
Reduced Retinoic Acid Receptor Beta (Rarβ) Affects Pancreatic β-Cell Physiology

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. mRNA Microarray and RNA-Sequencing Data
2.2. Analysis of Genetic Variants in Rarβ for the Association with T2D
2.3. Cell Culture
2.4. RNA Interference
2.5. Insulin Secretion Measurement
2.6. Western Blot Analysis
2.7. Methylthiazolyldiphenyl-Tetrazolium Bromide (MTT) Cell Viability Assay
2.8. Quantitative-PCR
2.9. Apoptosis Assay
2.10. Glucose Uptake Assay
2.11. ROS Production
2.12. Statistical Analysis
3. Results
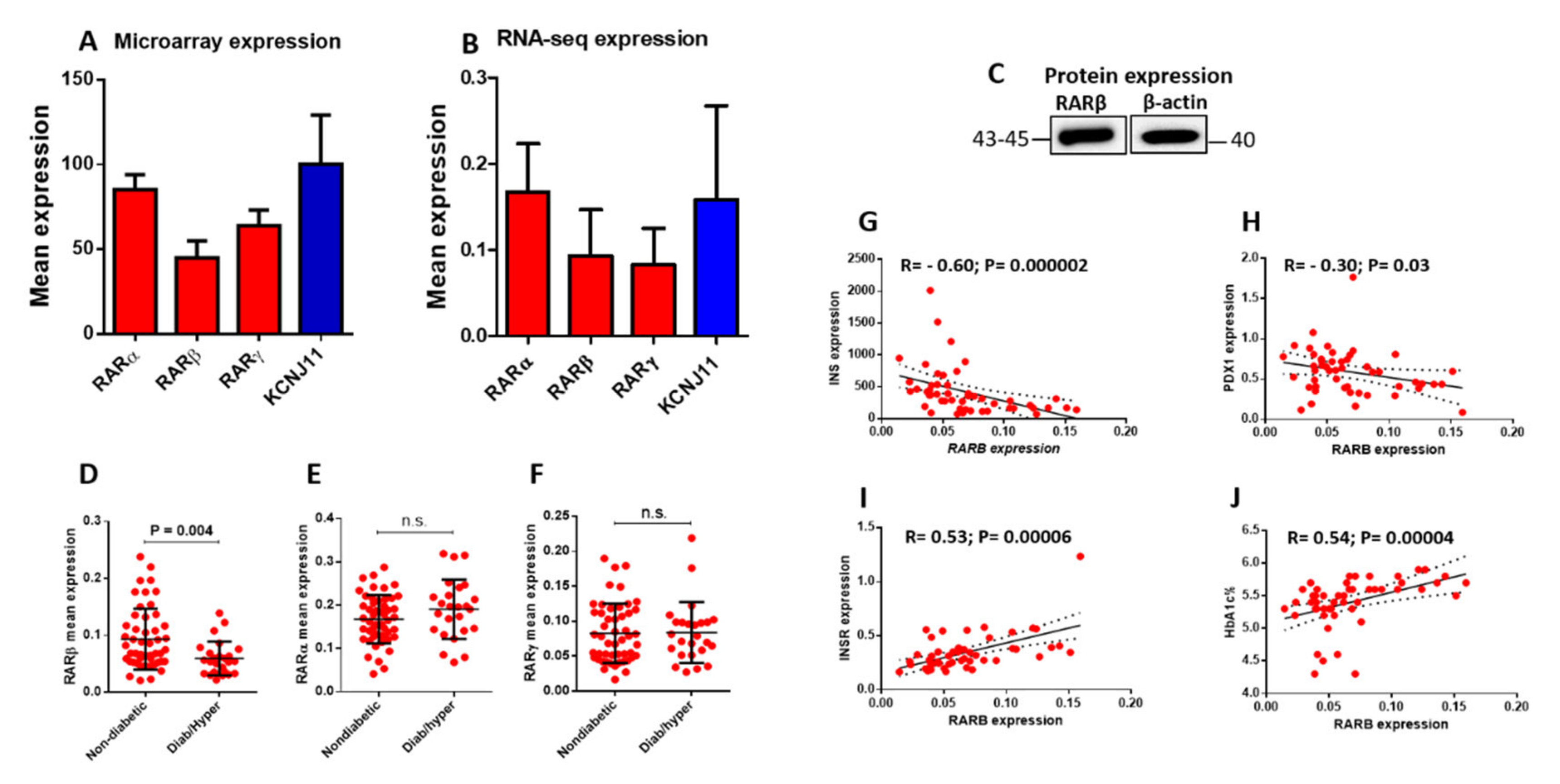
3.1. Expression of RARs in Human Pancreatic Islets
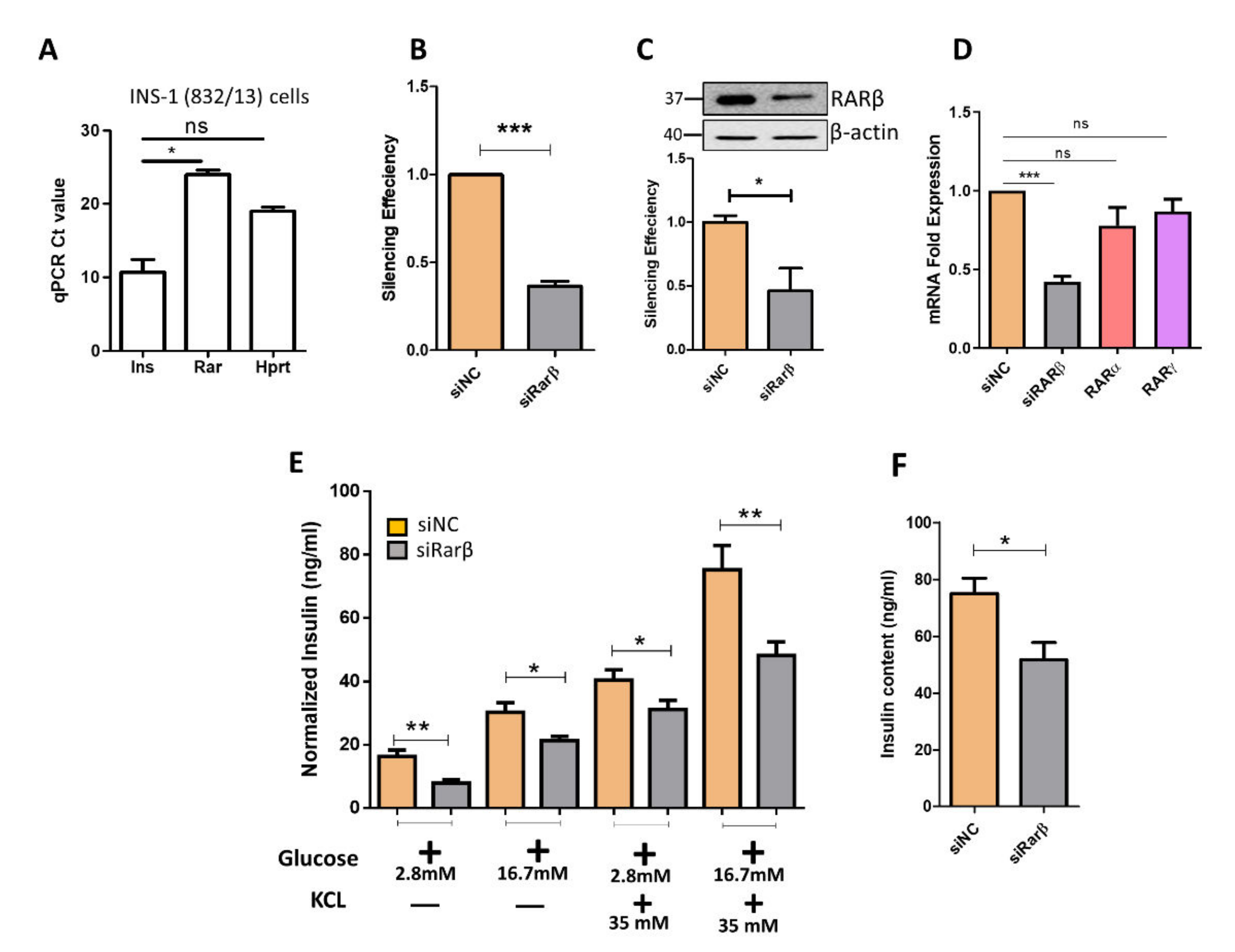
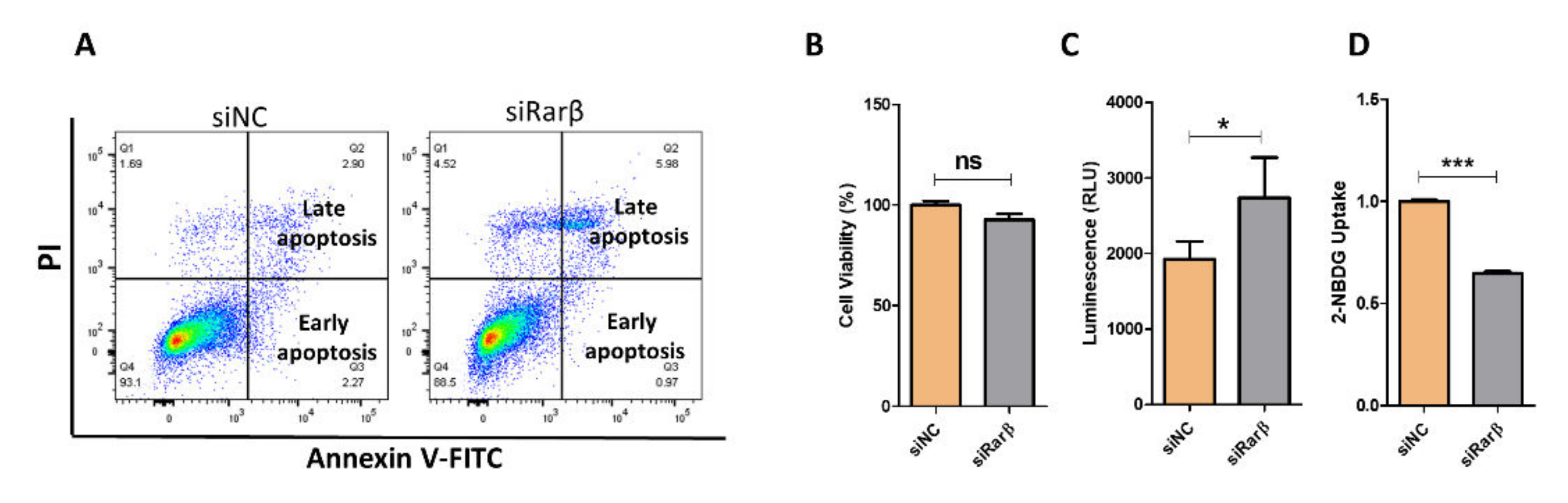
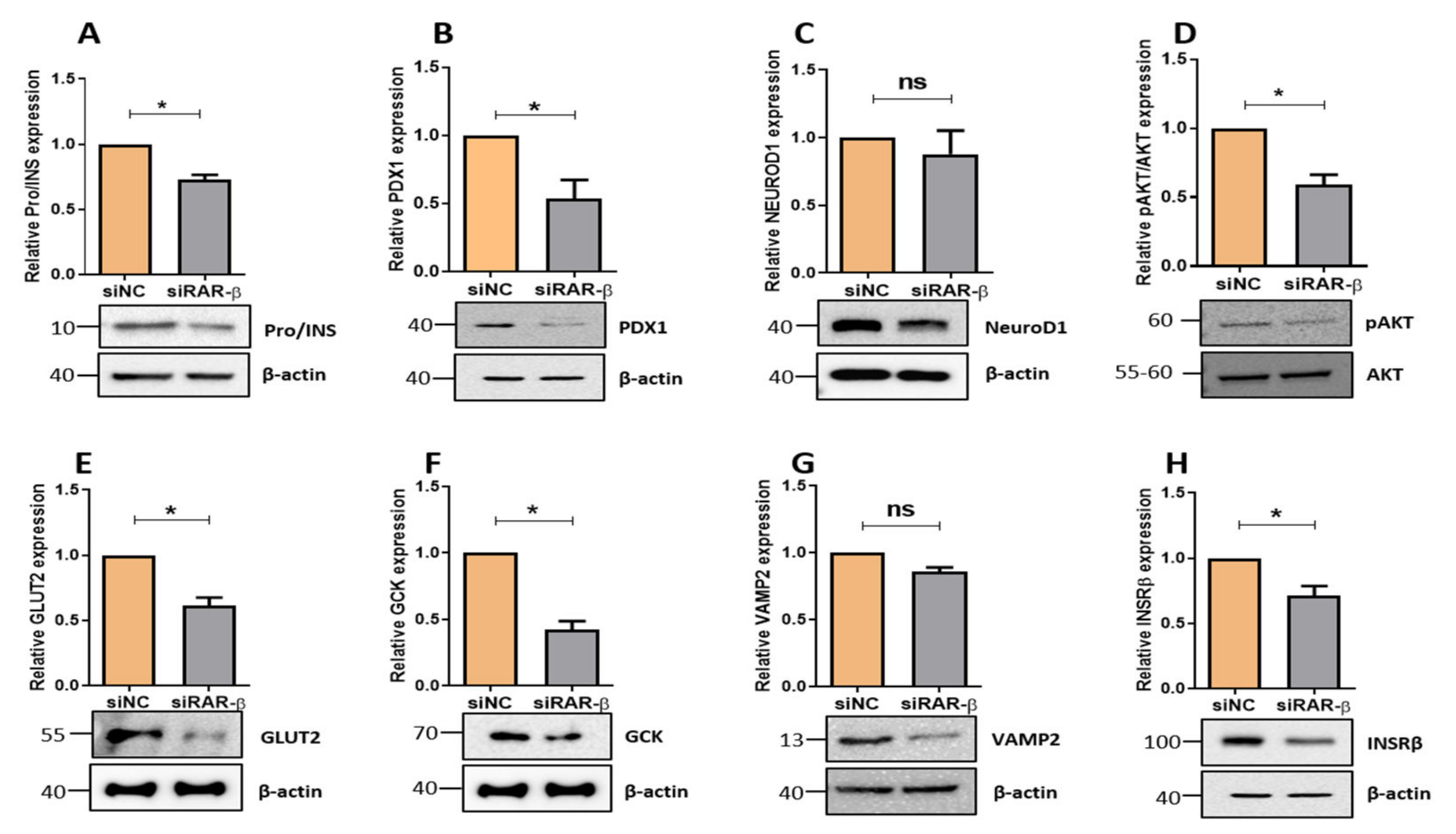
3.2. Silencing of Rarβ Impairs Insulin Secretion, ROS Production and Glucose Uptake
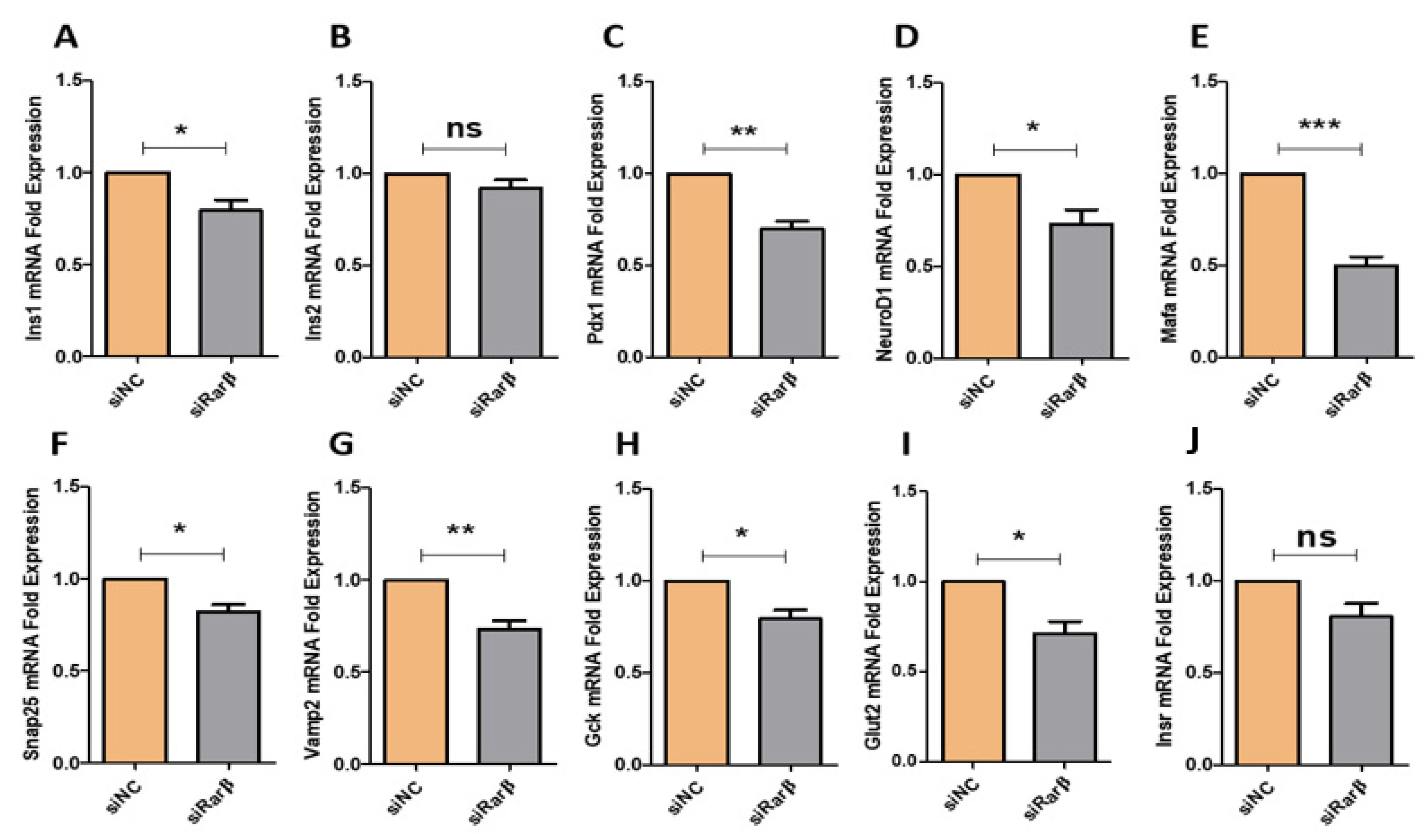
3.3. Silencing of Rarβ in INS-1 Cells Influences Key β-Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tremblay, J.; Hamet, P. Environmental and genetic contributions to diabetes. Metabolism 2019, 100, 153952. [Google Scholar] [CrossRef] [PubMed]
- Petroni, M.L.; Brodosi, L.; Marchignoli, F.; Sasdelli, A.S.; Caraceni, P.; Marchesini, G.; Ravaioli, F. Nutrition in Patients with Type 2 Diabetes: Present Knowledge and Remaining Challenges. Nutrients 2021, 13, 2748. [Google Scholar] [CrossRef] [PubMed]
- Khazrai, Y.M.; Defeudis, G.; Pozzilli, P. Effect of diet on type 2 diabetes mellitus: A review. Diabetes/Metab. Res. Rev. 2014, 30, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.; Lee, S.-M.; Heo, J.; Kwon, Y.M.; Chung, D.; Yu, W.-J.; Bae, S.S.; Choi, G.; Lee, D.-S.; Kim, Y. Retinaldehyde Dehydrogenase Inhibition-Related Adverse Outcome Pathway: Potential Risk of Retinoic Acid Synthesis Inhibition during Embryogenesis. Toxins 2021, 13, 739. [Google Scholar] [CrossRef]
