
Morphofunctional Alteration of Mesenteric Lymph Nodes in the Inflammation of the Abdominal Cavity

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Experiment Design
2.3. Morphological Research
2.4. Morphometric Analysis
2.5. Statistical Analysis
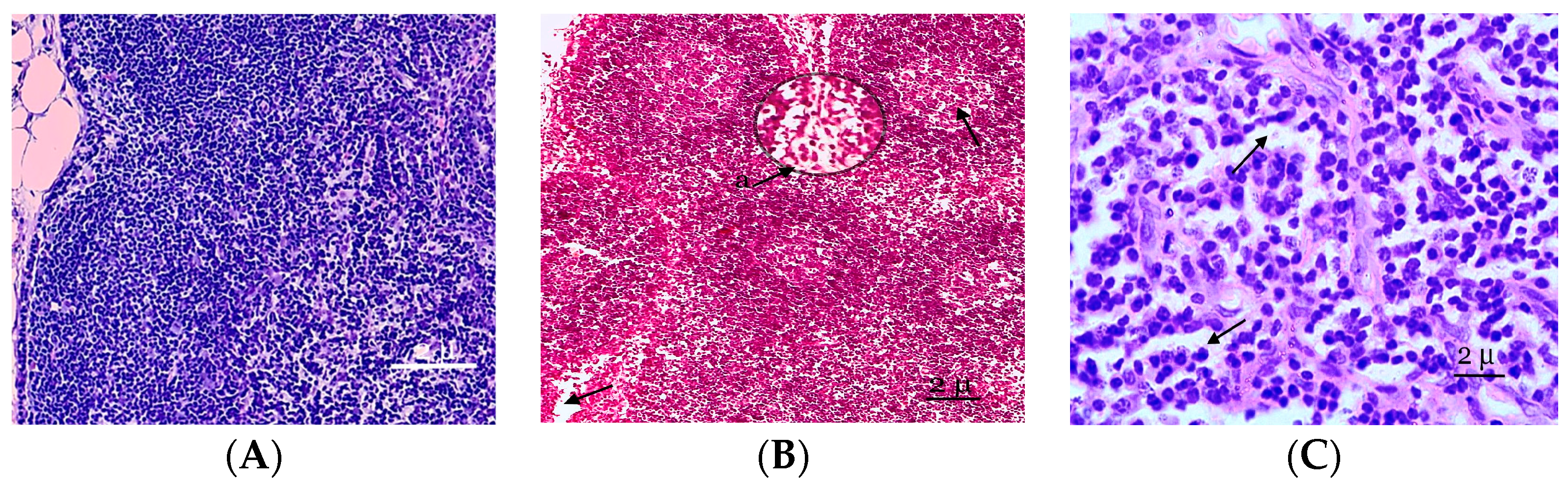
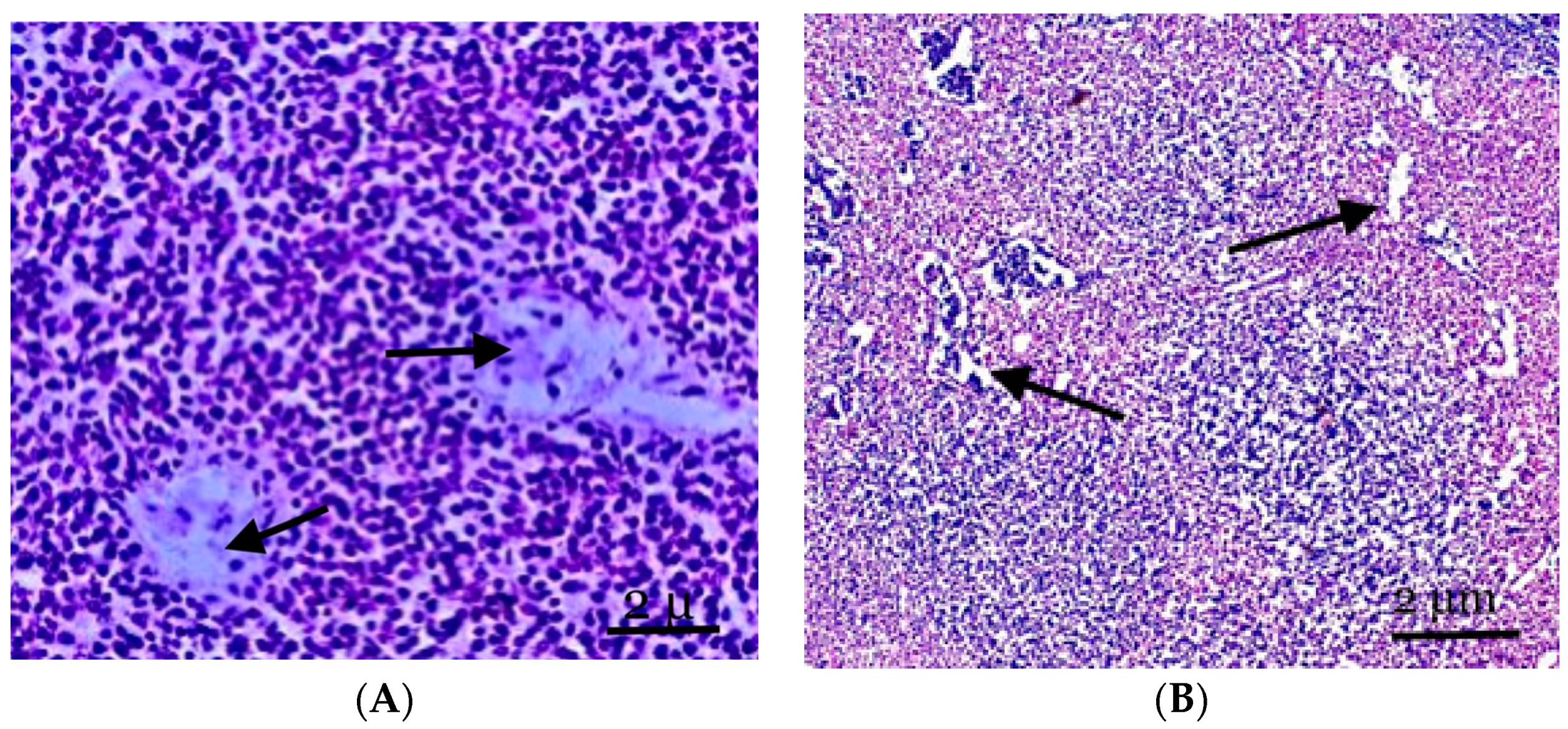
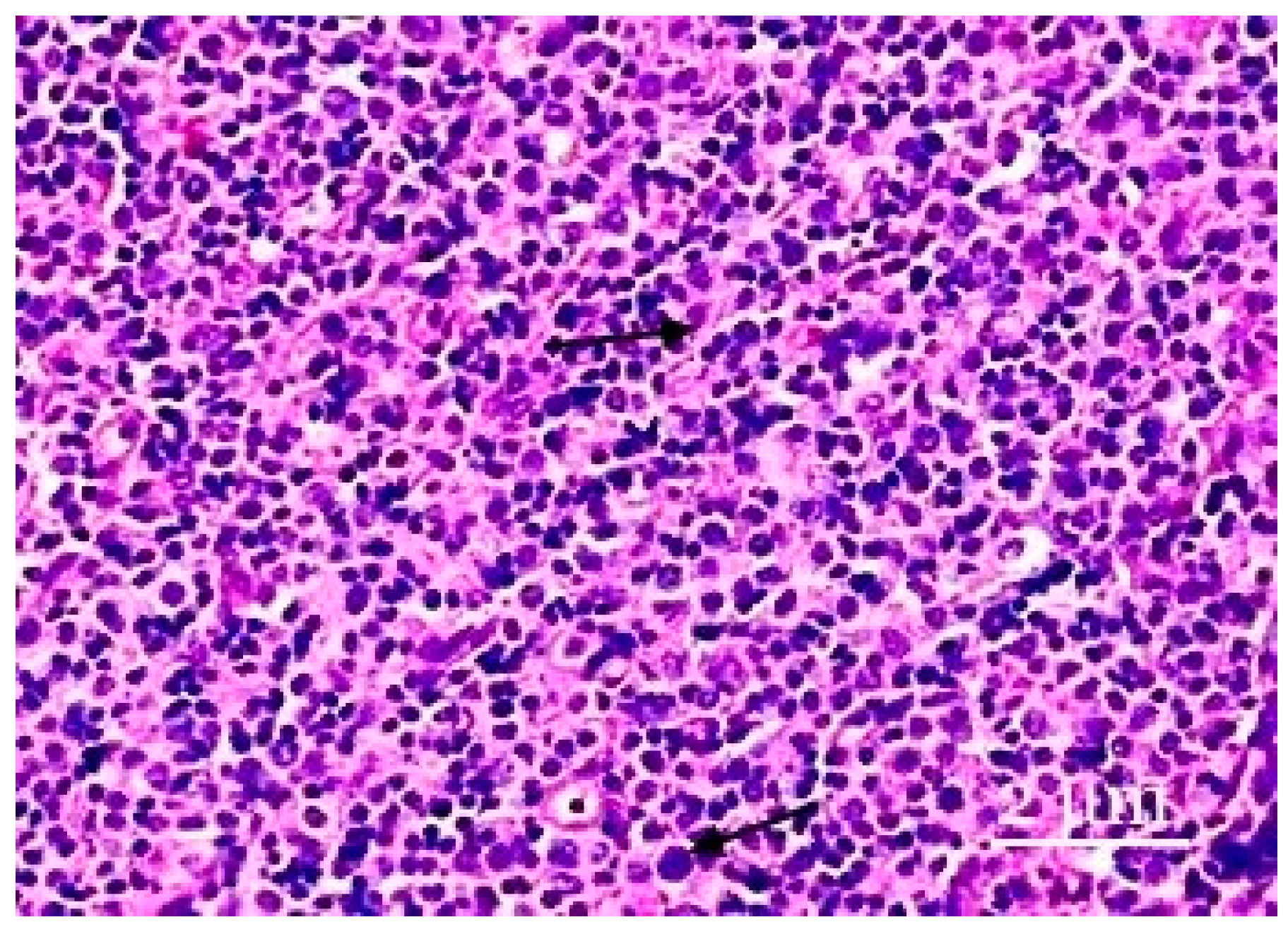
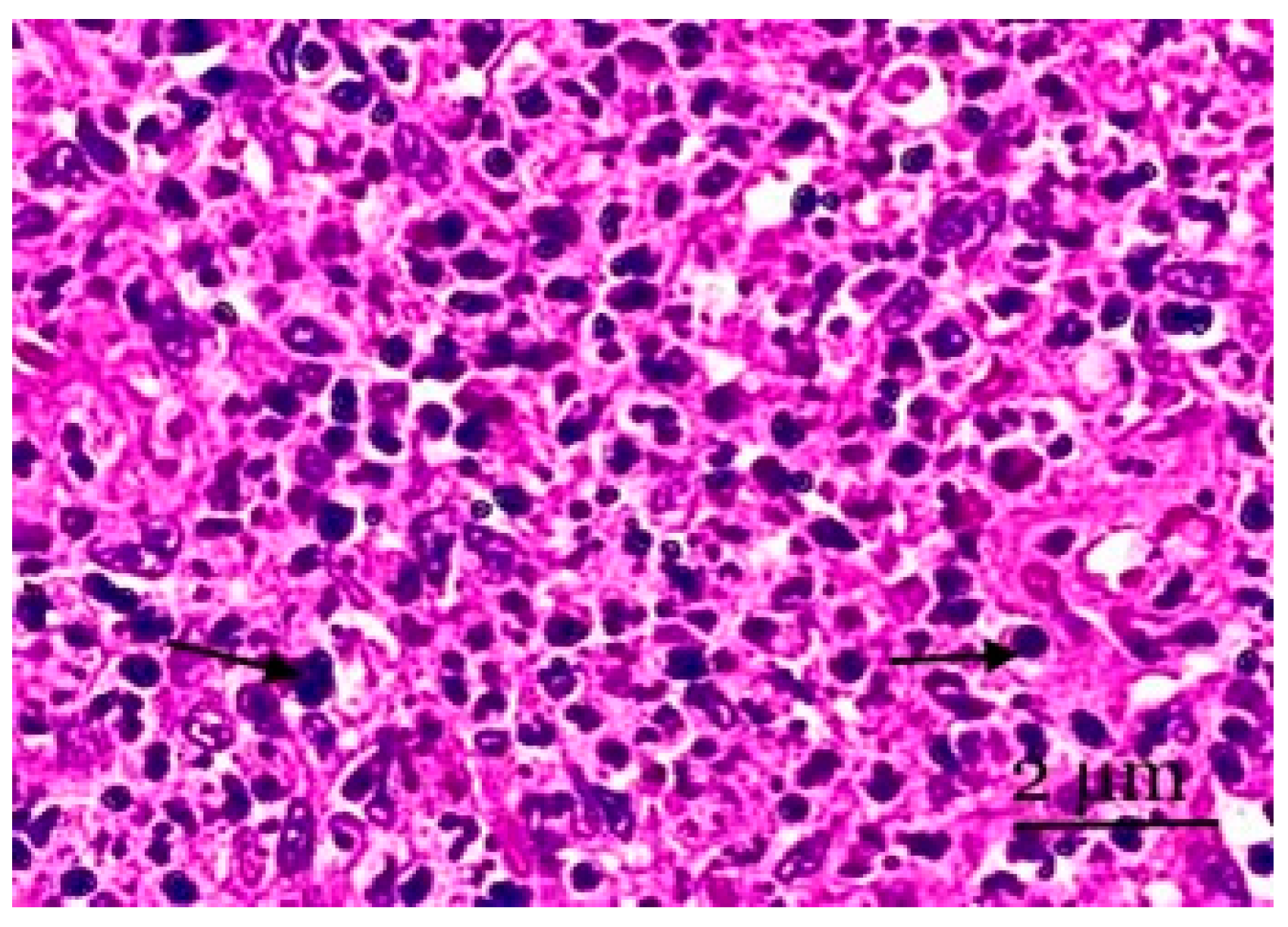
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silvia, M.; Francisco, O.N.; Kerstin, M.A.; Agneta, R.; Ricardo, F.; Miklos, L.; Anders, L.; Håkan, A.; Göran, H. Abdominal organ perfusion and inflammation in experimental sepsis: A magnetic resonance imaging study. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G187–G196. [Google Scholar]
- Khan, P.S.; Dar, L.A.; Hayat, H. Predictors of mortality and morbidity in peritonitis in a developing country. Turk. J. Surg. 2013, 29, 124–130. [Google Scholar] [CrossRef]
- Wacha, H.; Hau, T.; Dittmer, R.; Ohmann, C. Risk factors associated with intraabdominal infections: A prospective multicentre study. Peritonitis Study Group. Langenbeck’s Arch. Surg. 1999, 384, 24–32. [Google Scholar] [CrossRef]
- Karlos, A.O.; Juan, C.P. Management of Peritonitis in the Critically Ill Patient. Surg. Clin. N. Am. 2006, 86, 1323–1349. [Google Scholar]
- Bulesov, M.A.; Kausova, G.K.; Ashirov, B.O.; Ilimova, A.K. The structure of mortality in acute surgical diseases of the abdominal cavity and the role of endovideosurgical technology in its decline. Vestn. KazNMU Sci. Pract. J. Med. 2017, 4, 180–184. [Google Scholar]
- Lyust, V.I.; Igissinov, N.S.; Kozhakhmetov, S.K.; Abduov, M.; Zhumazhanov, N.; Bikhanov, N.; Toleubayev, M. Evaluation of changes in indicators of accident surgery in actue diseases of the abdominal cavity in the city of Astana (compared to 1998 from 2018). J. Med. 2019, 12, 24–28. [Google Scholar]
