

Exploring the Potential of the Model Cyanobacteria Synechococcus PCC 7002 and PCC 7942 for the Photoproduction of High-Value Terpenes: A Comparison with Synechocystis PCC 6803
 , , and
, , and 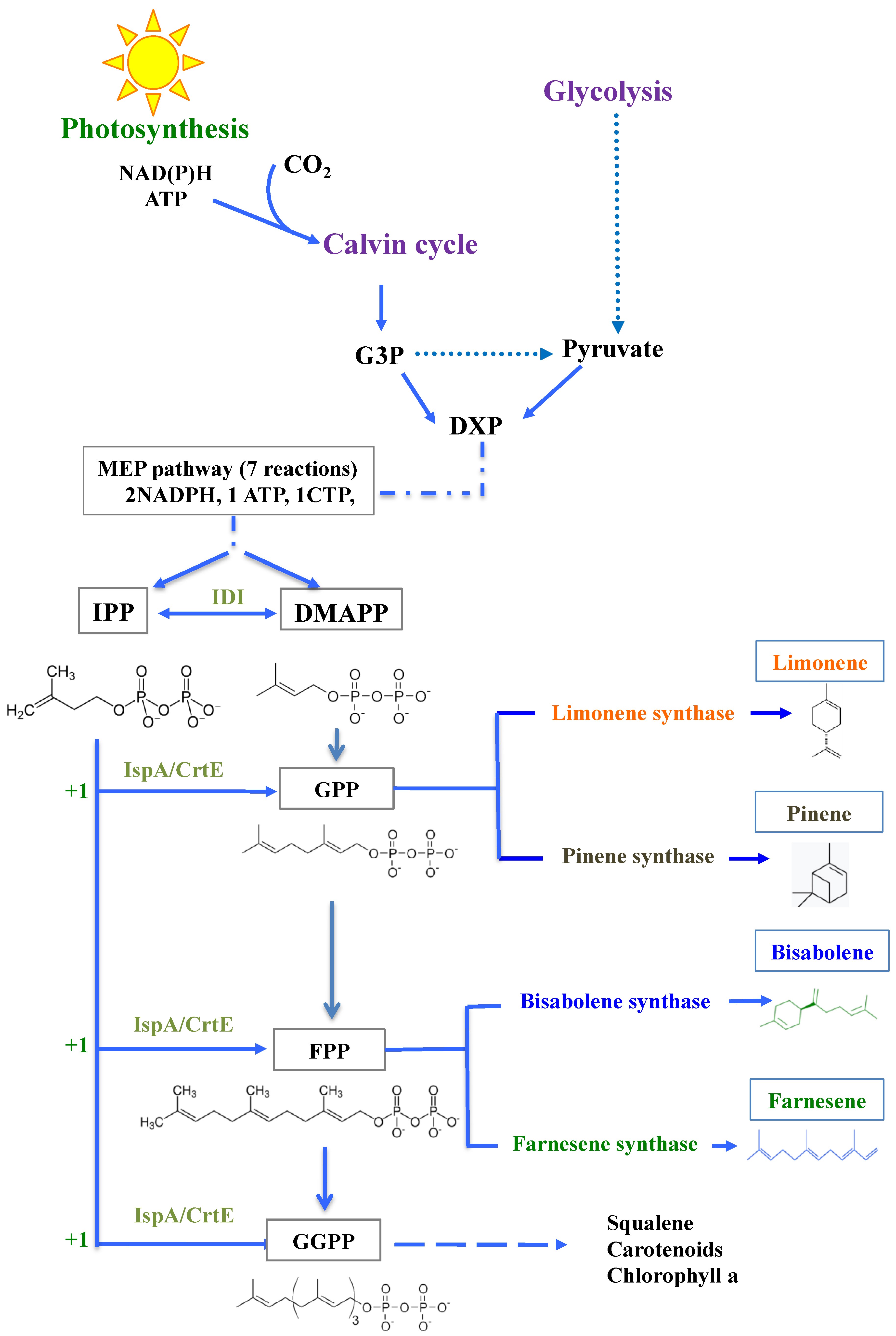{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Genetic Manipulations
2.3. Terpene Collection and Quantification by Gas Chromatography-Mass Spectrometry
3. Results and Discussion
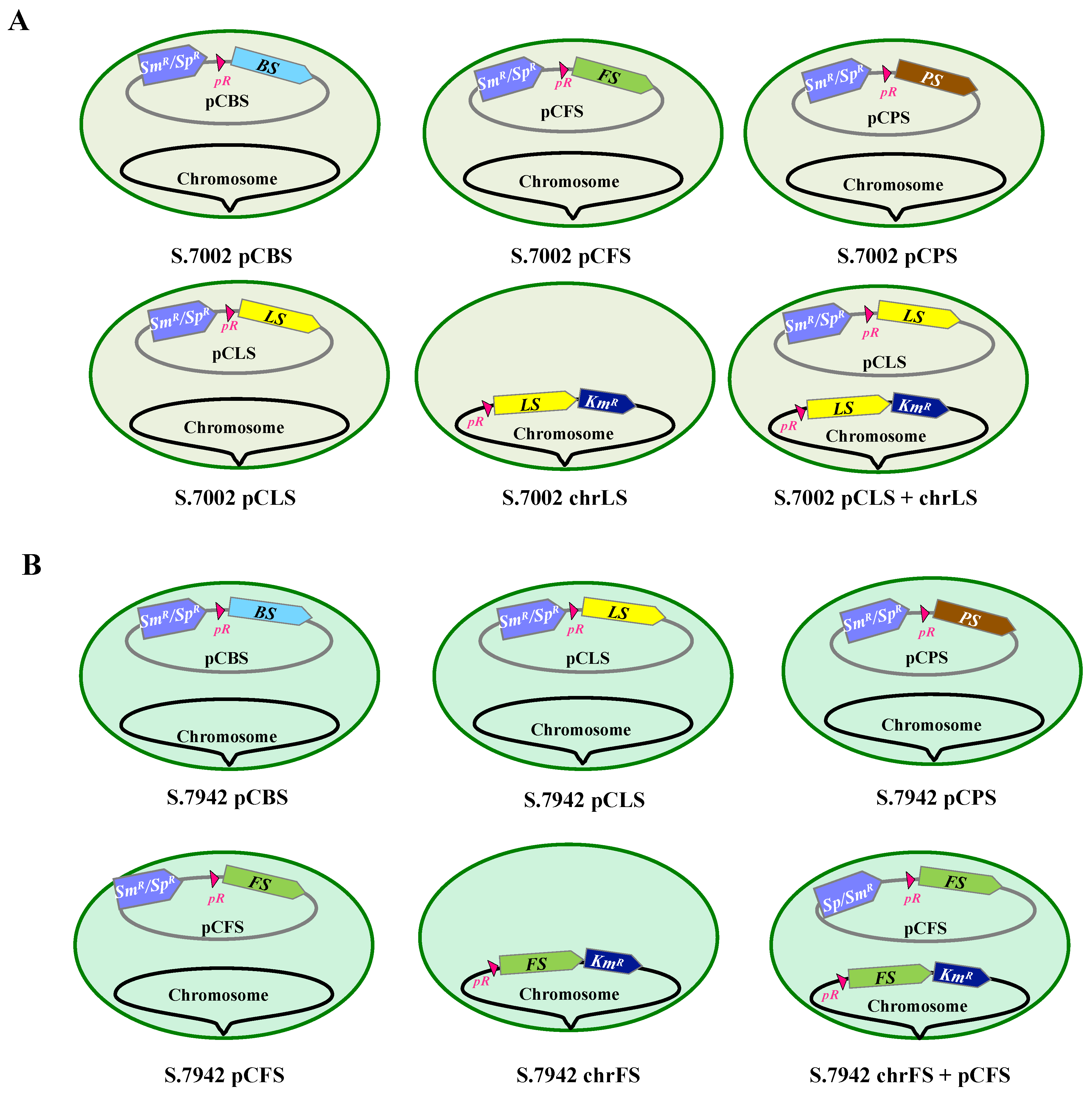
3.1. Construction of the Synechococcus PCC 7002 and the Synechococcus Elongatus PCC 7942 Strains Expressing the Studied Terpene Synthase Genes from the Strong Lambda-phage pR Promoter and Propagated in Either or Both a Neutral Chromosomal Site or a Replicative Plasmid
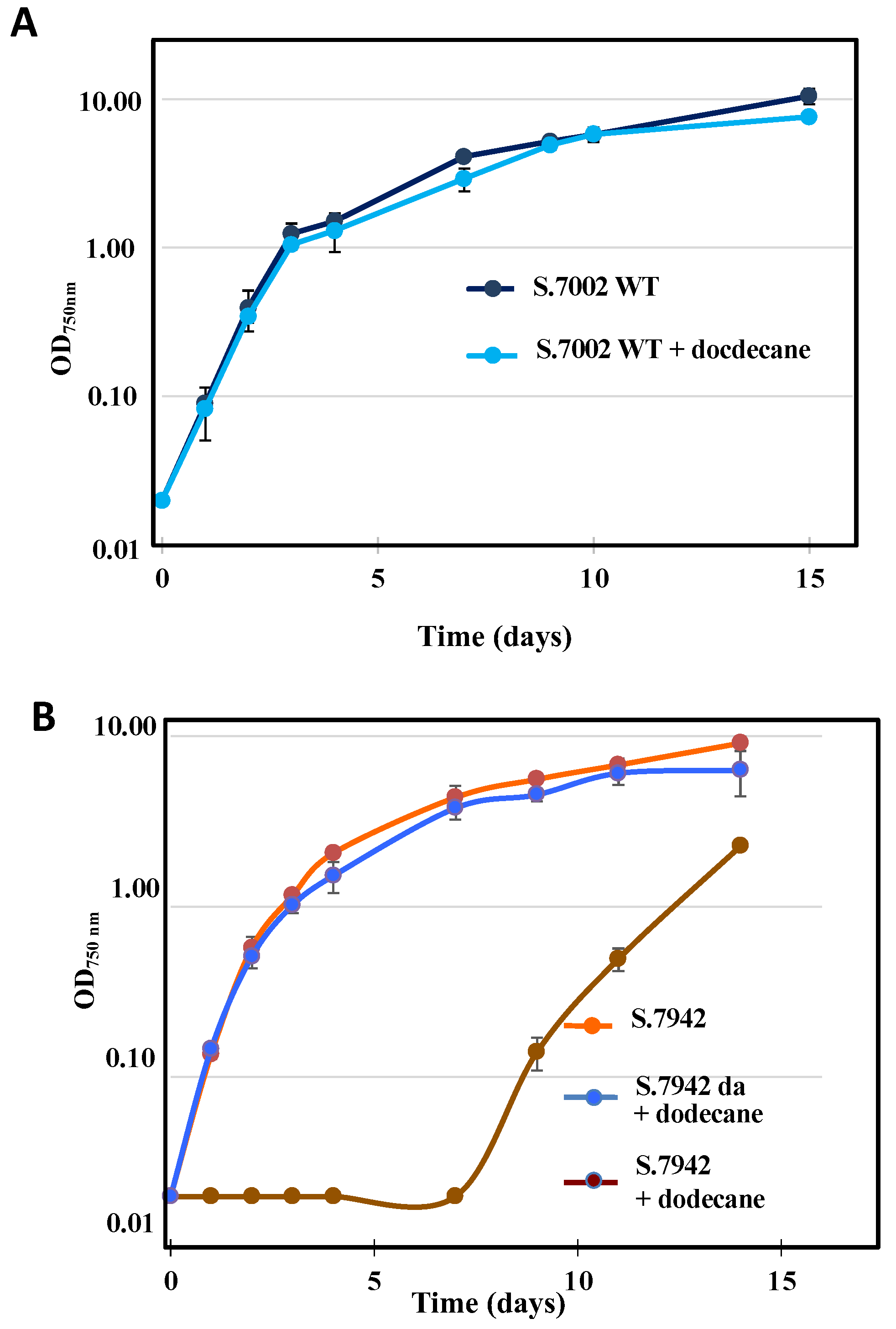
3.2. Unlike Synechococcus PCC 7002, the Growth of Synechococcus elongatus PCC 7942 Is Affected by the Dodecane Overlay Used to Trap Terpenes, but Dodecane-Adapted Strains Grow Healthy under Dodecane
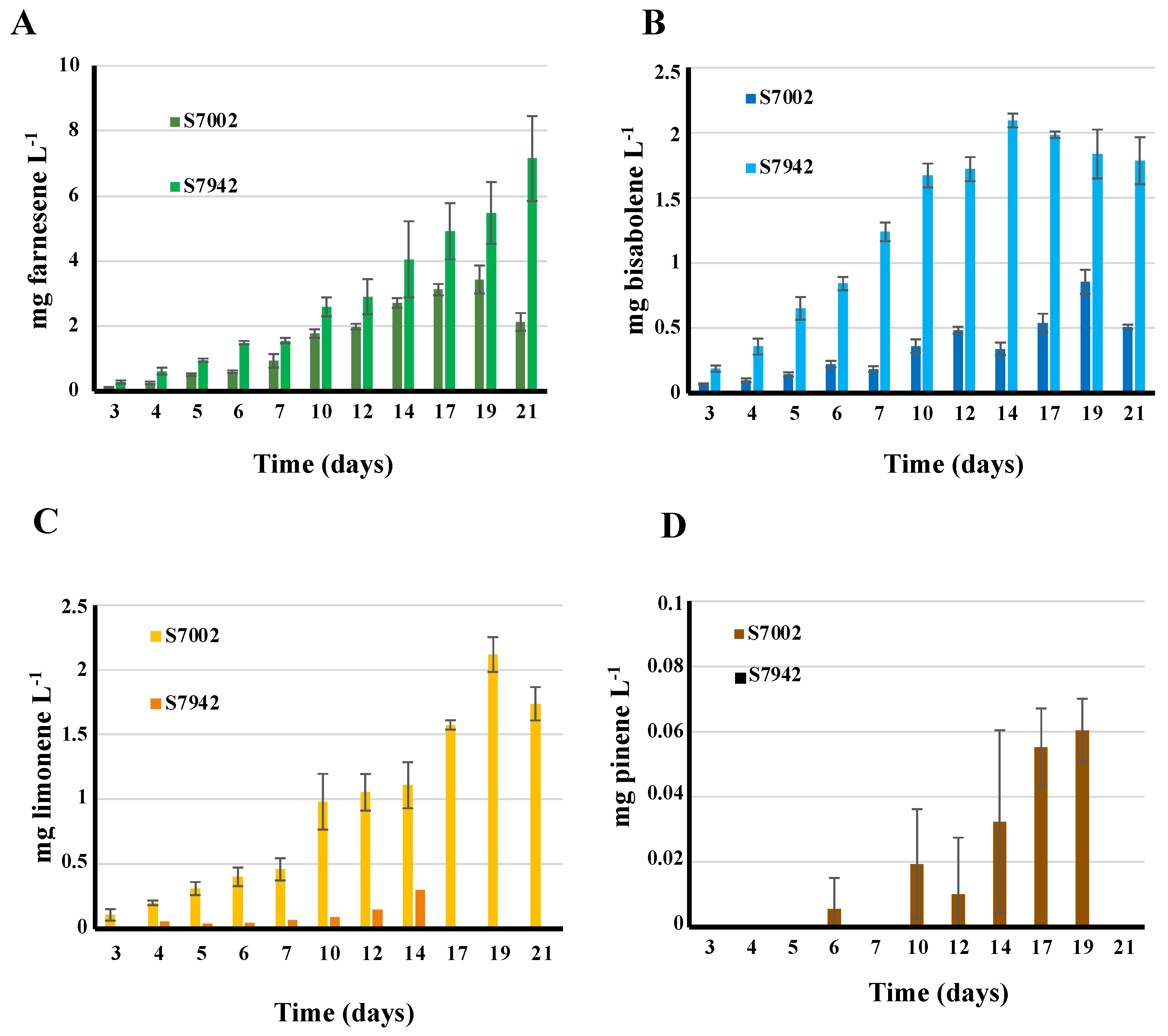
3.3. The First Report of Farnesene Production by Synechococcus PCC 7002: The Production Is Lower Than That of Synechococcus elongatus PCC 7942
3.4. First Report of Bisabolene Production by Synechococcus Elongatus PCC 7942: The Production Is Similar to That of Synechococcus PCC 7002
