1. Introduction
Autism spectrum disorders (ASDs) comprise a multifaceted, genetically heterogeneous group of neurodevelopmental conditions characterized by a range of behavioral impairments, including deficits in social interaction, repetitive behavior and increased sensitivity to sensory stimuli. ASDs have a complex and multifactorial etiology, involving genetic variations in several hundred genes as well as environmental factors. Prospective and retrospective clinical studies have identified valproic acid (VPA) as an environmental factor for ASDs. VPA is an anticonvulsant used to treat epilepsy, also used as a mood stabilizer to treat bipolar disorder, whose embryonic exposure has been associated with several instances of an increased risk for ASDs [
1,
2,
3]. Based on these observations, studies using embryonic exposure to VPA have also been conducted in many vertebrate species to model the core signs of ASDs [
4,
5,
6,
7,
8].
Several studies have analyzed social behavioral deficits in rodents prenatally exposed to VPA, describing lifelong impairments resembling the core signs of ASDs [
9]. In rats, a single administration of VPA in utero induces gender-specific social behavior abnormalities, modifies sensitivity to sensory stimuli, and increases repetitive behavior and anxiety [
4,
5,
6,
10]. Mice exposed to prenatal VPA also show cellular and molecular phenotypes in the medial prefrontal cortex [
11], the somatosensory cortex, the amygdala [
12], the brain stem and the cerebellum [
13]. In domestic chicks, VPA has been shown to produce alterations in early social-orienting responses and social interaction [
7,
8,
14].
The zebrafish is emerging as a popular model in biomedical research to study the effect of neurotoxicants involved in nervous system disorders. VPA exposure in zebrafish larvae has been shown to produce a plethora of effects on social behavior, locomotor activity and anxiety. Zimmerman et al. [
15] have reported changes in locomotor activity, increased anxiety and social interaction deficits in zebrafish larvae exposed to 48 µM VPA for 48 h. Liu et al. [
16] exposed 24 hpf zebrafish larvae to chronic (20 µM for 6 days) or acute (100 µM for 7 h) VPA treatment, reporting altered locomotor activity and social preference deficits. Molecular studies have also been conducted in VPA exposed zebrafish embryos to assess the neurobiological effect of VPA exposure. A recent study from Dwivedi et al. [
17] has reported altered levels of ASD-associated gene and protein expression in the brains of zebrafish larvae treated with 75 µM VPA for 5 days, accompanied by increased anxiety, social deficits and high circling behavior.
Histone deacetylases (HDACs) inhibition mediated by VPA has also been shown to directly inhibit
ascl1b expression, causing the selective failure of serotonergic identity [
18], a reduction of histaminergic neurons, changes in the expression of key genes of other monoaminergic systems, including dopamine, and decreased adult brain levels of noradrenaline and dopamine metabolites [
19].
Overall, the emerging picture on VPA exposure in zebrafish is very heterogeneous, with different doses, time and duration of exposure being reported. This poses serious limits to the interpretation of VPA action on neurodevelopment and to the use of the VPA zebrafish model to study ASD-relevant features.
To shed light on the neurotoxic effect of VPA on zebrafish larvae, we tested different micromolar concentrations of VPA, using exposure for 24 and 48 h in two different zebrafish inbred strains. Our data shows that, at 1 µM concentration, VPA mildly affects embryo survival and induces molecular alterations in neurodevelopmental genes, previously reported only using higher VPA doses, with substantial differences between strains.
