities. Currently, there are various RNA-based therapeutic modalities available, such as antisense oligonucleotides (ASOs), small interfering RNAs (siRNAs), short hairpin RNAs (shRNAs), ASO anti-microRNAs (antimiRs), miRNA mimics, miRNA sponges, therapeutic circular RNAs (circRNAs), and CRISPR-Cas9-based gene editing. Among these modalities, ASOs and siRNAs are more widely used [152].
There are several advantages to using miRNA-based therapies. Firstly, miRNAs are naturally occurring molecules in human cells, in contrast to man-made chemotherapeutic compounds or ASOs. They possess all the mechanisms for processing and downstream target selection. Secondly, miRNAs act by targeting multiple genes in a pathway, thereby eliciting a broader but specific response [153,154,155].
One notable characteristic of circRNAs is their closed covalent structure, which makes them less susceptible to degradation compared to other RNA molecules. This property allows circRNAs to easily accumulate within various types of cells or tissues and serve as drugs [156]. Understanding the molecular mechanisms by which circRNAs function has led to the utilization of various chemical and enzymatic synthesis strategies for producing circRNA drugs in vitro [157]. In contrast to conventional protein- or peptide-based drugs, circRNAs exhibit a sub-stoichiometric mode of action, thus facilitating drug delivery with lower toxicity concerns. Moreover, they have favorable pharmacokinetic potential compared to traditional vaccines based on pathogens and DNA/proteins/peptides since they do not require nucleotide modification; yet, they can achieve strong drug efficacy through simple steps [158].
Based on the advantages of targeted ncRNA therapy and the understanding of the molecular mechanism of ncRNA, the first clinical trial of miRNA therapy for tumors was conducted in 2013 [159]. The drug MRX34 is a synthetic double-stranded mir-34a mimic, which can induce tumor cell apoptosis, inhibit tumor cell survival, and extend the survival time of mice. However, severe immune-mediated adverse events occurred in patients in the phase I clinical trial, leading to trial failure [160]. Nevertheless, this experiment proved that improving tumor-specific delivery systems can reduce the off-target toxicity of miRNA drugs. Subsequently, a synthetic mimic of miR-16 called TargomiR entered clinical trials but showed insignificant effects and mild adverse events [161]. However, this clinical trial indicated that the drug delivery system has a greater responsibility for inflammation toxicity. With continuous improvement in drug delivery systems and other conditions, an analog of mir-193a-3p called INT-IB3 is currently undergoing phase I clinical trials [162]. In addition, as of the date of this paper, no circRNA therapeutic candidate has entered clinical trials. However, there has been some progress in pre-clinical studies utilizing circRNA-based mechanisms and nanoparticle delivery systems for cancer treatment. For instance, researchers have discovered that circEHMT1 can effectively inhibit the migration and invasion of breast cancer cells. In a study conducted on mice, nanoparticles were employed to deliver a plasmid expressing circEHMT1, resulting in a significant reduction in lung metastasis of breast cancer cells [163]. Although ncRNA-based therapeutics are still mainly in the research and development stage, it is believed that they will have a wide range of applications in the near future.
8. Conclusions and Future Perspectives
This review mainly summarizes the functions of miRNAs, lncRNAs, and circRNAs in tumor angiogenesis and highlights their molecular mechanisms. Meanwhile, we have summarized the advantages of ncRNA-based therapy. It is well understood for the significant roles of miRNAs, lncRNAs, and cirRNAs in tumor angiogenesis.
We have conducted a classification analysis of miRNA, lncRNA, and circRNA based on their molecular mechanisms in tumor angiogenesis. It can be observed that they are interconnected and compete with each other. For example, lncRNAs and circRNAs can inhibit the function of miRNAs by sequestering them. With regard to the summarized molecular mechanisms in this paper, progress has been made in recent years, which has advanced our understanding of tumor angiogenesis to some extent. However, considering the dual complexity of tumor angiogenesis and the potential involvement of various molecular mechanisms, our current understanding of the role of ncRNAs in tumor angiogenesis may only scratch the surface.
During the literature review process, we also focused on exploring the clinical applications of ncRNAs. Unfortunately, there are currently no real applications of ncRNAs in clinical therapy. We speculate on several reasons for this. Firstly, although substantial achievements have been made in understanding molecular mechanisms in humans, there is still a lack of breakthroughs due to similarities or redundancies among many studies. This might be limited by current molecular biology techniques. Secondly, each study highlights its ncRNA as having noteworthy clinical implications for tumor angiogenesis; however, identifying which ones occupy central positions within regulatory networks and serve as viable drug targets likely requires deep collaboration among fields such as artificial intelligence, molecular mechanism research, and drug development to integrate complex regulatory networks. Thirdly, tumor angiogenesis is an ongoing process occurring within a complex tumor microenvironment, which further complicates the application of ncRNAs in clinical settings. Therefore, more advanced models need to be developed for research purposes. Finally, the optimization of drug delivery systems plays a crucial role in reducing patient immune responses and determining the success of ncRNA therapy. Fortunately, the field of nanomaterials has experienced rapid development in recent years. With the continuous development of multiple disciplines, it is foreseeable that there will be more clinical applications based on ncRNA therapy in the future.
Author Contributions
All authors contributed to the study’s conception and design. Z.S., W.L. and Z.L. performed the investigation and wrote the first draft of the manuscript. L.H. illustrated the figures. S.W. and L.G. reviewed and edited the draft. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (82203773, 81972174), the Natural Science Foundation of Jiangsu Province (BK20231233), Lianyungang Key R&D Program (Social Development) (SF2207), Suzhou Science and Technology Program-Applied and Fundamental Medical Research-Key Clinical Technology Research (SKY2023009).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No additional data.
