
Soft-Tissue, Rare Earth Element, and Molecular Analyses of Dreadnoughtus schrani, an Exceptionally Complete Titanosaur from Argentina

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
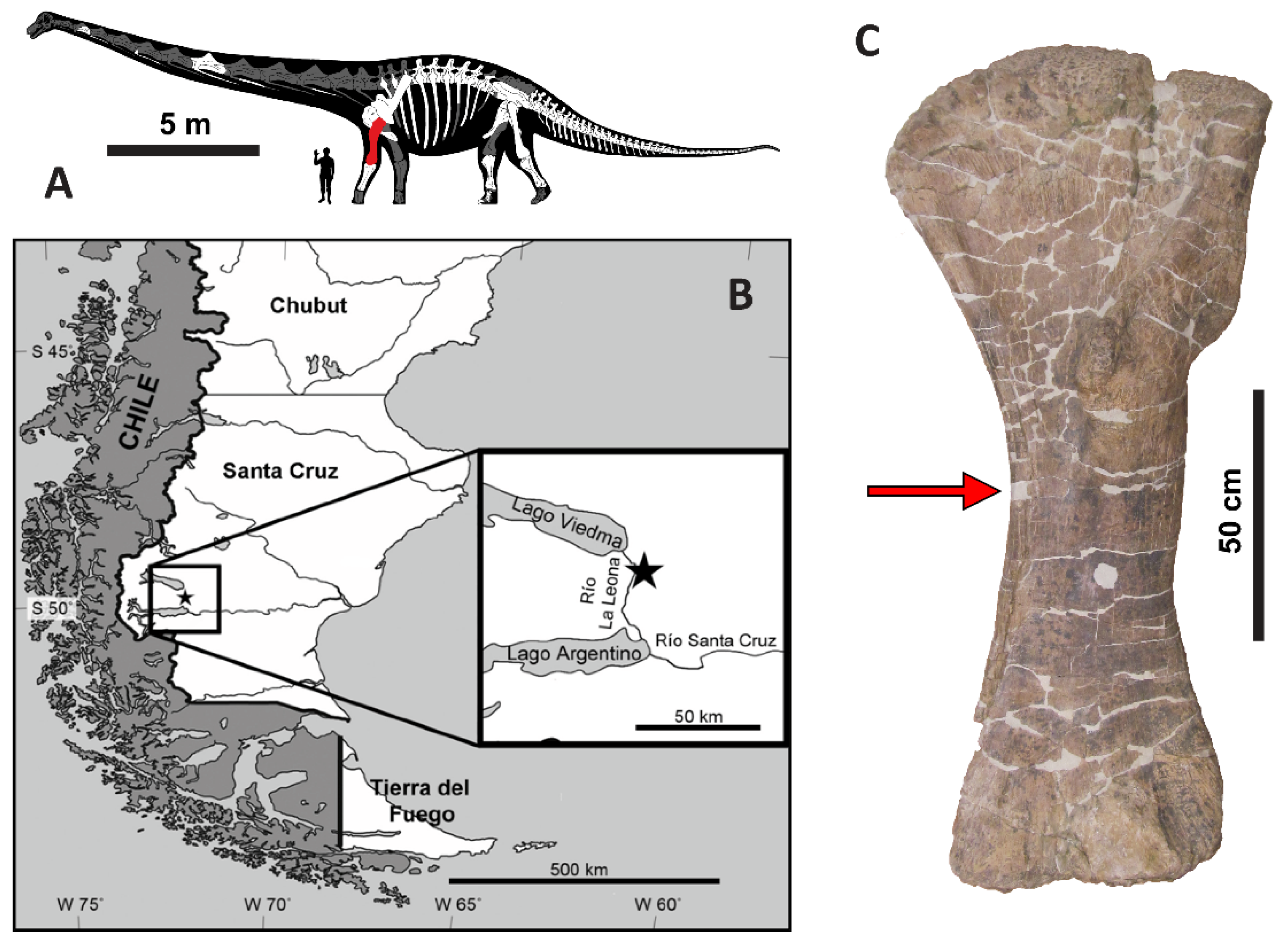
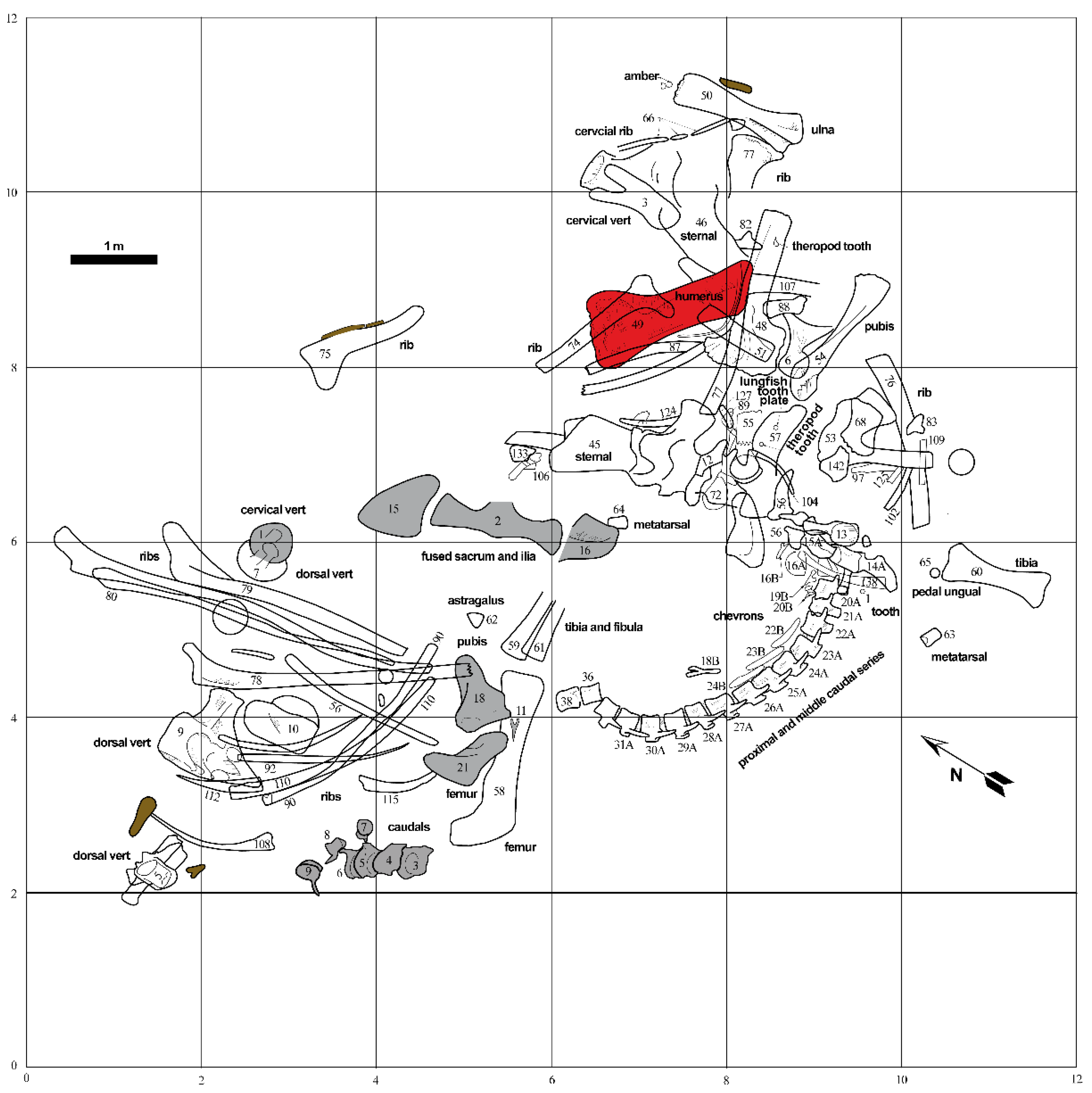
2. Geologic and Taphonomic Context
3. Methods
3.1. Sample Collection
3.2. Assessments of Preservational Quality and Geochemical History
3.2.1. Histological Analysis
3.2.2. X-ray Diffraction (XRD)
3.2.3. Laser Ablation Inductively Coupled Plasma Mass Spectrometry (LA-ICPMS)
3.3. Assessments of Soft Tissue Preservation
3.3.1. Demineralization and Evaluation of Morphological Structures
3.3.2. Immunofluorescence (IF)
3.3.3. Enzyme-Linked Immunosorbent Assay (ELISA)
4. Results
4.1. Preservational Quality
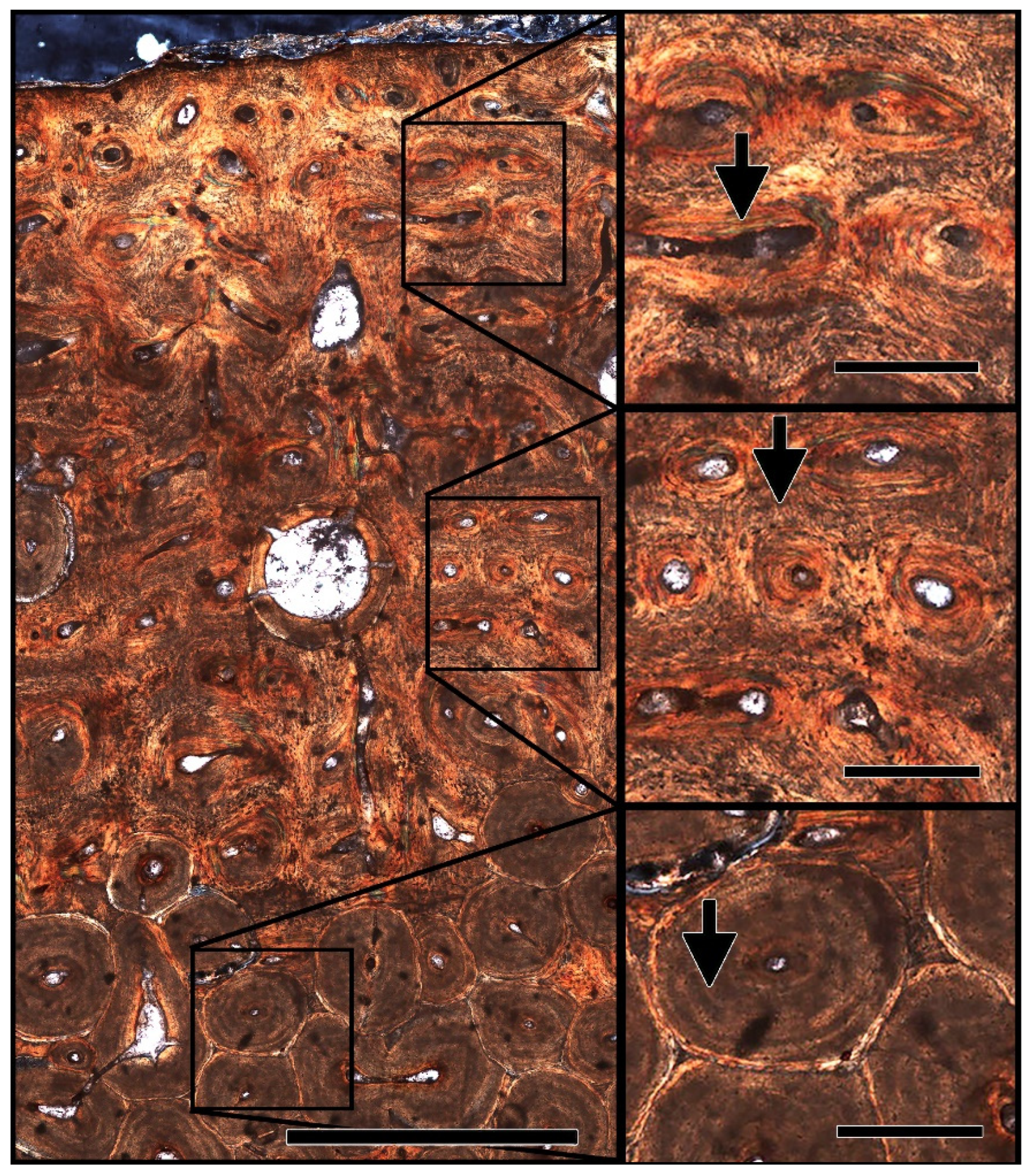
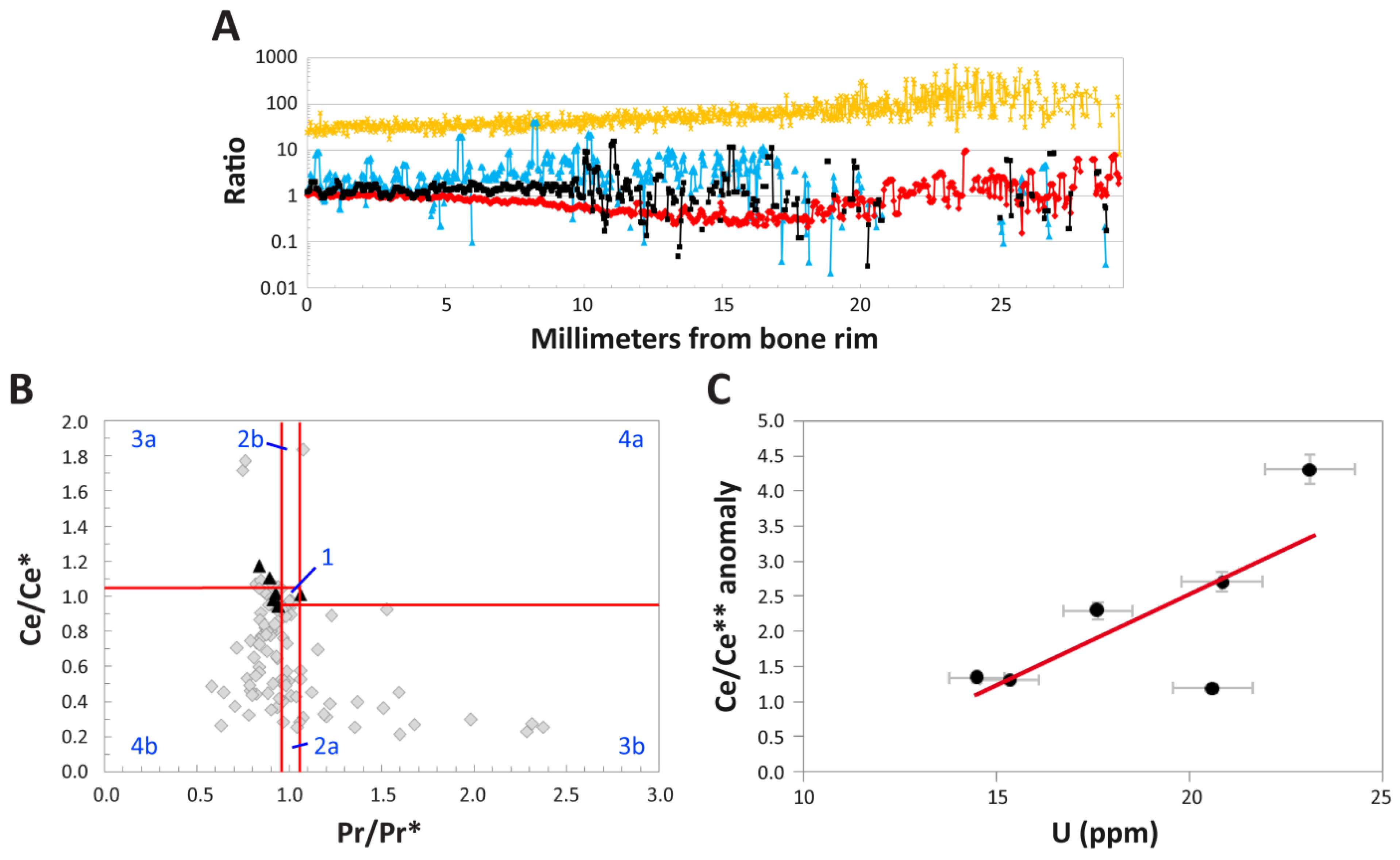
4.1.1. Histological Analysis
4.1.2. X-ray Diffraction (XRD)
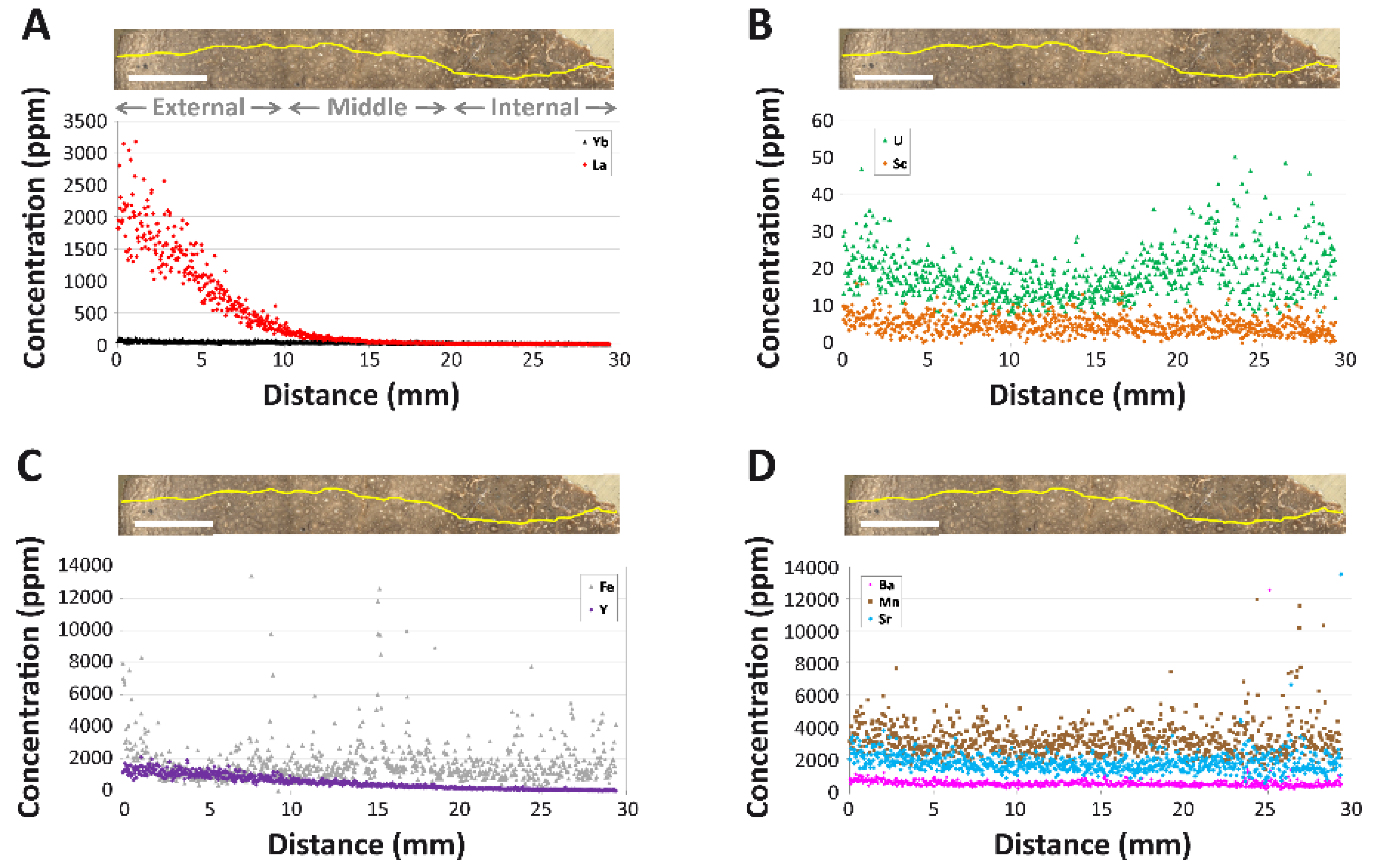
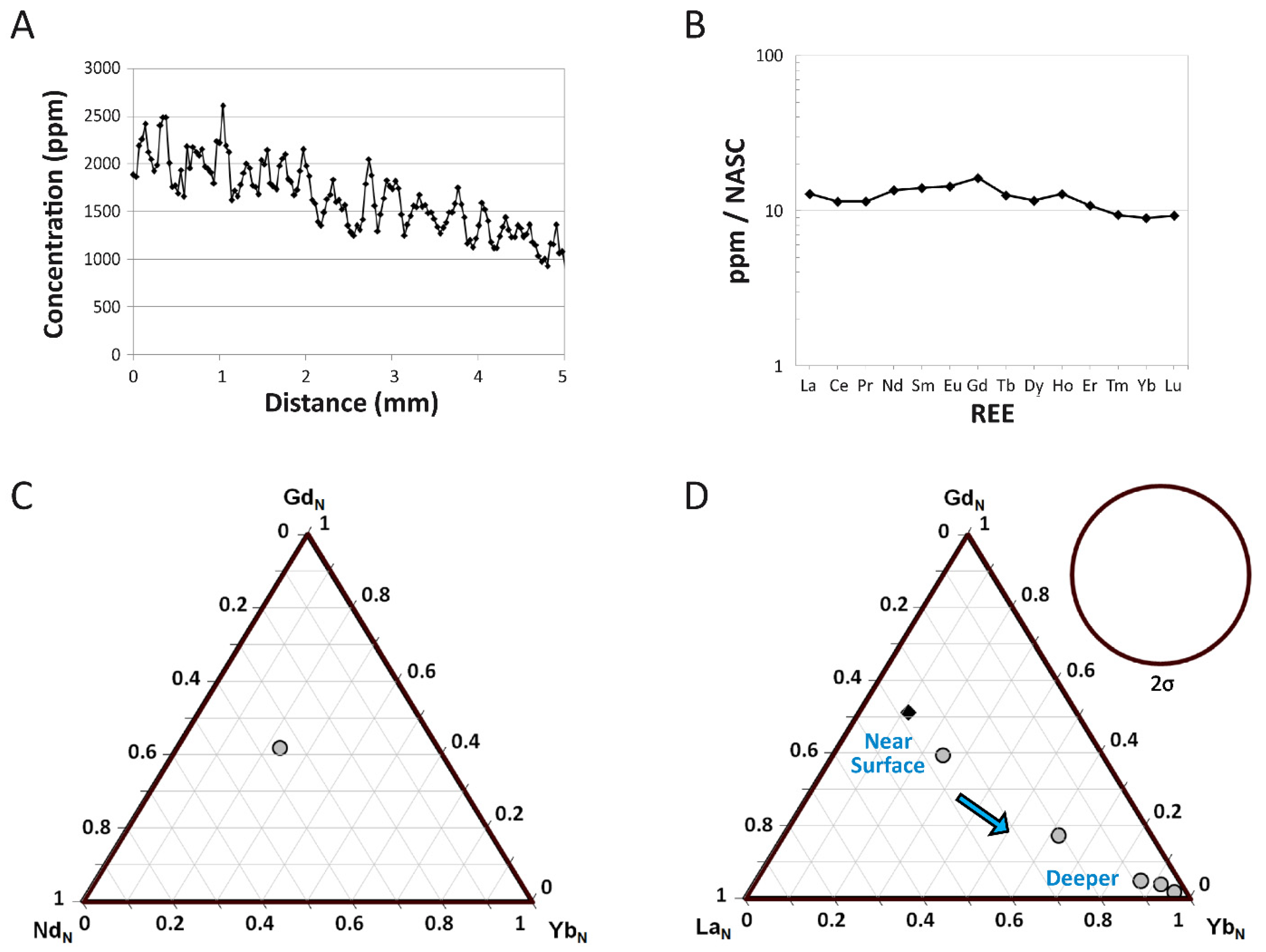
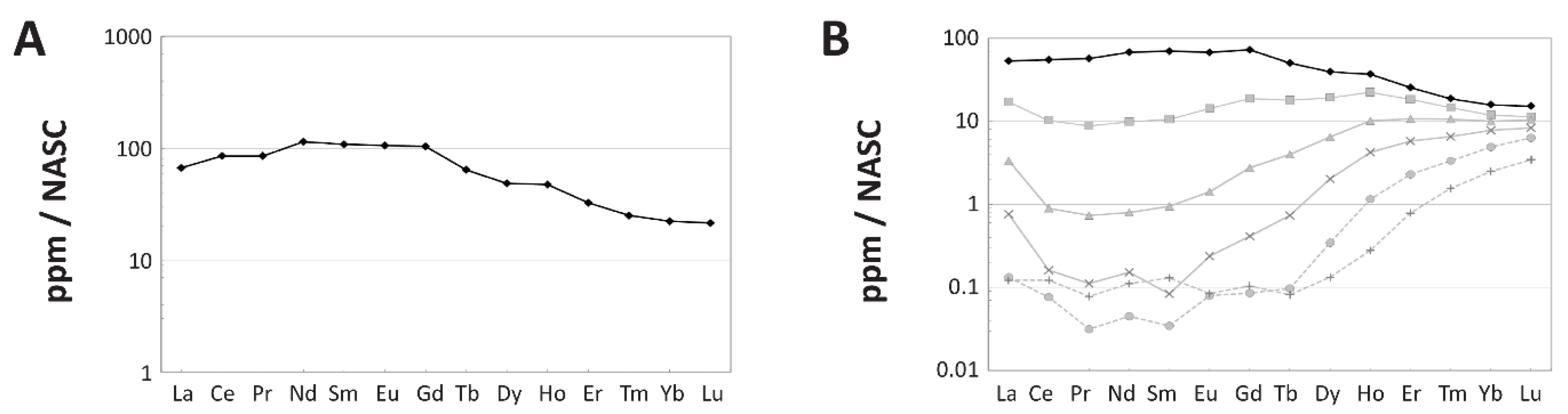
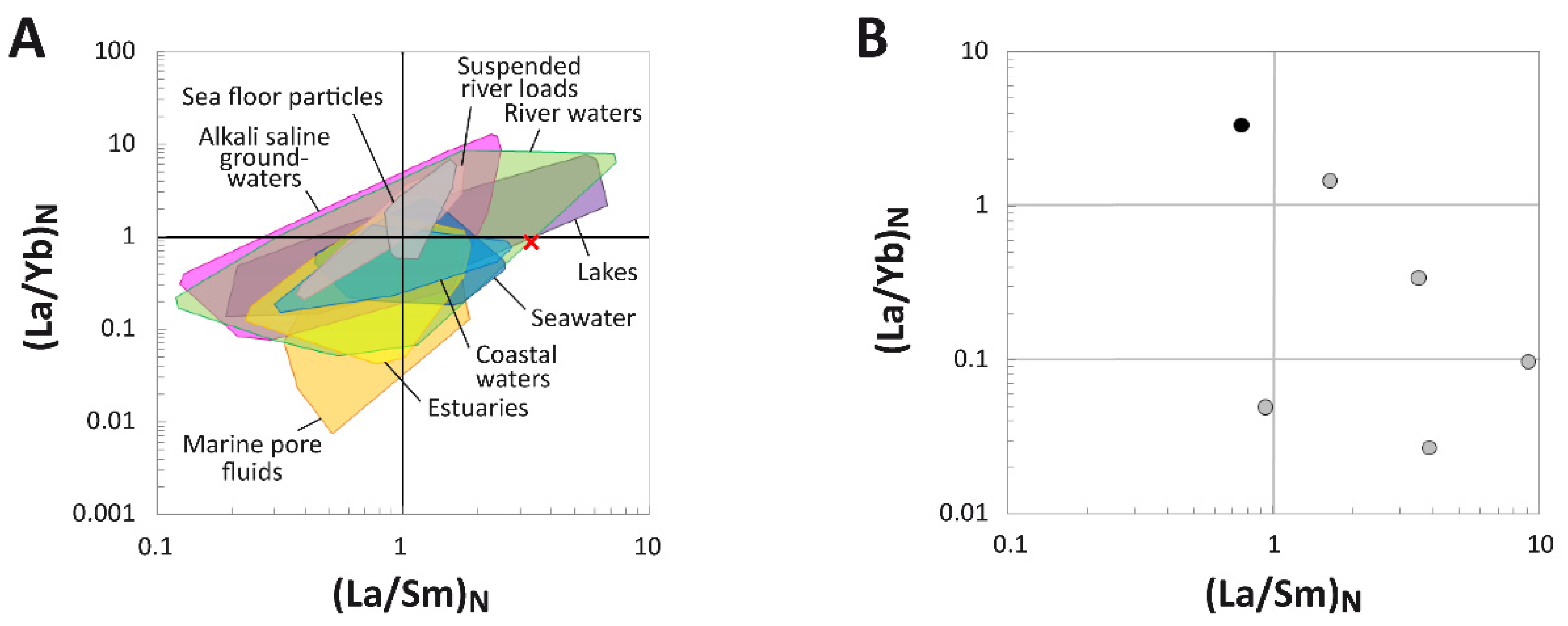
4.1.3. REE Analyses
4.2. Soft Tissue Preservation
4.2.1. Demineralization/Morphological Structures
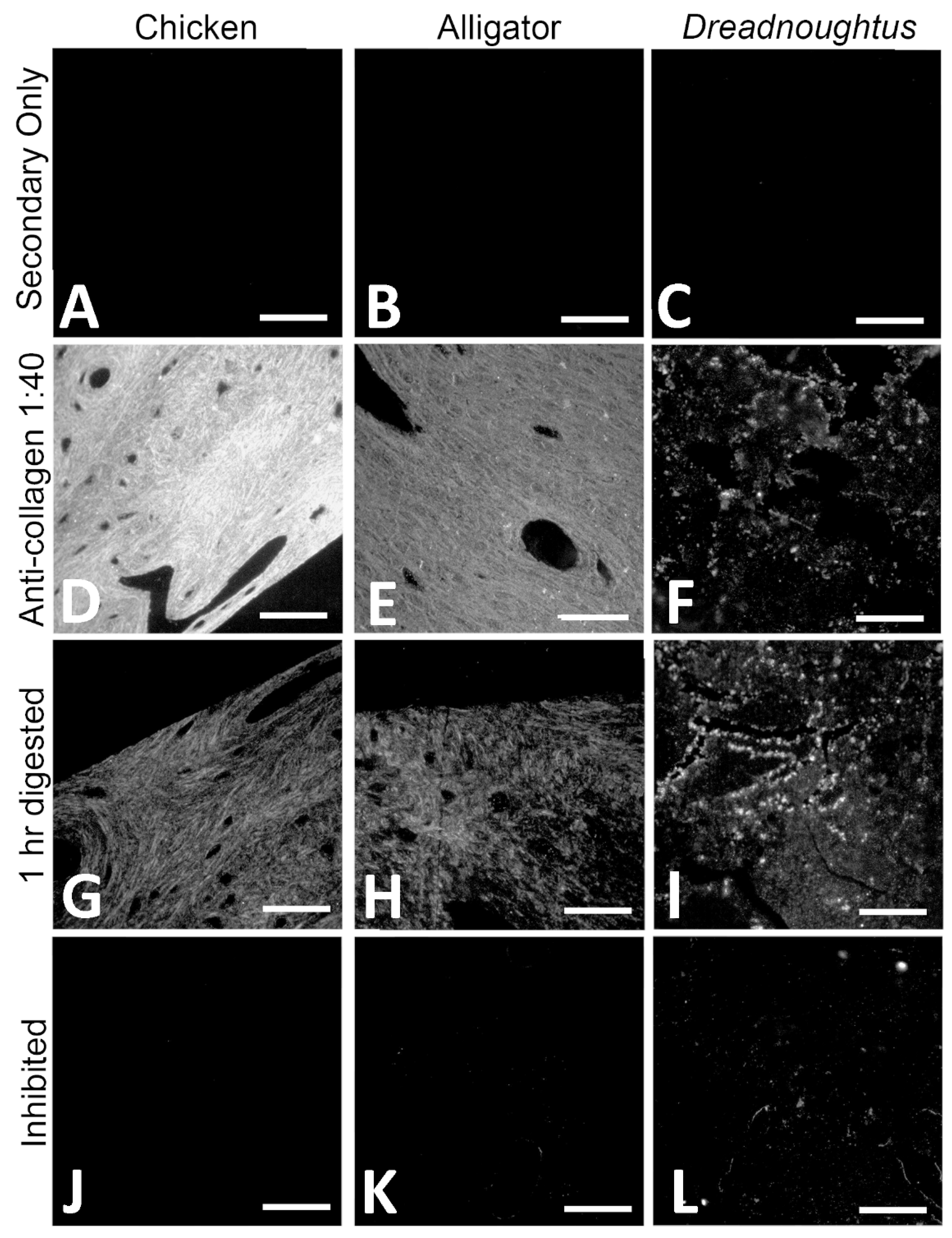
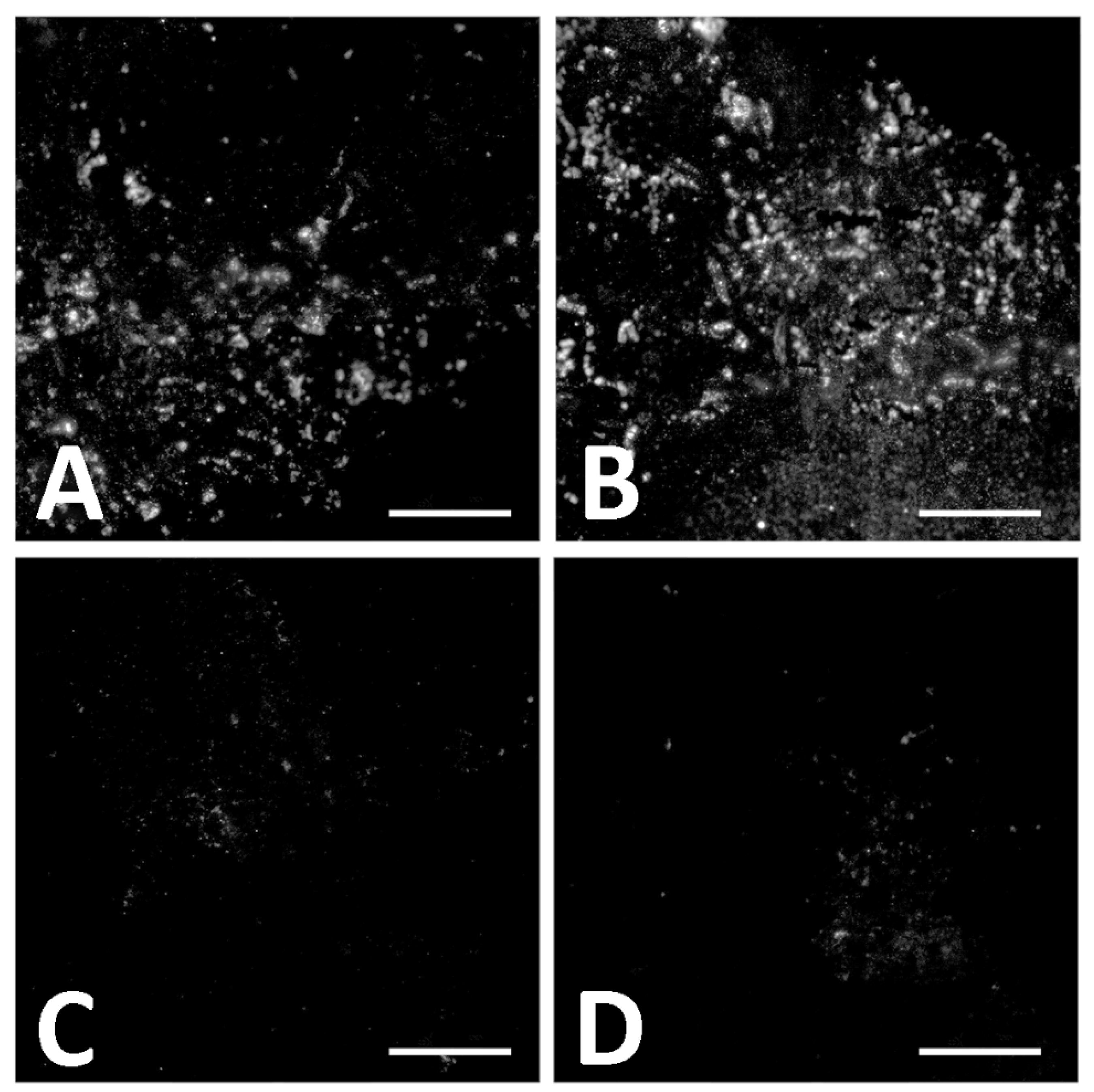
4.2.2. Immunofluorescence (IF)
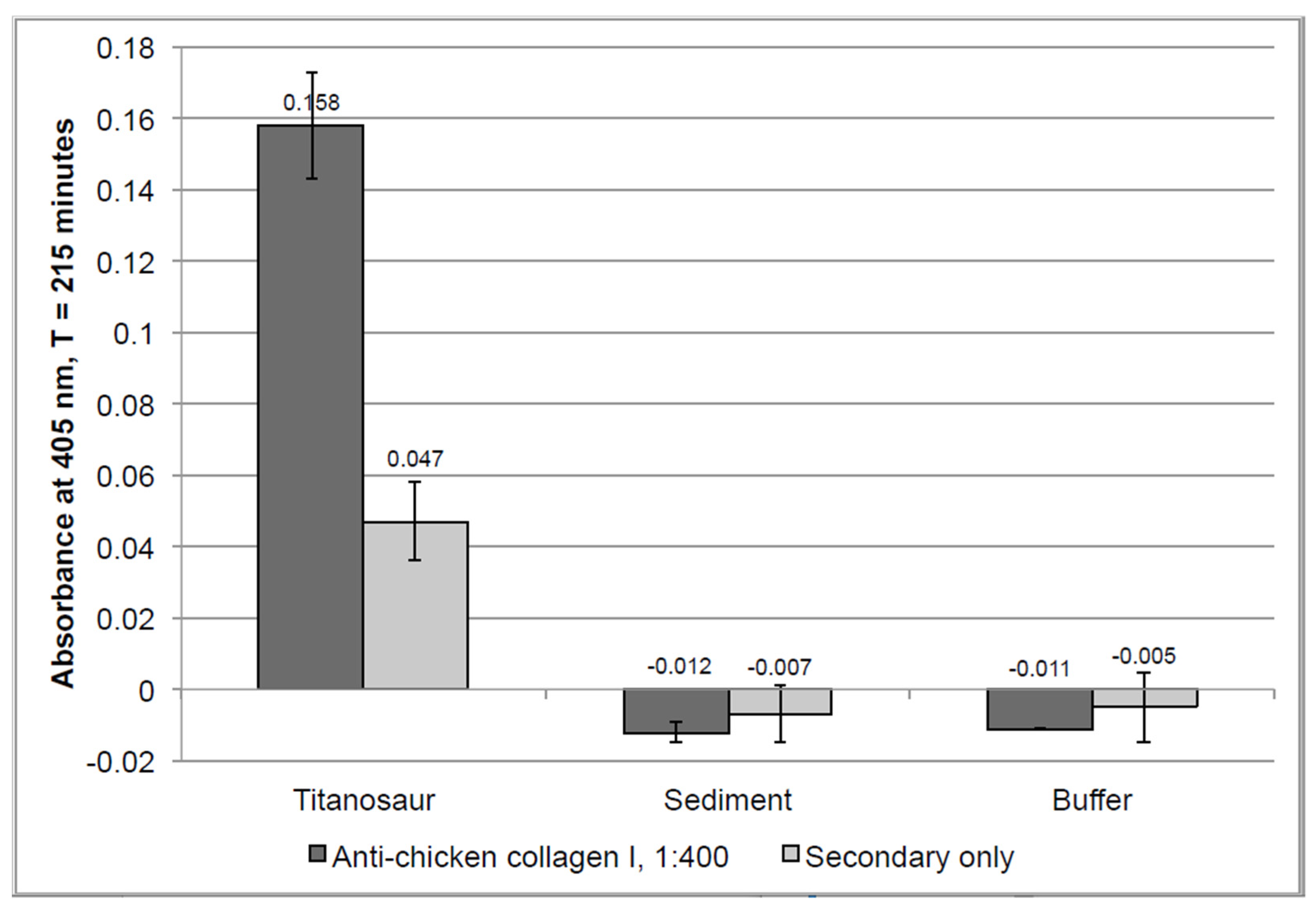
4.2.3. Enzyme-Linked Immunosorbent Assay (ELISA)
5. Discussion
5.1. Reconstructing the Geochemical History of MPM-PV 1156
5.2. Evaluating the Preservation of Soft Tissues
5.3. Implications for Future Paleomolecular Studies
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gioncada, A.; Collareta, A.; Gariboldi, K.; Lambert, O.; Di Celma, C.; Bonaccorsi, E.; Urbina, M.; Bianucci, G. Inside baleen: Exceptional microstructure preservation in a late Miocene whale skeleton from Peru. Geology 2016, 44, 839–842. [Google Scholar] [CrossRef]
- McCoy, V.E.; Gabbott, S.E.; Penkman, K.; Collins, M.J.; Presslee, S.; Holt, J.; Grossman, H.; Wang, B.; Solórzano, M.M.; Delclòs, X.; et al. Ancient amino acids from fossil feathers in amber. Sci. Rep. 2019, 9, 6420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiemann, J.; Fabbri, M.; Yang, T.R.; Stein, K.; Sander, P.M.; Norell, M.A.; Briggs, D.E.G. Fossilization transforms vertebrate hard tissue proteins into N-heterocyclic polymers. Nat. Commun. 2018, 9, 4741. [Google Scholar] [CrossRef] [Green Version]
- Wiemann, J.; Crawford, J.M.; Briggs, D.E.G. Phylogenetic and physiological signals in metazoan fossil biomolecules. Sci. Adv. 2020, 6, eaba6883. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Chiang, C.C.; Huang, P.Y.; Chung, C.Y.; Huang, T.D.; Wang, C.C.; Chen, C.I.; Chang, R.S.; Liao, C.H.; Reisz, R.R. Evidence of preserved collagen in an Early Jurassic sauropodomorph dinosaur revealed by synchrotron FTIR microspectroscopy. Nat. Commun. 2017, 8, 2–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reisz, R.R.; Huang, T.D.; Roberts, E.M.; Peng, S.; Sullivan, C.; Stein, K.; Leblanc, A.R.H.; Shieh, D.; Chang, R.; Chiang, C.; et al. Embryology of Early Jurassic dinosaur from China with evidence of preserved organic remains. Nature 2013, 496, 210–214. [Google Scholar] [CrossRef]
- Boatman, E.M.; Goodwin, M.B.; Holman, H.Y.N.; Fakra, S.; Zheng, W.; Gronsky, R.; Schweitzer, M.H. Mechanisms of soft tissue and protein preservation in Tyrannosaurus rex. Sci. Rep. 2019, 9, 15678. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Zheng, W.; Moyer, A.E.; O’Connor, J.K.; Wang, M.; Zheng, X.; Wang, X.; Schroeter, E.R.; Zhou, Z.; Schweitzer, M.H. Molecular evidence of keratin and melanosomes in feathers of the Early Cretaceous bird Eoconfuciusornis. Proc. Natl. Acad. Sci. USA 2016, 113, E7900–E7907. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Zheng, W.; Sawyer, R.H.; Pennington, M.W.; Zheng, X.; Wang, X.; Wang, M.; Hu, L.; O’Connor, J.; Zhao, T.; et al. The molecular evolution of feathers with direct evidence from fossils. Proc. Natl. Acad. Sci. USA 2019, 116, 3018–3023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, B.; Zhao, T.; Regnault, S.; Edwards, N.P.; Kohn, S.C.; Li, Z.; Wogelius, R.A.; Benton, M.J.; Hutchinson, J.R. Cellular preservation of musculoskeletal specializations in the Cretaceous bird Confuciusornis. Nat. Commun. 2017, 8, 14779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindgren, J.; Sjövall, P.; Thiel, V.; Zheng, W.; Ito, S.; Wakamatsu, K.; Hauff, R.; Kear, B.P.; Engdahl, A.; Alwmark, C.; et al. Soft-tissue evidence for homeothermy and crypsis in a Jurassic ichthyosaur. Nature 2018, 564, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Surmik, D.; Boczarowski, A.; Balin, K.; Dulski, M.; Szade, J.; Kremer, B.; Pawlicki, R. Spectroscopic studies on organic matter from triassic reptile bones, Upper Silesia, Poland. PLoS ONE 2016, 11, e0151143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweitzer, M.H.; Zheng, W.; Organ, C.L.; Avci, R.; Suo, Z.; Freimark, L.M.; Lebleu, V.S.; Duncan, M.B.; Heiden, M.G.V.; Neveu, J.M.; et al. Biomolecular characterization and protein sequences of the campanian hadrosaur b. canadensis. Science 2009, 324, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Schroeter, E.R.; DeHart, C.J.; Cleland, T.P.; Zheng, W.; Thomas, P.M.; Kelleher, N.L.; Bern, M.; Schweitzer, M.H. Expansion for the Brachylophosaurus canadensis Collagen I Sequence and Additional Evidence of the Preservation of Cretaceous Protein. J. Proteome Res. 2017, 16, 920–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckley, M.; Warwood, S.; van Dongen, B.; Kitchener, A.C.; Manning, P.L. A fossil protein chimera; difficulties in discriminating dinosaur peptide sequences from modern cross-contamination. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170544. [Google Scholar] [CrossRef]