3. Results
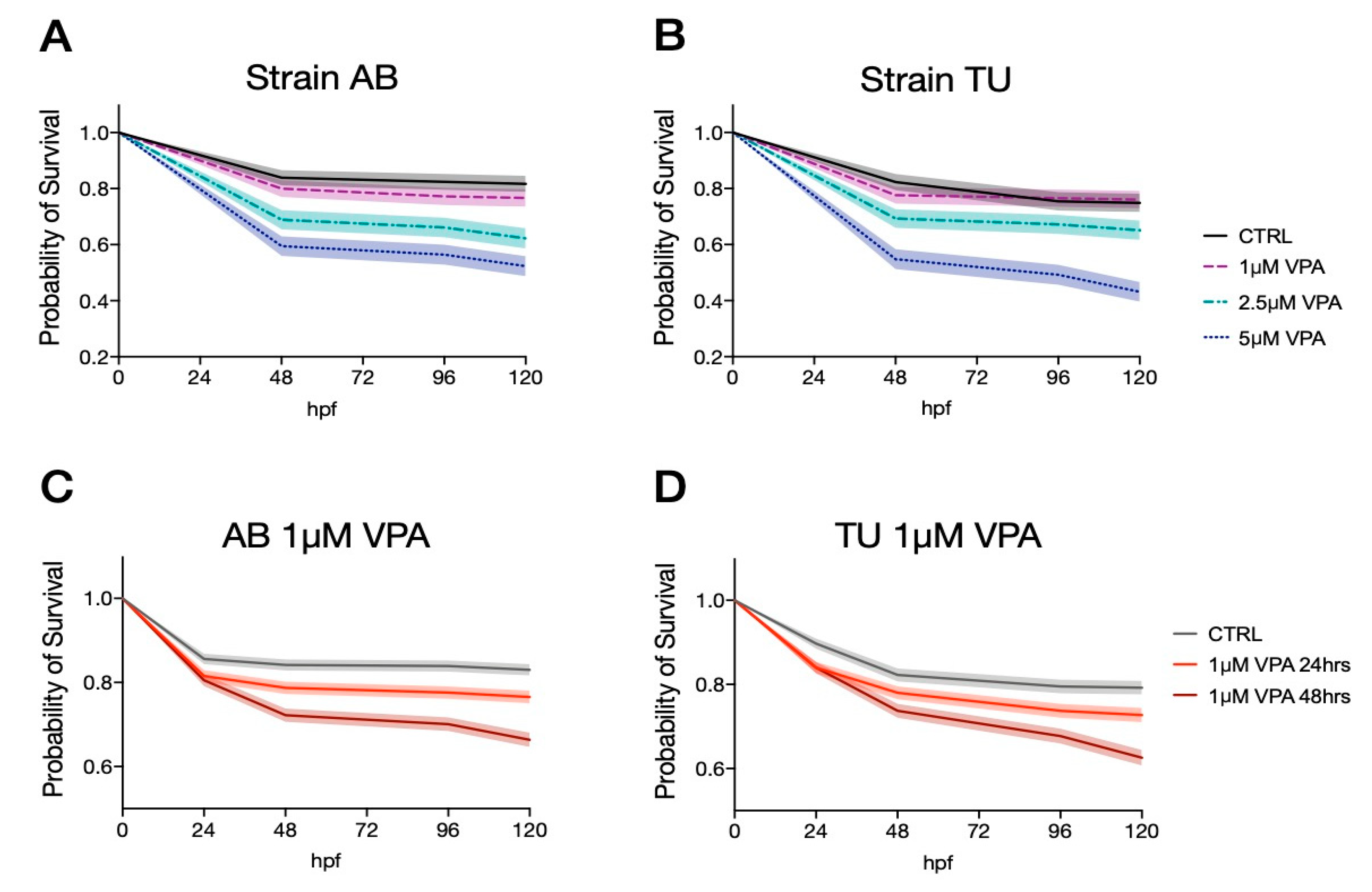
In order to identify the doses and administration regimens that have the lowest neurotoxic effects, we treated zebrafish larvae from two inbred strains (AB and TU) with different doses of VPA (1, 2.5 and 5 µM VPA) in five independent experiments (n = 735 and n = 756 for AB and TU, respectively). We administered 1, 2.5 and 5 µM VPA to zebrafish larvae for 24 h, starting from 5 hpf. The survival rates were analyzed at different time points (48, 96 and 120 hpf) using the log-rank (Mantel-Cox) test. AB zebrafish larvae treated with the vehicle showed a survival rate of 0.839 [95% C.I. 0.046, 0.062] at 48 hpf and at 96 hpf, and 0.817 [95% C.I. 0.049, 0.065] at 120 hpf. The survival rate for the three VPA concentrations (1 µM VPA: 0.800 [95% C.I. 0.051, 0.066] at 48 hpf, 0.772 [95% C.I. 0.055, 0.068] at 96 hpf and 0.767 [95% C.I. 0.055, 0.069] at 120 hpf; 2.5 µM VPA: 0.689 [95% C.I. 0.062, 0.073] at 48 hpf, 0.661 [95% C.I. 0.064, 0.074] at 96 hpf and 0.622 [95% C.I. 0.066, 0.075]; 5 µM VPA: 0.595 [95% C.I. 0.065, 0.072] at 48 hpf, 0.564 [95% C.I. 0.066, 0.073] at 96 hpf and 0.523 [95% C.I. 0.067, 0.072]) were significantly different from the vehicle-treated controls, as indicated by the log-rank (Mantel-Cox) test (
Figure 1A,
χ2(3,735) = 59.63,
p < 0.0001).
