
Quantifying Circulating IgY Antibody Responses against Select Opportunistic Bacterial Pathogens and Correlations with Body Condition Factors in Wild American Alligators, Alligator mississippiensis


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Alligators and Serum Collections
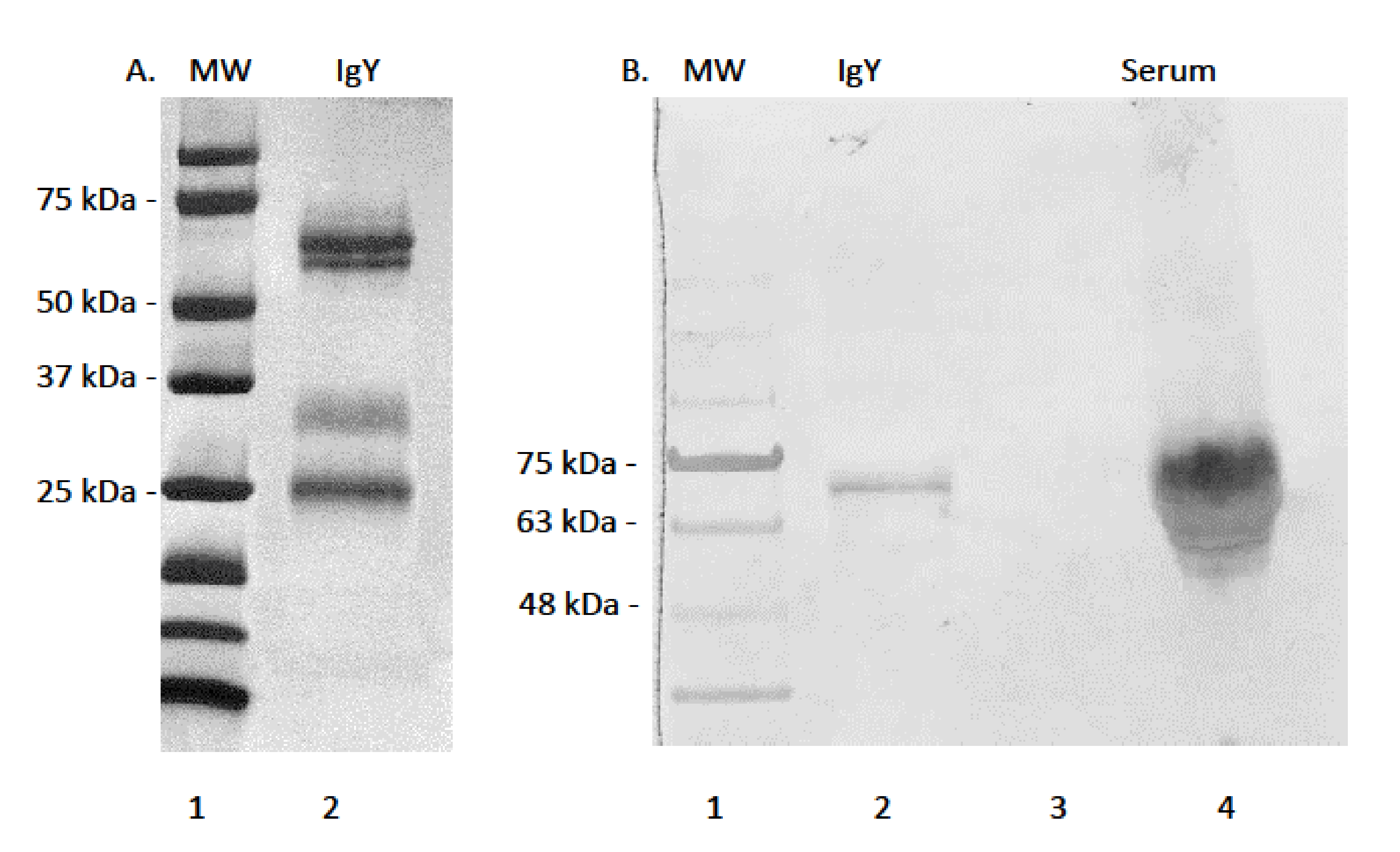
2.2. IgY Purification
2.3. Generation of Monoclonal Antibody against IgY
2.4. Development of ELISAs against Opportunistic Aquatic Bacteria
2.5. Statistical Analysis
3. Results
3.1. Serum Immunoglobulin Y Purification and Reactivity of mAb AMY-9
3.2. Antibody Titers and Condition Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woodward, A.R.; Elsey, R.M. American Alligator Alligator mississippiensis. In Crocodiles: Status Survey and Conservation Action Plan, 3rd ed.; Crocodile Specialist Group: Darwin, Australia, 2019; pp. 1–6. [Google Scholar]
- Chabreck, R.H. The American alligator—Past, present, and future. Proc. Annu. Conf. Southeast. Assoc. Game Fish Comm. 1967, 21, 554–557. [Google Scholar]
- Kahui, V.; Moyle, B.; Brunell, A.M. Alligator conservation and hunting efficiency. Nat. Resour. Model. 2018, 31, e12155. [Google Scholar] [CrossRef]
- Elsey, R.; Woodward, A.; Balaguera-Reina, S.A. Alligator Mississippiensis. The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2019. [Google Scholar]
- Gibbons, J.W.; Scott, D.E.; Ryan, T.J.; Buhlman, K.A.; Tuberville, T.D.; Metts, B.; Greene, J.L.; Mills, T.M.; Leiden, Y.; Poppy, S.M.; et al. The global decline of reptiles, deja’ vu amphibians. BioScience 2020, 50, 653–666. [Google Scholar] [CrossRef] [Green Version]
- Guillette, L.J., Jr.; Woodward, A.R.; Crain, D.A.; Masson, G.R.; Palmer, B.D.; Cox, M.C.; You-Xiang, Q.; Orlando, E.F. The reproductive cycle of the female American alligator (Alligator mississippiensis). Gen. Comp. Endocrinol. 1997, 108, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Mazzotti, F.J.; Best, G.R.; Brandt, L.A.; Cherkiss, M.S.; Jeffery, B.M.; Rice, K.G. Alligators and crocodiles as indicators for restoration of Everglades ecosystems. Ecol. Indic. 2009, 9, S137–S149. [Google Scholar] [CrossRef]
- McCoy, J.A.; Parrott, B.B.; Rainwater, T.R.; Wilkinson, P.M.; Guillette, L. Incubation history prior to the canonical thermosensitive period determines sex in the American alligator. Reproduction 2015, 150, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Rice, C.D. Fish immunotoxicology: Understanding mechanisms of action. In Target Organ Toxicity in Marine and Freshwater Teleosts; Schlenk, D., Benson, W.H., Eds.; Taylor and Francis: London, UK, 2001; pp. 96–138. [Google Scholar]
- Adelman, J.S.; Ardia, D.R.; Schat, K.A. Ecoimmunology. In Avian Immunology, 2nd ed.; Schat, K.A., Kaspers, B., Kaiser, P., Eds.; Academic Press: Boston, MA, USA, 2014; pp. 391–411. [Google Scholar]
- Brock, P.M.; Murdock, C.C.; Martin, L.B. The history of ecoimmunology and its integration with disease ecology. Integr. Comp. Biol. 2014, 54, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, L.; Vogel, L.; Bowden, R. Understanding the vertebrate immune system: Insights from the reptilian perspective. J. Exp. Biol. 2010, 213, 661–671. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, L.M. The reptilian perspective on vertebrate immunity: 10 years of progress. J. Exp. Biol. 2020, 223, jeb214171. [Google Scholar] [CrossRef]