- Bagnenko, S.F.; Mirzabaev, A.T.; Batotsyrenov, B.V.; Gorbachev, N.B.; Miroshnichenko, V.N.; Batotsyrenova, K.V.; Veliky, K.F. Pharmacological correction of free radical disturbances and endotoxicosis in patients with diffuse peritonitis at the postoperative period. Vestn. Khirurgii Im. II Grek. 2011, 170, 4–16. [Google Scholar]
- Mathikere, L.R.; Bellary, J.; Sathees, B.C. Chandra Clinical Study and Management of Secondary Peritonitis due to Perforated Hollow Viscous. Arch. Med. Sci. 2007, 3, 61–68. [Google Scholar]
- Gonzalez, L.M.; Moeser, A.J.; Blikslager, A.T. Animal models of ischemia-reperfusion-induced intestinal injury: Progress and promise for translational research. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Volkov, D.V.; Stadnikov, B.A.; Tarasenko, V.S.; Basov, F.V. Influence anti-ischemic and antioxidant therapy on a morphofunctional state of the small intestine with the syndrome enteral insufficiency in experimental peritonitis. Orenbg. Med. Her. 2015, 3, 14–19. [Google Scholar]
- Srinivas, D. Intraoperative Lavage in Peritonitis–Comparison between Saline and Metronidazole. Int. J. Sci. Study 2020, 8, 92–95. [Google Scholar]
- Johansson, M.E.V.; Sjövall, H.; Hansson, G.C. The gastrointestinal mucus system in health and disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 352–361. [Google Scholar] [CrossRef]
- Llamas, M.-A.; Aller, M.-A.; Marquina, D.; Nava, M.-P.; Arias, J. Bacterial translocation to mesenteric lymph nodes increases in chronic portal hypertensive rats. Dig. Dis. Sci. 2010, 55, 2244–2254. [Google Scholar] [CrossRef]