The vehicle treated TU larvae survival rates were 0.823 [95% C.I. 0.049, 0.065] at 48 hpf, 0.754 [95% C.I. 0.057, 0.071] at 96 hpf and 0.749 [95% C.I. 0.058, 0.071] at 120 hpf. Valproic acid treatment induced a significant reduction of the survival rates in all three concentrations (
Figure 1B, log-rank (Mantel-Cox) test
χ2(3,756) = 46.60,
p < 0.0001). The survival rates at the different concentration and time points were as follows: 1 µM VPA 0.776 [95% C.I. 0053, 0.066] at 48 and 96 hpf and 0.760 [95% C.I. 0.054, 0.067] at 120 hpf; 2.5 µM VPA 0.693 [95% C.I. 0.060, 0.071] at 48 hpf, 0.672 [95% C.I. 0.061, 0.071] at 96 hpf and 0.651 [95% C.I. 0.063, 0.072] at 120 hpf; 5 µM VPA 0.584 [95% C.I. 0.066, 0.072] at 48 hpf, 0.492 [95% C.I. 0.068, 0.072] at 96 hpf and 0.431 [95% C.I. 0.068, 0.070] at 120 hpf.
Overall, valproic acid seems to produce a similar toxic effect on the two strains, as indicated by the significant effect of VPA treatments in both strains. The two strains seem to have a similar survival rate in the vehicle-treated samples at 48 hpf but tend to diverge at later time points, especially when treated with the highest VPA concentration (log-rank (Mantel-Cox) test between AB and TU 5 µM VPA, χ2(1,392) = 3.0770 uncorrected p = 0.0794, data not shown).
Given the similar neurotoxic effect of the different VPA concentrations, we evaluated the lowest tested concentrations using two distinct regimens: treatment for 24 or 48 h. The survival rates for zebrafish larvae treated with the two regimens were similar in both strains, as indicated in
Figure 1C,D, representative of AB and TU strain, respectively. The survival rates for AB were for vehicle-treated larvae 0.856 [95% C.I. 0023, 0.027] at 24 hpf, 0.842 [95% C.I. 0.024, 0.028] at 48 hpf, 0.839 [95% C.I. 0.024, 0.028] at 96 hpf and 0.830 [95% C.I. 0.025, 0.028] at 120 hpf; 1 µM VPA for 24 h 0.815 [95% C.I. 0.025, 0.029] at 24 hpf, 0.787 [95% C.I. 0.027, 0.030] at 48 hpf, 0.776 [95% C.I. 0.028, 0.031] at 96 hpf and 0.766 [95% C.I. 0.028, 0.031] at 120 hpf; 1 µM VPA for 48 h 0.805 [95% C.I. 0.026, 0.029] at 24 hpf, 0.722 [95% C.I. 0.030, 0.033] at 48 hpf, 0.701 [95% C.I. 0.031, 0.033] at 96 hpf and 0.664 [95% C.I. 0.032, 0.034] at 120 hpf. Log-rank (Mantel-Cox) test indicated a significant effect of VPA treatment for both regimens compared to the vehicle (
Figure 1C, log-rank (Mantel-Cox) test
χ2(2, 2340) = 55.93,
p < 0.0001).
TU strain larvae showed comparable survival rates: vehicle-treated larvae 0.897 [95% C.I. 0021, 0.027] at 24 hpf, 0.823 [95% C.I. 0028, 0.032] at 48 hpf, 0.795 [95% C.I. 0030, 0.034] at 96 hpf and 0.792 [95% C.I. 0030, 0.034] at 120 hpf; 1 µM VPA for 24 h 0.840 [95% C.I. 0.025, 0.029] at 24 hpf, 0.780 [95% C.I. 0.029, 0.033] at 48 hpf, 0.737 [95% C.I. 0.031, 0.034] at 96 hpf and 0.727 [95% C.I. 0.031, 0.035] at 120 hpf; 1 µM VPA for 48 h 0.840 [95% C.I. 0.025, 0.029] at 24 hpf, 0.737 [95% C.I. 0.031, 0.034] at 48 hpf, 0.677 [95% C.I. 0.033, 0.036] at 96 hpf and 0.626 [95% C.I. 0.035, 0.037] at 120 hpf. As for AB strain larvae, a significant difference was observed in the survival rates among treatment regimens (
Figure 1D, log-rank (Mantel-Cox) test
χ2(2, 2020) = 42.10,
p < 0.0001).
Direct comparison between survival rates of embryos exposed to VPA for 24 and 48 h indicated a significant effect of treatment regimens in both strains (
Figure 1C,D, log-rank (Mantel-Cox) test:
χ2(1, 1570) = 18.19, uncorrected
p < 0.0001 for AB and
χ2(1, 1400) = 14.02, uncorrected
p < 0.0002 for TU).
To assess the neurodevelopmental effects of VPA treatments, we compared the expression levels of genes previously shown to be affected by VPA treatment at higher dosage and different treatment regimens. To assess the effect of treatment, age and transcript on gene expression levels, we used a linear mixed model (LMM), considering treatment, age and transcript as fixed factors and the experimental unit (experiment) as random factor. We compared a model with random-intercepts only to one with random slopes and intercepts, using the likelihood ratio, and found that the random slopes and intercepts approach fitted the data significantly better.
