
Calcium Binds to Transthyretin with Low Affinity

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Expression and Purification
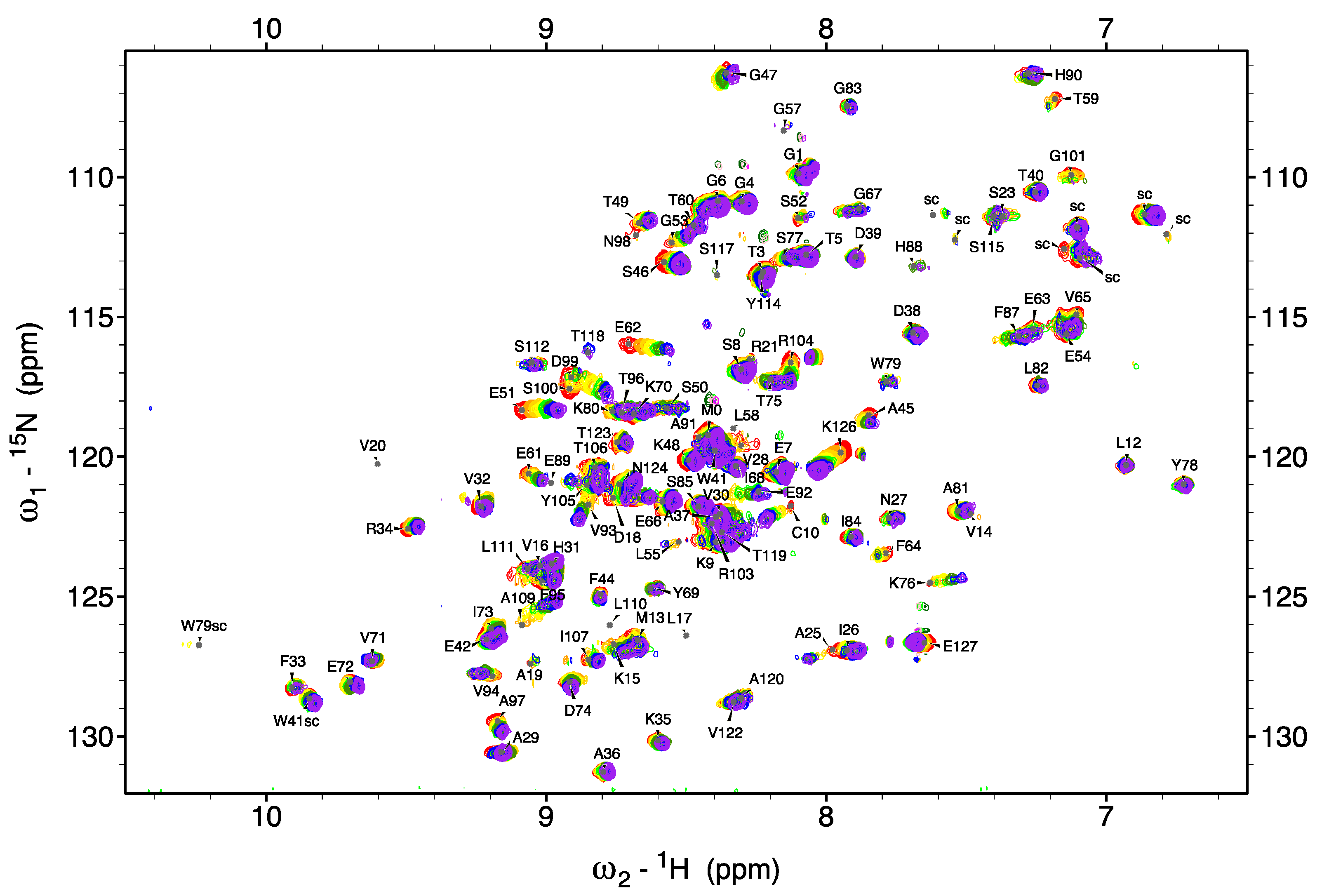
2.2. NMR Spectroscopy
2.3. Electrostatic Calculation
2.4. Molecular Dynamics (MD) Simulations
2.5. Aggregation
3. Results
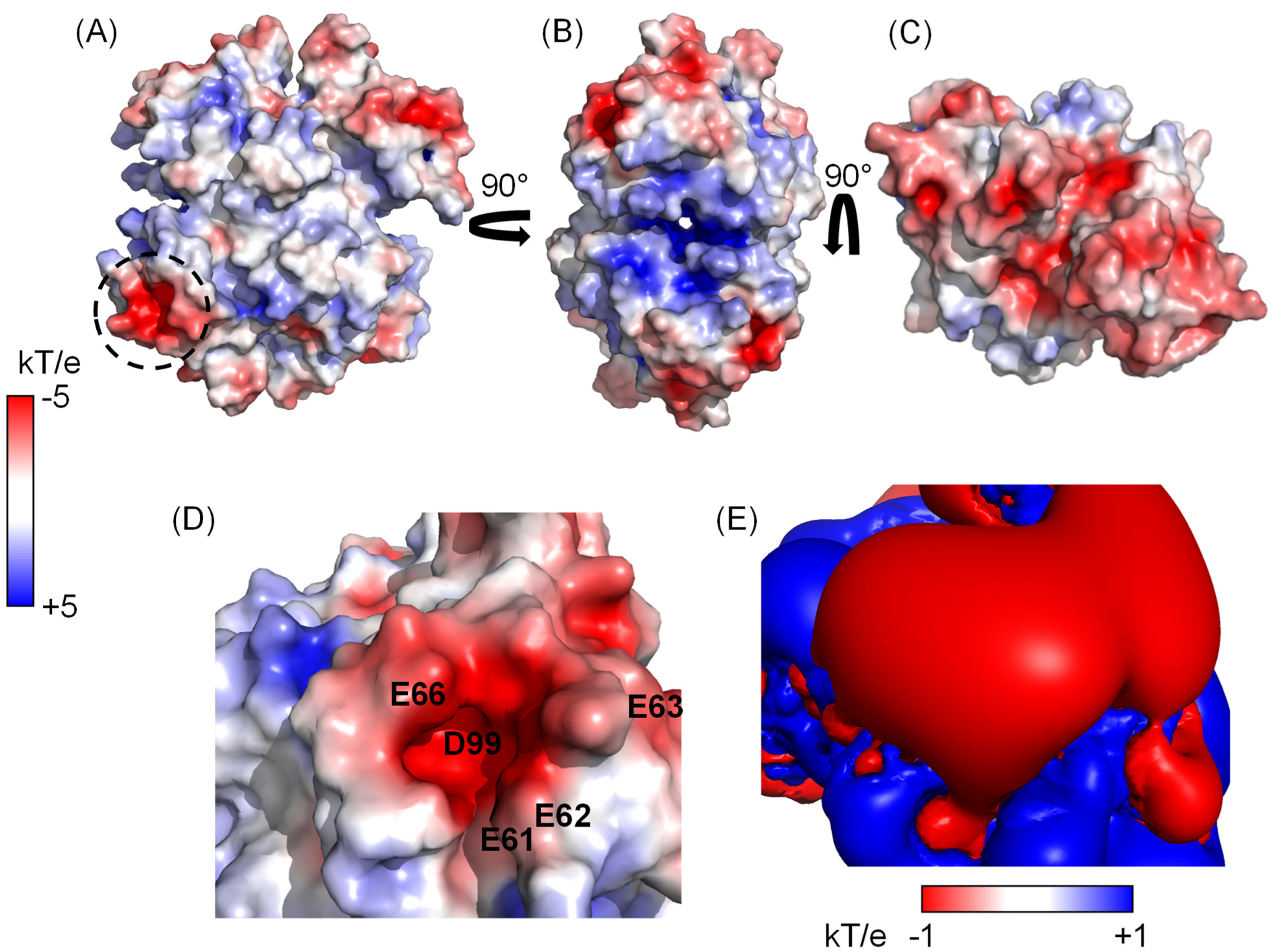
3.1. Electrostatics of TTR
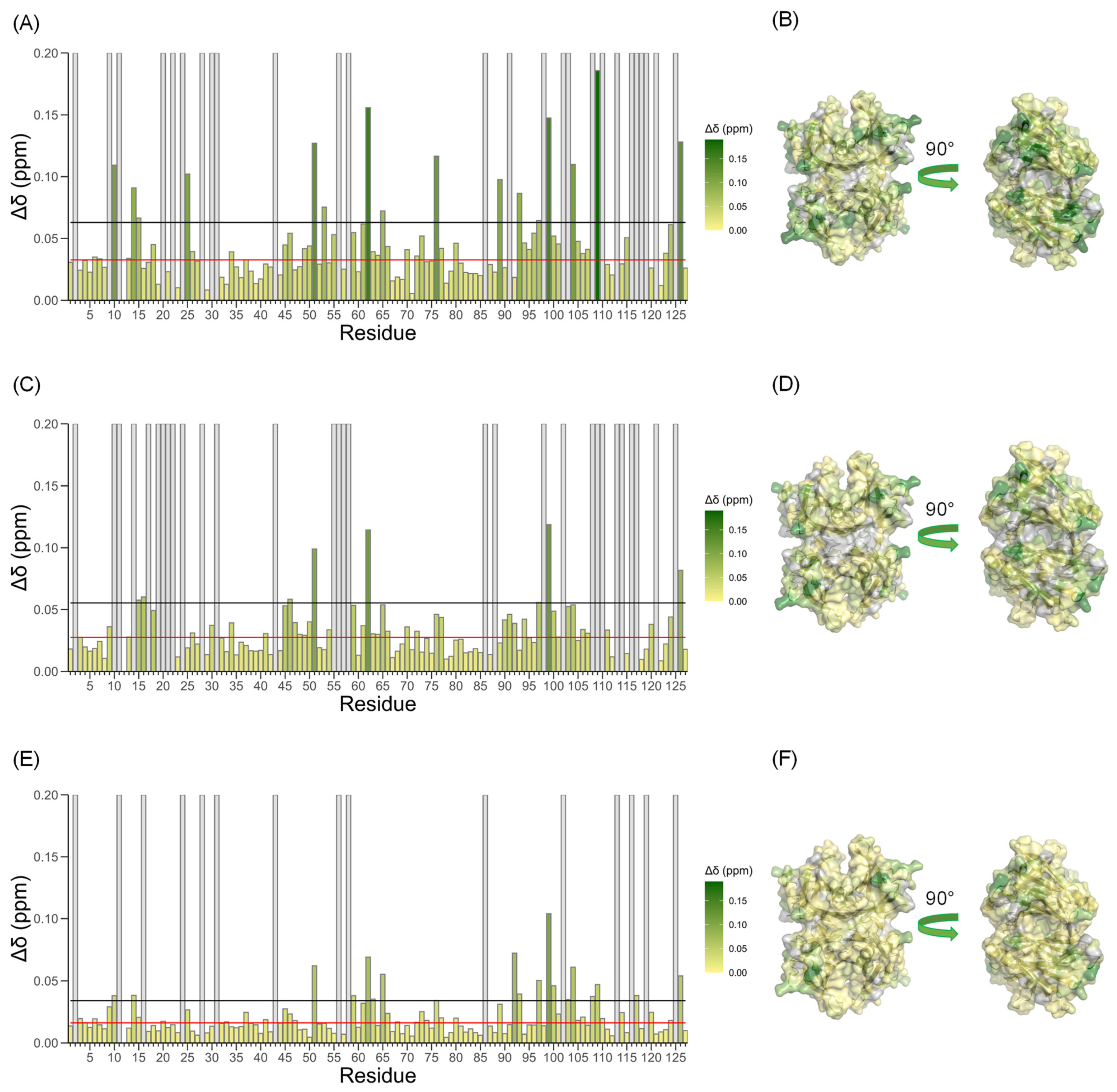
3.2. Does TTR Bind Ca2+?
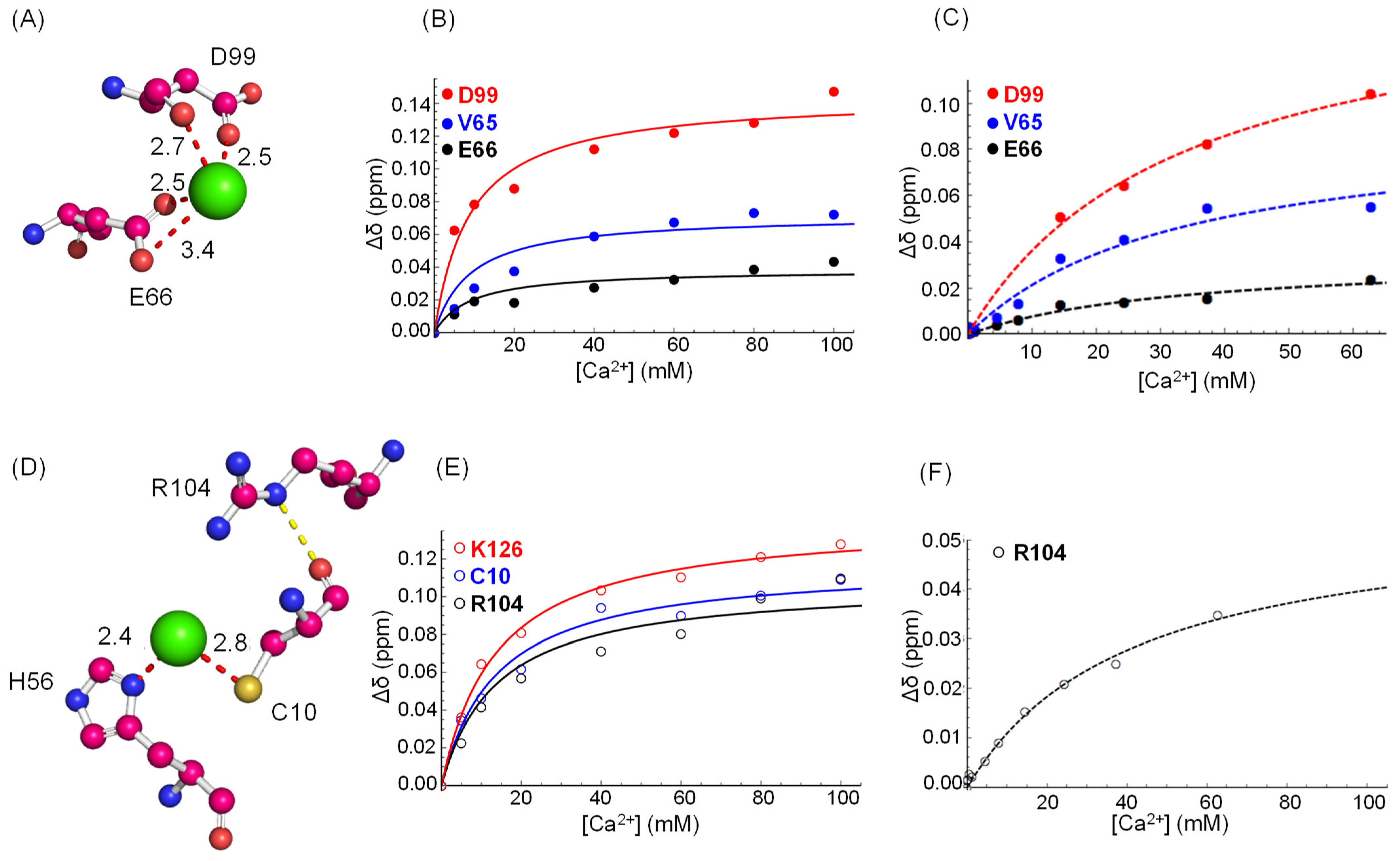
3.3. Binding Specificity
3.4. Molecular Dynamics Simulations
3.5. Ca2+ Favors TTR Aggregation under Physiological pH
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Westermark, P.; Sletten, K.; Johansson, B.; Cornwell, G.G. Fibril in Senile Systemic Amyloidosis Is Derived from Normal Transthyretin. Proc. Natl. Acad. Sci. USA 1990, 87, 2843–2845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarzman, A.L.; Gregori, L.; Vitek, M.P.; Lyubski, S.; Strittmatter, W.J.; Enghilde, J.J.; Bhasin, R.; Silverman, J.; Weisgraber, K.H.; Coyle, P.K. Tr ansthyretin Sequesters Amyloid Beta Protein and Prevents Amyloid Formation. Proc. Natl. Acad. Sci. USA 1994, 91, 8368–8372. [Google Scholar] [CrossRef] [Green Version]
- Colon, W.; Kelly, J.W. Partial Denaturation of Transthyretin Is Sufficient for Amyloid Fibril Formation in Vitro. Biochemistry 1992, 31, 8654–8660. [Google Scholar] [CrossRef]
- Thylen, C.; Wahlqvist, J.; Haettner, E.; Sandgren, O.; Holmgren, G.; Lundgren, E. Modifications of Transthyretin in Amyloid Fibrils: Analysis of Amyloid from Homozygous and Heterozygous Individuals with the Met30 Mutation. EMBO J. 1993, 12, 743–748. [Google Scholar] [CrossRef]
