Identification of a Biosynthetic Gene Cluster Responsible for the Production of a New Pyrrolopyrimidine Natural Product—Huimycin


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and BACs
2.2. DNA Manipulation
2.3. Metabolite Extraction and Analysis
2.4. Huimycin Isolation and NMR Data Acquisition
2.5. Genome Mining and Bioinformatics Analysis
3. Results and Discussion
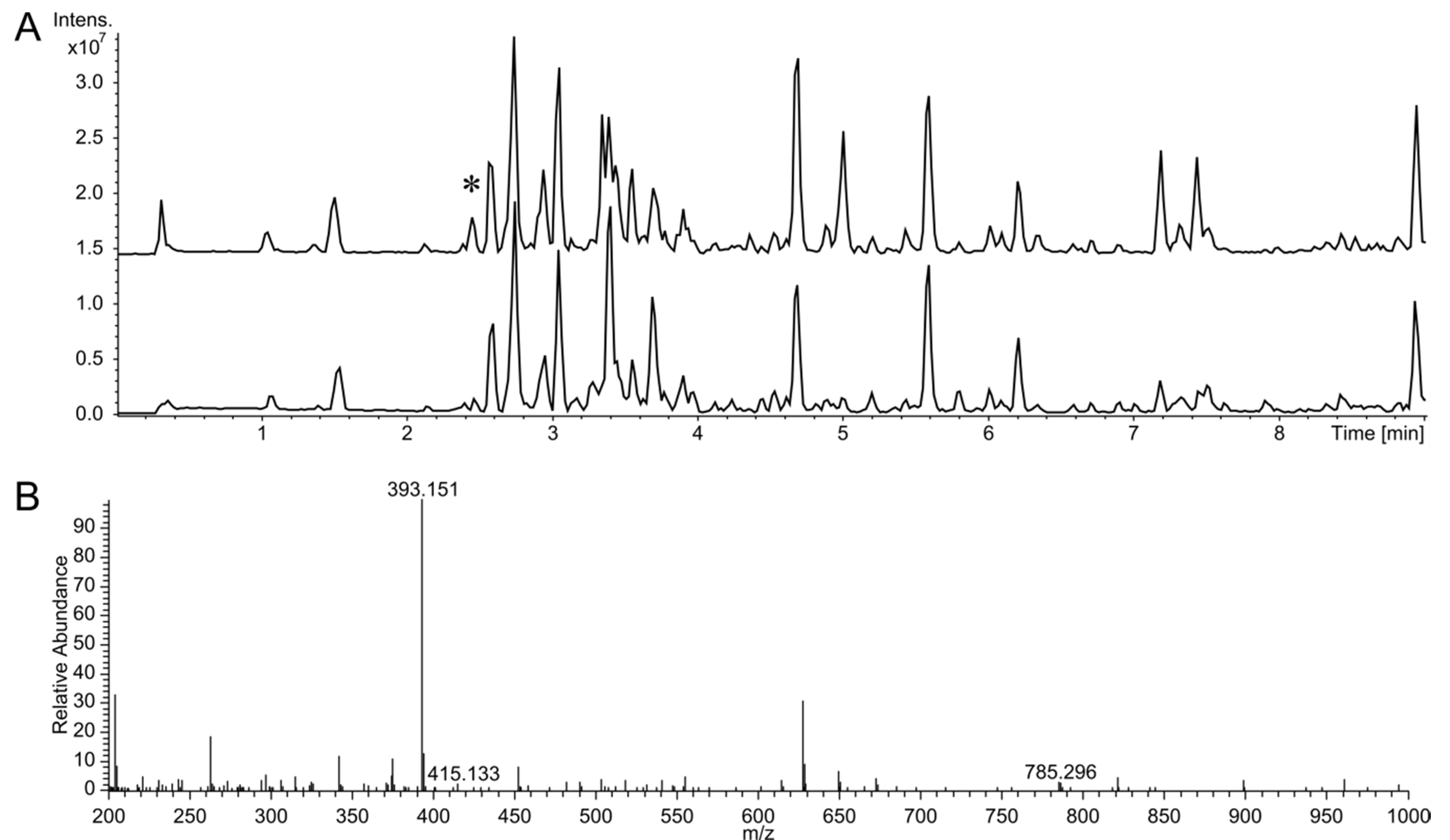
3.1. Identification of the Huimycin Gene Cluster Through Its Heterologous Expression in Streptomyces albus Del14
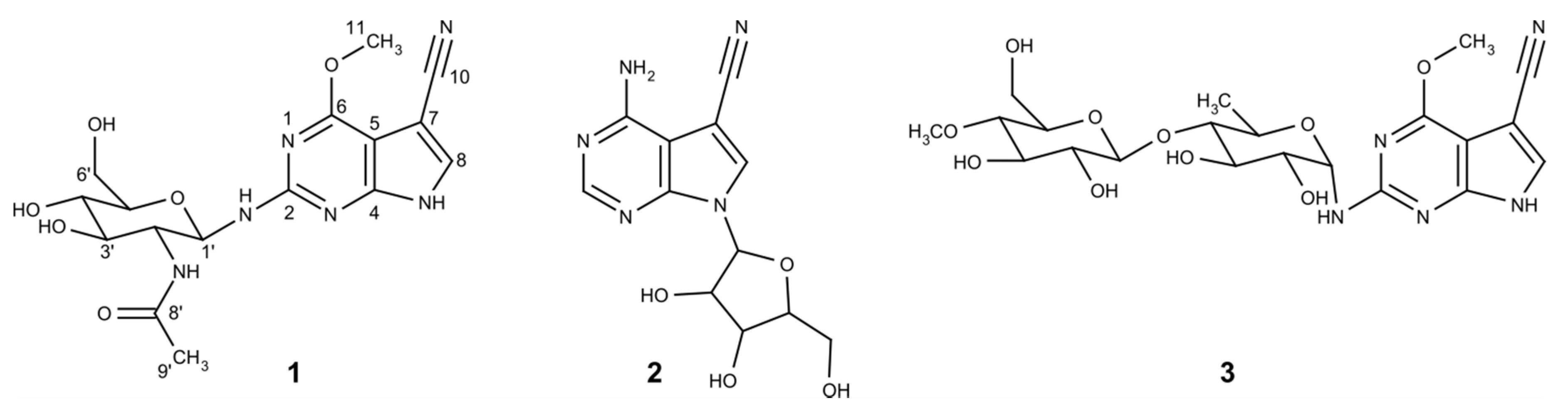
3.2. Isolation and Structure Elucidation of the Huimycin
3.3. Determination of the Minimal Huimycin Gene Cluster
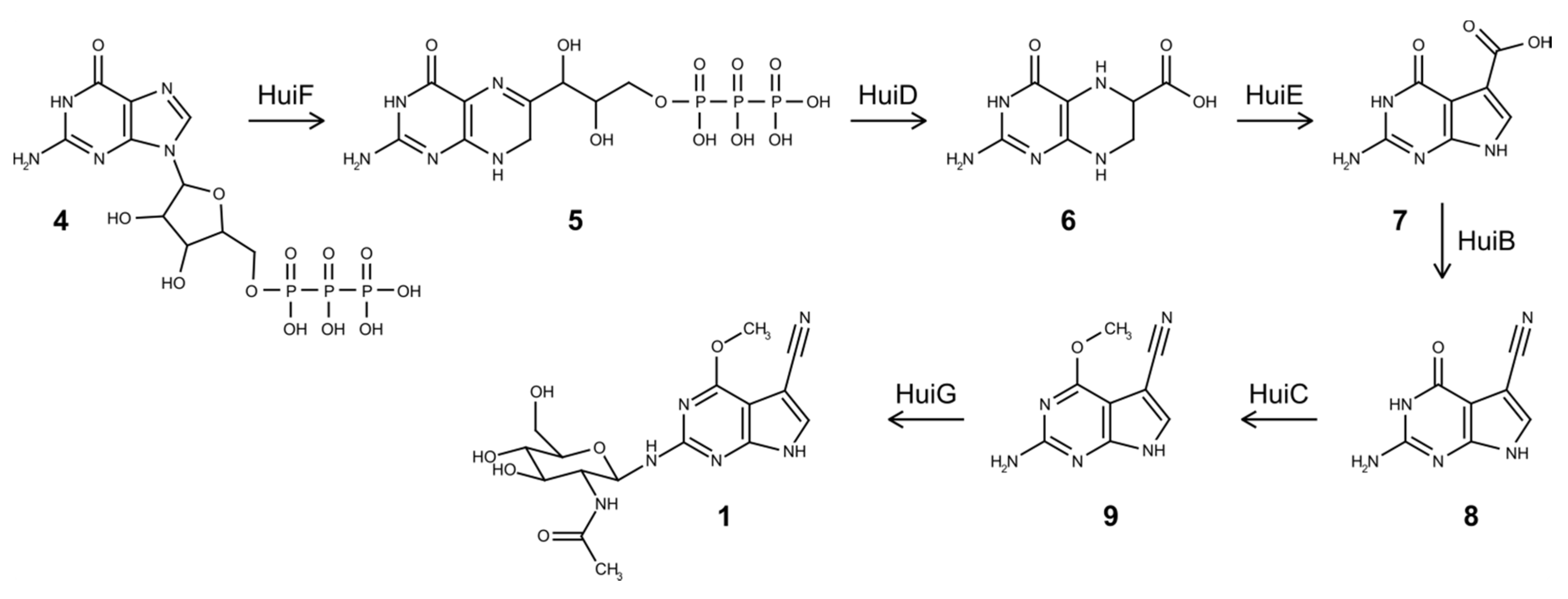
3.4. Biosynthesis of Huimycin
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McCarty, R.M.; Bandarian, V. Biosynthesis of pyrrolopyrimidines. Bioorg. Med. Chem. 2012, 43, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, H.; Katagiri, K.; Sato, K.; Mayama, M.; Shimaoka, N. Toyocamycin, a new anti-candida antibiotics. J. Antibiot. 1956, 9, 60–62. [Google Scholar] [PubMed]
- Anzai, K.; Nakamura, G.; Suzuki, S. A new antibiotic, tubercidin. J. Antibiot. 1957, 10, 201–204. [Google Scholar] [PubMed]
- Rao, K.V. Structure of sangivamycin. J. Med. Chem. 1968, 11, 939–941. [Google Scholar] [CrossRef]
- Vinayak, M.; Pathak, C. Queuosine modification of tRNA: Its divergent role in cellular machinery. Biosci. Rep. 2009, 30, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Harada, F.; Nishimura, S. Possible anticodon sequences of tRNA His, tRNA Asm, and tRNA Asp from Escherichia coli B. Universal presence of nucleoside Q in the first postion of the anticondons of these transfer ribonucleic acids. Biochemistry 1972, 11, 301–308. [Google Scholar] [CrossRef]
