
Hybridization between Crotalus aquilus and Crotalus polystictus Species: A Comparison of Their Venom Toxicity and Enzymatic Activities

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Phenetic Analysis
2.2. Venom Harvest
2.3. Polyacrylamide Gel Electrophoresis
2.4. Zymography Assays
2.4.1. Serine Proteases Zymography
2.4.2. Metalloprotease Zymography
2.4.3. PLA2 Zymography
2.4.4. Hyaluronidases Zymography
2.5. Enzymatic Assays
2.5.1. Serine Proteases
2.5.2. Metalloprotease Proteolytic Activity
2.5.3. Phospholipase A2 (PLA2) Activity
2.5.4. Hyaluronidase Activity
2.6. RP-HPLC
2.7. MALDI-TOF-MS
2.8. Acute Toxicological Assay
3. Results and Discussion
3.1. Phenetic Analysis
3.2. Enzymatic Activity and Zymography
3.2.1. Snake Venom Serine Proteases (SVSPs)
3.2.2. Gelatinolytic Activity
3.2.3. Snake Venom Hyaluronidases (SVHs)
3.2.4. Snake Venom Phospholipase A2 (SVPLA2) Activity
3.3. RP-HPLC and MALDI-TOF-MS

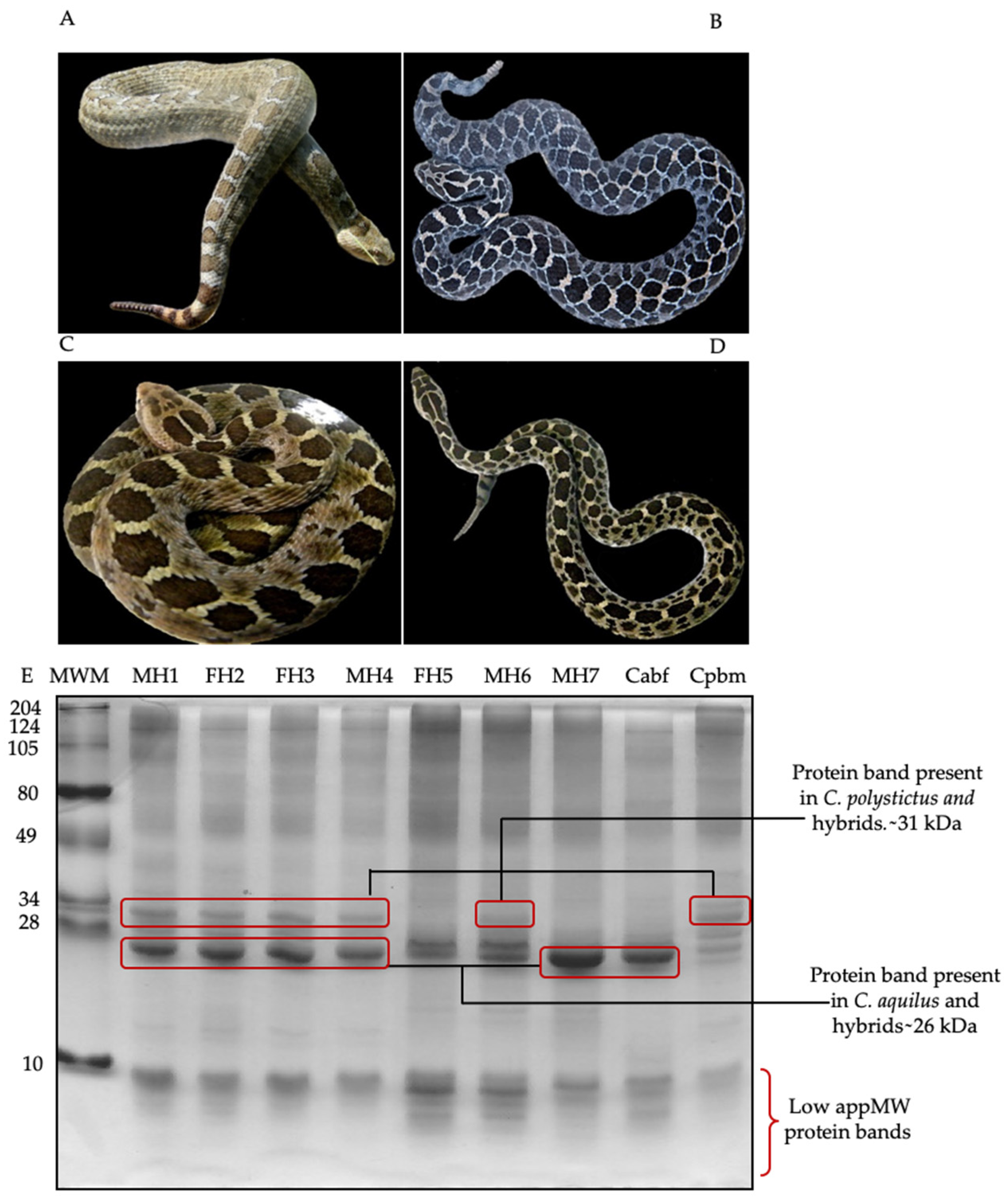{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Family | Data Reported | MCa-PV | FCp-PV | MH | FH |
|---|---|---|---|---|---|
| SVMP | Reported for C. polistictus in the range of 20 and 30–40 kDa [50,51] | 23.2 kDa | 23 kDa | 23 kDa | 23.4, 37.8, and 46.2 kDa |
| SVSP | Reported for C. molossus venom between 24 and 30 kDa [49] | 27 kDa | 24.7 and 29 kDa | 24.7 kDa | NF |
| SVPLA2 | Reported in the range of 13–18 kDa [50] | 12.4, 16.2, and 13.9 kDa | 11.5, 12.4, 13.9, and 16.2 kDa | 11.5, 12.3, and 13.6 kDa | 11.6, 13.9, and 17.6 kDa |
| MYO | Reported in the range of 4–5 kDa in snake venoms and one MYO of 10 kDa in C. molossus [49] | 6.9 and 7.7 kDa | 7.4 kDa | 3.7, 4.8, 6.8, and 7.4 kDa | 3.7, 5.8, 6.9, 7.7, 8.7, and 9.7 kDa |
3.4. Lethal Dose-50 (LD50)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casewell, N.R.; Jackson, T.N.W.; Laustsen, A.H.; Sunagar, K. Causes and Consequences of Snake Venom Variation. Trends Pharmacol. Sci. 2020, 41, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P.; Williams, V.; White, J. Snake Venom Variability: Methods of Study, Results and Interpretation. Toxicon 1991, 29, 1279–1303. [Google Scholar] [CrossRef]
- Zancolli, G.; Baker, T.G.; Barlow, A.; Bradley, R.K.; Calvete, J.J.; Carter, K.C.; de Jager, K.; Owens, J.B.; Price, J.F.; Sanz, L.; et al. Is Hybridization a Source of Adaptive Venom Variation in Rattlesnakes? A Test, Using a Crotalus Scutulatus × Viridis Hybrid Zone in Southwestern New Mexico. Toxins 2016, 8, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, V.J. Inventing an Arsenal: Adaptive Evolution and Neofunctionalization of Snake Venom Phospholipase A2 Genes. BMC Evol. Biol. 2007, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, D.M.; Tasima, L.J.; Bravo-Tobar, C.A.; Serino-Silva, C.; Tashima, A.K.; Rodrigues, C.F.B.; da Silva Aguiar, W.; da Costa Galizio, N.; de Lima, E.O.V.; Kavazoi, V.K.; et al. Venom Complexity of Bothrops Atrox (Common Lancehead) Siblings. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200018. [Google Scholar] [CrossRef]
- Massey, D.J.; Calvete, J.J.; Sánchez, E.E.; Sanz, L.; Richards, K.; Curtis, R.; Boesen, K. Venom Variability and Envenoming Severity Outcomes of the Crotalus Scutulatus Scutulatus (Mojave Rattlesnake) from Southern Arizona. J. Proteom. 2012, 75, 2576–2587. [Google Scholar] [CrossRef]
- Kindler, C.; Chèvre, M.; Ursenbacher, S.; Böhme, W.; Hille, A.; Jablonski, D.; Vamberger, M.; Fritz, U. Hybridization Patterns in Two Contact Zones of Grass Snakes Reveal a New Central European Snake Species. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zinenko, O.; Sovic, M.; Joger, U.; Gibbs, H.L. Hybrid Origin of European Vipers (Vipera Magnifica and Vipera Orlovi) from the Caucasus Determined Using Genomic Scale DNA Markers. BMC Evol. Biol. 2016, 16, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lewontin, R.C.; Birch, L.C. Hybridization as a Source of Variation for Adaptation to New Environments. Evolution 1966, 20, 315. [Google Scholar] [CrossRef] [Green Version]
- Harrison, R.G.; Larson, E.L. Hybridization, Introgression, and the Nature of Species Boundaries. J. Hered. 2014, 105, 795–809. [Google Scholar] [CrossRef] [Green Version]
- Santoro, M.L.; do Carmo, T.; Cunha, B.H.L.; Alves, A.F.; Zelanis, A.; Serrano, S.M.D.T.; Grego, K.F.; Sant’Anna, S.S.; Barbaro, K.C.; Fernandes, W. Ontogenetic Variation in Biological Activities of Venoms from Hybrids between Bothrops Erythromelas and Bothrops Neuwiedi Snakes. PLoS ONE 2015, 10, e0145516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowling, T.E.; Secor, C.L. The role of hybridization and introgression in the diversification of animals. Annu. Rev. Ecol. Syst. 1997, 28, 593–619. [Google Scholar] [CrossRef] [Green Version]
- Bullini, L. Origin and Evolution of Animal Hybrid Species. Trends Ecol. Evol. 1994, 9, 422–426. [Google Scholar] [CrossRef]
- Bailey, R.M. An Intergeneric Hybrid Rattlesnake. Am. Nat. 2015, 76, 376–385. [Google Scholar] [CrossRef]
- Aird, S.D.; Thirkhill, L.J.; Seebart, C.S.; Kaiser, I.I. Venoms and Morphology of Western Diamondback/Mojave Rattlesnake Hybrids. J. Herpetol. 1989, 23, 131. [Google Scholar] [CrossRef]