- Bouma, G.; Strober, W. The immunological and genetic basis of inflammatory bowel disease. Nat. Rev. Immunol. 2003, 3, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Opal, S.M.; Garber, G.E.; LaRosa, S.P.; Maki, D.G.; Freebairn, R.C.; Kinasewitz, G.T.; Dhainaut, J.-F.; Yan, S.B.; Williams, M.D.; Graham, D.E.; et al. Systemic host responses in severe sepsis analyzed by causative microorganism and treatment effects of drotrecogin alfa (activated). Clin. Infect. Dis. 2003, 37, 50–58. [Google Scholar] [CrossRef]
- Maclntosh, T. Emergency Management spontaneous Bacterial peritonitis—A clinical review. Cureus 2018, 10, e2253. [Google Scholar]
- Nieuwenhuijzen, G.A.; Deitch, E.A.; Goris, R.J. The relationship between gut-derived bacteria and the development of the multiple organ dysfunction syndrome. J. Anat. 1996, 189, 537–548. [Google Scholar]
- Ruthrauff, C.M.; Smith, J.; Glerum, L. Primary bacterial septic peritonitis in cats: 13 cases. J. Am. Anim. Hosp. Assoc. 2009, 45, 268–276. [Google Scholar] [CrossRef]
- Oliver, G.; Kipnis, J.; Randolph, G.J.; Harvey, N.L. The Lymphatic Vasculature in the 21st Century: Novel Functional Roles in Homeostasis and Disease. Cell 2020, 182, 270–296. [Google Scholar] [CrossRef]
- Hollenberg, M.S.; Ahrens, T.S.; Djillali, A.; Astiz, M.E.; Chalfin, D.B.; Dasta, J.F.; Heard, S.O.; Martin, C.; Napolitano, L.M.; Susla, G.M.; et al. Practice parameters for hemodynamic support of sepsis in adult patients: 2004 update. Crit. Care Med. 2004, 32, 1928–1948. [Google Scholar] [CrossRef]
- Steffers, C.P.; Dahn, M.S.; Lange, M.P. Oxygen transport-dependent splanchnic metabolism in the sepsis syndrome. Arch. Surg. 1994, 129, 46–52. [Google Scholar] [CrossRef]
- Phinney, S.D. Fatty acids, inflammation, and the metabolic syndrome. Am. J. Clin. Nutr. 2005, 82, 1151–1152. [Google Scholar] [CrossRef]
- Grandl, G.; Wolfrum, C. Hemostasis, endothelial stress, inflammation, and the metabolic syndrome. Semin. Immunopathol. 2018, 40, 215–224. [Google Scholar] [CrossRef]