Acknowledgments
Figures were created with BioRender (www.biorender.com) (accessed on 18 December 2023).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mattick, J.S.; Makunin, I.V. Non-coding RNA. Hum. Mol. Genet. 2006, 15, R17–R29. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and their Integrated Networks. J. Integr. Bioinform. 2019, 16, 20190027. [Google Scholar] [CrossRef] [PubMed]
- Voravud, N.; Charuruk, N. Tumor angiogenesis. J. Med. Assoc. Thai. 1999, 82, 394–404. [Google Scholar] [PubMed]
- Hida, K.; Maishi, N.; Torii, C.; Hida, Y. Tumor angiogenesis—Characteristics of tumor endothelial cells. Int. J. Clin. Oncol. 2016, 21, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.P.; Massague, J. Cancer metastasis: Building a framework. Cell 2006, 127, 679–695. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Yang, Z.; Xu, Y.; Chen, Y.; Yu, Q. Apoptosis, autophagy, necroptosis, and cancer metastasis. Mol. Cancer 2015, 14, 48. [Google Scholar] [CrossRef]
- Suresh, R.; Diaz, R.J. The remodelling of actin composition as a hallmark of cancer. Transl. Oncol. 2021, 14, 101051. [Google Scholar] [CrossRef]
- Tozer, G.M.; Kanthou, C.; Baguley, B.C. Disrupting tumour blood vessels. Nat. Rev. Cancer 2005, 5, 423–435. [Google Scholar] [CrossRef]
- Jayson, G.C.; Kerbel, R.; Ellis, L.M.; Harris, A.L. Antiangiogenic therapy in oncology: Current status and future directions. Lancet 2016, 388, 518–529. [Google Scholar] [CrossRef]
- Baru, O.; Nutu, A.; Braicu, C.; Cismaru, C.A.; Berindan-Neagoe, I.; Buduru, S.; Badea, M. Angiogenesis in Regenerative Dentistry: Are We Far Enough for Therapy? Int. J. Mol. Sci. 2021, 22, 929. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Coussens, L.M. Accessories to the crime: Functions of cells recruited to the tumor microenvironment. Cancer Cell 2012, 21, 309–322. [Google Scholar] [CrossRef]
- Kim, H.J.; Ji, Y.R.; Lee, Y.M. Crosstalk between angiogenesis and immune regulation in the tumor microenvironment. Arch. Pharm. Res. 2022, 45, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef] [PubMed]
- Arneth, B. Tumor Microenvironment. Medicina 2019, 56, 15. [Google Scholar] [CrossRef] [PubMed]
- Ke, Q.; Costa, M. Hypoxia-inducible factor-1 (HIF-1). Mol. Pharmacol. 2006, 70, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Balamurugan, K. HIF-1 at the crossroads of hypoxia, inflammation, and cancer. Int. J. Cancer 2016, 138, 1058–1066. [Google Scholar] [CrossRef]
- Nyberg, P.; Salo, T.; Kalluri, R. Tumor microenvironment and angiogenesis. Front. Biosci. 2008, 13, 6537–6553. [Google Scholar] [CrossRef]
- Ribatti, D.; Crivellato, E. Immune cells and angiogenesis. J. Cell Mol. Med. 2009, 13, 2822–2833. [Google Scholar] [CrossRef]
- Pardali, E.; ten Dijke, P. Transforming growth factor-beta signaling and tumor angiogenesis. Front. Biosci. 2009, 14, 4848–4861. [Google Scholar] [CrossRef]
- Haura, E.B.; Turkson, J.; Jove, R. Mechanisms of disease: Insights into the emerging role of signal transducers and activators of transcription in cancer. Nat. Clin. Pract. Oncol. 2005, 2, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Maishi, N.; Hida, K. Tumor endothelial cells accelerate tumor metastasis. Cancer Sci. 2017, 108, 1921–1926. [Google Scholar] [CrossRef] [PubMed]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Croce, C.M. MicroRNA dysregulation in cancer: Diagnostics, monitoring and therapeutics. A comprehensive review. EMBO Mol. Med. 2012, 4, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Zhao, Z.; Cai, Q.; Zhang, Y.; Zhang, P.; Shi, S.; Xie, H.; Peng, X.; Yin, W.; Tao, Y.; et al. miRNA-based biomarkers, therapies, and resistance in Cancer. Int. J. Biol. Sci. 2020, 16, 2628–2647. [Google Scholar] [CrossRef]
- Annese, T.; Tamma, R.; De Giorgis, M.; Ribatti, D. microRNAs Biogenesis, Functions and Role in Tumor Angiogenesis. Front. Oncol. 2020, 10, 581007. [Google Scholar] [CrossRef]
- Liao, Y.; Wang, C.; Yang, Z.; Liu, W.; Yuan, Y.; Li, K.; Zhang, Y.; Wang, Y.; Shi, Y.; Qiu, Y.; et al. Dysregulated Sp1/miR-130b-3p/HOXA5 axis contributes to tumor angiogenesis and progression of hepatocellular carcinoma. Theranostics 2020, 10, 5209–5224. [Google Scholar] [CrossRef]
- Li, H.; Liu, P.; Li, D.; Wang, Z.; Ding, Z.; Zhou, M.; Chen, X.; Miao, M.; Ding, J.; Lin, W.; et al. STAT3/miR-130b-3p/MBNL1 feedback loop regulated by mTORC1 signaling promotes angiogenesis and tumor growth. J. Exp. Clin. Cancer Res. 2022, 41, 297. [Google Scholar] [CrossRef]
- Qiu, S.; Xie, L.; Lu, C.; Gu, C.; Xia, Y.; Lv, J.; Xuan, Z.; Fang, L.; Yang, J.; Zhang, L.; et al. Gastric cancer-derived exosomal miR-519a-3p promotes liver metastasis by inducing intrahepatic M2-like macrophage-mediated angiogenesis. J. Exp. Clin. Cancer Res. 2022, 41, 296. [Google Scholar] [CrossRef]
- Chen, K.; Wang, Q.; Liu, X.; Wang, F.; Yang, Y.; Tian, X. Hypoxic pancreatic cancer derived exosomal miR-30b-5p promotes tumor angiogenesis by inhibiting GJA1 expression. Int. J. Biol. Sci. 2022, 18, 1220–1237. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.M.; Fu, Y.; Zeng, J.; Zhu, X.Y.; Gao, Y. Cancer-derived exosomal miR-197-3p confers angiogenesis via targeting TIMP2/3 in lung adenocarcinoma metastasis. Cell Death Dis. 2022, 13, 1032. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Cai, L.; Tian, X.; Li, W. MiR-543 promotes tumorigenesis and angiogenesis in non-small cell lung cancer via modulating metastasis associated protein 1. Mol. Med. 2020, 26, 44. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Hou, J.; Yang, C.; Wang, H.; Wu, S.; Wu, Y.; Zhao, X.; Lu, C. Extracellular vesicles secreted by hypoxia pre-challenged mesenchymal stem cells promote non-small cell lung cancer cell growth and mobility as well as macrophage M2 polarization via miR-21-5p delivery. J. Exp. Clin. Cancer Res. 2019, 38, 62. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Wei, K.; Yang, F.; Guo, Z.; Pan, C.; He, Y.; Wang, J.; Li, Z.; Chen, L.; Chen, Y.; et al. Tumor-derived exosomal miR-3157-3p promotes angiogenesis, vascular permeability and metastasis by targeting TIMP/KLF2 in non-small cell lung cancer. Cell Death Dis. 2021, 12, 840. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Park, S.; Kim, H.; Choi, Y.J.; Kim, S.Y.; Sung, K.J.; Sung, Y.H.; Choi, C.M.; Yun, M.; Yi, Y.S.; et al. Tumor-derived exosomal miR-619-5p promotes tumor angiogenesis and metastasis through the inhibition of RCAN1.4. Cancer Lett. 2020, 475, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; You, M.; Wang, F.; Wang, Z.; Gao, X.; Jing, C.; Liu, J.; Guo, M.; Li, J.; Luo, A.; et al. Multifunctional Graphdiyne-Cerium Oxide Nanozymes Facilitate MicroRNA Delivery and Attenuate Tumor Hypoxia for Highly Efficient Radiotherapy of Esophageal Cancer. Adv. Mater. 2021, 33, e2100556. [Google Scholar] [CrossRef]