Data from the two zebrafish inbred strains AB and TU were analyzed separately. We found that gene expression was significantly affected by treatment, age and transcript in both strains (LMM for AB: treatment χ2(2) = 30.78, p < 0.0001; age χ2(1) = 9.04, p = 0.0026; transcript χ2(9) = 1216.56, p < 0.0001. LMM for TU: treatment χ2(2) = 7.05, p = 0.0295; age χ2(1) = 38.11, p < 0.0001; transcript χ2(9) = 492.79, p < 0.0001). Significant interactions between the fixed factors were also assessed in both strains, in particular a triple interaction between treatment, age and transcript was observed for both AB and TU zebrafish larvae (LMM for AB: treatment*age χ2(2) = 1.84, p = 0.3978; treatment*transcript χ2(18) = 17.52, p = 0.4875; age*transcript: χ2(9) = 282.88, p < 0.0001; treatment*age*transcript: χ2(18) = 37.91, p = 0.0040. LMM for TU: treatment*age χ2(2) = 30.88, p < 0.0001; treatment*transcript χ2(18) = 26.54, p = 0.0880; age*transcript: χ2(9) = 580.06, p < 0.0001; treatment*age*transcript: χ2(18) = 56.13, p < 0.0001).
VPA has been shown to impair development of the 5HT system acting on 5HT neuronal progenitor differentiation [
18]. Therefore, we examined the effect of minimal doses of VPA (1 µM) on genes involved in 5HT neuron differentiation in the two different strains.
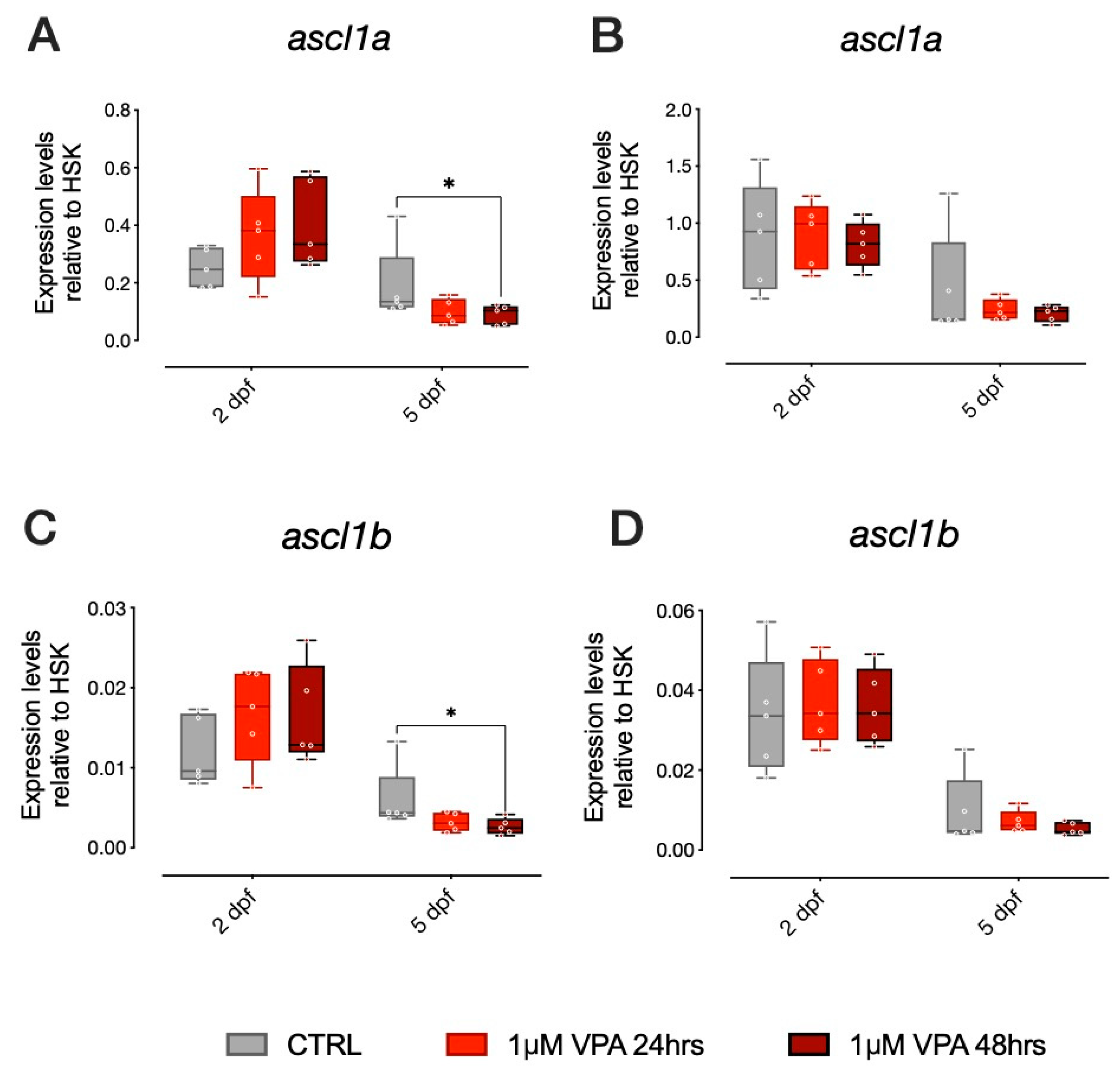
ascl1a and
ascl1b are bHLH transcription factors, homologs to the mammalian
Ascl1 (also known as
Mash1) involved in neural differentiation, that have been shown to be direct targets of VPA and to affect 5HT progenitor specification [
18]. In the AB strain larvae, we found that gene expression levels for