3.5. The Synechococcus PCC 7002 and Synechococcus elongatus PCC 7942 Strains Harboring the pCLS Plasmid Produce Limonene in This Order
3.6. The Synechococcus PCC 7002 and Synechococcus elongatus PCC 7942 Strains Harboring the pCPS Plasmid Produce Pinene Inefficiently
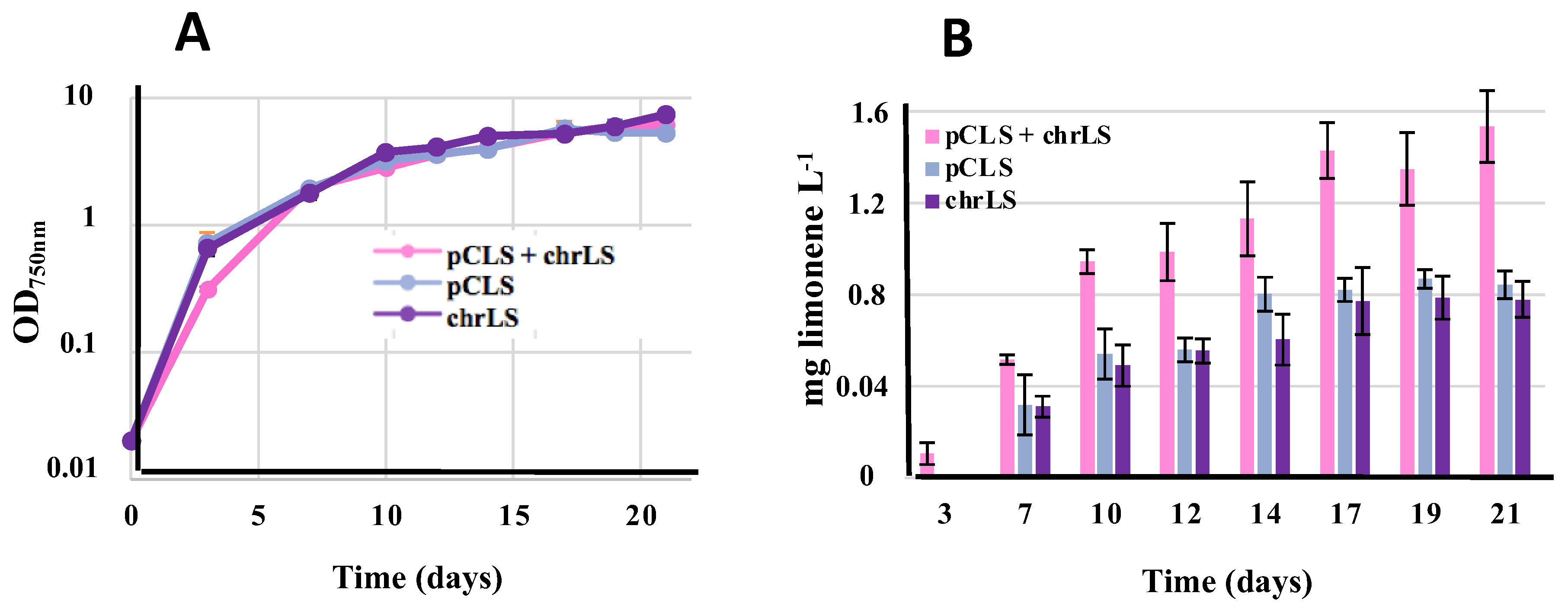
3.7. Comparison of Limonene Production in Synechococcus PCC 7002 Driven by the Recombinant LS Gene Propagated by Either or Both the pCLS Replicative Plasmid and the NS7002 Neutral Chromosomal Site
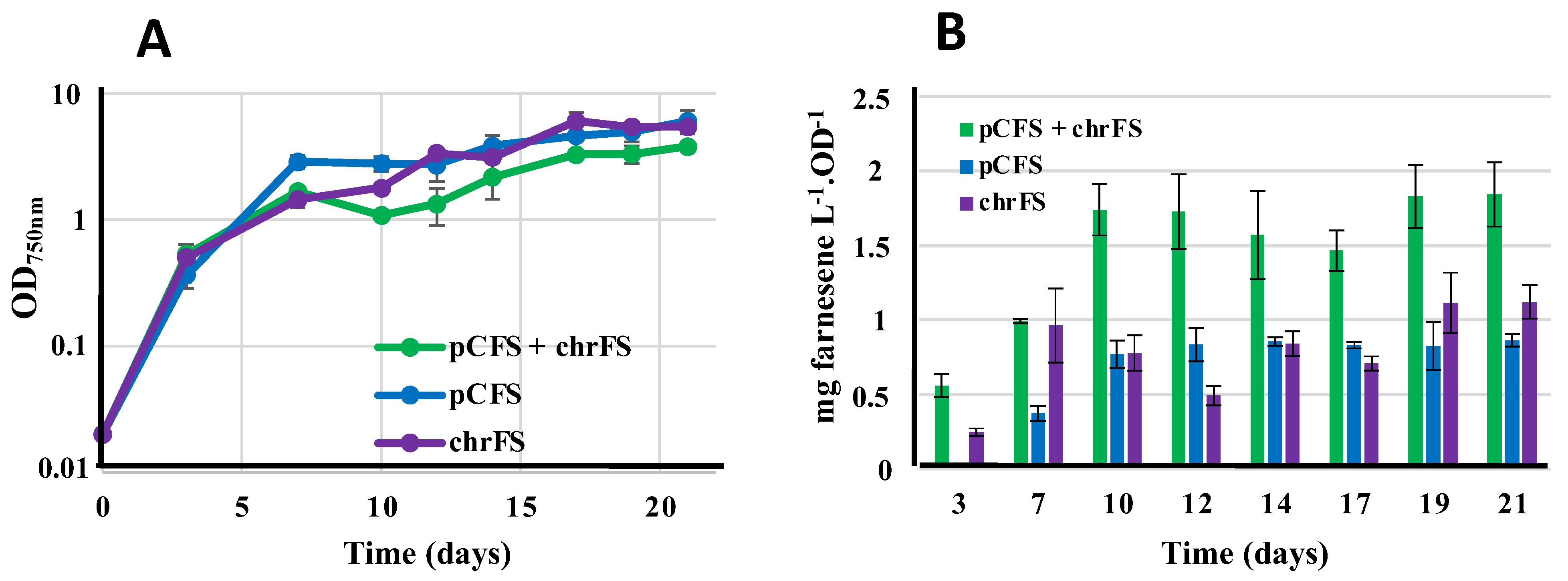
3.8. Comparison of Farnesene Production in Synechococcus Elongatus PCC 7942 Driven by the Recombinant FS Gene Propagated by Either or Both the pCFS Replicative Plasmid and the NSI7942 Neutral Chromosomal Site
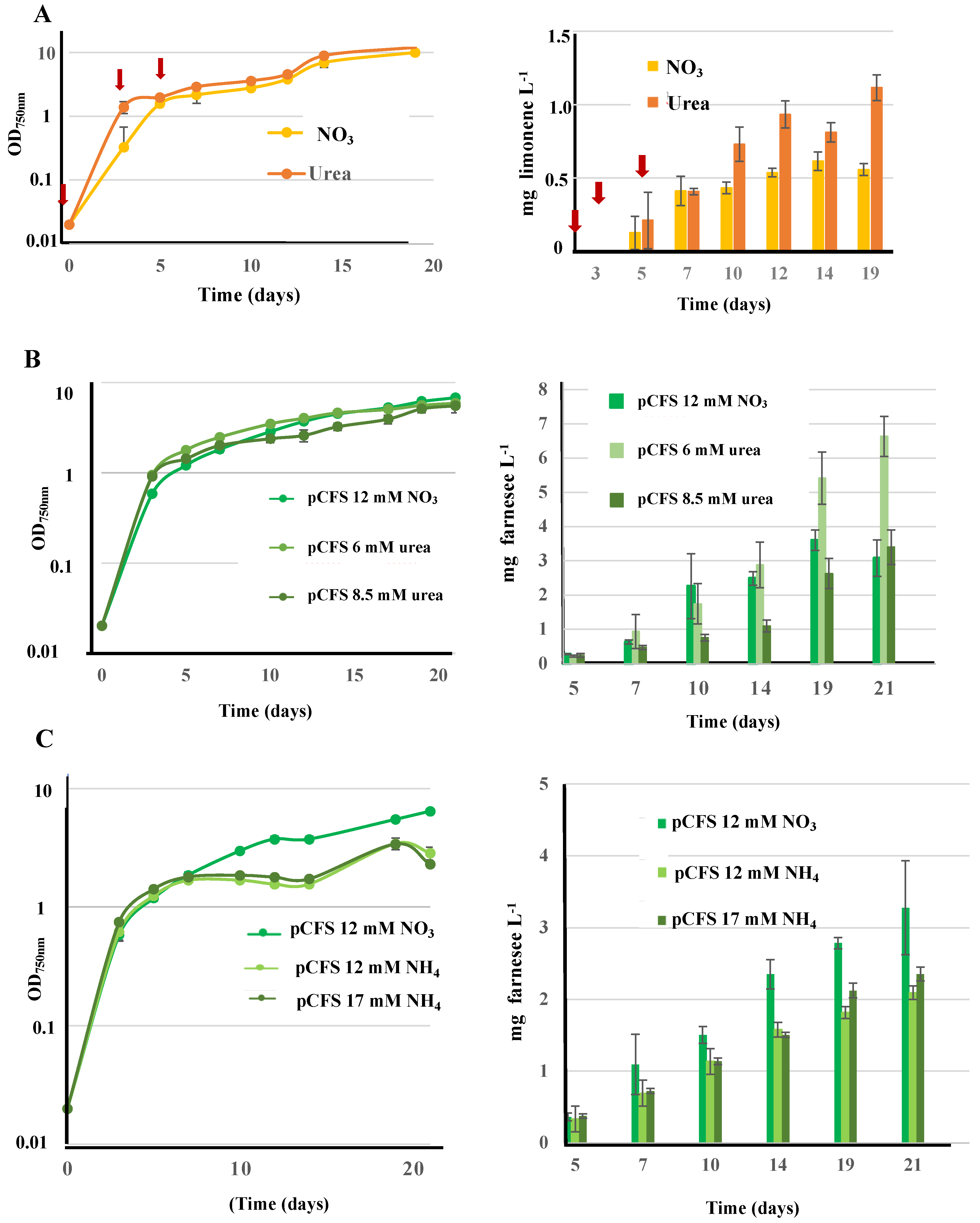
3.9. Influence of Growth Conditions on Terpene Production: First Report That Synechococcus PCC 7002 Cultivated on Urea Produces Terpenes More Efficiently Than Cultures Growing on Nitrate or Ammonium in This Order
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, P.C.; Pakrasi, H.B. Engineering cyanobacteria for production of terpenoids. Planta 2019, 249, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Rautela, A.; Kumar, S. Engineering plant family TPS into cyanobacterial host for terpenoids production. Plant Cell Rep. 2022, 41, 1791–1803. [Google Scholar] [CrossRef] [PubMed]