- He, L.; Zhu, W.; Chen, Q.; Yuan, Y.; Wang, Y.; Wang, J.; Wu, X. Ovarian cancer cell-secreted exosomal miR-205 promotes metastasis by inducing angiogenesis. Theranostics 2019, 9, 8206–8220. [Google Scholar] [CrossRef]
- He, Q.; Ye, A.; Ye, W.; Liao, X.; Qin, G.; Xu, Y.; Yin, Y.; Luo, H.; Yi, M.; Xian, L.; et al. Cancer-secreted exosomal miR-21-5p induces angiogenesis and vascular permeability by targeting KRIT1. Cell Death Dis. 2021, 12, 576. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, D.; Zhang, Z.X.; Zhang, J.J.; He, H.Y.; Liu, J.L.; Wang, T.; Yang, X.X.; Liu, B.D.; Yang, L.L.; et al. Colorectal cancer cell-secreted exosomal miRNA N-72 promotes tumor angiogenesis by targeting CLDN18. Am. J. Cancer Res. 2023, 13, 3482–3499. [Google Scholar]
- Sun, Z.; Zhang, Q.; Yuan, W.; Li, X.; Chen, C.; Guo, Y.; Shao, B.; Dang, Q.; Zhou, Q.; Wang, Q.; et al. MiR-103a-3p promotes tumour glycolysis in colorectal cancer via hippo/YAP1/HIF1A axis. J. Exp. Clin. Cancer Res. 2020, 39, 250. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Yan, T.; Huang, C.; Xu, Z.; Wang, L.; Jiang, E.; Wang, H.; Chen, Y.; Liu, K.; Shao, Z.; et al. Melanoma cell-secreted exosomal miR-155-5p induce proangiogenic switch of cancer-associated fibroblasts via SOCS1/JAK2/STAT3 signaling pathway. J. Exp. Clin. Cancer Res. 2018, 37, 242. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Zhao, X.; Shao, C.; Fu, B.; Huang, Y.; Zhang, N.; Dou, X.; Zhang, Z.; Qiu, Y.; Wang, R.; et al. STIM1 promotes angiogenesis by reducing exosomal miR-145 in breast cancer MDA-MB-231 cells. Cell Death Dis. 2021, 12, 38. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhu, C.; Chen, C.; Zong, Y.; Feng, H.; Liu, D.; Feng, W.; Zhao, J.; Lu, A. CCL19 suppresses angiogenesis through promoting miR-206 and inhibiting Met/ERK/Elk-1/HIF-1alpha/VEGF-A pathway in colorectal cancer. Cell Death Dis. 2018, 9, 974. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Chen, J.J.; Feng, Y.; Yang, J.L.; Huang, H.; Chung, W.Y.; Hu, Y.L.; Xue, W.J. DNMT1-induced miR-378a-3p silencing promotes angiogenesis via the NF-kappaB signaling pathway by targeting TRAF1 in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2021, 40, 352. [Google Scholar] [CrossRef]
- Li, D.; Wang, T.; Sun, F.F.; Feng, J.Q.; Peng, J.J.; Li, H.; Wang, C.; Wang, D.; Liu, Y.; Bai, Y.D.; et al. MicroRNA-375 represses tumor angiogenesis and reverses resistance to sorafenib in hepatocarcinoma. Cancer Gene Ther. 2021, 28, 126–140. [Google Scholar] [CrossRef]
- Chen, M.; Li, Z.; Cao, L.; Fang, C.; Gao, R.; Liu, C. miR-877-3p inhibits tumor growth and angiogenesis of osteosarcoma through fibroblast growth factor 2 signaling. Bioengineered 2022, 13, 8174–8186. [Google Scholar] [CrossRef]
- Raimondi, L.; Gallo, A.; Cuscino, N.; De Luca, A.; Costa, V.; Carina, V.; Bellavia, D.; Bulati, M.; Alessandro, R.; Fini, M.; et al. Potential Anti-Metastatic Role of the Novel miR-CT3 in Tumor Angiogenesis and Osteosarcoma Invasion. Int. J. Mol. Sci. 2022, 23, 705. [Google Scholar] [CrossRef]
- Li, Z.; Li, S.; Wen, Y.; Chen, J.; Liu, K.; Jia, J. MiR-495 Inhibits Cisplatin Resistance and Angiogenesis in Esophageal Cancer by Targeting ATP7A. Technol. Cancer Res. Treat. 2021, 20, 15330338211039127. [Google Scholar] [CrossRef]
- Cao, Y. Antiangiogenic cancer therapy. Semin Cancer Biol. 2004, 14, 139–145. [Google Scholar] [CrossRef]
- Gajeton, J.; Krukovets, I.; Muppala, S.; Verbovetskiy, D.; Zhang, J.; Stenina-Adognravi, O. Hyperglycemia-Induced miR-467 Drives Tumor Inflammation and Growth in Breast Cancer. Cancers 2021, 13, 1346. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, P.; Hultine, S.; Smith, L.M.; Dews, M.; Fox, J.L.; Biyashev, D.; Schelter, J.M.; Huang, Q.; Cleary, M.A.; Volpert, O.V.; et al. p53-responsive miR-194 inhibits thrombospondin-1 and promotes angiogenesis in colon cancers. Cancer Res. 2011, 71, 7490–7501. [Google Scholar] [CrossRef] [PubMed]
- Perez-Gutierrez, L.; Ferrara, N. Biology and therapeutic targeting of vascular endothelial growth factor A. Nat. Rev. Mol. Cell Biol. 2023, 24, 816–834. [Google Scholar] [CrossRef] [PubMed]
- Alhasan, L. MiR-126 Modulates Angiogenesis in Breast Cancer by Targeting VEGF-A-mRNA. Asian Pac. J. Cancer Prev. 2019, 20, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Peng, X.C.; Zheng, X.L.; Wang, J.; Qin, Y.W. MiR-126 restoration down-regulate VEGF and inhibit the growth of lung cancer cell lines in vitro and in vivo. Lung Cancer 2009, 66, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Sasahira, T.; Kurihara, M.; Bhawal, U.K.; Ueda, N.; Shimomoto, T.; Yamamoto, K.; Kirita, T.; Kuniyasu, H. Downregulation of miR-126 induces angiogenesis and lymphangiogenesis by activation of VEGF-A in oral cancer. Br. J. Cancer 2012, 107, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Kong, R.; Ma, Y.; Feng, J.; Li, S.; Zhang, W.; Jiang, J.; Zhang, J.; Qiao, Z.; Yang, X.; Zhou, B. The crucial role of miR-126 on suppressing progression of esophageal cancer by targeting VEGF-A. Cell. Mol. Biol. Lett. 2016, 21, 3. [Google Scholar] [CrossRef] [PubMed]
- D’Onofrio, N.; Prattichizzo, F.; Martino, E.; Anastasio, C.; Mele, L.; La Grotta, R.; Sardu, C.; Ceriello, A.; Marfella, R.; Paolisso, G.; et al. MiR-27b attenuates mitochondrial oxidative stress and inflammation in endothelial cells. Redox Biol. 2023, 62, 102681. [Google Scholar] [CrossRef]
- Zeng, Z.; Li, Y.; Pan, Y.; Lan, X.; Song, F.; Sun, J.; Zhou, K.; Liu, X.; Ren, X.; Wang, F.; et al. Cancer-derived exosomal miR-25-3p promotes pre-metastatic niche formation by inducing vascular permeability and angiogenesis. Nat. Commun. 2018, 9, 5395. [Google Scholar] [CrossRef]
- Zhao, S.; Mi, Y.; Guan, B.; Zheng, B.; Wei, P.; Gu, Y.; Zhang, Z.; Cai, S.; Xu, Y.; Li, X.; et al. Tumor-derived exosomal miR-934 induces macrophage M2 polarization to promote liver metastasis of colorectal cancer. J. Hematol. Oncol. 2020, 13, 156. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, Q.; Mi, J.; Wang, S.; Xu, Q.; Zhuang, D.; Chen, W.; Liu, C.; Zhang, L.; Guo, J.; et al. Exosomes derived from stem cells of human deciduous exfoliated teeth inhibit angiogenesis in vivo and in vitro via the transfer of miR-100-5p and miR-1246. Stem Cell Res. Ther. 2022, 13, 89. [Google Scholar] [CrossRef] [PubMed]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [PubMed]