- Mangione, P.P.; Porcari, R.; Gillmore, J.D.; Pucci, P.; Monti, M.; Porcari, M.; Giorgetti, S.; Marchese, L.; Raimondi, S.; Serpell, L.C.; et al. Proteolytic Cleavage of Ser52Pro Variant Transthyretin Triggers Its Amyloid Fibrillogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, 1539–1544. [Google Scholar] [CrossRef] [Green Version]
- Marcoux, J.; Mangione, P.P.; Porcari, R.; Degiacomi, M.T.; Verona, G.; Taylor, G.W.; Giorgetti, S.; Raimondi, S.; Sanglier-Cianférani, S.; Benesch, J.L.; et al. A Novel Mechano-Enzymatic Cleavage Mechanism Underlies Transthyretin Amyloidogenesis. EMBO Mol. Med. 2015, 7, 1337–1349. [Google Scholar] [CrossRef]
- Mangione, P.P.; Verona, G.; Corazza, A.; Marcoux, J.; Canetti, D.; Giorgetti, S.; Raimondi, S.; Stoppini, M.; Esposito, M.; Relini, A.; et al. Plasminogen Activation Triggers Transthyretin Amyloidogenesis in Vitro. J. Biol. Chem. 2018, 293, 14192–14199. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, E.; Kędracka-Krok, S.; Bystranowska, D.; Ptak, M.; Wiak, K.; Wygralak, Z.; Jankowska, U.; Ożyhar, A. Destabilisation of the Structure of Transthyretin Is Driven by Ca2+. Int. J. Biol. Macromol. 2021, 166, 409–423. [Google Scholar] [CrossRef]
- Carafoli, E.; Krebs, J. Why Calcium? How Calcium Became the Best Communicator. J. Biol. Chem. 2016, 291, 20849–20857. [Google Scholar] [CrossRef] [Green Version]
- Bushinsky, D.A.; Monk, R.D. Calcium. Lancet 1998, 352, 306–311. [Google Scholar] [CrossRef]
- Johnson, P.C.; Smith, W.O.; Wulff, B. Calcium-45 Binding by Human Prealbumin. J. Appl. Physiol. 1959, 14, 859–860. [Google Scholar] [CrossRef] [PubMed]
- Kragh-Hansen, U.; Vorum, H. Quantitative Analyses of the Interaction between Calcium Ions and Human Serum Albumin. Clin. Chem. 1993, 39, 202–208. [Google Scholar] [CrossRef]