- Schweitzer, M.H.; Zheng, W.; Cleland, T.P.; Goodwin, M.B.; Boatman, E.; Theil, E.; Marcus, M.A.; Fakra, S.C. A role for iron and oxygen chemistry in preserving soft tissues, cells and molecules from deep time. Proc. R. Soc. Ser. B 2014, 281, 20132741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwalt, D.E.; Goreva, Y.S.; Siljeström, S.M.; Rose, T.; Harbach, R.E. Hemoglobin-derived porphyrins preserved in a Middle Eocene blood-engorged mosquito. Proc. Natl. Acad. Sci. USA 2013, 110, 18496–18500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, J.E.; Lenczewski, M.E.; Scherer, R.P. Influence of microbial biofilms on the preservation of primary soft tissue in fossil and extant archosaurs. PLoS ONE 2010, 5, e13334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweitzer, M.H. Soft tissue preservation in terrestrial Mesozoic vertebrates. Annu. Rev. Earth Planet. Sci. 2011, 39, 187–216. [Google Scholar] [CrossRef]
- van Klinken, G.J.; Hedges, R.E.M. Experiments on Collagen-Humic Interactions: Speed of Humic Uptake, and Effects of Diverse Chemical Treatments. J. Archaeol. Sci. 1995, 22, 263–270. [Google Scholar] [CrossRef]
- Hedges, R.E.M. Bone diagenesis: An overview of processes. Archaeometry 2002, 44, 319–328. [Google Scholar] [CrossRef]
- Nielsen-Marsh, C.M.; Richards, M.P.; Hauschka, P.V.; Thomas-Oates, J.E.; Trinkaus, E.; Pettitt, P.B.; Karavanić, I.; Poinar, H.; Collins, M.J. Osteocalcin protein sequences of Neanderthals and modern primates. Proc. Natl. Acad. Sci. USA 2005, 102, 4409–4413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleland, T.P.; Schroeter, E.R. A Comparison of Common Mass Spectrometry Approaches for Paleoproteomics. J. Proteome Res. 2018, 17, 936–945. [Google Scholar] [CrossRef] [PubMed]
- Schroeter, E.R.; Cleland, T.P.; Schweitzer, M.H. Deep Time Paleoproteomics: Looking Forward. J. Proteome Res. 2022, 21, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Lacovara, K.J.; Lamanna, M.C.; Ibiricu, L.M.; Poole, J.C.; Schroeter, E.R.; Ullmann, P.V.; Voegele, K.K.; Boles, Z.M.; Carter, A.M.; Fowler, E.K.; et al. A gigantic, exceptionally complete titanosaurian sauropod dinosaur from southern Patagonia, Argentina. Sci. Rep. 2014, 4, 6196. [Google Scholar] [CrossRef] [Green Version]
- Curry Rogers, K. Titanosauria: A phylogenetic overview. In The Sauropods: Evolution and Paleobiology; Curry Rogers, K., Wilson, J.A., Eds.; University of California Press: Berkeley, CA, USA, 2005; pp. 50–103. ISBN 978-0-520-93233-3. [Google Scholar]
- Lingham-Soliar, T. A unique cross section through the skin of the dinosaur Psittacosaurus from China showing a complex fibre architecture. Proc. R. Soc. B Biol. Sci. 2008, 275, 775–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweitzer, M.H.; Horner, J.R.H. Intravascular microstructures in trabecular bone tissues of Tyrannosaurus rex. Ann. Paleontol. 1999, 85, 179–192. [Google Scholar] [CrossRef]
- Eglinton, G.; Logan, G.A. Molecular preservation. Philos. Trans.-R. Soc. Lond. B 1991, 333, 315–328. [Google Scholar] [CrossRef]
- Chin, K.; Eberth, D.A.; Schweitzer, M.H.; Rando, T.A.; Sloboda, W.J.; Horner, J.R. Remarkable Preservation of Undigested Muscle Tissue within a Late Cretaceous Tyrannosaurid Coprolite from Alberta, Canada. Palaios 2003, 18, 286–294. [Google Scholar] [CrossRef]
- Zhou, Z.; Barrett, P.M.; Hilton, J. An exceptionally preserved lower cretaceous ecosystem. Nature 2003, 421, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Wegweiser, M.; Hartman, S.; Lovelace, D. Duckbill dinosaur chin skin scales: Ups, downs and arounds of surficial morphology of Upper Cretaceous Lance Formation dinosaur skin. New Mex. Mus. Nat. Hist. Sci. Bull. 2006, 35, 119–126. [Google Scholar]
- Young, G.A.; Rudkin, D.M.; Dobrzanski, E.P.; Robson, S.P.; Nowlan, G.S. Exceptionally preserved late Ordovician biotas from Manitoba, Canada. Geology 2007, 35, 883–886. [Google Scholar] [CrossRef]
- Manning, P.L.; Morris, P.M.; McMahon, A.; Jones, E.; Gize, A.; Macquaker, J.H.S.; Wolff, G.; Thompson, A.; Marshall, J.; Taylor, K.G.; et al. Mineralized soft-tissue structure and chemistry in a mummified hadrosaur from the Hell Creek Formation, North Dakota (USA). Proc. R. Soc. B Biol. Sci. 2009, 276, 3429–3437. [Google Scholar] [CrossRef] [PubMed]
- Davis, M. Census of dinosaur skin reveals lithology may not be the most important factor in increased preservation of hadrosaurid skin. Acta Palaeontol. Pol. 2014, 59, 601–605. [Google Scholar] [CrossRef] [Green Version]
- Schroeter, E.R.; Egerton, V.M.; Ibiricu, L.M.; Lacovara, K.J. Lamniform shark teeth from the late cretaceous of southernmost South America (Santa Cruz Province, Argentina). PLoS ONE 2014, 9, e104800. [Google Scholar] [CrossRef] [PubMed]
- Sickmann, Z.T.; Schwartz, T.M.; Graham, S.A. Refining stratigraphy and tectonic history using detrital zircon maximum depositional age: An example from the Cerro Fortaleza Formation, Austral Basin, southern Patagonia. Basin Res. 2018, 30, 708–729. [Google Scholar] [CrossRef]
- Ullmann, P.V.; Lacovara, K.J. Appendicular osteology of Dreadnoughtus schrani, a giant titanosaurian (Sauropoda, Titanosauria) from the Upper Cretaceous of Patagonia, Argentina. J. Vertebr. Paleontol. 2016, 36, e1225303. [Google Scholar] [CrossRef]
- Macellari, C.E.; Barrio, C.A.; Manassero, M.J. Upper Cretaceous to Paleocene depositional sequences and sandstone petrography of southwestern Patagonia (Argentina and Chile). J. S. Am. Earth Sci. 1989, 2, 223–239. [Google Scholar] [CrossRef]
- Marenssi, S.A.; Casadio, S.; Santillana, S.N. Estratigrafía y sedimentología de las unidades del Cretácico superior-Paleógeno aflorantes en la margen sureste del lago Viedma, provincia de Santa Cruz, Argentina. Rev. Asoc. Geol. Argent. 2003, 58, 403–416. [Google Scholar]
- Egerton, V.M.; Williams, C.J.; Lacovara, K.J. A new Late Cretaceous (late Campanian to early Maastrichtian) wood flora from southern Patagonia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 441, 305–316. [Google Scholar] [CrossRef]