- Batista, P.J.; Chang, H.Y. Long noncoding RNAs: Cellular address codes in development and disease. Cell 2013, 152, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Ulitsky, I.; Bartel, D.P. lincRNAs: Genomics, evolution, and mechanisms. Cell 2013, 154, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Chi, Y.; Wang, D.; Wang, J.; Yu, W.; Yang, J. Long Non-Coding RNA in the Pathogenesis of Cancers. Cells 2019, 8, 1015. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tan, L.; Yu, X.; Cao, X.; Jia, B.; Chen, R.; Li, J. lncRNA ZNRD1-AS1 promotes malignant lung cell proliferation, migration, and angiogenesis via the miR-942/TNS1 axis and is positively regulated by the m(6)A reader YTHDC2. Mol. Cancer 2022, 21, 229. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, A.; Wang, Z.; Wang, B.; Chai, X.; Lu, W.; Cao, T.; Li, R.; Wu, M.; Lu, Z.; et al. LINC00173.v1 promotes angiogenesis and progression of lung squamous cell carcinoma by sponging miR-511-5p to regulate VEGFA expression. Mol. Cancer 2020, 19, 98. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Ruan, S.; Wang, J.; Xia, Y.; Le, K.; Xiao, X.; Hu, T.; Wang, Q. M2 macrophage-induced lncRNA PCAT6 facilitates tumorigenesis and angiogenesis of triple-negative breast cancer through modulation of VEGFR2. Cell Death Dis. 2020, 11, 728. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, T.; Wang, Z.; Kuang, X.; Shao, N.; Lin, Y. lncRNA NR2F1-AS1 promotes breast cancer angiogenesis through activating IGF-1/IGF-1R/ERK pathway. J. Cell Mol. Med. 2020, 24, 8236–8247. [Google Scholar] [CrossRef]
- Sun, D.S.; Guan, C.H.; Wang, W.N.; Hu, Z.T.; Zhao, Y.Q.; Jiang, X.M. LncRNA NORAD promotes proliferation, migration and angiogenesis of hepatocellular carcinoma cells through targeting miR-211-5p/FOXD1/VEGF-A axis. Microvasc. Res. 2021, 134, 104120. [Google Scholar] [CrossRef]
- Teng, F.; Zhang, J.X.; Chang, Q.M.; Wu, X.B.; Tang, W.G.; Wang, J.F.; Feng, J.F.; Zhang, Z.P.; Hu, Z.Q. LncRNA MYLK-AS1 facilitates tumor progression and angiogenesis by targeting miR-424-5p/E2F7 axis and activating VEGFR-2 signaling pathway in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2020, 39, 235. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Qiu, X. Corrigendum to “LncRNA miR503HG inhibits epithelial-mesenchymal transition andangiogenesis in hepatocellular carcinoma by enhancing PDCD4 viaregulation of miR-15b”. Dig. Liver Dis. 2021, 53, 1219. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Pang, X.; Lei, L.; Zhang, J.; Zhang, X.; Chen, Z.; Zhu, J.; Jiang, Y.; Chen, G.; Wu, Y.; et al. LncRNA CRART16/miR-122-5p/FOS axis promotes angiogenesis of gastric cancer by upregulating VEGFD expression. Aging 2022, 14, 4137–4157. [Google Scholar] [CrossRef] [PubMed]
- Teng, F.; Zhang, J.X.; Chen, Y.; Shen, X.D.; Su, C.; Guo, Y.J.; Wang, P.H.; Shi, C.C.; Lei, M.; Cao, Y.O.; et al. LncRNA NKX2-1-AS1 promotes tumor progression and angiogenesis via upregulation of SERPINE1 expression and activation of the VEGFR-2 signaling pathway in gastric cancer. Mol. Oncol. 2021, 15, 1234–1255. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Z.; Tian, Y.F.; Wu, H.; Ouyang, S.Y.; Kuang, W.L. LncRNA H19 promotes glioma angiogenesis through miR-138/HIF-1alpha/VEGF axis. Neoplasma 2020, 67, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Liu, Z.Z.; Wu, H.; Kuang, W.L. LncRNA H19 Promotes Cell Proliferation, Migration, and Angiogenesis of Glioma by Regulating Wnt5a/beta-Catenin Pathway via Targeting miR-342. Cell Mol. Neurobiol. 2022, 42, 1065–1077. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Yang, F.; Qi, X.; Li, Q.; Wang, D.; Yi, T.; Yin, R.; Zhao, X.; Zhong, X.; Bian, C. LncRNA DANCR promotes tumor growth and angiogenesis in ovarian cancer through direct targeting of miR-145. Mol. Carcinog. 2019, 58, 2286–2296. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Tang, G.; Qian, H.; Chen, J.; Cheng, B.; Zhou, C.; Shen, Y. LncRNA LOC100129620 promotes osteosarcoma progression through regulating CDK6 expression, tumor angiogenesis, and macrophage polarization. Aging 2021, 13, 14258–14276. [Google Scholar] [CrossRef]
- Guo, Y.P.; Wang, Z.F.; Li, N.; Lei, Q.Q.; Cheng, Q.; Shi, L.G.; Zhou, S.L.; Wang, X.H.; Sun, Y.; Kong, L.F. Suppression of lncRNA HOTAIR alleviates RCC angiogenesis through regulating miR-126/EGFL7 axis. Am. J. Physiol. Cell Physiol. 2021, 320, C880–C891. [Google Scholar] [CrossRef]
- Cao, J.; Yang, Z.; An, R.; Zhang, J.; Zhao, R.; Li, W.; Xu, L.; Sun, Y.; Liu, M.; Tian, L. lncRNA IGKJ2-MALLP2 suppresses LSCC proliferation, migration, invasion, and angiogenesis by sponging miR-1911-3p/p21. Cancer Sci. 2020, 111, 3245–3257. [Google Scholar] [CrossRef]
- Zeng, H.; Hou, Y.; Zhou, X.; Lang, L.; Luo, H.; Sun, Y.; Wan, X.; Yuan, T.; Wang, R.; Liu, Y.; et al. Cancer-associated fibroblasts facilitate premetastatic niche formation through lncRNA SNHG5-mediated angiogenesis and vascular permeability in breast cancer. Theranostics 2022, 12, 7351–7370. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Bao, L.; Chen, Y.; Wang, C.; Luo, M.; Zhang, B.; Zhou, M.; Wang, J.E.; Fang, Y.V.; Kumar, A.; et al. HIF2-Induced Long Noncoding RNA RAB11B-AS1 Promotes Hypoxia-Mediated Angiogenesis and Breast Cancer Metastasis. Cancer Res. 2020, 80, 964–975. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Xu, Z.; Chen, L.; Wei, Q.; Huang, Z.; Liu, G.; Li, W.; Wang, J.; Tang, Q.; Pu, J. Long non-coding RNA PAARH promotes hepatocellular carcinoma progression and angiogenesis via upregulating HOTTIP and activating HIF-1alpha/VEGF signaling. Cell Death Dis. 2022, 13, 102. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, H.; Hou, Y. LncRNA MAGI2-AS3 inhibits tumor progression and angiogenesis by regulating ACY1 via interacting with transcription factor HEY1 in clear cell renal cell carcinoma. Cancer Gene Ther. 2022, 29, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Sun, H.; Luo, D.; Gan, L.; Mo, S.; Dai, W.; Liang, L.; Yang, Y.; Xu, M.; Li, J.; et al. Lnc-RP11-536 K7.3/SOX2/HIF-1alpha signaling axis regulates oxaliplatin resistance in patient-derived colorectal cancer organoids. J. Exp. Clin. Cancer Res. 2021, 40, 348. [Google Scholar] [CrossRef]
- Wang, X.; Li, L.; Zhao, K.; Lin, Q.; Li, H.; Xue, X.; Ge, W.; He, H.; Liu, D.; Xie, H.; et al. A novel LncRNA HITT forms a regulatory loop with HIF-1alpha to modulate angiogenesis and tumor growth. Cell Death Differ. 2020, 27, 1431–1446. [Google Scholar] [CrossRef] [PubMed]
- Jia, P.; Cai, H.; Liu, X.; Chen, J.; Ma, J.; Wang, P.; Liu, Y.; Zheng, J.; Xue, Y. Long non-coding RNA H19 regulates glioma angiogenesis and the biological behavior of glioma-associated endothelial cells by inhibiting microRNA-29a. Cancer Lett. 2016, 381, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, G.; Wang, B.; Yuan, Z.; Liu, G.; Niu, B.; Chen, Y.; Zhou, S.; He, J.; Xue, H. Long non-coding RNA BZRAP1-AS1 silencing suppresses tumor angiogenesis in hepatocellular carcinoma by mediating THBS1 methylation. J. Transl. Med. 2019, 17, 421. [Google Scholar] [CrossRef]
- Zhao, J.; Du, P.; Cui, P.; Qin, Y.; Hu, C.; Wu, J.; Zhou, Z.; Zhang, W.; Qin, L.; Huang, G. LncRNA PVT1 promotes angiogenesis via activating the STAT3/VEGFA axis in gastric cancer. Oncogene 2018, 37, 4094–4109. [Google Scholar] [CrossRef]