- Kartasheva-Ebertz, D.M.; Pol, S.; Lagaye, S. Retinoic Acid: A New Old Friend of IL-17A in the Immune Pathogeny of Liver Fibrosis. Front. Immunol. 2021, 12, 2295. [Google Scholar] [CrossRef]
- Dollé, P.; Niederreither, K. The Retinoids: Biology, Biochemistry, and Disease; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Yang, H.Y.; Liu, M.; Sheng, Y.; Zhu, L.; Jin, M.M.; Jiang, T.X.; Yang, L.; Liu, P.H.; Liu, X.D.; Liu, L. All-trans retinoic acid impairs glucose-stimulated insulin secretion by activating the RXR/SREBP-1c/UCP2 pathway. Acta Pharmacol. Sin. 2022, 43, 1441–1452. [Google Scholar] [CrossRef]
- Matthews, K.A.; Rhoten, W.B.; Driscoll, H.K.; Chertow, B.S. Vitamin A deficiency impairs fetal islet development and causes subsequent glucose intolerance in adult rats. J. Nutr. 2004, 134, 1958–1963. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, S.; Naseem, I. Role of vitamin A in type 2 diabetes mellitus biology: Effects of intervention therapy in a deficient state. Nutrition 2015, 31, 901–907. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, J.; Sun, B.; Xu, W.; Zhong, M.; Li, Y.; He, C.; Chen, Y.; Wang, X.; Jones, P.; et al. Vitamin A deficiency causes islet dysfunction by inducing islet stellate cell activation via cellular retinol binding protein 1. Int. J. Biol. Sci. 2020, 16, 947–956. [Google Scholar] [CrossRef]
- Lorberbaum, D.S.; Kishore, S.; Rosselot, C.; Sarbaugh, D.; Brooks, E.P.; Aragon, E.; Xuan, S.; Simon, O.; Ghosh, D.; Mendelsohn, C.; et al. Retinoic acid signaling within pancreatic endocrine progenitors regulates mouse and human β cell specification. Development 2020, 147, dev189977. [Google Scholar] [CrossRef]