- Han, M.S.; White, A.; Perry, R.J.; Camporez, J.-P.; Hidalgo, J.; Shulman, G.I.; Davis, R.J. Regulation of adipose tissue inflammation by interleukin 6. Proc. Natl. Acad. Sci. USA 2020, 117, 2751–2760. [Google Scholar] [CrossRef]
- Cl`aria, J.; Dalli, J.; Yacoubian, S.; Gao, F.; Serhan, C.N. Resolvin D1 and resolvin D2 govern local inflammatory tone in obese fat 1. J. Immunol. 2012, 89, 2597–2605. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Panzarini, E.; Rajkovic, J.; Fokou, P.V.T.; Azzini, E.; Peluso, I.; Mishra, A.P.; Nigam, M.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Hokari, R.; Tomioka, A. The role of lymphatics in intestinal inflammation. Inflamm. Regen. 2021, 41, 25. [Google Scholar] [CrossRef]
- Nikolakis, D.; de Voogd, F.A.E.; Pruijt, M.J.; Grootjans, J.; van de Sande, M.G.; D’Haens, G.R. The Role of the Lymphatic System in the Pathogenesis and Treatment of Inflammatory Bowel Disease. Int. J. Mol. Sci. 2022, 23, 1854. [Google Scholar] [CrossRef]
- Lynskey, N.; Banerji, S.; Johnson, L.; Holder, K.; Reglinski, M.; Wing, P.; Rigby, D.; Jackson, D.; Sriskandan, S. Rapid Lymphatic Dissemination of Encapsulated Group A Streptococci via Lymphatic Vessel Endothelial Receptor-1 Interaction. PloS Pathog. 2015, 11, e1005137. [Google Scholar] [CrossRef]
- Földi, M.; Kubik, S. Lehrbuch der Lymphologie; Gustav Fischer Verlag: Stuttgart, Germany; Jena, Germany; New York, NY, USA, 1991; p. 225. [Google Scholar]
- Abdreshov, S.N.; Bulekbaeva, L.E.; Demchenko, G.A. Lympho- and Hemodynamics in Dogs with Acute Experimental Pancreatitis. Bull. Exp. Biol. Med. 2015, 159, 32–34. [Google Scholar] [CrossRef]
- Liao, S.; Weid, P.Y. Lymphatic System: An Active Pathway for Immune Protection. Semin. Cell Dev. Biol. 2015, 38, 83–89. [Google Scholar] [CrossRef]