- Meier, F.; Suter, B.; Grosjean, H.; Keith, G.; Kubli, E. Queuosine modification of the wobble base in tRNAHis influences “In Vivo” decoding properties. EMBO J. 1985, 4, 823–827. [Google Scholar] [CrossRef]
- Urbonavicius, J.; Qian, Q.; Durand, J.M.; Hagervall, T.G.; Björk, G.R. Improvement of reading frame maintenance is a common function for several tRNA modifications. EMBO J. 2001, 20, 4863–4873. [Google Scholar] [CrossRef] [Green Version]
- Kilpatrick, M.W.; Walker, R.T. The nucleotide sequence of the tRNAMMet from the archaebacterium Thermoplasma acidophilum. Nucleic Acids Res. 1981, 9, 4387–4390. [Google Scholar] [CrossRef] [Green Version]
- Pathania, S.; Rawal, R.K. Pyrrolopyrimidines: An update on recent advancements in their medicinal attributes. Eur. J. Med. Chem. 2018, 157, 503–526. [Google Scholar] [CrossRef]
- Saneyoshi, M.; Tokuzen, R.; Fukuoka, F. Antitumor Activities and Structural Relationship of Tubercidine, Toyocamycin, and Their Derivatives. GANN Jpn. J. Cancer Res. 1965, 56, 219–222. [Google Scholar] [CrossRef]
- Acs, G.; Reich, E.; Mori, M. Biological and biochemical properties of the analogue antibiotic tubercidin. Proc. Natl. Acad. Sci. USA 1964, 52, 493–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindberg, B.; Klenow, H.; Hansen, K. Some properties of partially purified mammalian adenosine kinase. J. Biol. Chem. 1967, 242, 350–356. [Google Scholar] [CrossRef]
- Suhadolnik, R.J.; Uematsu, T.; Uematsu, H. Toyocamycin: Phosphorylation and incorporation into RNA and DNA and the biochemical properties of the triphosphate. Biochim. Biophys. Acta 1967, 149, 41–49. [Google Scholar] [CrossRef]
- Uretsky, S.C.; Acs, G.; Reich, E.; Mori, M.; Altwerger, L. Pyrrolopyrimidine nucleotides and protein synthesis. J. Biol. Chem. 1968, 243, 306–312. [Google Scholar] [PubMed]