- Hou, Y.; Jia, H.; Cao, Y.; Zhang, S.; Zhang, X.; Wei, P.; Xie, J.; Dong, W.; Wang, B. LncRNA EPIC1 promotes tumor angiogenesis via activating the Ang2/Tie2 axis in non-small cell lung cancer. Life Sci. 2021, 267, 118933. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, S.; Zhu, X.; Zhang, L.; Deng, J.; Li, F.; Guo, B.; Zhang, S.; Wu, R.; Zhang, Z.; et al. LncRNA-encoded polypeptide ASRPS inhibits triple-negative breast cancer angiogenesis. J. Exp. Med. 2020, 217, e20190950. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, J.; Zhang, H.; Zhang, Y.; Wang, X.; Yang, H.; Zhou, Z.; Hao, X.; Ying, G.; Ba, Y. Gastric cancer derived exosomes mediate the delivery of circRNA to promote angiogenesis by targeting miR-29a/VEGF axis in endothelial cells. Biochem. Biophys. Res. Commun. 2021, 560, 37–44. [Google Scholar] [CrossRef]
- Du, Y.; Zhang, J.Y.; Gong, L.P.; Feng, Z.Y.; Wang, D.; Pan, Y.H.; Sun, L.P.; Wen, J.Y.; Chen, G.F.; Liang, J.; et al. Hypoxia-induced ebv-circLMP2A promotes angiogenesis in EBV-associated gastric carcinoma through the KHSRP/VHL/HIF1alpha/VEGFA pathway. Cancer Lett. 2022, 526, 259–272. [Google Scholar] [CrossRef]
- Xie, M.; Yu, T.; Jing, X.; Ma, L.; Fan, Y.; Yang, F.; Ma, P.; Jiang, H.; Wu, X.; Shu, Y.; et al. Exosomal circSHKBP1 promotes gastric cancer progression via regulating the miR-582-3p/HUR/VEGF axis and suppressing HSP90 degradation. Mol. Cancer 2020, 19, 112. [Google Scholar] [CrossRef]
- Li, F.; Cai, Y.; Deng, S.; Yang, L.; Liu, N.; Chang, X.; Jing, L.; Zhou, Y.; Li, H. A peptide CORO1C-47aa encoded by the circular noncoding RNA circ-0000437 functions as a negative regulator in endometrium tumor angiogenesis. J. Biol. Chem. 2021, 297, 101182. [Google Scholar] [CrossRef]
- Shi, P.; Liu, Y.; Yang, H.; Hu, B. Breast cancer derived exosomes promoted angiogenesis of endothelial cells in microenvironment via circHIPK3/miR-124-3p/MTDH axis. Cell. Signal. 2022, 95, 110338. [Google Scholar] [CrossRef]
- Ma, J.; Huang, L.; Gao, Y.B.; Li, M.X.; Chen, L.L.; Yang, L. M2 macrophage facilitated angiogenesis in cutaneous squamous cell carcinoma via circ_TNFRSF21/miR-3619-5p/ROCK axis. Kaohsiung J. Med. Sci. 2022, 38, 761–771. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhou, J.; Zhao, J.; Zhang, H.; Li, L.; Li, H.; Chen, L.; Hu, J.; Zheng, W.; Jing, Z. The U2AF2 /circRNA ARF1/miR-342-3p/ISL2 feedback loop regulates angiogenesis in glioma stem cells. J. Exp. Clin. Cancer Res. 2020, 39, 182. [Google Scholar] [CrossRef]
- Deng, L.; Gong, K.; Wang, G. Hsa_circ_0008344 Promotes Glioma Tumor Progression and Angiogenesis Presumably by Regulating miR-638/SZRD1 Pathway. Neurotox. Res. 2022, 40, 825–836. [Google Scholar] [CrossRef]
- Liu, X.; Shen, S.; Zhu, L.; Su, R.; Zheng, J.; Ruan, X.; Shao, L.; Wang, D.; Yang, C.; Liu, Y. SRSF10 inhibits biogenesis of circ-ATXN1 to regulate glioma angiogenesis via miR-526b-3p/MMP2 pathway. J. Exp. Clin. Cancer Res. 2020, 39, 121. [Google Scholar] [CrossRef]
- Guo, Y.; Guo, Y.; Chen, C.; Fan, D.; Wu, X.; Zhao, L.; Shao, B.; Sun, Z.; Ji, Z. Circ3823 contributes to growth, metastasis and angiogenesis of colorectal cancer: Involvement of miR-30c-5p/TCF7 axis. Mol. Cancer 2021, 20, 93. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, Y.; Liu, L.; Si, C.; Xu, Y.; Wu, X.; Wang, C.; Sun, Z.; Kang, Q. Exosomal circTUBGCP4 promotes vascular endothelial cell tipping and colorectal cancer metastasis by activating Akt signaling pathway. J. Exp. Clin. Cancer Res. 2023, 42, 46. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Han, C.; Zhai, T.; Zhang, X.; Chen, C.; Lian, L. Circ_0030998 promotes tumor proliferation and angiogenesis by sponging miR-567 to regulate VEGFA in colorectal cancer. Cell Death Discov. 2021, 7, 160. [Google Scholar] [CrossRef]
- Wang, J.; Lin, Y.; Jiang, D.H.; Yang, X.; He, X.G. CircRNA ZNF609 promotes angiogenesis in nasopharyngeal carcinoma by regulating miR-145/STMN1 axis. Kaohsiung J. Med. Sci. 2021, 37, 686–698. [Google Scholar] [CrossRef]
- Yu, L.; Zhu, H.; Wang, Z.; Huang, J.; Zhu, Y.; Fan, G.; Wang, Y.; Chen, X.; Zhou, G. Circular RNA circFIRRE drives osteosarcoma progression and metastasis through tumorigenic-angiogenic coupling. Mol. Cancer 2022, 21, 167. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, Z.; Wu, B.; Sun, F. Circular RNA circFOXP1 promotes angiogenesis by regulating microRNA -127-5p/CDKN2AIP signaling pathway in osteosarcoma. Bioengineered 2021, 12, 9991–9999. [Google Scholar] [CrossRef]
- Zhang, X.; Tan, P.; Zhuang, Y.; Du, L. hsa_circRNA_001587 upregulates SLC4A4 expression to inhibit migration, invasion, and angiogenesis of pancreatic cancer cells via binding to microRNA-223. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G703–G717. [Google Scholar] [CrossRef]
- Liu, X.; Zhong, L.; Jiang, W.; Wen, D. Repression of circRNA_000684 inhibits malignant phenotypes of pancreatic ductal adenocarcinoma cells via miR-145-mediated KLF5. Pancreatology 2021, 21, 406–417. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhu, Z.; Xiao, W.; Zong, G.; Wang, C.; Jiang, W.; Li, K.; Shen, J.; Guo, X.; Cui, J.; et al. Hypoxia-induced circRNF13 promotes the progression and glycolysis of pancreatic cancer. Exp. Mol. Med. 2022, 54, 1940–1954. [Google Scholar] [CrossRef]
- Guo, X.Y.; Liu, T.T.; Zhu, W.J.; Liu, H.T.; Zhang, G.H.; Song, L.; Zhao, R.N.; Chen, X.; Gao, P. CircKDM4B suppresses breast cancer progression via the miR-675/NEDD4L axis. Oncogene 2022, 41, 1895–1906. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, J.; Wang, J.; Li, Y.; Hu, G.; Li, H. Circ_0001667 accelerates breast cancer proliferation and angiogenesis through regulating CXCL10 expression by sponging miR-6838-5p. Thorac. Cancer 2023, 14, 881–892. [Google Scholar] [CrossRef]
- Liu, C.; Yang, J.; Zhu, F.; Zhao, Z.; Gao, L. Exosomal circ_0001190 Regulates the Progression of Gastric Cancer via miR-586/SOSTDC1 Axis. Biochem. Genet. 2022, 60, 1895–1913. [Google Scholar] [CrossRef]
- Ma, H.; Qu, S.; Zhai, Y.; Yang, X. circ_0025033 promotes ovarian cancer development via regulating the hsa_miR-370-3p/SLC1A5 axis. Cell. Mol. Biol. Lett. 2022, 27, 94. [Google Scholar] [CrossRef]
- Ye, H.; Wang, R.Y.; Yu, X.Z.; Wu, Y.K.; Yang, B.W.; Ao, M.Y.; Xi, M.R.; Hou, M.M. Exosomal circNFIX promotes angiogenesis in ovarian cancer via miR-518a-3p/TRIM44 axis. Kaohsiung J. Med. Sci. 2023, 39, 26–39. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Zhou, J.H.; Shen, F.R.; Shi, X.; Chen, Y.G. CircATRNL1 activates Smad4 signaling to inhibit angiogenesis and ovarian cancer metastasis via miR-378. Mol. Oncol. 2021, 15, 1217–1233. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, Y.; Lin, Q.; Zhou, J.; Lv, X.; Song, Y. Hsa_circ_0111738 Inhibits Tumor Progression and Angiogenesis in Multiple Myeloma by Sponging miR-1233-3p to Regulate HIF-1 Signaling Pathway. Arch. Med. Res. 2023, 54, 299–309. [Google Scholar] [CrossRef]