- Cassier-Chauvat, C.; Blanc-Garin, V.; Chauvat, F. Genetic, Genomics, and Responses to Stresses in Cyanobacteria: Biotechnological Implications. Genes 2021, 12, 500. [Google Scholar] [CrossRef] [PubMed]
- Veaudor, T.; Cassier-Chauvat, C.; Chauvat, F. Genomics of Urea Transport and Catabolism in Cyanobacteria: Biotechnological Implications. Front. Microbiol. 2019, 10, 2052. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, M.; Bryant, D.A. Acclimation of the global transcriptome of the cyanobacterium Synechococcus sp. strain PCC 7002 to nutrient limitations and different nitrogen sources. Front. Microbiol. 2012, 11, 354. [Google Scholar] [CrossRef] [Green Version]
- Veaudor, T.; Blanc-Garin, V.; Chenebault, C.; Diaz-Santos, E.; Sassi, J.F.; Cassier-Chauvat, C.; Chauvat, F. Recent advances in the photoautotrophic metabolism of cyanobacteria: Biotechnological implications. Life 2020, 10, 71. [Google Scholar] [CrossRef]
- Cassier-Chauvat, C.; Veaudor, T.; Chauvat, F. Comparative genomics of DNA recombination and repair in cyanobacteria: Biotechnological implications. Front. Microbiol. 2016, 7, 1809. [Google Scholar] [CrossRef] [Green Version]