- Rombola, C.F.; Greppi, C.D.; Pujana, R.R.; García Massini, J.L.; Bellosi, E.S.; Marenssi, S.A. Brachyoxylon fossil woods with traumatic resin canals from the Upper Cretaceous Cerro Fortaleza Formation, southern Patagonia (Santa Cruz Province, Argentina). Cretac. Res. 2022, 130, 105065. [Google Scholar] [CrossRef]
- Voegele, K.K.; Ullmann, P.V.; Lamanna, M.C.; Lacovara, K.J. Appendicular myological reconstruction of the forelimb of the giant titanosaurian sauropod dinosaur Dreadnoughtus schrani. J. Anat. 2020, 237, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Voegele, K.K.; Ullmann, P.V.; Lamanna, M.C.; Lacovara, K.J. Myological reconstruction of the pelvic girdle and hind limb of the giant titanosaurian sauropod dinosaur Dreadnoughtus schrani. J. Anat. 2021, 238, 576–597. [Google Scholar] [CrossRef] [PubMed]
- Gates, T.A. The Late Jurassic Cleveland-Lloyd dinosaur quarry as a drought-induced assemblage. Palaios 2005, 20, 363–375. [Google Scholar] [CrossRef]
- Green, J.L.; Schweitzer, M.H.; Lamm, E.T. Limb bone histology and growth in Placerias hesternus (Therapsida: Anomodontia) from the Upper Triassic of North America. Palaeontology 2010, 53, 347–364. [Google Scholar] [CrossRef]
- Boyd, C.A.; Cleland, T.P.; Novas, F. Osteogenesis, homology, and function of the intercostal plates in ornithischian dinosaurs (Tetrapoda, Sauropsida). Zoomorphology 2011, 130, 305–313. [Google Scholar] [CrossRef]
- Cleland, T.P.; Stoskopf, M.K.; Schweitzer, M.H. Histological, chemical, and morphological reexamination of the “heart” of a small Late Cretaceous Thescelosaurus. Naturwissenschaften 2011, 98, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Ullmann, P.V.; Grandstaff, D.E.; Ash, R.D.; Lacovara, K.J. Geochemical taphonomy of the Standing Rock Hadrosaur Site: Exploring links between rare earth elements and cellular and soft tissue preservation. Geochim. Cosmochim. Acta 2020, 269, 223–237. [Google Scholar] [CrossRef]
- Gromet, L.P.; Haskin, L.A.; Korotev, R.L.; Dymek, R.F. The “North American shale composite”: Its compilation, major and trace element characteristics. Geochim. Cosmochim. Acta 1984, 48, 2469–2482. [Google Scholar] [CrossRef]
- Haskin, L.A.; Haskin, M.A.; Frey, F.A.; Wildeman, T.R. Relative and absolute terrestrial abundances of the rare earths. In Origin and Distribution of the Elements; Ahrens, L.H., Ed.; Pergamon: Oxford, UK, 1968; pp. 889–912. [Google Scholar]
- Herwartz, D.; Tütken, T.; Jochum, K.P.; Sander, P.M. Rare earth element systematics of fossil bone revealed by LA-ICPMS analysis. Geochim. Cosmochim. Acta 2013, 103, 161–183. [Google Scholar] [CrossRef]
- Jans, M.M.E.; Nielsen-Marsh, C.M.; Smith, C.I.; Collins, M.J.; Kars, H. Characterisation of microbial attack on archaeological bone. J. Archaeol. Sci. 2004, 31, 87–95. [Google Scholar] [CrossRef]
- Hedges, R.E.M.; Millard, A.R.; Pike, A.W.G. Measurements and Relationships of Diagenetic Alteration of Bone from Three Archaeological Sites. J. Archaeol. Sci. 1995, 22, 201–209. [Google Scholar] [CrossRef]
- Nielsen-marsh, C.; Gernaey, A.; Turner-Walker, G.; Hedges, R.; Pike, A.; Collins, M. The Chemical Degradation of Bone. In Human Osteology: In Archaeology and Forensic Science; Cambridge University Press: Cambridge, UK, 2000; pp. 439–454. [Google Scholar]
- Kohn, M.J. Models of diffusion-limited uptake of trace elements in fossils and rates of fossilization. Geochim. Cosmochim. Acta 2008, 72, 3758–3770. [Google Scholar] [CrossRef]
- Grandstaff, D.E.; Terry, D.O. Rare earth element composition of Paleogene vertebrate fossils from Toadstool Geologic Park, Nebraska, USA. Appl. Geochem. 2009, 24, 733–745. [Google Scholar] [CrossRef]
- Bau, M.; Dulski, P. Distribution of yttrium and rare-earth elements in the Penge and Kuruman iron-formations, Transvaal Supergroup, South Africa. Precambrian Res. 1996, 79, 37–55. [Google Scholar] [CrossRef]
- Pack, A.; Russell, S.S.; Shelley, J.M.G.; van Zuilen, M. Geo- and cosmochemistry of the twin elements yttrium and holmium. Geochim. Cosmochim. Acta 2007, 71, 4592–4608. [Google Scholar] [CrossRef]
- Schweitzer, M.H.; Wittmeyer, J.L.; Horner, J.R. Soft tissue and cellular preservation in vertebrate skeletal elements from the Cretaceous to the present. Proc. R. Soc. Lond. B 2007, 274, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Schweitzer, M.H.; Wittmeyer, J.L.; Horner, J.R.; Toporski, J.K. Soft-tissue vessels and cellular preservation in Tyrannosaurus rex. Science 2005, 307, 1952–1955. [Google Scholar] [CrossRef]
- Bartnicki-Garcia, S.; Bracker, C.E.; Gierz, G.; López-Franco, R.; Haisheng, L. Mapping the growth of fungal hyphae: Orthogonal cell wall expansion during tip growth and the role of turgor. Biophys. J. 2000, 79, 2382–2390. [Google Scholar] [CrossRef] [Green Version]
- Harris, S.D. Branching of fungal hyphae: Regulation, mechanisms and comparison with other branching systems. Mycologia 2008, 100, 823–832. [Google Scholar] [CrossRef] [Green Version]
- Nesse, W. Optical mineralogy. In Introduction to Mineralogy; Oxford University Press: New York, NY, USA, 2000; pp. 114–159. [Google Scholar]
- Perkins, D.; Henke, K.R. Minerals in Thin Section; Prentice Hall: Upper Saddle River, NJ, USA, 2003. [Google Scholar]
- Cadena, E.A.; Schweitzer, M.H. Variation in osteocytes morphology vs. bone type in turtle shell and their exceptional preservation from the Jurassic to the present. Bone 2012, 51, 614–620. [Google Scholar] [CrossRef]
- Schweitzer, M.H.; Zheng, W.; Cleland, T.P.; Bern, M. Molecular analyses of dinosaur osteocytes support the presence of endogenous molecules. Bone 2013, 52, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Armitage, M.H.; Anderson, K.L. Soft sheets of fibrillar bone from a fossil of the supraorbital horn of the dinosaur Triceratops horridus. Acta Histochem. 2013, 115, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Lowenstein, J.M.; Scheuenstuhl, G. Immunological methods in molecular palaeontology. Philos. Trans.-R. Soc. Lond. B 1991, 333, 375–380. [Google Scholar] [CrossRef]