- Yokoyama, T.; Kosaka, Y.; Mizuguchi, M. Crystal Structures of Human Transthyretin Complexed with Glabridin. J. Med. Chem. 2014, 57, 1090–1096. [Google Scholar] [CrossRef] [PubMed]
- Pervushin, K.; Riek, R.; Wider, G.; Wüthrich, K. Attenuated T2 Relaxation by Mutual Cancellation of Dipole–Dipole Coupling and Chemical Shift Anisotropy Indicates an Avenue to NMR Structures of Very Large Biological Macromolecules in Solution. Proc. Natl. Acad. Sci. USA 1997, 94, 12366–12371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulder, F.A.A.; Schipper, D.; Bott, R.; Boelens, R. Altered Flexibility in the Substrate-Binding Site of Related Native and Engineered High-Alkaline Bacillus Subtilisins11Edited by P. E. Wright. J. Mol. Biol. 1999, 292, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Tonelli, M.; Markley, J.L. NMRFAM-SPARKY: Enhanced Software for Biomolecular NMR Spectroscopy. Bioinformatics 2015, 31, 1325–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, J.A.; Steinrauf, L.K.; Braden, B.C.; Liepnieks, J.; Benson, M.D.; Holmgren, G.; Sandgren, O.; Steen, L. The X-Ray Crystal Structure Refinements of Normal Human Transthyretin and the Amyloidogenic Val-30–>Met Variant to 1.7-A Resolution. J. Biol. Chem. 1993, 268, 2416–2424. [Google Scholar] [CrossRef]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K.; et al. Improvements to the APBS Biomolecular Solvation Software Suite. Protein Sci. 2018, 27, 112–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yee, A.W.; Moulin, M.; Breteau, N.; Haertlein, M.; Mitchell, E.P.; Cooper, J.B.; Boeri Erba, E.; Forsyth, V.T. Impact of Deuteration on the Assembly Kinetics of