- Davies, F.K.; Work, V.H.; Beliaev, A.S.; Posewitz, M.C. Engineering Limonene and Bisabolene Production in Wild Type and a Glycogen-Deficient Mutant of Synechococcus sp. PCC 7002. Front. Bioeng. Biotechnol. 2014, 2, 21. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Zhu, L.; Li, T.; Zhu, L.; Ye, Z.; Zhang, D. Photosynthetic Conversion of CO2 Into Pinene Using Engineered Synechococcus sp. PCC 7002. Front. Bioeng. Biotechnol. 2021, 9, 779437. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, J.; Lee, S.-M.; Um, Y.; Kim, Y.; Sim, S.J.; Choi, J.-I.; Woo, H.M. Direct Conversion of CO2 to α-Farnesene Using Metabolically Engineered Synechococcus elongatus PCC 7942. J. Agric. Food Chem. 2017, 60, 10424–10428. [Google Scholar] [CrossRef]
- Pattharaprachayakul, N.; Lee, H.J.; Incharoensakdi, A.; Woo, H.M. Evolutionary Engineering of Cyanobacteria to Enhance the Production of α-Farnesene from CO2. J. Agric. Food Chem. 2019, 67, 13658–13664. [Google Scholar] [CrossRef]
- Lee, H.J.; Choi, J.I.; Woo, H.M. Biocontainment of Engineered Synechococcus elongatus PCC 7942 for Photosynthetic Production of α-Farnesene from CO2. J. Agric. Food Chem. 2021, 69, 698–703. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, W.; Xin, C.; Zheng, Y.; Cheng, Y.; Sun, S.; Li, R.; Zhu, X.-G.; Dai, S.Y.; Rentzepis, P.M.; et al. Enhanced limonene production in cyanobacteria reveals photosynthesis limitations. Proc. Natl. Acad. Sci. USA 2016, 113, 14225–14230. [Google Scholar] [CrossRef] [Green Version]