- Xue, L.; Jia, T.; Zhu, Y.; Zhao, L.; Mao, J. Down-regulation of circ_0058058 suppresses proliferation, angiogenesis and metastasis in multiple myeloma through miR-338-3p/ATG14 pathway. J. Orthop. Surg. Res. 2021, 16, 723. [Google Scholar] [CrossRef]
- Zhou, T.; Zhao, S.; Tang, S.; Wang, Y.; Wu, R.; Zeng, X.; Yang, P.; Zhang, X.; Tian, X. Guggulsterone Promotes Nasopharyngeal Carcinoma Cells Exosomal Circfip1L1 to Mediate miR-125a-5p/VEGFA Affecting Tumor Angiogenesis. Curr. Mol. Pharmacol. 2023, 16, 870–880. [Google Scholar] [CrossRef]
- Ren, M.; Song, X.; Niu, J.; Tang, G.; Sun, Z.; Li, Y.; Kong, F. The malignant property of circHIPK2 for angiogenesis and chemoresistance in non-small cell lung cancer. Exp. Cell Res. 2022, 419, 113276. [Google Scholar] [CrossRef]
- Yang, C.; Shi, J.; Wang, J.; Hao, D.; An, J.; Jiang, J. Circ_0006988 promotes the proliferation, metastasis and angiogenesis of non-small cell lung cancer cells by modulating miR-491-5p/MAP3K3 axis. Cell Cycle 2021, 20, 1334–1346. [Google Scholar] [CrossRef]
- Zhuang, Q.; Huang, Z.; Zhuang, W.; Hong, Y.; Huang, Y. Knockdown of circ-RAD23B inhibits non-small cell lung cancer progression via the miR-142-3p/MAP4K3 axis. Thorac. Cancer 2022, 13, 750–760. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, F.Q.; Sun, C.M.; Huang, J.H.; Zhang, H.M.; Li, X.; Wang, G.C.; Zhang, N.; Che, J.P.; Zhang, W.T.; et al. circPRRC2A promotes angiogenesis and metastasis through epithelial-mesenchymal transition and upregulates TRPM3 in renal cell carcinoma. Theranostics 2020, 10, 4395–4409. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Dai, X.; Chen, X.; Chen, Y.; Wu, S.; Zhou, Q.; Yang, C.; Jiang, H. Circ-AFAP1 promote clear cell renal cell carcinoma growth and angiogenesis by the Circ-AFAP1/miR-374b-3p/VEGFA signaling axis. Cell Death Discov. 2022, 8, 68. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lv, J.; Hou, L.; Guo, X. Circ_0001955 Acts as a miR-646 Sponge to Promote the Proliferation, Metastasis and Angiogenesis of Hepatocellular Carcinoma. Dig. Dis. Sci. 2022, 67, 2257–2268. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, H.; Zhang, Y.; Wang, Q.; Li, S.; Wang, Z.; Shi, X. Circular RNA hsa_circ_0000519 contributes to angiogenesis and tumor progression in hepatocellular carcinoma through the miR-1296/E2F7 axis. Hum. Cell 2023, 36, 738–751. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Jia, W.; Feng, Y.; Wang, J.; Wang, J.; Zhu, D.; Xu, C.; Liang, L.; Ding, W.; Zhou, Y.; et al. Exosome-transported circHDAC1_004 Promotes Proliferation, Migration, and Angiogenesis of Hepatocellular Carcinoma by the miR-361-3p/NACC1 Axis. J. Clin. Transl. Hepatol. 2023, 11, 1079–1093. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Yang, D.; Guo, P. Circ_0000144 acts as a miR-1178-3p decoy to promote cell malignancy and angiogenesis by increasing YWHAH expression in papillary thyroid cancer. J. Otolaryngol. Head Neck Surg. 2022, 51, 28. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, W.; Su, Z.; Zhang, J.; Cao, H. Circ_0011058 facilitates proliferation, angiogenesis and radioresistance in papillary thyroid cancer cells by positively regulating YAP1 via acting as miR-335-5p sponge. Cell. Signal. 2021, 88, 110155. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, H.; Wang, J. Circular RNA CircSHKBP1 accelerates the proliferation, invasion, angiogenesis, and stem cell-like properties via modulation of microR-766-5p/high mobility group AT-hook 2 axis in laryngeal squamous cell carcinoma. Bioengineered 2022, 13, 11551–11563. [Google Scholar] [CrossRef]
- Ren, X.; Cheng, J.; Zhu, M.; Chen, X.; Jiang, M.; Hu, X.; Lu, Y. Circular RNA circ_0062019 exerts oncogenic properties in prostate cancer via mediating miR-1253/NRBP1 axis. Andrologia 2022, 54, e14343. [Google Scholar] [CrossRef]
- Wang, D.; Yan, S.; Wang, L.; Li, Y.; Qiao, B. circSLC8A1 Acts as a Tumor Suppressor in Prostate Cancer via Sponging miR-21. Biomed. Res. Int. 2021, 2021, 6614591. [Google Scholar] [CrossRef] [PubMed]
- Han, T.; Shi, M.; Chen, G.; Hao, J. Circ_0008726 promotes malignant progression of ESCC cells through miR-206/HOXA13 pathway. Gen. Thorac. Cardiovasc. Surg. 2023, 71, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhao, J.; Xu, J.; Zhang, H.; Zhou, J.; Li, H.; Zhang, G.; Xu, K.; Jing, Z. Glioblastoma-associated microglia-derived exosomal circKIF18A promotes angiogenesis by targeting FOXC2. Oncogene 2022, 41, 3461–3473. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, S.; Liu, R.; Chen, H.; Xu, B.; Xu, W.; Chen, M. Circular RNA circPOLR2A promotes clear cell renal cell carcinoma progression by facilitating the UBE3C-induced ubiquitination of PEBP1 and, thereby, activating the ERK signaling pathway. Mol. Cancer 2022, 21, 146. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, C.; Zhang, H.; Li, Y.; Hou, D.; Liu, D.; Xu, R.; Cheng, J.; Liu, L.; Fu, Y.; et al. circFNDC3B Accelerates Vasculature Formation and Metastasis in Oral Squamous Cell Carcinoma. Cancer Res. 2023, 83, 1459–1475. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, M.; Bi, X.; Hao, L.; Liu, R.; Zhang, H. ESR1 mediated circ_0004018 suppresses angiogenesis in hepatocellular carcinoma via recruiting FUS and stabilizing TIMP2 expression. Exp. Cell Res. 2021, 408, 112804. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, D.; Caponnetto, A.; Brex, D.; Mirabella, F.; Barbagallo, C.; Lauretta, G.; Morrone, A.; Certo, F.; Broggi, G.; Caltabiano, R.; et al. CircSMARCA5 Regulates VEGFA mRNA Splicing and Angiogenesis in Glioblastoma Multiforme Through the Binding of SRSF1. Cancers 2019, 11, 194. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, J.; Hou, Y. Classification, function, and advances in tsRNA in non-neoplastic diseases. Cell Death Dis. 2023, 14, 748. [Google Scholar] [CrossRef]
- Zhou, J.; Wan, F.; Wang, Y.; Long, J.; Zhu, X. Small RNA sequencing reveals a novel tsRNA-26576 mediating tumorigenesis of breast cancer. Cancer Manag. Res. 2019, 11, 3945–3956. [Google Scholar] [CrossRef]
- Tao, E.W.; Wang, H.L.; Cheng, W.Y.; Liu, Q.Q.; Chen, Y.X.; Gao, Q.Y. A specific tRNA half, 5′tiRNA-His-GTG, responds to hypoxia via the HIF1alpha/ANG axis and promotes colorectal cancer progression by regulating LATS2. J. Exp. Clin. Cancer Res. 2021, 40, 67. [Google Scholar] [CrossRef]