- Smith, C.F.; Mackessy, S.P. The Effects of Hybridization on Divergent Venom Phenotypes: Characterization of Venom from Crotalus Scutulatus Scutulatus × Crotalus Oreganus Helleri Hybrids. Toxicon 2016, 120, 110–123. [Google Scholar] [CrossRef]
- Aird, S.D.; Aggarwal, S.; Villar-Briones, A.; Tin, M.M.-Y.; Terada, K.; Mikheyev, A.S. Snake Venoms Are Integrated Systems, but Abundant Venom Proteins Evolve More Rapidly. BMC Genom. 2015, 16, 647. [Google Scholar] [CrossRef] [Green Version]
- McCranie, J.R. Crotalus Polystictus. Soc. Study Amphib. Reptiles 1976, 180, 1–2. [Google Scholar]
- Campbell, J.A.; Lamar, W.W. The Venomous Reptiles of Latin America. Q. Rev. Biol. 1989, 65, 516–517. [Google Scholar] [CrossRef]
- Meik, J.; Mociño-Deloya, E.; Setser, K. New Distribution Records for the Queretero Dusky Rattlesnake Crotalus Aquilus (Viperidae), with Comments on Morphology and Habitat Use. West. N. Am. Nat. 2007, 67, 601–604. [Google Scholar] [CrossRef] [Green Version]
- Oukkache, N.; Lalaoui, M.; Ghalim, N. General Characterization of Venom from the Moroccan Snakes Macrovipera Mauritanica and Cerastes Cerastes. J. Venom. Anim. Toxins Incl. Trop. Dis. 2012, 18, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Alencar, L.R.V.; Quental, T.B.; Grazziotin, F.G.; Alfaro, M.L.; Martins, M.; Venzon, M.; Zaher, H. Diversification in Vipers: Phylogenetic Relationships, Time of Divergence and Shifts in Speciation Rates. Mol. Phylogenetics Evol. 2016, 105, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Maruñak, S.L.; Ruíz de Torrent, R.M.; Teibler, G.P.; Gay, C.C.; Leiva, L.; Acosta De Pérez, O. Acción Del Veneno de Bothrops Jararacussu de Argentina Sobre La Coagulación Sanguínea; Universidad de Buenos Aires, Facultad de Ciencias Veterinarias: Corrientes, Argentina, 2005; Volume 8. [Google Scholar]
- Slagboom, J.; Kool, J.; Harrison, R.A.; Casewell, N.R. Haemotoxic Snake Venoms: Their Functional Activity, Impact on Snakebite Victims and Pharmaceutical Promise. Br. J. Haematol. 2017, 177, 947–959. [Google Scholar] [CrossRef] [Green Version]
- Calvete, J.J. Proteomic Tools against the Neglected Pathology of Snake Bite Envenoming. Expert Rev. Proteom. 2011, 8, 739–758. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; Huang, C. Synergistic Strategies of Predominant Toxins in Snake Venoms. Toxicol. Lett. 2018, 287, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Rohlf, F. NTSYS-Pc: Microcomputer Programs for Numerical Taxonomy and Multivariate Analysis. Am. Stat. 1987, 41, 330. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Ohlsson, B.G.; Weström, B.R.; Karlsson, B.W. Enzymoblotting: A Method for Localizing Proteinases and Their Zymogens Using Para-Nitroanilide Substrates after Agarose Gel Electrophoresis and Transfer to Nitrocellulose. Anal. Biochem. 1986, 152, 239–244. [Google Scholar] [CrossRef]