- Gorchakova, O.; Gorchakov, V.; Kolmogorov, Y.; Nurmakhanova, B.; Demchenko, G.; Abdreshov, S. Microelement profile and structure of regional lymph nodes during senile in volution of lymphoid tissue. Arch. Euromedica 2021, 11, 48–51. [Google Scholar] [CrossRef]
- Bridenbaugh, E.A.; Nizamutdinova, I.T.; Jupiter, D.; Nagai, T.; Thangaswamy, S.; Chatterjee, V.; Gashev, A.A. Lymphatic muscle cells in rat mesenteric lymphatic vessels of various ages. Lymphat. Res. Biol. 2013, 11, 35–42. [Google Scholar] [CrossRef]
- Abdreshov, S.N.; Bulekbaeva, L.E.; Demchenko, G.A. Contractile activity of pectoral channel and lymph nodes in rats under antiorthostatic influences. Ross. Fiziol. Zhurnal Im. IM Sechenova 2011, 97, 509–514. [Google Scholar]
- Sapin, M.R. Lymphatic system and its significance in immune processes. Morfologiia 2007, 131, 18–22. [Google Scholar]
- Borodi, Y.I. Regional lymphatic drainage and lymphodetoxication. Morfologiia 2005, 128, 25–28. [Google Scholar]
- Deitch, E.A.; Xu, D.; Kaise, V.L. Role of the gut in the development of injury- and shock induced SIRS and MODS: The gut-lymph hypothesis, a review. Front. Biosci. 2006, 11, 520–528. [Google Scholar] [CrossRef]
- Lemole, G.M. The importance of the lymphatic system in vascular disease. J. Integr. Cardiol. 2016, 2, 413–416. [Google Scholar]
- Abdreshow, S.N.; Demshenko, G.A. Shifts in the lymph flow and the lymph composition in toxic hepatitis and their correction by protective substances. Bull. Exp. Biol. Med. 2009, 148, 403–405. [Google Scholar] [CrossRef]
- Kholova, I.; Dragneva, G.; Ermakova, P.; Laidinen, S.; Kaskenpää, N.; Hazes, T.; Cermáková, E.; Steiner, I.; Ylä-Herttuala, S. Lymphatic vasculature is increased in heart valves, ischaemic and inflamed hearts and in cholesterol-rich and calcified atherosclerotic lesions. Eur. J. Clin. Investig. 2011, 41, 487–497. [Google Scholar] [CrossRef]