- Filipowicz, W.; Pelczar, P.; Pogacic, V.; Dragon, F. Structure and biogenesis of small nucleolar RNAs acting as guides for ribosomal RNA modification. Acta Biochim. Pol. 1999, 46, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Mourksi, N.E.; Morin, C.; Fenouil, T.; Diaz, J.J.; Marcel, V. snoRNAs Offer Novel Insight and Promising Perspectives for Lung Cancer Understanding and Management. Cells 2020, 9, 541. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.P.; Liao, J.P.; Shen, J.; Yu, L.; Liu, B.L.; Liu, L.; Li, R.Y.; Ji, L.; Dorsey, S.G.; Jiang, Z.R.; et al. Small nucleolar RNA 42 acts as an oncogene in lung tumorigenesis. Oncogene 2012, 31, 2794–2804. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, J.; Yao, Y.; Liu, Y.; Xia, P.; Zhang, H.; Yin, M.; Qin, Z.; Ma, W.; Yuan, Y. Small nucleolar RNA 42 promotes the growth of hepatocellular carcinoma through the p53 signaling pathway. Cell Death Discov. 2021, 7, 347. [Google Scholar] [CrossRef] [PubMed]
- Okugawa, Y.; Toiyama, Y.; Toden, S.; Mitoma, H.; Nagasaka, T.; Tanaka, K.; Inoue, Y.; Kusunoki, M.; Boland, C.R.; Goel, A. Clinical significance of SNORA42 as an oncogene and a prognostic biomarker in colorectal cancer. Gut 2017, 66, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Li, G.; Liao, J.; Huang, Z.; Wen, J.; Wang, Y.; Chen, Z.; Cai, G.; Xu, W.; Ding, Z.; et al. Non-coding small nucleolar RNA SNORD17 promotes the progression of hepatocellular carcinoma through a positive feedback loop upon p53 inactivation. Cell Death Differ. 2022, 29, 988–1003. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.N.; Li, Y.; Xia, S.Q.; Zhang, Y.Y.; Zheng, J.H.; Li, W. PIWI Proteins and PIWI-Interacting RNA: Emerging Roles in Cancer. Cell Physiol. Biochem. 2017, 44, 1–20. [Google Scholar] [CrossRef]
- Liu, P.; Dong, Y.; Gu, J.; Puthiyakunnon, S.; Wu, Y.; Chen, X.G. Developmental piRNA profiles of the invasive vector mosquito Aedes albopictus. Parasit. Vectors 2016, 9, 524. [Google Scholar] [CrossRef]
- Liu, Y.; Dou, M.; Song, X.; Dong, Y.; Liu, S.; Liu, H.; Tao, J.; Li, W.; Yin, X.; Xu, W. The emerging role of the piRNA/piwi complex in cancer. Mol. Cancer 2019, 18, 123. [Google Scholar] [CrossRef]
- Yan, H.; Wu, Q.L.; Sun, C.Y.; Ai, L.S.; Deng, J.; Zhang, L.; Chen, L.; Chu, Z.B.; Tang, B.; Wang, K.; et al. piRNA-823 contributes to tumorigenesis by regulating de novo DNA methylation and angiogenesis in multiple myeloma. Leukemia 2015, 29, 196–206. [Google Scholar] [CrossRef]
- Zhao, Q.; Qian, L.; Guo, Y.; Lu, J.; Li, D.; Xie, H.; Wang, Q.; Ma, W.; Liu, P.; Liu, Y.; et al. IL11 signaling mediates piR-2158 suppression of cell stemness and angiogenesis in breast cancer. Theranostics 2023, 13, 2337–2349. [Google Scholar] [CrossRef]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA therapeutics—Challenges and potential solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Fabbri, M.; Calin, G.A. MicroRNAs and other non-coding RNAs as targets for anticancer drug development. Nat. Rev. Drug Discov. 2013, 12, 847–865. [Google Scholar] [CrossRef]
- Slack, F.J.; Chinnaiyan, A.M. The Role of Non-coding RNAs in Oncology. Cell 2019, 179, 1033–1055. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.Y.; Ferrajoli, A.; Sood, A.K.; Lopez-Berestein, G.; Calin, G.A. microRNA Therapeutics in Cancer—An Emerging Concept. EBioMedicine 2016, 12, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xue, W.; Li, X.; Zhang, J.; Chen, S.; Zhang, J.L.; Yang, L.; Chen, L.L. The Biogenesis of Nascent Circular RNAs. Cell Rep. 2016, 15, 611–624. [Google Scholar] [CrossRef]
- Obi, P.; Chen, Y.G. The design and synthesis of circular RNAs. Methods 2021, 196, 85–103. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Zhou, S.; Dain, L.; Mei, L.; Zhu, G. Circular RNA: An emerging frontier in RNA therapeutic targets, RNA therapeutics, and mRNA vaccines. J. Control Release 2022, 348, 84–94. [Google Scholar] [CrossRef]
- Bouchie, A. First microRNA mimic enters clinic. Nat. Biotechnol. 2013, 31, 577. [Google Scholar] [CrossRef]
- Hong, D.S.; Kang, Y.K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.L.; Kim, T.Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- van Zandwijk, N.; Pavlakis, N.; Kao, S.C.; Linton, A.; Boyer, M.J.; Clarke, S.; Huynh, Y.; Chrzanowska, A.; Fulham, M.J.; Bailey, D.L.; et al. Safety and activity of microRNA-loaded minicells in patients with recurrent malignant pleural mesothelioma: A first-in-man, phase 1, open-label, dose-escalation study. Lancet Oncol. 2017, 18, 1386–1396. [Google Scholar] [CrossRef] [PubMed]
- Telford, B.J.; Yahyanejad, S.; de Gunst, T.; den Boer, H.C.; Vos, R.M.; Stegink, M.; van den Bosch, M.T.J.; Alemdehy, M.F.; van Pinxteren, L.A.H.; Schaapveld, R.Q.J.; et al. Multi-modal effects of 1B3, a novel synthetic miR-193a-3p mimic, support strong potential for therapeutic intervention in oncology. Oncotarget 2021, 12, 422–439. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Wu, Y.; Zeng, B.; Sun, J.; Li, Y.; Luo, J.; Wang, L.; Yi, Z.; Li, H.; Ren, G. CircEHMT1 inhibits metastatic potential of breast cancer cells by modulating miR-1233-3p/KLF4/MMP2 axis. Biochem. Biophys. Res. Commun. 2020, 526, 306–313. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mechanisms of miRNA regulation of tumor angiogenesis. miRNAs can play a role in tumor cells and vascular endothelial cells to regulate tumor angiogenesis. In the process of tumor angiogenesis, miRNAs primarily function by binding to the 3′-UTR of specific mRNAs, leading to mRNA degradation or translation inhibition. These target genes mainly include pro-angiogenic or anti-angiogenic factors, as well as genes involved in angiogenesis signaling pathways. A similar mechanism also exists in vascular endothelial cells. For example, miR-27b-3p targets the 3′-UTR region of FOXO1 mRNA and downregulates its expression, thereby regulating the expression of VEGFR. In addition, exosomal miRNAs have a significant effect on angiogenesis. For example, miR-25-3p targets and silences KLF2 and KLF4, ultimately regulating the expression of genes associated with VEGFR2 and ZO-1 in endothelial cells.
Figure 1.
Mechanisms of miRNA regulation of tumor angiogenesis. miRNAs can play a role in tumor cells and vascular endothelial cells to regulate tumor angiogenesis. In the process of tumor angiogenesis, miRNAs primarily function by binding to the 3′-UTR of specific mRNAs, leading to mRNA degradation or translation inhibition. These target genes mainly include pro-angiogenic or anti-angiogenic factors, as well as genes involved in angiogenesis signaling pathways. A similar mechanism also exists in vascular endothelial cells. For example, miR-27b-3p targets the 3′-UTR region of FOXO1 mRNA and downregulates its expression, thereby regulating the expression of VEGFR. In addition, exosomal miRNAs have a significant effect on angiogenesis. For example, miR-25-3p targets and silences KLF2 and KLF4, ultimately regulating the expression of genes associated with VEGFR2 and ZO-1 in endothelial cells.