- Shinde, S.; Singapuri, S.; Jiang, Z.; Long, B.; Wilcox, D.; Klatt, C.; Jones, J.A.; Yuan, J.S.; Wang, X. Thermodynamics contributes to high limonene productivity in cyanobacteria. Metab. Eng. Commun. 2022, 14, e00193. [Google Scholar] [CrossRef] [PubMed]
- Mermet-Bouvier, P.; Chauvat, F. A conditional expression vector for the cyanobacteria Synechocystis sp. strains PCC6803 and PCC6714 or Synechococcus sp. strains PCC7942 and PCC6301. Curr. Microbiol. 1994, 28, 145–148. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Domain, F.; Houot, L.; Chauvat, F.; Cassier-Chauvat, C. Function and regulation of the cyanobacterial genes lexA, recA and ruvB: LexA is critical to the survival of cells facing inorganic carbon starvation. Mol. Microbiol. 2004, 53, 65–80. [Google Scholar] [CrossRef]
- Stevens, S.E.; Pat Paterson, C.O.; Myers, J. The production of hydrogen peroxide by blue-green algae: A survey. J. Phycol. 1973, 9, 427–430. [Google Scholar] [CrossRef]
- Mermet-Bouvier, P.; Cassier-Chauvat, C.; Marraccini, P.; Chauvat, F. Transfer and replication of RSF1010-derived plasmids in several cyanobacteria of the genera Synechocystis and Synechococcus. Curr. Microbiol. 1993, 27, 323–327. [Google Scholar] [CrossRef]