Transthyretin Monitored by Native Mass Spectrometry and Implications for Amyloidoses. Angew. Chem. Int. Ed. 2016, 55, 9292–9296. [Google Scholar] [CrossRef]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and Better Reference Data for Improved All-Atom Structure Validation. Protein Sci. 2018, 27, 293–315. [Google Scholar] [CrossRef]
- Rodrigues, J.P.G.L.M.; Teixeira, J.M.C.; Trellet, M.; Bonvin, A.M.J.J. Pdb-Tools: A Swiss Army Knife for Molecular Structures. F1000Research 2018, 7, 1961. [Google Scholar] [CrossRef]
- Dolinsky, T.J.; Nielsen, J.E.; McCammon, J.A.; Baker, N.A. PDB2PQR: An Automated Pipeline for the Setup of Poisson-Boltzmann Electrostatics Calculations. Nucleic Acids Res. 2004, 32, W665–W667. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.C.; Hardy, D.J.; Maia, J.D.C.; Stone, J.E.; Ribeiro, J.V.; Bernardi, R.C.; Buch, R.; Fiorin, G.; Hénin, J.; Jiang, W.; et al. Scalable Molecular Dynamics on CPU and GPU Architectures with NAMD. J. Chem. Phys. 2020, 153, 044130. [Google Scholar] [CrossRef]
- MacKerell, A.D.; Bashford, D.; Bellott, M.; Dunbrack, R.L.; Evanseck, J.D.; Field, M.J.; Fischer, S.; Gao, J.; Guo, H.; Ha, S.; et al. All-Atom Empirical Potential for Molecular Modeling and Dynamics Studies of Proteins. J. Phys. Chem. B 1998, 102, 3586–3616. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of Simple Potential Functions for Simulating Liquid Water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Project, E.; Nachliel, E.; Gutman, M. The Dynamics of Ca2+ Ions within the Solvation Shell of Calbindin D9k. PLoS ONE 2011, 6, e14718. [Google Scholar] [CrossRef] [Green Version]