- Loomis, C.R.; Bell, R.M. Sangivamycin, a nucleoside analogue, is a potent inhibitor of protein kinase C. J. Biol. Chem. 1988, 263, 1682–1692. [Google Scholar] [PubMed]
- Osada, H.; Sonoda, T.; Tsunoda, K.; Isono, K. A new biological role of sangivamycin; inhibition of protein kinases. J. Antibiot. 1989, 42, 102–106. [Google Scholar] [CrossRef]
- Roskoski, R. Properties of FDA-approved small molecule protein kinase inhibitors. Pharmacol. Res. 2019, 144, 19–50. [Google Scholar] [CrossRef]
- Mogul, A.; Corsi, K.; McAuliffe, L. Baricitinib: The Second FDA-Approved JAK Inhibitor for the Treatment of Rheumatoid Arthritis. Ann. Pharmacother. 2019, 53, 947–953. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 2012. [Google Scholar]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics; John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Rodríguez Estévez, M.; Myronovskyi, M.; Gummerlich, N.; Nadmid, S.; Luzhetskyy, A. Heterologous Expression of the Nybomycin Gene Cluster from the Marine Strain Streptomyces albus subsp. chlorinus NRRL B-24108. Mar. Drugs 2018, 16, 435. [Google Scholar] [CrossRef] [Green Version]
- Muyrers, J.P.P.; Zhang, Y.; Benes, V.; Testa, G.; Rientjes, J.M.J.; Stewart, A.F. ET recombination: DNA engineering using homologous recombination in E. coli. Methods Mol. Biol. 2004, 256, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebets, Y.; Tokovenko, B.; Lushchyk, I.; Rückert, C.; Zaburannyi, N.; Bechthold, A.; Kalinowski, J.; Luzhetskyy, A. Complete genome sequence of producer of the glycopeptide antibiotic Aculeximycin Kutzneria albida DSM 43870T, a representative of minor genus of Pseudonocardiaceae. BMC Genom. 