- Blanc-Garin, V.; Chenebault, C.; Diaz-Santos, E.; Vincent, M.; Sassi, J.F.; Cassier-Chauvat, C.; Chauvat, F. Exploring the potential of the model cyanobacterium Synechocystis PCC 6803 for the photosynthetic production of various high-value terpenes. Biotechnol. Biofuels 2022, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Chauvat, F.; Astier, C.; Vedel, F.; Joset-Espardellier, F. Transformation in the cyanobacterium Synechococcus R2: Improvement of efficiency; Role of the pUH24 plasmid. Mol. Gen. Genet. 1983, 191, 39–45. [Google Scholar] [CrossRef]
- Labarre, J.; Chauvat, F.; Thuriaux, P. Insertional mutagenesis by random cloning of antibiotic resistance genes into the genome of the cyanobacterium Synechocystis strain PCC 6803. J. Bacteriol. 1989, 171, 3449–3457. [Google Scholar] [CrossRef] [Green Version]
- Chenebault, C.; Diaz-Santos, E.; Kammerscheit, X.; Görgen, S.; Ilioaia, C.; Streckaite, S.; Gall, A.; Robert, B.; Marcon, E.; Buisson, D.-A.; et al. A Genetic Toolbox for the New Model Cyanobacterium Cyanothece PCC 7425: A Case Study for the Photosynthetic Production of Limonene. Front. Microbiol. 2020, 11, 586–601. [Google Scholar] [CrossRef] [PubMed]