ascl1a were different in vehicle-treated control larvae at 5 dpf compared to larvae treated with 1 µM VPA for 48 h, as shown in
Figure 2A (Tukey post-hoc pairwise comparison at 5 dpf: CTRL-1 µM VPA for 48 h t
(236) = 2.528,
p = 0.0324). For
ascl1b, we found a similar effect of treatment in the AB strain larvae. The
ascl1b gene expression levels were different from vehicle-treated control larvae at 5 dpf in larvae treated with 1 µM VPA for 48 h (
Figure 2C; Tukey post-hoc pairwise comparison at 5 dpf: CTRL-1 µM VPA for 48 h t
(226) = 2.839,
p = 0.0136). Thus, exposure to 1 µM VPA for 48 h seems to be sufficient to affect the expression of proneural genes involved in differentiation and specification of progenitor neurons in the AB strain. In the TU strain, however, VPA did not produce the same effect on
ascl1a and
ascl1b. We did not observe any significant difference in the expression levels of
ascl1a or
ascl1b in the TU strain (
Figure 2B,D, respectively).
Notice that our data also indicates that ascl1a and ascl1b expression is significantly affected by age in both AB and TU strains, as it decreases between 2 and 5 dpf, independent of VPA treatment. Overall, we observed a different effect of VPA treatment at 1 µM concentration in the two zebrafish strains, as ascl1a and ascl1b expression seemed to be affected by VPA treatment only in larvae from the AB strain.
VPA-mediated changes in
ascl1a and
ascl1b have been previously shown to affect the brain serotonin system through direct inhibition of serotonin progenitor differentiation. We thus analyzed the expression of
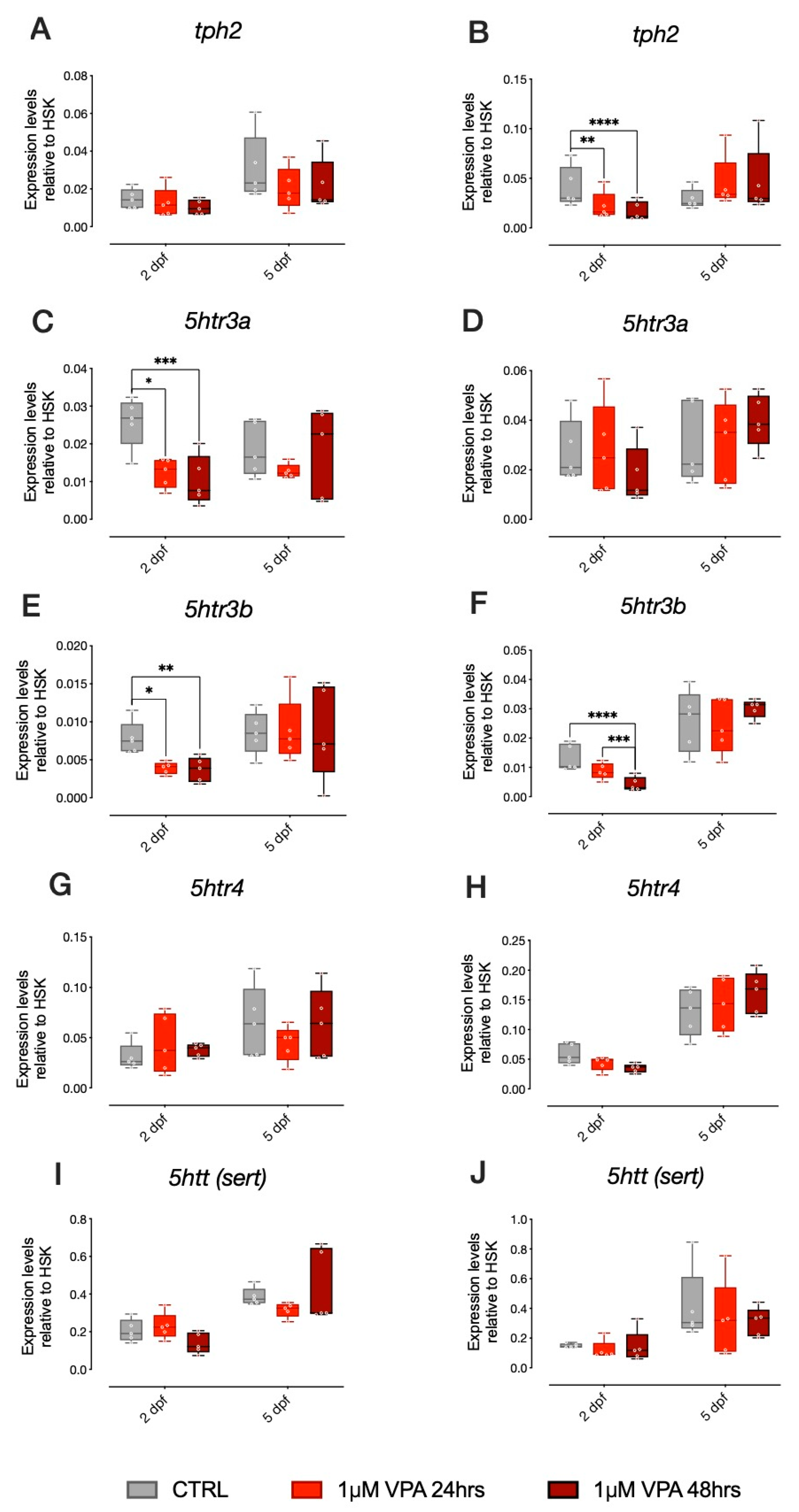
tph2, the rate-limiting enzyme for brain serotonin synthesis, as well as other genes involved in serotonergic neurotransmission, such as the serotonin transporter (