- Trasino, S.E.; Benoit, Y.D.; Gudas, L.J. Vitamin A Deficiency Causes Hyperglycemia and Loss of Pancreatic β-Cell Mass. J. Biol. Chem. 2015, 290, 1456–1473. [Google Scholar] [CrossRef] [Green Version]
- Kane, M.A.; Folias, A.E.; Pingitore, A.; Perri, M.; Obrochta, K.M.; Krois, C.R.; Cione, E.; Ryu, J.Y.; Napoli, J.L. Identification of 9-cis-retinoic acid as a pancreas-specific autacoid that attenuates glucose-stimulated insulin secretion. Proc. Natl. Acad. Sci. USA 2010, 107, 21884–21889. [Google Scholar] [CrossRef] [Green Version]
- Raghow, R. Metabolic balancing acts of vitamin A in type-2 diabetes and obesity. World J. Diabetes 2012, 3, 174–177. [Google Scholar] [CrossRef]
- Blaner, W.S. Vitamin A signaling and homeostasis in obesity, diabetes, and metabolic disorders. Pharmacol. Ther. 2019, 197, 153–178. [Google Scholar] [CrossRef]
- Taneera, J.; Awadallah, S.; Mohammed, A.K.; Unnikannan, H.; Sulaiman, N. Vitamin A levels are decreased but not influenced by glucose- or lipid-lowering medications in subjects with type 2 diabetes. Saudi J. Biol. Sci. 2020, 28, 572–577. [Google Scholar] [CrossRef]
- Trasino, S.E.; Gudas, L.J. Vitamin A: A missing link in diabetes? Diabetes Manag. 2015, 5, 359. [Google Scholar] [CrossRef] [Green Version]
- Chertow, B.S.; Blaner, W.S.; Baranetsky, N.G.; Sivitz, W.; Cordle, M.B.; Thompson, D.; Meda, P. Effects of vitamin A deficiency and repletion on rat insulin secretion in vivo and in vitro from isolated islets. J. Clin. Investig. 1987, 79, 163–169. [Google Scholar] [CrossRef]
- Fernandez-Mejia, C.; German, M.; Matschinsky, F.; Magnuson, M. Regulation of glucokinase by vitamins and hormones. Front. Diabetes 2004, 16, 240–248. [Google Scholar]