- Frank, S.A. Classification by antigenicity and phylogeny. In Immunology and Evolution of Infectious Disease; Princeton University Press: Princeton, NJ, USA, 2002; pp. 175–187. [Google Scholar]
- Hedges, S.B.; Dudley, J.; Kumar, S. TimeTree: A public knowledge-base of divergence times among organisms. Bioinformatics 2006, 22, 2971–2972. [Google Scholar] [CrossRef]
- Brochu, C.A. Progress and future directions in archosaur phylogenetics. J. Paleontol. 2001, 75, 1185–1201. [Google Scholar] [CrossRef]
- Ramos-Vara, J.A.; Beissenherz, M.E. Optimization of immunohistochemical methods using two different antigen retrieval methods on formalin-fixed, paraffin-embedded tissues: Experience with 63 markers. J. Vet. Diagn. Investig. 2000, 12, 307–311. [Google Scholar] [CrossRef] [Green Version]
- Harper, E. Collagenases. Annu. Rev. Biochem. 1980, 49, 1063–1078. [Google Scholar] [CrossRef]
- Harper, E.; Kang, A.H. Studies on the specificity of bacterial collagenases. Biochem. Biophys. Res. Commun. 1970, 41, 482–487. [Google Scholar] [CrossRef]
- Avci, R.; Schweitzer, M.H.; Boyd, R.D.; Wittmeyer, J.L.; Arce, F.T.; Calvo, J.O. Preservation of bone collagen from the Late Cretaceous period studied by immunological techniques and atomic force microscopy. Am. Chem. Soc. 2005, 21, 3584–3591. [Google Scholar] [CrossRef]
- Werner, M.; Chott, A.; Fabiano, A.; Battifora, H. Effect of formalin tissue fixation and processing on immunohistochemistry. Am. J. Surg. Pathol. 2000, 24, 1016–1019. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, M.H.; Avci, R.; Collier, T.C.; Goodwin, M.B. Microscopic, chemical and molecular methods for examining fossil preservation. Comptes Rendus Palevol 2008, 7, 159–184. [Google Scholar] [CrossRef]
- Saper, C.B. A guide to the perplexed on the specificity of antibodies. J. Histochem. Cytochem. 2009, 57, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Paulie, S.; Perlmann, H.; Perlmann, P. Enzyme-linked Immunosorbent Assay. Encycl. Life Sci. 2006, 21, 165–209. [Google Scholar] [CrossRef]
- Avci, R.; Schweitzer, M.; Boyd, R.D.; Wittmeyer, J.; Steeled, A.; Toporski, J.; Beech, I.; Arce, F.T.; Spangler, B.; Cole, K.M.; et al. Comparison of antibody-antigen interactions on collagen measured by conventional immunological techniques and atomic force microscopy. Langmuir 2004, 20, 11053–11063. [Google Scholar] [CrossRef] [PubMed]
- Lechtzier, V.; Hutoran, M.; Levy, T.; Kotler, M.; Brenner, T.; Steinitz, M. Sodium dodecyl sulphate-treated proteins as ligands in ELISA. J. Immunol. Methods 2002, 270, 19–26. [Google Scholar] [CrossRef]
- Appiah, A.S.; Amoatey, H.M.; Klu, G.Y.; Afful, N.T.; Azu, E.; Owusu, G.K. Spread of African cassava mosaic virus from cassava (Manihot esculenta Crantz) to physic nut (Jatropha curcas L.) in Ghana. J. Phytol. 2012, 4, 31–37. [Google Scholar]
- Ostlund, E.N.; Crom, R.L.; Pedersen, D.D.; Johnson, D.J.; Williams, W.O.; Schmitt, B.J. Equine West Nile encephalitis, United States. Emerg. Infect. Dis. 2001, 7, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Tabatabai, L.B.; Deyoe, B.L. Specific enzyme-linked immunosorbent assay for detection of bovine antibody to Brucella abortus. J. Clin. Microbiol. 1984, 20, 209–213. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.I.; Craig, O.E.; Prigodich, R.V.; Nielsen-Marsh, C.M.; Jans, M.M.E.; Vermeer, C.; Collins, M.J. Diagenesis and survival of osteocalcin in archaeological bone. J. Archaeol. Sci. 2005, 32, 105–113. [Google Scholar] [CrossRef]
- Nielsen-Marsh, C.M.; Hedges, R.E.M. Patterns of diagenesis in bone I: The effects of site environments. J. Archaeol. Sci. 2000, 27, 1139–1150. [Google Scholar] [CrossRef]
- Nielsen-Marsh, C.M.; Hedges, R.E.M. Patterns of diagenesis in bone II: Effects of acetic acid treatment and the removal of diagenetic CO32−. J. Archaeol. Sci. 2000, 27, 1151–1159. [Google Scholar] [CrossRef]
- Martin, J.E.; Patrick, D.; Kihm, A.J.; Foit, F.F.; Grandstaff, D.E. Lithostratigraphy, tephrochronology, and rare earth element geochemistry of fossils at the classical pleistocene Fossil Lake area, south central Oregon. J. Geol. 2005, 113, 139–155. [Google Scholar] [CrossRef] [Green Version]
- Trueman, C.N. Rare earth element geochemistry and taphonomy of terrestrial vertebrate assemblages. Palaios 1999, 14, 555–568. [Google Scholar] [CrossRef]
- Suarez, C.A.; Suarez, M.B.; Terry, D.O.; Grandstaff, D.E. Rare earth element geochemistry and taphonomy of the early Cretaceous Crystal Geyser Dinosaur Quarry, east-central Utah. Palaios 2007, 22, 500–512. [Google Scholar] [CrossRef]
- Titus, A.L.; Knoll, K.; Sertich, J.J.W.; Yamamura, D.; Suarez, C.A.; Glasspool, I.J.; Ginouves, J.E.; Lukacic, A.K.; Roberts, E.M. Geology and taphonomy of a unique tyrannosaurid bonebed from the upper Campanian Kaiparowits Formation of southern Utah: Implications for tyrannosaurid gregariousness. PeerJ 2021, 9, e11013. [Google Scholar] [CrossRef]
- Rogers, R.R.; Fricke, H.C.; Addona, V.; Canavan, R.R.; Dwyer, C.N.; Harwood, C.L.; Koenig, A.E.; Murray, R.; Thole, J.T.; Williams, J. Using laser ablation-inductively coupled plasma-mass spectrometry (LA-ICP-MS) to explore geochemical taphonomy of vertebrate fossils in the upper cretaceous two medicine and Judith River formations of Montana. Palaios 2010, 25, 183–195. [Google Scholar] [CrossRef]
- Kowal-Linka, M.; Jochum, K.P.; Surmik, D. LA-ICP-MS analysis of rare earth elements in marine reptile bones from the Middle Triassic bonebed (Upper Silesia, S Poland): Impact of long-lasting diagenesis, and factors controlling the uptake. Chem. Geol. 2014, 363, 213–228. [Google Scholar] [CrossRef]
- McCormack, J.M.; Bahr, A.; Gerdes, A.; Tütken, T.; Prinz-Grimm, P. Preservation of successive diagenetic stages in Middle Triassic bonebeds: Evidence from in situ trace element and strontium isotope analysis of vertebrate fossils. Chem. Geol. 2015, 410, 108–123. [Google Scholar] [CrossRef]