sert) and three of the most important serotonin receptors (
5htr3a, 5htr3b and
5ht4). For
tph2, we did not observe any significant effect of treatment in AB strain larvae (
Figure 3A). Despite the absence of significant changes in
ascl1a and
ascl1b expression levels, in the TU strain larvae, we observed that
tph2 expression levels were significantly affected by our VPA treatment regimen, but only at 2 dpf (
Figure 3B; Tukey post-hoc pairwise comparison at 2 dpf: CTRL-1 µM VPA 24 h t
(236) = 2.978,
p = 0.0090; CTRL-1 µM VPA 48 h F
(2, 8) = 4.230,
p = 0.0001).
Concerning the expression of 5HT receptors, in AB strain larvae,
5htr3a expression levels were significantly affected by treatment at 2 dpf (
Figure 3C; Tukey post-hoc pairwise comparison at 2 dpf: CTRL-1 µM VPA 24 h t
(236) = 2.898,
p = 0.0114; CTRL-1 µM VPA 48 h t
(236) = 4.087,
p = 0.0002). In the TU strain larvae,
5htr3a expression levels were not significantly affected by treatment (
Figure 3D). For
5htr3b we observed a significant effect of treatment in AB strain larvae at 2 dpf (
Figure 3E; Tukey post-hoc pairwise comparison at 2 dpf: CTRL-1 µM VPA 24 h t
(236) = 2.464,
p = 0.0235; CTRL-1 µM VPA 48 h t
(236) = 3.112,
p = 0.0059). In TU strain larvae, we also observed a significant effect of treatment on the expression levels of
5htr3b at 2 dpf (
Figure 3F; Tukey post-hoc pairwise comparison at 2 dpf: CTRL-1 µM VPA 48 h t
(236) = 5.507,
p < 0.0001; 1 µM VPA 24 h–1 µM VPA 48 h t
(236) = 3.642,
p = 0.0010). For
5htr4 and
sert, we did not observe any effect of treatment in AB or TU strain larvae (
Figure 3G,H,I,J, respectively).