- Perissi, V.; Rosenfeld, M.G. Controlling nuclear receptors: The circular logic of cofactor cycles. Nat. Rev. Mol. Cell Biol. 2005, 6, 542–554. [Google Scholar] [CrossRef]
- Brun, P.J.; Grijalva, A.; Rausch, R.; Watson, E.; Yuen, J.J.; Das, B.C.; Shudo, K.; Kagechika, H.; Leibel, R.L.; Blaner, W.S. Retinoic acid receptor signaling is required to maintain glucose-stimulated insulin secretion and β-cell mass. FASEB J. 2015, 29, 671–683. [Google Scholar] [CrossRef] [Green Version]
- Pérez, R.J.; Benoit, Y.D.; Gudas, L.J. Deletion of retinoic acid receptor β (RARβ) impairs pancreatic endocrine differentiation. Exp. Cell Res. 2013, 319, 2196–2204. [Google Scholar] [CrossRef] [Green Version]
- Chertow, B.S.; Goking, N.Q.; Driscoll, H.K.; Primerano, D.A.; Matthews, K.A. Effects of All-trans-Retinoic Acid (ATRA) and Retinoic Acid Receptor (RAR) Expression on Secretion, Growth, and Apoptosis of Insulin-Secreting RINm5F Cells. Pancreas 1997, 15, 122–131. [Google Scholar] [CrossRef]
- Chertow, B.S.; Driscoll, H.K.; Goking, N.Q.; Primerano, D.; Cordle, M.B.; Matthews, K.A. Retinoid-X receptors and the effects of 9-cis-retinoic acid on insulin secretion from RINm5F cells. Metabolism 1997, 46, 656–660. [Google Scholar] [CrossRef]
- Fadista, J.; Vikman, P.; Laakso, E.O.; Mollet, I.G.; Esguerra, J.L.; Taneera, J.; Storm, P.; Osmark, P.; Ladenvall, C.; Prasad, R.B.; et al. Global genomic and transcriptomic analysis of human pancreatic islets reveals novel genes influencing glucose metabolism. Proc. Natl. Acad. Sci. USA 2014, 111, 13924–13929. [Google Scholar] [CrossRef] [Green Version]
- Taneera, J.; Fadista, J.; Ahlqvist, E.; Atac, D.; Ottosson-Laakso, E.; Wollheim, C.B.; Groop, L. Identification of novel genes for glucose metabolism based upon expression pattern in human islets and effect on insulin secretion and glycemia. Hum. Mol. Genet. 2014, 24, 1945–1955. [Google Scholar] [CrossRef] [Green Version]