- Vinokurov, K.S.; Oppert, B.; Elpidina, E.N. Notes & Tips an Overlay Technique for Postelectrophoretic Analysis of Proteinase Spectra in Complex Mixtures Using p-Nitroanilide Substrates. Anal. Biochem. 2005, 337, 164–166. [Google Scholar] [CrossRef]
- Rice, K.; Peralta, R.; Bast, D.; De Azavedo, J.; McGavin, M.J. Description of Staphylococcus Serine Protease (Ssp) Operon in Staphylococcus Aureus and Nonpolar Inactivation of SspA-Encoded Serine Protease. Infect. Immun. 2001, 69, 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossignol, G.; Merieau, A.; Guerillon, J.; Veron, W.; Lesouhaitier, O.; Feuilloley, M.G.J.; Orange, N. Involvement of a Phospholipase C in the Hemolytic Activity of a Clinical Strain of Pseudomonas Fluorescens. BMC Microbiol. 2008, 8, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guntenhöner, M.W.; Pogrel, M.A.; Stern, R. A Substrate-Gel Assay for Hyaluronidase Activity. Matrix 1992, 12, 388–396. [Google Scholar] [CrossRef]
- Erlanger, B.F.; Kokowsky, N.; Cohen, W. The Preparation and Properties of Two New Chromogenic Substrates of Trypsin. Arch. Biochem. Biophys. 1961, 95, 271–278. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Sanz, L.; Escolano, J.; Fernández, J.; Lomonte, B.; Angulo, Y.; Rucavado, A.; Warrell, D.A.; Calvete, J.J. Snake Venomics of the Lesser Antillean Pit Vipers Bothrops Caribbaeus and Bothrops Lanceolatus: Correlation with Toxicological Activities and Immunoreactivity of a Heterologous Antivenom. J. Proteome Res. 2008, 7, 4396–4408. [Google Scholar] [CrossRef]
- Ponce-Soto, L.A.; Bonfim, V.L.; Novello, J.C.; Navarro Oviedo, R.; Yarlequé Chocas, A.; Marangoni, S. Isolation and Characterization of a Serine Protease, Ba III-4, from Peruvian Bothrops Atrox Venom. Protein J. 2007, 26, 387–394. [Google Scholar] [CrossRef]
- Di Ferrante, N. Turbidimetric Measurement of Acid Mucopolysaccharides and Hyaluronidase Activity. J. Biol. Chem. 1956, 220, 303–306. [Google Scholar] [CrossRef]
- NOM-062-ZOO Norma Oficial Mexicana. Especificaciones Técnicas Para La Producción, Cuidado y Uso de Los Animales de Laboratorio (NOM-062-ZOO 1999). D. Of. Fed. 2001, 107–165. Available online: http://www.anmm.org.mx/bgmm/1864_2007/2002-138-3-295-298.pdf (accessed on 20 February 2022).