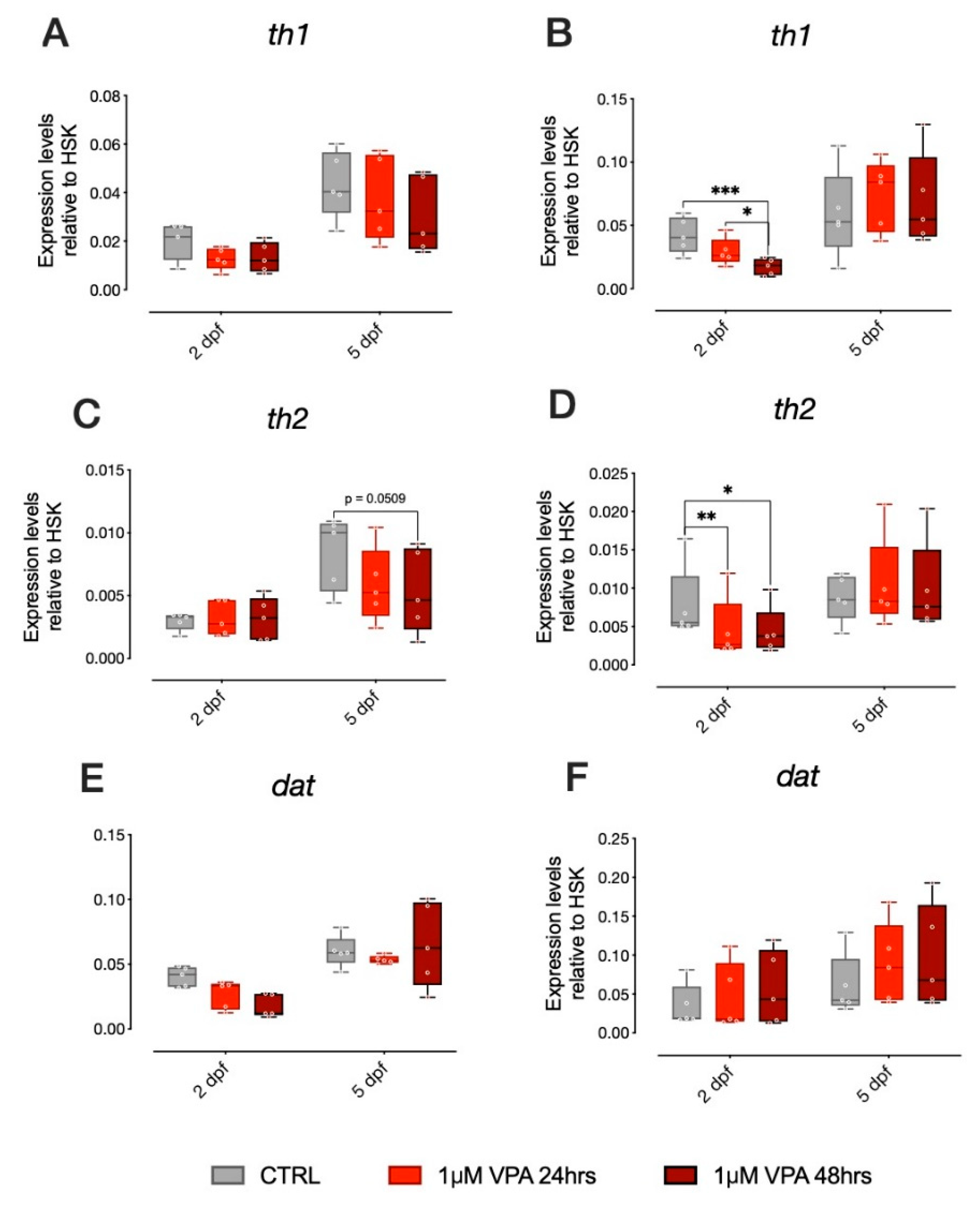
In addition to several reports on the effect of VPA on the serotonergic system, studies on zebrafish have shown that VPA treatment also affects key genes in other monoaminergic systems, in particular dopamine [
19]. We thus analyzed genes involved in dopamine synthesis and metabolism in zebrafish larvae from the two different strains treated with our administration regimens (1 µM VPA for 24 or 48 h). Expression of
th1,
th2 and
dat were analyzed and statistical evaluation of the changes in gene expression did not reveal any significant effect of treatment in
th1 and
dat in the larvae of the AB strain (
Figure 4A,E, respectively), although an effect of VPA treatment on
th2 expression was observed (
Figure 4C; Tukey post-hoc pairwise comparison at 5 dpf:
th1, CTRL-1 µM VPA 48 h t
(236) = 2.351,
p = 0.0509). In the TU strain larvae, however, we found a significant effect of treatment on the expression of both
th1 and
th2, but not on
dat (
Figure 4B,D,F, respectively; Tukey post-hoc pairwise comparison at 2 dpf:
th1, CTRL-1 µM VPA 24 h t
(236) = 4.122,
p = 0.0002; CTRL-1 µM VPA 48 h t
(236) = 2.477,
p = 0.04;
th2, CTRL-1 µM VPA 24 h t
(236) = 3.010,
p = 0.0081; CTRL-1 µM VPA 48 h t
(236) = 2.851,
p = 0.0131).
4. Discussion
The administration of VPA in zebrafish represents a powerful model to study neurobiological and molecular changes relevant for ASDs. However, the use of heterogeneous experimental paradigms (dose and time of administration) limit comparison between studies, hindering the use of zebrafish models to investigate the neurobiological mechanisms underlying ASDs.