- Taneera, J.; Mohammed, A.K.; Dhaiban, S.; Hamad, M.; Prasad, R.B.; Sulaiman, N.; Salehi, A. RORB and RORC associate with human islet dysfunction and inhibit insulin secretion in INS-1 cells. Islets 2019, 11, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Taneera, J.; Mohammed, I.; Mohammed, A.K.; Hachim, M.; Dhaiban, S.; Malek, A.; Dunér, P.; Elemam, N.M.; Sulaiman, N.; Hamad, M.; et al. Orphan G-protein coupled receptor 183 (GPR183) potentiates insulin secretion and prevents glucotoxicity-induced β-cell dysfunction. Mol. Cell. Endocrinol. 2019, 499, 110592. [Google Scholar] [CrossRef]
- Voloboueva, L.A.; Liu, J.; Suh, J.H.; Ames, B.N.; Miller, S.S. (R)-α-Lipoic Acid Protects Retinal Pigment Epithelial Cells from Oxidative Damage. Investig. Opthalmol. Vis. Sci. 2005, 46, 4302–4310. [Google Scholar] [CrossRef] [Green Version]
- Ashcroft, F.M.; Harrison, D.E.; Ashcroft, S.J.H. Glucose induces closure of single potassium channels in isolated rat pancreatic β-cells. Nature 1984, 312, 446–448. [Google Scholar] [CrossRef]
- Berry, D.C.; Noy, N. All- trans -Retinoic Acid Represses Obesity and Insulin Resistance by Activating both Peroxisome Proliferation-Activated Receptor β/δ and Retinoic Acid Receptor. Mol. Cell. Biol. 2009, 29, 3286–3296. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.C.; Kim, C.-K.; Axe, D.; Cook, A.; Lee, M.; Li, T.; Smallwood, N.; Chiang, J.Y.; Hardwick, J.P.; Moore, D.D.; et al. All-trans-retinoic acid ameliorates hepatic steatosis in mice by a novel transcriptional cascade. Hepatology 2013, 59, 