- Klauber, L.M. Rattlesnakes: Their Habits, Life Histories, and Influence on Mankind; Zoological Society of San Diego, University of California Press: Berkeley, CA, USA, 1972; ISBN 0520017757. [Google Scholar]
- Reyes-Velasco, J.; Meik, J.M.; Smith, E.N.; Castoe, T.A. Phylogenetic Relationships of the Enigmatic Longtailed Rattlesnakes (Crotalus Ericsmithi, C. Lannomi, and C. Stejnegeri). Mol. Phylogenetics Evol. 2013, 69, 524–534. [Google Scholar] [CrossRef]
- Blair, C.; Sánchez-Ramírez, S. Diversity-Dependent Cladogenesis throughout Western Mexico: Evolutionary Biogeography of Rattlesnakes (Viperidae: Crotalinae: Crotalus and Sistrurus). Mol. Phylogenetics Evol. 2016, 97, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Mallet, J. Hybridization as an Invasion of the Genome. Trends Ecol. Evol. 2005, 20, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Roldán-Padrón, O.; Castro-Guillén, J.L.; García-Arredondo, J.A.; Cruz-Pérez, M.S.; Díaz-Peña, L.F.; Saldaña, C.; Blanco-Labra, A.; García-Gasca, T. Snake Venom Hemotoxic Enzymes: Biochemical Comparison between Crotalus Species from Central Mexico. Molecules 2019, 24, 1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casewell, N.R.; Wagstaff, S.C.; Wuster, W.; Cook, D.A.N.; Bolton, F.M.S.; King, S.I.; Pla, D.; Sanz, L.; Calvete, J.J.; Harrison, R.A. Medically Important Differences in Snake Venom Composition Are Dictated by Distinct Postgenomic Mechanisms. Proc. Natl. Acad. Sci. USA 2014, 111, 9205–9210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marangoni, F.A.; Ponce-Soto, L.A.; Marangoni, S.; Landucci, E.C.T. Unmasking Snake Venom of Bothrops Leucurus: Purification and Pharmacological and Structural Characterization of New PLA2 Bleu TX-III. Biomed. Res. Int. 2013, 2013, 941467. [Google Scholar] [CrossRef] [Green Version]
- Lomonte, B.; Calvete, J.J. Strategies in ‘Snake Venomics’ Aiming at an Integrative View of Compositional, Functional, and Immunological Characteristics of Venoms. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 26. [Google Scholar] [CrossRef] [Green Version]
- Menaldo, D.L.; Bernardes, C.P.; Santos-Filho, N.A.; Moura, L.D.A.; Fuly, A.L.; Arantes, E.C.; Sampaio, S.V. Biochemical Characterization and Comparative Analysis of Two Distinct Serine Proteases from Bothrops Pirajai Snake Venom. Biochimie 2012, 94, 2545–2558. [Google Scholar] [CrossRef]
- Borja, M.; Neri-Castro, E.; Pérez-Morales, R.; Strickland, J.; Ponce-López, R.; Parkinson, C.; Espinosa-Fematt, J.; Sáenz-Mata, J.; Flores-Martínez, E.; Alagón, A.; et al. Ontogenetic Change in the Venom of Mexican Black-Tailed Rattlesnakes (Crotalus Molossus Nigrescens). Toxins 2018, 10, 501. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Manjunatha Kini, R.; Doley, R. Snake Venom Phospholipase A2 Enzymes. In Handbook of Venoms and Toxins of Reptiles; CRC Press: Boca Raton, FL, USA, 2009; pp. 173–205. ISBN 978-0-8493-9165-1. [Google Scholar]
- Rael, E.D.; Rivas, J.Z.; Chen, T.; Maddux, N.; Huizar, E.; Lieb, C.S. Differences in Fibrinolysis and Complement Inactivation by Venom from Different Northern Blacktailed Rattlesnakes (Crotalus Molossus Molossus). Toxicon 1997, 35, 505–513. [Google Scholar] [CrossRef]
- Rivas, E.; Neri-Castro, E.; Bénard-Valle, M.; Hernánez-Dávila, A.I.; Zamudio, F.; Alagón, A. General Characterization of the Venoms from Two Species of Rattlesnakes and an Intergrade Population (C. Lepidus × Aquilus) from Aguascalientes and Zacatecas, Mexico. Toxicon 2017, 138, 191–195. [Google Scholar] [CrossRef]