- Protein NMR Spectroscopy—2nd Edition. Available online: https://www.elsevier.com/books/protein-nmr-spectroscopy/cavanagh/978-0-12-164491-8 (accessed on 24 February 2022).
- Grzesiek, S.; Anglister, J.; Ren, H.; Bax, A. Carbon-13 Line Narrowing by Deuterium Decoupling in Deuterium/Carbon-13/Nitrogen-15 Enriched Proteins. Application to Triple Resonance 4D J Connectivity of Sequential Amides. J. Am. Chem. Soc. 1993, 115, 4369–4370. [Google Scholar] [CrossRef]
- Corazza, A.; Verona, G.; Waudby, C.A.; Mangione, P.P.; Bingham, R.; Uings, I.; Canetti, D.; Nocerino, P.; Taylor, G.W.; Pepys, M.B.; et al. Binding of Monovalent and Bivalent Ligands by Transthyretin Causes Different Short- and Long-Distance Conformational Changes. J. Med. Chem. 2019, 62, 8274–8283. [Google Scholar] [CrossRef] [Green Version]
- Williamson, M.P. Using Chemical Shift Perturbation to Characterise Ligand Binding. Prog. Nucl. Magn. Reson. Spectrosc. 2013, 73, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kukic, P.; O’Meara, F.; Hewage, C.; Erik Nielsen, J. Coupled Effect of Salt and PH on Proteins Probed with NMR Spectroscopy. Chem. Phys. Lett. 2013, 579, 114–121. [Google Scholar] [CrossRef]
- Castro-Rodrigues, A.F.; Gales, L.; Saraiva, M.J.; Damas, A.M. Structural Insights into a Zinc-Dependent Pathway Leading to Leu55Pro Transthyretin Amyloid Fibrils. Acta Crystallogr. Sect. D 2011, 67, 1035–1044. [Google Scholar] [CrossRef]
- Wilkinson-White, L.E.; Easterbrook-Smith, S.B. Characterization of the Binding of Cu(II) and Zn(II) to Transthyretin: Effects on Amyloid Formation. Biochemistry 2007, 46, 9123–9132. [Google Scholar] [CrossRef]