- Abdreshov, S.N.; Demchenko, G.A.; Mamataeva, A.T.; Atanbaeva, G.K.; Mankibaeva, S.A.; Akhmetbaeva, N.A.; Kozhaniyazova, U.N.; Nauryzbai, U.B. Condition of adrenergic innervation apparatus of the thyroid gland, blood and lymph vessels, and lymph nodes during correction of hypothyrosis. Bull. Exp. Biol. Med. 2021, 171, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Mascena, G.V.; Filho, C.A.F.; de Lima Júnior, M.A.X.; Oliveira, T.K.B.; Gadelha, D.N.B.; Melo, M.C.S.C.; Brandt, C.T. Fecal peritonitis in aging rat model. Therapeutic response to different antibiotic strategies. Acta Cir. Bras. 2018, 33, 446–453. [Google Scholar] [CrossRef] [PubMed]
- Ham, A.W.; Cormack, D.H. Histology, 8th ed.; Lippincott Co.: Philadelphia, PA, USA; Toronto, ON, Canada, 1979. [Google Scholar]
- Merkulov, G.A. Course of Pathohistological Techniques; Medicine: Leningrad, Russia, 1969; 423p. [Google Scholar]
- Avtandilov, G.G. Medical Morphometry; Medicine: Moscow, Russia, 1990; 384p, SHBA.SPB.RU/YUZ. [Google Scholar]
- Khmelnitsky, O.K.; Tretyakova, M.S. Thyroid gland as an object of morphometric research. Arch. Pathol. 1998, 4, 47–49. [Google Scholar]
- Rogers, L.F. Magnetic resonance images of reactive lymphadenitis. Lymphology 2006, 39, 53–54. [Google Scholar] [PubMed]
- Reid, I.M. Morphometric Methods in Veterinary Pathology: A Review. Vet. Pathol. 1980, 17, 522–543. [Google Scholar] [CrossRef]
- Tanasiychuk, I.S. Cytomorphological characteristics of the cellular composition of the lymph nodes are normal. J. Cytol. Genet. 2004, 38, 60–66. [Google Scholar]
- Suttie, A.E. (Ed.) Boorman’s Pathology of the Rat: Reference and Atlas, 2nd ed.; Academic Press, an Imprint of Elsevier: London, UK; San Diego, CA, USA, 2018; 728p. [Google Scholar]
- Avtandilov, G.G. Fundamentals of Quantitative Pathological Anatomy. Moscow, Medicine. 2002. 240p. Available online: http://www.mmbook.ru (accessed on 25 February 2002).