1750–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chertow, B.S.; Blaner, W.S.; Rajan, N.; Primerano, D.A.; Meda, P.; Cirulli, V.; Krozowski, Z.; Smith, R.; Cordle, M.B. Retinoic acid receptor, cytosolic retinol-binding and retinoic acid-binding protein mRNA transcripts and proteins in rat insulin-secreting cells. Diabetes 1993, 42, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Gudas, L.J. Cellular biology and biochemistry of the retinoids. Retinoids 1994, 443–520. [Google Scholar]
- Ottosson-Laakso, E.; Krus, U.; Storm, P.; Prasad, R.B.; Oskolkov, N.; Ahlqvist, E.; Fadista, J.; Hansson, O.; Groop, L.; Vikman, P. Glucose-Induced Changes in Gene Expression in Human Pancreatic Islets: Causes or Consequences of Chronic Hyperglycemia. Diabetes 2017, 66, 3013–3028. [Google Scholar] [CrossRef] [Green Version]
- Rossetti, S.; Sacchi, N. Emerging Cancer Epigenetic Mechanisms Regulated by All-Trans Retinoic Acid. Cancers 2020, 12, 2275. [Google Scholar] [CrossRef]
- El-Metwally, T.H.; Hussein, M.R.; Abd-El-Ghaffar, S.K.; Abo-El-Naga, M.M.; Ulrich, A.B.; Pour, P.M. Retinoic acid can induce markers of endocrine transdifferentiation in pancreatic ductal adenocarcinoma: Preliminary observations from an in vitro cell line model. J. Clin. Pathol. 2006, 59, 603–610. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Guleria, R.S.; Zhu, S.; Baker, K.M. Molecular Mechanisms of Retinoid Receptors in Diabetes-Induced Cardiac Remodeling. J. Clin. Med. 2014, 3, 566–594. [Google Scholar] [CrossRef] [Green Version]
- Clark, A.R.; Wilson, M.E.; London, N.J.M.; James, R.F.L.; Docherty, K. Identification and characterization of a functional retinoic acid/thyroid hormone-response element upstream of the human insulin gene enhancer. Biochem. J. 1995, 309, 863–870. [Google Scholar] [CrossRef] [Green Version]