- Suarez, C.A.; Morschhauser, E.M.; Suarez, M.B.; You, H.; Li, D.; Dodson, P. Rare earth element geochemistry of bone beds from the Lower Cretaceous Zhonggou Formation of Gansu Province, China. J. Vertebr. Paleontol. 2018, 38, 22–35. [Google Scholar] [CrossRef]
- Suarez, C.A.; Kohn, M.J. Caught in the act: A case study on microscopic scale physicochemical effects of fossilization on stable isotopic composition of bone. Geochim. Cosmochim. Acta 2020, 268, 277–295. [Google Scholar] [CrossRef]
- Botfalvai, G.; Csiki-Sava, Z.; Kocsis, L.; Albert, G.; Magyar, J.; Bodor, E.R.; Tabara, D.; Ulyanov, A.; Makádi, L. ‘X’ marks the spot! Sedimentological, geochemical and palaeontological investigations of Upper Cretaceous (Maastrichtian) vertebrate fossil localities from the Valioara valley (Densus-Ciula Formation, Hațeg Basin, Romania). Cretac. Res. 2021, 123, 104781. [Google Scholar] [CrossRef]
- Ferrante, C.; Cavin, L.; Vennemann, T.; Martini, R. Histology and Geochemistry of Allosaurus (Dinosauria: Theropoda) from the Cleveland-Lloyd Dinosaur Quarry (Late Jurassic, Utah): Paleobiological Implications. Front. Earth Sci. 2021, 9, 225. [Google Scholar] [CrossRef]
- Ullmann, P.V.; Macauley, K.; Ash, R.D.; Shoup, B.; Scannella, J.B. Taphonomic and diagenetic pathways to protein preservation, part i: The case of tyrannosaurus rex specimen mor 1125. Biology 2021, 10, 1193. [Google Scholar] [CrossRef]
- Trueman, C.N.; Palmer, M.R.; Field, J.; Privat, K.; Ludgate, N.; Chavagnac, V.; Eberth, D.A.; Cifelli, R.; Rogers, R.R. Comparing rates of recrystallisation and the potential for preservation of biomolecules from the distribution of trace elements in fossil bones. Comptes Rendus-Palevol 2008, 7, 145–158. [Google Scholar] [CrossRef]
- Herwartz, D.; Tütken, T.; Münker, C.; Jochum, K.P.; Stoll, B.; Sander, P.M. Timescales and mechanisms of REE and Hf uptake in fossil bones. Geochim. Cosmochim. Acta 2011, 75, 82–105. [Google Scholar] [CrossRef]
- Trueman, C.N.; Kocsis, L.; Palmer, M.R.; Dewdney, C. Fractionation of rare earth elements within bone mineral: A natural cation exchange system. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 310, 124–132. [Google Scholar] [CrossRef]
- Wings, O. Authigenic minerals in fossil bones from the Mesozoic of England: Poor correlation with depositional environments. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2004, 204, 15–32. [Google Scholar] [CrossRef]
- Janssens, K.; Vincze, L.; Vekemans, B.; Williams, C.T.; Radtke, M.; Haller, M.; Knöchel, A. The non-destructive determination of REE in fossilized bone using synchrotron radiation induced K-line X-ray microfluorescence analysis. Fresenius. J. Anal. Chem. 1999, 363, 413–420. [Google Scholar] [CrossRef]
- Hinz, E.A.; Kohn, M.J. The effect of tissue structure and soil chemistry on trace element uptake in fossils. Geochim. Cosmochim. Acta 2010, 74, 3213–3231. [Google Scholar] [CrossRef]
- Suarez, C.A.; Macpherson, G.L.; González, L.A.; Grandstaff, D.E. Heterogeneous rare earth element (REE) patterns and concentrations in a fossil bone: Implications for the use of REE in vertebrate taphonomy and fossilization history. Geochim. Cosmochim. Acta 2010, 74, 2970–2988. [Google Scholar] [CrossRef]
- Kohn, M.J.; Moses, R.J. Trace element diffusivities in bone rule out simple diffusive uptake during fossilization but explain in vivo uptake and release. Proc. Natl. Acad. Sci. USA 2013, 110, 419–424. [Google Scholar] [CrossRef] [Green Version]
- Elderfield, H.; Upstill-Goddard, R.; Sholkovitz, E.R. The rare earth elements in rivers, estuaries, and coastal seas and their significance to the composition of ocean waters. Geochim. Cosmochim. Acta 1990, 54, 971–991. [Google Scholar] [CrossRef]
- Johannesson, K.H.; Tang, J.; Daniels, J.M.; Bounds, W.J.; Burdige, D.J. Rare earth element concentrations and speciation in organic-rich blackwaters of the Great Dismal Swamp, Virginia, USA. Chem. Geol. 2004, 209, 271–294. [Google Scholar] [CrossRef]
- Smith, C.; Liu, X.M. Spatial and temporal distribution of rare earth elements in the Neuse River, North Carolina. Chem. Geol. 2018, 488, 34–43. [Google Scholar] [CrossRef]
- Luo, Y.R.; Byrne, R.H. Carbonate complexation of yttrium and the rare earth elements in natural waters. Geochim. Cosmochim. Acta 2004, 68, 691–699. [Google Scholar] [CrossRef]
- Pourret, O.; Davranche, M.; Gruau, G.; Dia, A. Rare earth elements complexation with humic acid. Chem. Geol. 2007, 243, 128–141. [Google Scholar] [CrossRef] [Green Version]
- Byrne, R.H.; Liu, X.; Schijf, J. The influence of phosphate coprecipitation on rare earth distributions in natural waters. Geochim. Cosmochim. Acta 1996, 60, 3341–3346. [Google Scholar] [CrossRef]
- Metzger, C.A.; Terry, D.O.; Grandstaff, D.E. Effect of paleosol formation on rare earth element signatures in fossil bone. Geology 2004, 32, 497–500. [Google Scholar] [CrossRef]
- Company, J. Bone histology of the titanosaur Lirainosaurus astibiae (Dinosauria: Sauropoda) from the Latest Cretaceous of Spain. Naturwissenschaften 2011, 98, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Klein, N.; Christian, A.; Sander, P.M. Histology shows that elongated neck ribs in sauropod dinosaurs are ossified tendons. Biol. Lett. 2012, 8, 1032–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, K.; Csiki, Z.; Curry Rogers, K.; Weishampel, D.B.; Redelstorff, R.; Carballido, J.L.; Sander, P.M. Small body size and extreme cortical bone remodeling indicate phyletic dwarfism in Magyarosaurus dacus (Sauropoda: Titanosauria). Proc. Natl. Acad. Sci. USA 2010, 107, 9258–9263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, D.H. Introduction to the Fungi. In Fungal Physiology; Griffin, D.H., Ed.; Wiley-Liss, Inc.: New York, NY, USA, 1994; pp. 1–23. [Google Scholar]
- Kaye, T.G.; Gaugler, G.; Sawlowicz, Z. Dinosaurian soft tissues interpreted as bacterial biofilms. PLoS ONE 2008, 3, e2808. [Google Scholar] [CrossRef] [Green Version]
- Buckley, M.; Wadsworth, C. Proteome degradation in ancient bone: Diagenesis and phylogenetic potential. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 416, 69–79. [Google Scholar] [CrossRef]