- Palmieri, L.D.C.; Lima, L.M.T.; Freire, J.B.; Bleicher, L.; Polikarpov, I.; Almeida, F.C.; Foguel, D. Novel Zn2+-Binding Sites in Human Transthyretin: Implications for Amyloidogenesis and Retinol-Binding Protein Recognition. J. Biol. Chem. 2010, 285, 31731–31741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciccone, L.; Fruchart-Gaillard, C.; Mourier, G.; Savko, M.; Nencetti, S.; Orlandini, E.; Servent, D.; Stura, E.A.; Shepard, W. Copper Mediated Amyloid-β Binding to Transthyretin. Sci. Rep. 2018, 8, 13744. [Google Scholar] [CrossRef] [PubMed]
- Ghadami, S.A.; Chia, S.; Ruggeri, F.S.; Meisl, G.; Bemporad, F.; Habchi, J.; Cascella, R.; Dobson, C.M.; Vendruscolo, M.; Knowles, T.P.J.; et al. Transthyretin Inhibits Primary and Secondary Nucleations of Amyloid-β Peptide Aggregation and Reduces the Toxicity of Its Oligomers. Biomacromolecules 2020, 21, 1112–1125. [Google Scholar] [CrossRef]
- Zhang, Y.; Cremer, P.S. Interactions between Macromolecules and Ions: The Hofmeister Series. Curr. Opin. Chem. Biol. 2006, 10, 658–663. [Google Scholar] [CrossRef]
- Nijst, P.; Verbrugge, F.H.; Grieten, L.; Dupont, M.; Steels, P.; Tang, W.H.W.; Mullens, W. The Pathophysiological Role of Interstitial Sodium in Heart Failure. J. Am. Coll. Cardiol. 2015, 65, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Khiabani, N.P.; Bahramian, A.; Ejtehadi, M.R.; Chen, P.; Pourafshary, P.; Goddard, W.A. Calcium Chloride Adsorption at Liquid-Liquid Interfaces: A Molecular Dynamics Simulation Study. Colloids Surf. A Physicochem. Eng. Asp. 2017, 527, 70–80. [Google Scholar] [CrossRef]
- Thelander, U.; Westermark, G.T.; Antoni, G.; Estrada, S.; Zancanaro, A.; Ihse, E.; Westermark, P. Cardiac Microcalcifications in Transthyretin (ATTR) Amyloidosis. Int. J. Cardiol. 2022, 352, 84–91. [Google Scholar] [CrossRef]