- Jara, M.A.; Werneck-De-Castro, J.P.; Lubaczeuski, C.; Johnson, J.D.; Bernal-Mizrachi, E. Pancreatic and duodenal homeobox-1 (PDX1) contributes to β-cell mass expansion and proliferation induced by Akt/PKB pathway. Islets 2020, 12, 32–40. [Google Scholar] [CrossRef]
- Jonsson, J.; Carlsson, L.; Edlund, T.; Edlund, H. Insulin-promoter-factor 1 is required for pancreas development in mice. Nature 1994, 371, 606–609. [Google Scholar] [CrossRef]
- Wang, J.; Gu, W.; Chen, C. Knocking down Insulin Receptor in Pancreatic Beta Cell lines with Lentiviral-Small Hairpin RNA Reduces Glucose-Stimulated Insulin Secretion via Decreasing the Gene Expression of Insulin, GLUT2 and Pdx. Int. J. Mol. Sci. 2018, 19, 985. [Google Scholar] [CrossRef] [Green Version]
- Aljaibeji, H.; Mukhopadhyay, D.; Mohammed, A.K.; Dhaiban, S.; Hachim, M.; Elemam, N.M.; Sulaiman, N.; Salehi, A.; Taneera, J. Reduced Expression of PLCXD3 Associates With Disruption of Glucose Sensing and Insulin Signaling in Pancreatic β-Cells. Front. Endocrinol. 2019, 10, 735. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Tu, J.; Zhang, S.; Cai, B.; Liu, Z.; Hou, S.; Zhong, Q.; Hu, X.; Liu, W.; Li, G.; et al. Different regions of synaptic vesicle membrane regulate VAMP2 conformation for the SNARE assembly. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.C.; Min, L.; Pessin, J.E. Insulin Granule Biogenesis, Trafficking and Exocytosis. Vitam. Horm. 2009, 80, 473–506. [Google Scholar] [CrossRef] [Green Version]
- Widerska, E.; Strycharz, J.; Wróblewski, A.; Szemraj, J.; Drzewoski, J.; Śliwińska, A. Role of PI3K/AKT pathway in insulin-mediated glucose uptake. Blood Glucose Levels 2018, 1, 1–18. [Google Scholar]