- Brandt, E.; Weichmann, I.; Grupe, G. Howe reliable are immunological tools for the detection of ancient proteins in fossil bones? Int. J. Osteoarchaeol. 2002, 12, 307–316. [Google Scholar] [CrossRef]
- Ullmann, P.V.; Voegele, K.K.; Grandstaff, D.E.; Ash, R.D.; Zheng, W.; Schroeter, E.R.; Schweitzer, M.H.; Lacovara, K.J. Molecular tests support the viability of rare earth elements as proxies for fossil biomolecule preservation. Sci. Rep. 2020, 10, 15566. [Google Scholar] [CrossRef] [PubMed]
- Hubert, J.F.; Panish, P.T.; Chure, D.J.; Prostak, K.S. Chemistry, microstructure, petrology, and diagenetic model of jurassic dinosaur bones, dinosaur national monument, Utah. J. Sediment. Res. 1996, 66, 531–547. [Google Scholar]
- Tuross, N. Albumin preservation in the Taima-taima mastodon skeleton. Appl. Geochem. 1989, 4, 255–259. [Google Scholar] [CrossRef]
- Grupe, G. Preservation of collagen in bone from dry, sandy soil. J. Archaeol. Sci. 1995, 22, 193–199. [Google Scholar] [CrossRef]
- Koenig, A.E.; Rogers, R.R.; Trueman, C.N. Visualizing fossilization using laser ablation-inductively coupled plasma-mass spectrometry maps of trace elements in Late Cretaceous bones. Geology 2009, 37, 511–514. [Google Scholar] [CrossRef]
- Schweitzer, M.H. Molecular paleontology: Some current advances and problems. Ann. Paleontol. 2004, 90, 81–102. [Google Scholar] [CrossRef]
- Gatti, L.; Lugli, F.; Sciutto, G.; Zangheri, M.; Prati, S.; Mirasoli, M. Combining elemental and immunochemical analyses to characterize diagenetic alteration patterns in ancient skeletal remains. Sci. Rep. 2022, 12, 5112. [Google Scholar] [CrossRef] [PubMed]
- Abbott, A.N.; Haley, B.A.; McManus, J.; Reimers, C.E. The sedimentary flux of dissolved rare earth elements to the ocean. Geochim. Cosmochim. Acta 2015, 154, 186–200. [Google Scholar] [CrossRef]
- Åström, M.; Corin, N. Distribution of rare earth elements in anionic, cationic and particulate fractions in boreal humus-rich streams affected by acid sulphate soils. Water Res. 2003, 37, 273–280. [Google Scholar] [CrossRef]
- Barroux, G.; Sonke, J.E.; Boaventura, G.; Viers, J.; Godderis, Y.; Bonnet, M.-P.; Sondag, F.; Gardoll, S.; Lagane, C.; Seyler, P. Seasonal dissolved rare earth element dynamics of the Amazon River main stem, its tributaries, and the Curuaí floodplain. Geochem. Geophys. Geosyst. 2006, 7, Q12005. [Google Scholar] [CrossRef]
- Bau, M.; Dulski, P. Anthropogenic origin of positive gadolinium anomalies in river waters. Earth Planet. Sci. Lett. 1996, 143, 245–255. [Google Scholar] [CrossRef]
- Bau, M.; Knappe, A.; Dulski, P. Anthropogenic gadolinium as a micropollutant in river waters in Pennsylvania and in Lake Erie, northeastern United States. Geochemistry 2006, 66, 143–152. [Google Scholar] [CrossRef]
- Bayon, G.; Birot, D.; Ruffine, L.; Caprais, J.-C.; Ponzevera, E.; Bollinger, C.; Donval, J.-P.; Charlou, J.-L.; Voisset, M.; Grimaud, S. Evidence for intense REE scavenging at cold seeps from the Niger Delta margin. Earth Planet. Sci. Lett. 2011, 312, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Biddau, R.; Cidu, R.; Frau, F. Rare earth elements in waters from the albitite-bearing granodiorites of Central Sardinia, Italy. Chem. Geol. 2002, 182, 1–14. [Google Scholar] [CrossRef]
- Bwire Ojiambo, S.; Lyons, W.B.; Welch, K.A.; Poreda, R.J.; Johannesson, K.H. Strontium isotopes and rare earth elements as tracers of groundwater-lake water interactions, Lake Naivasha, Kenya. Appl. Geochem. 2003, 18, 1789–1805. [Google Scholar] [CrossRef]
- Censi, P.; Sposito, F.; Inguaggiato, C.; Zuddas, P.; Inguaggiato, S.; Venturi, M. Zr, Hf and REE distribution in river water under different ionic strength conditions. Sci. Total Environ. 2018, 645, 837–853. [Google Scholar] [CrossRef] [PubMed]
- Centeno, L.M.; Faure, G.; Lee, G.; Talnagi, J. Fractionation of chemical elements including the REEs and 226Ra in stream contaminated with coal-mine effluent. Appl. Geochem. 2004, 19, 1085–1095. [Google Scholar] [CrossRef]
- Chevis, D.A.; Johannesson, K.H.; Burdige, D.J.; Tang, J.; Moran, S.B.; Kelly, R.P. Submarine groundwater discharge of rare earth elements to a tidally-mixed estuary in Southern Rhode Island. Chem. Geol. 2015, 397, 128–142. [Google Scholar] [CrossRef] [Green Version]
- de Baar, H.J.W.; Bruland, K.W.; Schijf, J.; van Heuven, S.M.A.C.; Behrens, M.K. Low cerium among the dissolved rare earth elements in the central North Pacific Ocean. Geochim. Cosmochim. Acta 2018, 236, 5–40. [Google Scholar] [CrossRef]
- de Baar, H.J.W.; Bacon, M.P.; Brewer, P.G. Rare-earth distributions with a positive Ce anomaly in the Western North Atlantic Ocean. Nature 1983, 301, 324–327. [Google Scholar] [CrossRef]
- De Carlo, E.H.; Green, W.J. Rare earth elements in the water column of Lake Vanda, McMurdo Dry Valleys, Antarctica. Geochim. Cosmochim. Acta 2002, 66, 1323–1333. [Google Scholar] [CrossRef]
- Elderfield, H.; Greaves, M.J. The rare earth elements in seawater. Nature 1982, 296, 214–219. [Google Scholar] [CrossRef]
- Elderfield, H.; Sholkovitz, E.R. Rare earth elements in the pore waters of reducing nearshore sediments. Earth Planet. Sci. Lett. 1987, 82, 280–288. [Google Scholar] [CrossRef]
- Esmaeili-Vardanjani, M.; Shamsipour-Dehkordi, R.; Eslami, A.; Moosaei, F.; Pazand, K. A study of differentiation pattern and rare earth elements migration in geochemical and hydrogeochemical environments of Airekan and Cheshmeh Shotori areas (Central Iran). Environ. Earth Sci. 2013, 68, 719–732. [Google Scholar] [CrossRef]
- Gammons, C.H.; Wood, S.A.; Pedrozo, F.; Varekamp, J.C.; Nelson, B.J.; Shope, C.L.; Baffico, G. Hydrogeochemistry and rare earth element behavior in a volcanically acidified watershed in Patagonia, Argentina. Chem. Geol. 2005, 222, 249–267. [Google Scholar] [CrossRef]
- Gammons, C.H.; Wood, S.A.; Nimick, D.A. Diel behavior of rare earth elements in a mountain stream with acidic to neutral pH. Geochim. Cosmochim. Acta 2005, 69, 3747–3758. [Google Scholar] [CrossRef]