2014, 15, 885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shomura, T.; Nishizawa, N.; Iwata, M.; Yoshida, J.; Ito, M.; Amano, S.; Koyama, M.; Kojima, M.; Inouye, S. Studies on a new nucleoside antibiotic, dapiramicin. I. Producing organism, assay method and fermentation. J. Antibiot. 1983, 36, 1300–1304. [Google Scholar] [CrossRef]
- Nishizawa, N.; Kondo, Y.; Koyama, M.; Omoto, S.; Iwata, M.; Tsuruoka, T.; Inouye, S. Studies on a new nucleoside antibiotic, dapiramicin. II. Isolation, physico-chemical and biological characterization. J. Antibiot. 1984, 37, 1–5. [Google Scholar] [CrossRef] [Green Version]
- McCarty, R.; Bandarian, V. Rosetta stone for deciphering deazapurine biosynthesis: Pathway for pyrrolopyrimidine nucleosides toyocamycin and sangivamycin. Chem. Biol. 2008, 15, 790–798. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Song, Z.; Xu, X.; Ma, Z.; Bechthold, A.; Yu, X. ToyA, a positive pathway-specific regulator for toyocamycin biosynthesis in Streptomyces diastatochromogenes 1628. Appl. Microbiol. Biotechnol. 2019, 103, 7071–7084. [Google Scholar] [CrossRef]
- Iijima, M.; Kubota, Y.; Sawa, R.; Kubota, Y.; Hatano, M.; Igarashi, M.; Kawada, M.; Momose, I.; Takekawa, M.; Shibasaki, M. A guanine derivative as a new MEK inhibitor produced by Streptomyces sp. MK63-43F2. J. Antibiot. 2017. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δCa | δH mult. (J, Hz) b | HMBC c |
|---|---|---|---|
| 2 | 160.3 | ||
| 4 | 155.4 | ||
| 5 | 98.7 | ||
| 6 | 164.5 | ||
| 7 | 84.5 | ||
| 8 | 131.0 | 7.60, s | 2 e, 4, 5, 6 e, 7, 10 |
| 10 | 116.3 | ||
| 11 | 54.2 | 4.05, s | 5 e, 6 |
| 1′ | 83.6 | 5.28, d 9.6 | 2, 2′, 3′, 5′ |
| 2′ | 56.1 | 3.87 d | 1′, 3′, 8′ |
| 3′ | 76.3 | 3.55, m | 2′, 4′ |
| 4′ | 71.9 | 3.38 d | 5′, 6′ |
| 5′ | 79.2 | 3.39 d | |
| 6′ a | 62.6 | 3.69, m | 4′ |
| 6′ b | 3.85 d | ||