- Harapko, T.; Mateshuk-Vatseba, L. Effects of MSG on the lymph nodes of the albino rat: Ultrastructural and morphometric studies. Eur. J. Anat. 2021, 25, 75–81. [Google Scholar]
- Johnson, L.A.; Clasper, S.; Holt, A.P.; Lalor, P.F.; Baban, D.; Jackson, D.G. An inflammation-induced mechanism for leukocyte transmigration across lymphatic vessel endothelium. J. Exp. Med. 2006, 203, 2763–2777. [Google Scholar] [CrossRef]
- Abdreshov, S.N.; Demchenko, G.A.; Gorchakov, V.N.; Yeshmukhanbet, A.N.; Yessenova, M.A. Lymph flow and cellular composition, rheological properties of lymph and blood in animals with experimental peritonitis. News NAS RK Ser. Biol. Med. 2021, 4, 5–13. [Google Scholar] [CrossRef]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Gut, G. Microbiota: Role in Pathogen Colonization, Immune Responses and Inflammatory Disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef]
- Földi, M.; Kubik, S. Lehrbuch der Lymphologie; Gustav Fischer Verlag: Stuttgart, Germany, 1989; 409p, EAN: 9783437112485. [Google Scholar]
- Zhdanov, D.A. General Anatomy and Physiology of the Lymphatic System; Medgiz: Leningrad, Russia, 1952. [Google Scholar]
- Kuka, M.; Iannacone, M. The role of lymph node sinus macrophages in host defense. Ann. N. Y. Acad. Sci. 2014, 1319, 38–46. [Google Scholar] [CrossRef]
- Benahmed, F.; Ely, S.; Lu, T. Lymph node vascular-stromal growth and function as a potential target for controlling immunity. Clin. Immunol. 2012, 144, 109–116. [Google Scholar] [CrossRef]
- Gray, E.E.; Cyster, J.G. Lymph Node Macrophages. J. Innate Immun. 2012, 4, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Kastenmuller, W.; Torabi-Parizi, P.; Subramanian, N.; Lammermann, T.; Germain, R.N. A spatially-organized multicellular innate immune response in lymph nodes limits systemic pathogen spread. Cell 2012, 150, 1235–1248. [Google Scholar] [CrossRef] [PubMed]
- Bajénoff, M.; Glaichenhaus, N.; Germain, R.N. Fibroblastic reticular cells guide T lymphocyte entry into and migration within the splenic T cell zone. J. Immunol. 2008, 181, 3947–3954. [Google Scholar] [CrossRef] [PubMed]
- Von Der Weid, P.-Y.; Rainey, K.J. Review article: Lymphatic system and associated adipose tissue in the development of inflammatory bowel disease. Aliment. Pharmacol. Ther. 2010, 32, 697–711. [Google Scholar] [CrossRef]
- Abdreshov, S.N.; Bulekbayeva, L.E.; Demshenko, G.A. Lymph Flow and Contractile Activity of Mesenteric Lymph Nodes in Rats with Toxic Hepatitis Effects of Antioxidants. Bull. Exp. Biol. Med. 2013, 155, 22–25. [Google Scholar] [CrossRef]
- Bogoslowski, A.; Kubes, P. Lymph Nodes: The Unrecognized Barrier against Pathogens. ACS Infect. Dis. 2018, 4, 1158–1161. [Google Scholar] [CrossRef]
- Willard-Mack, C.L. Normal structure, function, and histology of lymph nodes. Toxicol. Pathol. 2006, 34, 409–424. [Google Scholar] [CrossRef]