- Garcia-Solsona, E.; Jeandel, C.; Labatut, M.; Lacan, F.; Vance, D.; Chavagnac, V.; Pradoux, C. Rare earth elements and Nd isotopes tracing water mass mixing and particle-seawater interactions in the SE Atlantic. Geochim. Cosmochim. Acta 2014, 125, 351–372. [Google Scholar] [CrossRef]
- German, C.R.; Masuzawa, T.; Greaves, M.J.; Elderfield, H.; Edmond, J.M. Dissolved rare earth elements in the Southern Ocean: Cerium oxidation and the influence of hydrography. Geochim. Cosmochim. Acta 1995, 59, 1551–1558. [Google Scholar] [CrossRef]
- German, C.R.; Holliday, B.P.; Elderfield, H. Redox cycling of rare earth elements in the suboxic zone of the Black Sea. Geochim. Cosmochim. Acta 1991, 55, 3553–3558. [Google Scholar] [CrossRef]
- Goldstein, S.J.; Jacobsen, S.B. Rare earth elements in river waters. Earth Planet. Sci. Lett. 1988, 89, 35–47. [Google Scholar] [CrossRef]
- Grenier, M.; Jeandel, C.; Lacan, F.; Vance, D.; Venchiarutti, C.; Cros, A.; Cravatte, S. From the subtropics to the central equatorial Pacific Ocean: Neodymium isotopic composition and rare earth element concentration variations. J. Geophys. Res.-Oceans 2013, 118, 592–618. [Google Scholar] [CrossRef] [Green Version]
- Haley, B.A.; Klinkhammer, G.P.; McManus, J. Rare earth elements in pore waters of marine sediments. Geochim. Cosmochim. Acta 2004, 68, 1265–1279. [Google Scholar] [CrossRef]
- Hathorne, E.C.; Stichel, T.; Brück, B.; Frank, M. Rare earth element distribution in the Atlantic sector of the Southern Ocean: The balance between particle scavenging and vertical supply. Mar. Chem. 2015, 177, 157–171. [Google Scholar] [CrossRef] [Green Version]
- Hongo, Y.; Obata, H.; Alibo, D.S.; Nozaki, Y. Spatial variations of rare earth elements in North Pacific surface water. J. Oceanogr. 2006, 62, 441–455. [Google Scholar] [CrossRef]
- Hoyle, J.; Elderfield, H.; Gledhill, A.; Greaves, M. The behaviour of the rare earth elements during mixing of river and sea waters. Geochim. Cosmochim. Acta 1984, 48, 143–149. [Google Scholar] [CrossRef]
- Jeandel, C.; Delattre, H.; Grenier, M.; Pradoux, C.; Lacan, F. Rare earth element concentrations and Nd isotopes in the Southeast Pacific Ocean. Geochem. Geophys. Geosyst. 2013, 14, 328–341. [Google Scholar] [CrossRef] [Green Version]
- Johannesson, K.H.; Palmore, C.D.; Fackrell, J.; Prouty, N.G.; Swarzenski, P.W.; Chevis, D.A.; Telfeyan, K.; White, C.D.; Burdige, D.J. Rare earth element behavior during groundwater-seawater mixing along the Kona Coast of Hawaii. Geochim. Cosmochim. Acta 2017, 198, 229–258. [Google Scholar] [CrossRef] [Green Version]
- Johannesson, K.H.; Farnham, I.M.; Guo, C.; Stetzenbach, K.J. Rare earth element fractionation and concentration variations along a groundwater flow path within a shallow, basin-fill aquifer, southern Nevada, USA. Geochim. Cosmochim. Acta 1999, 63, 2697–2708. [Google Scholar] [CrossRef]
- Johannesson, K.H.; Stetzenbach, K.J.; Hodge, V.F. Rare earth elements as geochemical tracers of regional groundwater mixing. Geochim. Cosmochim. Acta 1997, 61, 3605–3618. [Google Scholar] [CrossRef]
- Johannesson, K.H.; Lyons, W.B.; Stetzenbach, K.J.; Byrne, R.H. The solubility control of rare earth elements in natural terrestrial waters and the significance of PO43- and CO32- in limiting dissolved rare earth concentrations: A review of recent information. Aquat. Geochem. 1995, 1, 157–173. [Google Scholar] [CrossRef]
- Johannesson, K.H.; Lyons, W.B. Rare-earth element geochemistry of Colour Lake, an acidic freshwater lake on Axel Heiberg Island, Northwest Territories, Canada. Chem. Geol. 1995, 119, 209–223. [Google Scholar] [CrossRef]
- Kalender, L.; Aytimur, G. REE geochemistry of Euphrates River, Turkey. J. Chem. 2016, 2016, 1012021. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Torres, M.E.; Haley, B.A.; Kastner, M.; Pohlman, J.W.; Riedel, M.; Lee, Y.J. The effect of diagenesis and fluid migration on rare earth element distribution in fluids of the northern Cascadia accretionary margin. Chem. Geol. 2011, 291, 152–165. [Google Scholar] [CrossRef]
- Kulaksiz, S.; Bau, M. Anthropogenic gadolinium as a microcontaminant in tap water used as drinking water in urban areas and megacities. Appl. Geochem. 2011, 26, 1877–1885. [Google Scholar] [CrossRef]
- Kulaksiz, S.; Bau, M. Contrasting behaviour of anthropogenic gadolinium and natural rare earth elements in estuaries and the gadolinium input into the North Sea. Earth Planet. Sci. Lett. 2007, 260, 361–371. [Google Scholar] [CrossRef]
- Leybourne, M.I.; Goodfellow, W.D.; Boyle, D.R.; Hall, G.M. Rapid development of negative Ce anomalies in surface waters and contrasting REE patterns in groundwaters associated with Zn-Pb massive sulphide deposits. Appl. Geochem. 2000, 15, 695–723. [Google Scholar] [CrossRef]
- Liu, H.; Guo, H.; Xing, L.; Zhan, Y.; Li, F.; Shao, J.; Niu, H.; Liang, X.; Li, C. Geochemical behaviors of rare earth elements in groundwater along a flow path in the North China Plain. J. Asian Earth Sci. 2016, 117, 33–51. [Google Scholar] [CrossRef]
- Merschel, G.; Bau, M.; Schmidt, K.; Münker, C.; Dantas, E.L. Hafnium and neodymium isotopes and REY distribution in the truly dissolved, nanoparticulate/colloidal and suspended loads of rivers in the Amazon Basin, Brazil. Geochim. Cosmochim. Acta 2017, 213, 383–399. [Google Scholar] [CrossRef]
- Nozaki, Y.; Lerche, D.; Alibo, D.S.; Tsutsumi, M. Dissolved indium and rare earth elements in three Japanese rivers and Tokyo Bay: Evidence for anthropogenic Gd and In. Geochim. Cosmochim. Acta 2000, 64, 3975–3982. [Google Scholar] [CrossRef]
- Osborne, A.H.; Hathorne, E.C.; Schijf, J.; Plancherel, Y.; Böning, P.; Frank, M. The potential of sedimentary foraminiferal rare earth element patterns to trace water masses in the past. Geochem. Geophys. Geosyst. 2017, 18, 1550–1568. [Google Scholar] [CrossRef]