- Bastien, J.; Adam-Stitah, S.; Plassat, J.-L.; Chambon, P.; Rochette-Egly, C. The Phosphorylation Site Located in the A Region of Retinoic X Receptor α Is Required for the Antiproliferative Effect of Retinoic Acid (RA) and the Activation of RA Target Genes in F9 Cells. J. Biol. Chem. 2002, 277, 28683–28689. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Reference Allele | Alternate Allele | p-Value | Beta |
|---|---|---|---|---|
| rs6804842 | A | G | 1.2 × 10−25 | ▲0.0156 |
| rs10510554 | T | C | 5.6 × 10−23 | ▲0.0156 |
| rs7619139 | T | A | 2.6 × 10−21 | ▲0.0154 |
| rs4858697 | A | G | 1.6 × 10−20 | ▲0.0149 |
| rs1609783 | G | A | 2.7 × 10−20 | ▲0.0149 |
| rs12632128 | G | T | 8.3 × 10−20 | ▲0.0146 |
| rs6767671 | G | T | 2.4 × 10−19 | ▲0.0147 |
| rs4858696 | C | T | 2 × 10−18 | ▲0.0146 |
| rs9847186 | G | A | 3.3 × 10−18 | ▼−0.0140 |
| rs4353774 | A | C | 3.3 × 10−18 | ▲0.0143 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalique, A.; Mohammed, A.K.; Al-khadran, N.M.; Gharaibeh, M.A.; Abu-Gharbieh, E.; El-Huneidi, W.; Sulaiman, N.; Taneera, J. Reduced Retinoic Acid Receptor Beta (Rarβ) Affects Pancreatic β-Cell Physiology. Biology 2022, 11, 1072. https://doi.org/10.3390/biology11071072
Khalique A, Mohammed AK, Al-khadran NM, Gharaibeh MA, Abu-Gharbieh E, El-Huneidi W, Sulaiman N, Taneera J. Reduced Retinoic Acid Receptor Beta (Rarβ) Affects Pancreatic β-Cell Physiology. Biology. 2022; 11(7):1072. https://doi.org/10.3390/biology11071072
Chicago/Turabian StyleKhalique, Anila, Abdul Khader Mohammed, Nujood Mohammed Al-khadran, Mutaz Al Gharaibeh, Eman Abu-Gharbieh, Waseem El-Huneidi, Nabil Sulaiman, and Jalal Taneera. 2022. "Reduced Retinoic Acid Receptor Beta (Rarβ) Affects Pancreatic β-Cell Physiology" Biology 11, no. 7: 1072. https://doi.org/10.3390/biology11071072