- Yücel, Y.H.; Johnston, M.G.; Ly, T.; Patel, M.; Drake, B.; Gümüş, E.; Fraenkl, S.A.; Moore, S.; Tobbia, D.; Armstrong, D.; et al. Identification of lymphatics in the ciliary body of the human eye: A novel “uveolymphatic” outflow pathway. Exp. Eye Res. 2009, 89, 810–819. [Google Scholar] [CrossRef]
- Abdreshov, S.N.; Akhmetbaeva, N.A.; Atanbaeva, G.K.; Mamataeva, A.T.; Nauryzbai, U.B. Adrenergic Innervation of the Thyroid Gland, Blood and Lymph Vessels, and Lymph Nodes in Hypothyroidism. Bull. Exp. Biol. Med. 2013, 168, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Mateshuk-Vatseba, L.R.; Holovatskyi, A.S.; Harapko, T.V.; Foros, A.I.; Lytvak, Y.V. Changes in the structural organization of lymph nodes duringshort-term exposure to monosodium glutamate. Rep. Morphol. 2022, 28, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Yeo, K.P.; Angeli, V. Bidirectional crosstalk between lymphatic endothelial cell and T cell and its implications in tumor immunity. Front. Immunol. 2017, 8, 83. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Structures of the Mesenteric Lymph Nodes | Control | Two Days after Inflammation | Five Days after Inflammation |
|---|---|---|---|
| 1 | 2 | 3 | |
| Capsule | 0.98 ± 0.07 | 1.13 ± 0.08 | 1.18 ± 0.04 |
| Subcapsular sinus | 0.14 ± 0.04 | 0.51 ± 0.09 ** | 0.45 ± 0.06 ** |
| Cortical plateau | 1.06 ± 0.07 | 0.70 ± 0.04 * | 0.71 ± 0.08 * |
| Lymphoid nodule without a germinal center (F1) | 1.33 ± 0.011 | 1.72 ± 0.05 | 1.68 ± 0.13 *• |
| Lymphoid nodule with germinal center (F2) | 1.72 ± 0.02 | 1.02 ± 0.07 * | 0.93 ± 0.02 *• |
| Paracortex | 4.46 ± 0.35 | 3.27 ± 0.42 * | 3.11 ± 0.21 * |
| Pulp strands | 5.41 ± 0.12 | 4.78 ± 0.08 * | 6.74 ± 0.18 *• |
| Cerebral sinus | 2.5 ± 0.11 | 4.35 ± 0.25 * | 3.23 ± 0.25 *• |
| Total area | 17.58 ± 1.99 | 19.51 ± 2.11 | 17.98 ± 2.12 |
| Cortical brain index | 0.84 ± 0.013 | 0.51 ±0.16 | 0.49 ± 0.20 |
| F2/F1 | 1.29 ± 0.012 | 0.59 ± 0.04 | 0.58 ± 0.06 |
| T-zone | 5.52 ± 0.21 | 3.97 ± 0.23 * | 3.89 ± 0.29 * |
| B-zone | 8.46 ± 0.35 | 7.51 ± 0.38 | 8.52 ± 0.24 |
| Index T/B | 0.65 ± 0.26 | 0.53 ± 0.29 | 0.46 ± 0.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdreshov, S.N.; Demchenko, G.A.; Yeshmukhanbet, A.N.; Yessenova, M.A.; Mankibaeva, S.A.; Atanbaeva, G.K.; Kulbayeva, M.S. Morphofunctional Alteration of Mesenteric Lymph Nodes in the Inflammation of the Abdominal Cavity. Biology 2024, 13, 166. https://doi.org/10.3390/biology13030166
Abdreshov SN, Demchenko GA, Yeshmukhanbet AN, Yessenova MA, Mankibaeva SA, Atanbaeva GK, Kulbayeva MS. Morphofunctional Alteration of Mesenteric Lymph Nodes in the Inflammation of the Abdominal Cavity. Biology. 2024; 13(3):166. https://doi.org/10.3390/biology13030166
Chicago/Turabian StyleAbdreshov, Serik N., Georgii A. Demchenko, Anar N. Yeshmukhanbet, Makpal A. Yessenova, Sandugash A. Mankibaeva, Gulshat K. Atanbaeva, and Marzhan S. Kulbayeva. 2024. "Morphofunctional Alteration of Mesenteric Lymph Nodes in the Inflammation of the Abdominal Cavity" Biology 13, no. 3: 166. https://doi.org/10.3390/biology13030166