- Mackessy, S. Venom Composition in Rattlesnakes: Trends and Biological Significance. Biol. Ratt. 2008, 495, 510. [Google Scholar]
- Mackessy, S.P.; Leroy, J.; Mociño-Deloya, E.; Setser, K.; Bryson, R.W.; Saviola, A.J. Venom Ontogeny in the Mexican Lance-Headed Rattlesnake (Crotalus Polystictus). Toxins 2018, 10, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amazonas, D.R.; Freitas-de-Sousa, L.A.; Orefice, D.P.; Sousa, L.F.D.; Martinez, M.G.; Mourão, R.H.; Chalkidis, H.M.; Camargo, P.B.; Moura-da-Silva, A.M. Evidence for Snake Venom Plasticity in a Long-Term Study with Individual Captive Bothrops Atrox. Toxins 2019, 11, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex Cocktails: The Evolutionary Novelty of Venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J. Venomics: Integrative Venom Proteomics and Beyond. Biochem. J. 2017, 474, 611–634. [Google Scholar] [CrossRef]
- Margres, M.J.; Wray, K.P.; Hassinger, A.T.B.; Ward, M.J.; McGivern, J.J.; Moriarty Lemmon, E.; Lemmon, A.R.; Rokyta, D.R. Quantity, Not Quality: Rapid Adaptation in a Polygenic Trait Proceeded Exclusively through Expression Differentiation. Mol. Biol. Evol. 2017, 34, 3099–3110. [Google Scholar] [CrossRef] [Green Version]
- Amazonas, D.R.; Portes-Junior, J.A.; Nishiyama-Jr, M.Y.; Nicolau, C.A.; Chalkidis, H.M.; Mourão, R.H.V.; Grazziotin, F.G.; Rokyta, D.R.; Gibbs, H.L.; Valente, R.H.; et al. Molecular Mechanisms Underlying Intraspecific Variation in Snake Venom. J. Proteom. 2018, 181, 60–72. [Google Scholar] [CrossRef]









| Scales | MCa+ | FCa+ | Cabf | MCp * | FCp * | Cpbm | MHs | FHs |
|---|---|---|---|---|---|---|---|---|
| Dorsal spotting pattern | 24–35 | 26–32 | 27–30 | 30–47 | 30–47 | ND | 32–39 | 31–34 |
| Tail band pattern | 2–4 | 4–6 | 4 | ND | ND | 5 | 4–7 | 3–5 |
| Scales around the rattle | 8–10 | 8–10 | 10 | ND | ND | 12 | 10–12 | 10–12 |
| Subcaudal | 18–24 | 23–28 | 25–26 | 24–29 | 17–25 | 20 | 24–27 | 19–21 |
| Ventral | 141–150 | 136–144 | 143–152 | 161–177 | 167–187 | 165 | 155–158 | 157–164 |
| Supralabial | 11–13 | 10–13 | 11–12 | 12–15 | 12–15 | 14 | 13–14 | 12–14 |
| Infralabial | 11–12 | 10–12 | 11–12 | 11–16 | 11–16 | 14 | 11–14 | 12–14 |
| Interrectal | 20–25 | 20–26 | ND | ND | ND | ND | ND | ND |
| Middle body scales | 21–24 | 21–23 | 23 | 25–28 | 25–28 | 25 | 25–26 | 25 |
| Intersupraocular | 3 | 2–4 | ND | ND | ND | 3 | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roldán-Padrón, O.; Cruz-Pérez, M.S.; Castro-Guillén, J.L.; García-Arredondo, J.A.; Mendiola-Olaya, E.; Saldaña-Gutiérrez, C.; Herrera-Paniagua, P.; Blanco-Labra, A.; García-Gasca, T. Hybridization between Crotalus aquilus and Crotalus polystictus Species: A Comparison of Their Venom Toxicity and Enzymatic Activities. Biology 2022, 11, 661. https://doi.org/10.3390/biology11050661
Roldán-Padrón O, Cruz-Pérez MS, Castro-Guillén JL, García-Arredondo JA, Mendiola-Olaya E, Saldaña-Gutiérrez C, Herrera-Paniagua P, Blanco-Labra A, García-Gasca T. Hybridization between Crotalus aquilus and Crotalus polystictus Species: A Comparison of Their Venom Toxicity and Enzymatic Activities. Biology. 2022; 11(5):661. https://doi.org/10.3390/biology11050661
Chicago/Turabian StyleRoldán-Padrón, Octavio, Martha Sandra Cruz-Pérez, José Luis Castro-Guillén, José Alejandro García-Arredondo, Elizabeth Mendiola-Olaya, Carlos Saldaña-Gutiérrez, Patricia Herrera-Paniagua, Alejandro Blanco-Labra, and Teresa García-Gasca. 2022. "Hybridization between Crotalus aquilus and Crotalus polystictus Species: A Comparison of Their Venom Toxicity and Enzymatic Activities" Biology 11, no. 5: 661. https://doi.org/10.3390/biology11050661