| 8′ | 174.5 | ||
| 9′ | 22.7 | 1.96, s | 8′ |
| Gene | Proposed Function | Homolog in Toy Pathway | Identities/Positives |
|---|---|---|---|
| KALB_4064 | Enhanced intracellular survival protein | ||
| KALB_4065 | Hypothetical protein | ||
| KALB_4066 | Hypothetical protein | ||
| huiA; KALB_4067 | Pathway-specific regulator | toyA | 33%/46% |
| huiB; KALB_4068 | 7-cyano-7-deazaguanine synthase | toyM | 72%/82% |
| huiC; KALB_4069 | SAM-dependent methyltransferase | ||
| huiD; KALB_4070 | 6-carboxytetrahydropterin synthase | toyB | 65%/76% |
| huiE; KALB_4071 | 7-carboxy-7-deazaguanine synthase | toyC | 50%/55% |
| huiF; KALB_4072 | GTP cyclohydrolase I | toyD | 66%/75% |
| huiG; KALB_4073 | Glycosyltransferase | ||
| KALB_4074 | Carbamoyltransferase | ||
| KALB_4075 | Pyridoxamine 5′-phosphate oxidase | ||
| KALB_4076 | SAM-dependent methyltransferase | ||
| KALB_4077 | Glycosyltransferases | ||
| KALB_4078 | Glycosyltransferases |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shuai, H.; Myronovskyi, M.; Nadmid, S.; Luzhetskyy, A. Identification of a Biosynthetic Gene Cluster Responsible for the Production of a New Pyrrolopyrimidine Natural Product—Huimycin. Biomolecules 2020, 10, 1074. https://doi.org/10.3390/biom10071074
Shuai H, Myronovskyi M, Nadmid S, Luzhetskyy A. Identification of a Biosynthetic Gene Cluster Responsible for the Production of a New Pyrrolopyrimidine Natural Product—Huimycin. Biomolecules. 2020; 10(7):1074. https://doi.org/10.3390/biom10071074
Chicago/Turabian StyleShuai, Hui, Maksym Myronovskyi, Suvd Nadmid, and Andriy Luzhetskyy. 2020. "Identification of a Biosynthetic Gene Cluster Responsible for the Production of a New Pyrrolopyrimidine Natural Product—Huimycin" Biomolecules 10, no. 7: 1074. https://doi.org/10.3390/biom10071074