Figure 2.
Mechanisms of lncRNA regulation of tumor angiogenesis. In certain tumor cells and specific tissues, some lncRNAs carry specific sequences that can adsorb miRNA, acting in a similar way to sponges in order to bind with miRNA, thereby preventing miRNA from binding to its target mRNA. On the other hand, lncRNA can interact with certain proteins, affecting their post-translational modifications, protein stability, transcription, and translation activities, ultimately affecting tumor angiogenesis through the regulation of downstream target genes.
Figure 2.
Mechanisms of lncRNA regulation of tumor angiogenesis. In certain tumor cells and specific tissues, some lncRNAs carry specific sequences that can adsorb miRNA, acting in a similar way to sponges in order to bind with miRNA, thereby preventing miRNA from binding to its target mRNA. On the other hand, lncRNA can interact with certain proteins, affecting their post-translational modifications, protein stability, transcription, and translation activities, ultimately affecting tumor angiogenesis through the regulation of downstream target genes.

Figure 3.
Mechanisms of circRNA regulation of tumor angiogenesis. Some circRNAs function as sponges, absorbing miRNAs, inhibiting the binding of miRNAs to target genes, reducing the degradation of target mRNA, and thereby affecting tumor angiogenesis. Certain circRNAs have binding sites with the protein KHSRP, influencing the degradation of target mRNA and impacting tumor angiogenesis. The small peptides encoded by circRNAs play a role in influencing tumor angiogenesis.
Figure 3.
Mechanisms of circRNA regulation of tumor angiogenesis. Some circRNAs function as sponges, absorbing miRNAs, inhibiting the binding of miRNAs to target genes, reducing the degradation of target mRNA, and thereby affecting tumor angiogenesis. Certain circRNAs have binding sites with the protein KHSRP, influencing the degradation of target mRNA and impacting tumor angiogenesis. The small peptides encoded by circRNAs play a role in influencing tumor angiogenesis.

Table 1.
Targets and dysregulation of miRNA associated with tumor angiogenesis.
| MiRNA | Target Genes | Cancer Types | The Role in Tumor Angiogenesis | References |
|---|---|---|---|---|
| miR-130b-3p | HOXA5 | Hepatocellular Carcinoma | Promoter | [28] |
| miR-130b-3p | MBNL1 | Head and Neck Squamous Cell Carcinoma | Promoter | [29] |
| miR-519a-3p | DUSP2 | Gastric Cancer | Promoter | [30] |
| miR-30b-5p | GJA1 | Pancreatic Cancer | Promoter | [31] |
| miR-197-3p | TIMP2/3 | Lung Adenocarcinoma Metastasis | Promoter | [32] |
| miR-543 | MTA1 | Non-small Cell Lung Cancer | Promoter | [33] |
| miR-21-5p | PTEN, PDCD4, and RECK | Non-small Cell Lung Cancer | Promoter | [34] |
| miR-3157-3p | TIMP/KLF2 | Non-small Cell Lung Cancer | Promoter | [35] |
| miR-619-5p | RCAN1.4 | Non-small Cell Lung Cancer | Promoter | [36] |
| miR-181a | RAD17 | Esophageal Cancer | Promoter | [37] |
| miR-205 | PTEN | Ovarian Cancer | Promoter | [38] |
| miR-21-5p | KRIT1 | Colorectal Cancer | Promoter | [39] |
| miR-N-72 | CLDN18 | Colorectal Cancer | Promoter | [40] |
| miR-103a-3p | LATS2 and SAV1 | Colorectal Cancer | Promoter | [41] |
| miR-155-5p | SOCS1 | Melanoma | Promoter | [42] |
| miR-145 | IRS1 | Breast Cancer | Inhibitor | [43] |
| miR-206 | Met | Colorectal Cancer | Inhibitor | [44] |
| miR-378a-3p | TRAF1 | Hepatocellular Carcinoma | Promoter | [45] |
| miR-375 | PDGFC | Hepatocarcinoma | Inhibitor | [46] |
| miR-877-3p | FGF2 | Osteosarcoma | Inhibitor | [47] |
| miR-CT3 | VEGF-A | Osteosarcoma | Inhibitor | [48] |
| miR-495 | ATP7A | Esophageal Cancer | Inhibitor | [49] |
Table 2.
Targets and dysregulation of lncRNA associated with tumor angiogenesis.
| Basic Mechanisms | LncRNA | Target Genes | Cancer Types | The Role in Tumor Angiogenesis | References |
|---|---|---|---|---|---|
| Sponging miRNAs (lncRNA-miRNA-targeted gene) | LncRNA ZNRD1-AS1 | miR-942/TNS1 | Lung Cancer | Promoter | [66] |
| LINC00173.v1 | miR-511-5p/VEGF-A | Lung Squamous cell carcinoma | Promoter | [67] | |
| LncRNA PCAT6 | miR-4723-5p/VEGFR2 | TNBC | Promoter | [68] | |
| LncRNA NR2F1-AS1 | miRNA-338-3p/IGF-1 | Breast Cancer | Promoter | [69] | |
| LncRNA NORAD | miR-211-5p/FOXD1 | Hepatocellular Carcinoma Cells | Promoter | [70] | |
| LncRNA MYLK-AS1 | miR-424-5p/E2F7 | Hepatocellular Carcinoma | Promoter | [71] | |
| LncRNA miR503HG | miR-15b/PDCD4 | Hepatocellular Carcinoma | Inhibitor | [72] | |
| LncRNA CRART16 | miR-122-5p/FOS | Gastric Cancer | Promoter | [73] | |
| LncRNA NKX2-1-AS1 | miR-145-5p/SERPINE1 | Gastric Cancer | Promoter | [74] | |
| LncRNA H19 | miR-138/HIF-1α | Glioma | Promoter | [75] | |
| LncRNA H19 | miR-342/Wnt5a | Glioma | Promoter | [76] | |
| LncRNA DANCR | miR-145/VEGF | Ovarian Cancer | Promoter | [77] | |
| LncRNA LOC100129620 | miR-335-3p/CDK6 | Osteosarcoma | Promoter | [78] | |
| LncRNA HOTAIR | miR-126/EGFL7 | Renal Cell Carcinoma | Promoter | [79] | |
| LncRNA IGKJ2-MALLP2 | miR-1911-3p/p21 | Laryngeal Squamous Cell Carcinoma | Inhibitor | [80] | |
| Interacting (lncRNA- Proteins) | LncRNA SNHG5 | IGF2BP2 | Breast Cancer | Promoter | [81] |
| LncRNA RAB11B-AS1 | RNA Pol II | Breast Cancer | Promoter | [82] | |
| LncRNA PAARH | HIF-1α | Hepatocellular Carcinoma | Promoter | [83] | |
| LncRNA MAGI2-AS3 | HEY1 | Clear Cell Renal Cell Carcinoma | Inhibitor | [84] | |
| LncRNA RP11-536 K7.3 | SOX2 | colorectal cancer | Promoter | [85] | |
| LncRNA HITT | YB-1 | Colon Cancer | Inhibitor | [86] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Su, Z.; Li, W.; Lei, Z.; Hu, L.; Wang, S.; Guo, L. Regulation of Angiogenesis by Non-Coding RNAs in Cancer. Biomolecules 2024, 14, 60. https://doi.org/10.3390/biom14010060
AMA Style
Su Z, Li W, Lei Z, Hu L, Wang S, Guo L. Regulation of Angiogenesis by Non-Coding RNAs in Cancer. Biomolecules. 2024; 14(1):60. https://doi.org/10.3390/biom14010060
Chicago/Turabian StyleSu, Zhiyue, Wenshu Li, Zhe Lei, Lin Hu, Shengjie Wang, and Lingchuan Guo. 2024. "Regulation of Angiogenesis by Non-Coding RNAs in Cancer" Biomolecules 14, no. 1: 60. https://doi.org/10.3390/biom14010060
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.