- Piepgras, D.J.; Jacobsen, S.B. The behavior of rare earth elements in seawater: Precise determination of variations in the North Pacific water column. Geochim. Cosmochim. Acta 1992, 56, 1851–1862. [Google Scholar] [CrossRef]
- Pokrovsky, O.S.; Viers, J.; Shirokova, L.S.; Shevchenko, V.P.; Filipov, A.S.; Dupré, B. Dissolved, suspended, and colloidal fluxes of organic carbon, major and trace elements in the Severnaya Dvina River and its tributary. Chem. Geol. 2010, 273, 136–149. [Google Scholar] [CrossRef]
- Reynard, B.; Lecuyer, C.; Grandjean, P. Crystal-chemical controls on rare-earth element concentrations in fossil biogenic apatites and implications for paleoenvironmental reconstructions. Chem. Geol. 1999, 155, 233–241. [Google Scholar] [CrossRef]
- Rousseau, T.C.C.; Sonke, J.E.; Chmeleff, J.; van Beek, P.; Souhat, M.; Boaventura, G.; Seyler, P.; Jeandel, C. Rapid neodymium release to marine waters from lithogenic sediments in the Amazon estuary. Nat. Commun. 2015, 6, 7592. [Google Scholar] [CrossRef] [PubMed]
- Sholkovitz, E.R. The geochemistry of rare earth elements in the Amazon River estuary. Geochim. Cosmochim. Acta 1993, 57, 2181–2190. [Google Scholar] [CrossRef]
- Sholkovitz, E.R.; Landing, W.M.; Lewis, B.L. Ocean particle chemistry: The fractionation of rare earth elements between suspended particles and seawater. Geochim. Cosmochim. Acta 1994, 58, 1567–1579. [Google Scholar] [CrossRef]
- Smedley, P.L. The geochemistry of rare earth elements in groundwater from the Carnmenellis area, southwest England. Geochim. Cosmochim. Acta 1991, 55, 2767–2779. [Google Scholar] [CrossRef]
- Tang, J.; Johannesson, K.H. Controls on the geochemistry of rare earth elements along a groundwater flow path in the Carrizo Sand aquifer, Texas, USA. Chem. Geol. 2006, 225, 156–171. [Google Scholar] [CrossRef]
- Tang, J.; Johannesson, K.H. Speciation of rare earth elements in natural terrestrial waters: Assessing the role of dissolved organic matter from the modeling approach. Geochim. Cosmochim. Acta 2003, 67, 2321–2339. [Google Scholar] [CrossRef]
- Trueman, C.N.; Behrensmeyer, A.K.; Potts, R.; Tuross, N. High-resolution records of location and stratigraphic provenance from the rare earth element composition of fossil bones. Geochim. Cosmochim. Acta 2006, 70, 4343–4355. [Google Scholar] [CrossRef]
- van de Flierdt, T.; Pahnke, K.; Amakawa, H.; Andersson, P.; Basak, C.; Coles, B.; Colin, C.; Crocket, K.; Frank, M.; Frank, N.; et al. GEOTRACES intercalibration of neodymium isotopes and rare earth element concentrations in seawater and suspended particles. Part 1: Reproducibility of results for the international intercomparison. Limnol. Oceanogr. Methods 2012, 10, 234–251. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.-L.; Yamada, M. Geochemistry of dissolved rare earth elements in the Equatorial Pacific Ocean. Environ. Geol. 2007, 52, 779–787. [Google Scholar] [CrossRef]
- Zhang, J.; Nozaki, Y. Rare earth elements and yttrium in seawater: ICP-MS determinations in the East Caroline, Coral Sea, and South Fiji basins of the western South Pacific Ocean. Geochim. Cosmochim. Acta 1996, 60, 4631–4644. [Google Scholar] [CrossRef]
- Zheng, W.; Schweitzer, M.H. Chemical Analyses of Fossil Bone. In Forensic Microscopy for Skeletal Tissues: Methods and Protocols; Bell, L.S., Ed.; Humana Press: Totowa, NJ, USA, 2012; pp. 153–172. ISBN 978-1-61779-976-1. [Google Scholar]
- Zheng, X.-Y.; Plancherel, Y.; Saito, M.A.; Scott, P.M.; Henderson, G.M. Rare earth elements (REEs) in the tropical South Atlantic and quantitative deconvolution of their non-conservative behavior. Geochim. Cosmochim. Acta 2016, 177, 217–237. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ref. Code | Score | Mineral | Displacement [°2Th.] | Scale Factor | Chemical Formula | Semi-Quantitative Analysis |
|---|---|---|---|---|---|---|
| 01-071-1663 | 43 | Dolomite | 0.000 | 0.334 | Mg0.1Ca0.9CO3 | 5% |
| 01-084-1999 | 29 | Chloroapatite | 0.000 | 0.437 | Ca5(PO4)3F0.09Cl0.88 | 17% |
| 01-071-0880 | 62 | Fluoroapatite | 0.000 | 0.934 | Ca5(PO4)3F | 41% |
| 01-073-1731 | 67 | Hydroxyapatite | 0.000 | 0.866 | Ca5(PO4)3(OH) | 36% |
| Element | Concentration |
|---|---|
| Sc | 4.96 |
| Mn | 3233 |
| Fe | 0.19 |
| Sr | 1747 |
| Y | 505 |
| Ba | 447 |
| Th | 0.23 |
| U | 18.64 |
| Light Rare Earth Elements (LREEs) | |
| La | 398.95 |
| Ce | 762.75 |
| Pr | 90.67 |
| Nd | 371.52 |
| Middle Rare Earth Elements (MREEs) | |
| Sm | 78.35 |
| Eu | 16.91 |
| Gd | 84.46 |
| Heavy Rare Earth Elements (HREEs) | |
| Tb | 10.67 |
| Dy | 63.76 |
| Ho | 13.32 |
| Er | 36.67 |
| Tm | 4.69 |
| Yb | 27.38 |
| Lu | 4.23 |
| ∑REE | 1964 |
| (Ce/Ce*)N | 0.94 |
| (Pr/Pr*)N | 0.92 |
| (Ce/Ce**)N | 1.22 |
| (La/La*)N | 1.75 |
| Y/Ho | 37.90 |
| Clear DMD Kink for LREE? | Relative Noise in Outer Cortex for La | REE Suggest Flow in Marrow Cavity? | Relative ∑REE Content (Whole Bone) | Relative U Content (Whole Bone) | Relative Porosity of the Cortex |
|---|---|---|---|---|---|
| No | Low | No | Moderate | Low | Low |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schroeter, E.R.; Ullmann, P.V.; Macauley, K.; Ash, R.D.; Zheng, W.; Schweitzer, M.H.; Lacovara, K.J. Soft-Tissue, Rare Earth Element, and Molecular Analyses of Dreadnoughtus schrani, an Exceptionally Complete Titanosaur from Argentina. Biology 2022, 11, 1158. https://doi.org/10.3390/biology11081158
Schroeter ER, Ullmann PV, Macauley K, Ash RD, Zheng W, Schweitzer MH, Lacovara KJ. Soft-Tissue, Rare Earth Element, and Molecular Analyses of Dreadnoughtus schrani, an Exceptionally Complete Titanosaur from Argentina. Biology. 2022; 11(8):1158. https://doi.org/10.3390/biology11081158
Chicago/Turabian StyleSchroeter, Elena R., Paul V. Ullmann, Kyle Macauley, Richard D. Ash, Wenxia Zheng, Mary H. Schweitzer, and Kenneth J. Lacovara. 2022. "Soft-Tissue, Rare Earth Element, and Molecular Analyses of Dreadnoughtus schrani, an Exceptionally Complete Titanosaur from Argentina" Biology 11, no. 8: 1158. https://doi.org/10.3390/biology11081158