- Roijers, R.B.; Debernardi, N.; Cleutjens, J.P.M.; Schurgers, L.J.; Mutsaers, P.H.A.; van der Vusse, G.J. Microcalcifications in Early Intimal Lesions of Atherosclerotic Human Coronary Arteries. Am. J. Pathol. 2011, 178, 2879–2887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stats, M.A.; Stone, J.R. Varying Levels of Small Microcalcifications and Macrophages in ATTR and AL Cardiac Amyloidosis: Implications for Utilizing Nuclear Medicine Studies to Subtype Amyloidosis. Cardiovasc. Pathol. 2016, 25, 413–417. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Kd (mM) | R2; p-Value |
|---|---|---|
| Site-1 | 8.7 ± 2.1 | 0.993; 6.3 × 10−3 |
| Site-2 | 13.5 ± 1.3 | 0.999; 5.0 × 10−5 |
| Site-3 | 16.1 ± 2.2 | 0.998; 3.3 × 10−4 |
| Site-4 | 16.2 ± 3.8 | 0.993; 5.4 × 10−3 |
| Site-5 | 19.5 ± 6.2 | 0.973; 6.9 × 10−3 |
| Site-6 | 15.4 ± 2.1 | 0.979; 2.4 × 10−9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cantarutti, C.; Mimmi, M.C.; Verona, G.; Mandaliti, W.; Taylor, G.W.; Mangione, P.P.; Giorgetti, S.; Bellotti, V.; Corazza, A. Calcium Binds to Transthyretin with Low Affinity. Biomolecules 2022, 12, 1066. https://doi.org/10.3390/biom12081066
Cantarutti C, Mimmi MC, Verona G, Mandaliti W, Taylor GW, Mangione PP, Giorgetti S, Bellotti V, Corazza A. Calcium Binds to Transthyretin with Low Affinity. Biomolecules. 2022; 12(8):1066. https://doi.org/10.3390/biom12081066
Chicago/Turabian StyleCantarutti, Cristina, Maria Chiara Mimmi, Guglielmo Verona, Walter Mandaliti, Graham W. Taylor, P. Patrizia Mangione, Sofia Giorgetti, Vittorio Bellotti, and Alessandra Corazza. 2022. "Calcium Binds to Transthyretin with Low Affinity" Biomolecules 12, no. 8: 1066. https://doi.org/10.3390/biom12081066