- Veaudor, T.; Cassier-Chauvat, C.; Chauvat, F. Overproduction of the cyanobacterial hydrogenase and selection of a mutant thriving on urea, as a possible step towards the future production of hydrogen coupled with water treatment. PLoS ONE 2018, 13, e0198836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc-Garin, V.; Veaudor, T.; Sétif, P.; Gontero, B.; Lemaire, S.D.; Chauvat, F.; Cassier-Chauvat, C. First in vivo analysis of the regulatory protein CP12 of the model cyanobacterium Synechocystis PCC 6803: Biotechnological implications. Front. Plant Sci. 2022, 13, 999672. [Google Scholar] [CrossRef]
- Vogel, A.I.M.; Lale, R.; Hohmann-Marriott, M.F. Streamlining recombination-mediated genetic engineering by validating three neutral integration sites in Synechococcus sp. PCC 7002. J. Biol. Eng. 2017, 11, 19. [Google Scholar] [CrossRef] [Green Version]
- Gibson, D.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchinson, C.A.; Smith, H. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Bustos, S.A.; Golden, S.S. Light-regulated expression of the psbD gene family in Synecbococcus sp. strain PCC 7942: Evidence for the role of duplicated psbD genes in cyanobacteria. Mol. Gen. Genet. 1992, 232, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Betterle, N.; Melis, A. Photosynthetic generation of heterologous terpenoids in cyanobacteria. Biotechnol. Bioeng. 2019, 116, 2041–2051. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chenebault, C.; Blanc-Garin, V.; Vincent, M.; Diaz-Santos, E.; Goudet, A.; Cassier-Chauvat, C.; Chauvat, F. Exploring the Potential of the Model Cyanobacteria Synechococcus PCC 7002 and PCC 7942 for the Photoproduction of High-Value Terpenes: A Comparison with Synechocystis PCC 6803. Biomolecules 2023, 13, 504. https://doi.org/10.3390/biom13030504
Chenebault C, Blanc-Garin V, Vincent M, Diaz-Santos E, Goudet A, Cassier-Chauvat C, Chauvat F. Exploring the Potential of the Model Cyanobacteria Synechococcus PCC 7002 and PCC 7942 for the Photoproduction of High-Value Terpenes: A Comparison with Synechocystis PCC 6803. Biomolecules. 2023; 13(3):504. https://doi.org/10.3390/biom13030504
Chicago/Turabian StyleChenebault, Célia, Victoire Blanc-Garin, Marine Vincent, Encarnación Diaz-Santos, Amélie Goudet, Corinne Cassier-Chauvat, and Franck Chauvat. 2023. "Exploring the Potential of the Model Cyanobacteria Synechococcus PCC 7002 and PCC 7942 for the Photoproduction of High-Value Terpenes: A Comparison with Synechocystis PCC 6803" Biomolecules 13, no. 3: 504. https://doi.org/10.3390/biom13030504