In order to identify the doses and administration regimens that had the lowest neurotoxic effects, we extended previous studies and tested micromolar doses of VPA, analyzing the animal survival rates and the expression of neurodevelopmental genes in two different wild type inbred strains: the AB and TU. We exposed AB and TU zebrafish embryos at 5 hpf to 1, 2.5 and 5 µM of VPA for 24 h and followed their survival rates for 5 days. We found that VPA already had a significant toxic effect at 1 µM concentration on both strains. We also characterized the effect of exposure to 1 µM VPA for 24 or 48 h on the expression of genes involved in neurodevelopment and known to impact neurotransmitter phenotypes.
The survival data in our study was in line with previous reports [
22], since 1 µM VPA produced an increase in mortality in both AB and TU zebrafish strains. The discrepancy among our study and previous studies that did not report neurotoxic effect of VPA at higher concentrations, may be explained by the genetic differences of the inbred strains used in the studies. Zimmermann et al. [
15] did not report the strain of the animals used in their study, while Baronio et al. [
19] used the Turku strain. This study reported a high mortality rate (above 50%) in larvae treated with 50 µM, a high level of deformities and arrested development using 35 µM VPA, while no significant effects on mortality were reported by 25 µM VPA treatment [
19]. Our data is also supported by Li et al. [
22], who used a mixed AB-TU strain. Indeed, Li et al. [
22] showed that, already at concentration ranges between 1 and 10 µM, VPA induced malformations in the endocrine and exocrine pancreas and the liver, also affecting craniofacial cartilage formation and vascular development.
We analyzed the expression of several neurodevelopmental genes at 2 and 5 dpf and observed an overall effect of age, independent of treatment. As expected, neurodevelopmental programs shape neural circuits in the first 5 days of development, affecting the expression levels of many genes. Globally, gene expression seemed to be mostly affected by VPA at 2 dpf in the TU strain larvae. Instead, for the AB strain, we observed gene expression changes both at 2 and at 5 dpf.
Most importantly, we observed striking differences induced by VPA in the expression of some of the genes between AB and TU strains. Given the equivalent effect of VPA at 1 µM concentration on survival rate in the two strains, we hypothesize that different genetic programs may activate distinct gene expression pathways in the two strains. Further studies, extending the set of genes analyzed through whole transcriptome sequencing, may be necessary to obtain a deeper understanding of the molecular programs that are affected by VPA in the AB compared to the TU strain.
Expression of the early neurogenic genes,
ascl1a and
ascl1b, was only affected by VPA in the AB strain. Previous reports analyzed the effect of HDAC inhibitors on neurogenic programs, showing the prominent role of HDAC in coordinating transcription and regulating cell fate in the developing nervous system through
ascl1a/b [
23]. HDAC inhibition mediated by VPA, directly inhibits
ascl1b, causing the selective failure of serotonergic identity [
18]. A similar effect was observed only in the AB strain larvae exposed to 1 µM VPA at 5 dpf, but not in the TU larvae, suggesting a difference in this specific neurodevelopmental program in the two strains. Despite the fact that VPA did not affect
ascl1a/b expression in TU larvae, we still found significant changes in serotonergic markers in this strain. This data suggests that VPA may be sufficient to alter the serotonergic system development, independent of the genetic background, but indicates that different programs could be exploited to obtain similar effects on the serotonergic system in the two strains. Inbred strains may differ substantially due to inbreeding and genetic drift [
24]. We tested two inbred, commercially available strains and showed that VPA exposure affected different molecular pathways in the two strains, suggesting different susceptibility to the drug and/or different mechanism of action.
Previous studies have analyzed the effect of VPA exposure on other monoaminergic system development, including the dopaminergic system [
19], demonstrating a reduction of
th1 expression. We confirmed previous data with our dosage and administration regimens, analyzing the expression of
th1,
th2 and
dat in both strains.
th1 and
th2 expression was significantly affected by VPA only in the TU strain, although an effect of VPA treatment on
th2 expression was also observed also in the AB strain (
Figure 4C). This data further highlights the potential differential effect of VPA on larvae of the two strains.
Recent studies have revealed alterations in the dopaminergic systems of mice embryonically exposed to VPA [
25]. Epidemiological and animal model studies have also suggested that perinatal alterations in 5HT, either above or below typical levels, may cause social behavioral deficits resembling ASDs [
26].
The lack of behavioral tests and detailed neuroanatomical analyses represents a limit to the interpretation of the molecular mechanisms underlying the neurotoxic action of VPA at micromolar doses used in this study. Previous reports have shown that zebrafish larvae do not exhibit social behavioral responses until 3 or 4 weeks of age [
16,
27], limiting the possibility to test social behavior at early stages. Future studies will determine whether low doses of VPA have long-lasting effects on social behavior and locomotor activity.

{kind=link}
{kind=link}
{kind=link}
{kind=link}