- Mateo, M.R.; Roberts, E.D.; Enright, F.M. Inflammation induced by subcutaneous turpentine inoculation of young American alligators (Alligator mississippiensis). Am. J. Vet. Res. 1984, 45, 1870–1875. [Google Scholar]
- Riera Romo, M.; Pérez-Martínez, D.; Castillo Ferrer, C. Innate immunity in vertebrates: An overview. Immunology 2016, 148, 125–139. [Google Scholar] [CrossRef]
- Stewart, J.R.; Gast, R.J.; Fujioka, R.S.; Solo-Gabriele, H.M.; Meschke, J.S.; Amaral-Zettler, L.A.; del Castillo, E.; Polz, M.F.; Collier, T.K.; Strom, M.S.; et al. The coastal environment and human health: Microbial indicators, pathogens, sentinels and reservoirs. Environ. Health 2008, 7, S3. [Google Scholar] [CrossRef] [Green Version]
- Loge, F.J.; Thompson, D.E.; Call, D.R. PCR detection of specific pathogens in water: A risk-based analysis. Environ. Sci. Technol. 2002, 36, 2754–2759. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Huygens, F.; Goonetilleke, A.; Gardner, T. Real-Time PCR Detection of Pathogenic Microorganisms in Roof-Harvested Rainwater in Southeast Queensland, Australia. Appl. Environ. Microbiol. 2008, 74, 5490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhu, Y.; Wu, X.; Hoffmann, M.R. Rapid Detection Methods for Bacterial Pathogens in Ambient Waters at the Point of Sample Collection: A Brief Review. Clin. Infect. Dis. 2020, 71, S84–S90. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Wu, Q.; Ren, L.; Hu, X.; Guo, Y.; Warr, G.W.; Hammarström, L.; Li, N.; Zhao, Y. Expression of IgM, IgD, and IgY in a Reptile, Anolis carolinensis. J. Immunol. 2009, 183, 3858–3864. [Google Scholar] [CrossRef] [Green Version]
- Delene, K.; Lemma, A.; Fesseha, H. Major diseases of nile crocodile (Crocodylus niloticus) with focus on current status in Arba Minch crocodile ranch, Ethiopia. J. World’s Poult. Res. 2020, 10, 98–110. [Google Scholar] [CrossRef]
- Zemanova, M.A. Towards more compassionate wildlife research through the 3Rs principles: Moving from invasive to non-invasive methods. Wildl. Biol. 2020, 2020, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Myburgh, J.G.; Kirberger, R.M.; Steyl, J.C.; Soley, J.T.; Booyse, D.G.; Huchzermeyer, F.W.; Lowers, R.H.; Guillette, L.J., Jr. The post-occipital spinal venous sinus of the Nile crocodile Crocodylus niloticus: Its anatomy and use for blood sample collection and intravenous infusions. J. S. Afr. Vet. Assoc. 2014, 85, e1–e10. [Google Scholar] [CrossRef] [Green Version]
- Hamlin, H.J.; Lowers, R.H.; Kohno, S.; Mitsui-Watanabe, N.; Amano, H.; Hara, A.; Ohta, Y.; Miyagawa, S.; Iguchi, T.; Guillette, L.J., Jr. The reproductive hormone cycle of adult female American alligators from a barrier island population. Reproduction 2014, 147, 855–863. [Google Scholar] [CrossRef] [Green Version]
- Parrott, B.B.; Bowden, J.A.; Kohno, S.; Cloy-McCoy, J.A.; Hale, M.D.; Bangma, J.T.; Rainwater, T.R.; Wilkinson, P.M.; Kucklick, J.R.; Guillette, L.J., Jr. Influence of tissue, age, and environmental quality on DNA methylation in Alligator mississippiensis. Reproduction 2014, 147, 503–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsen, F.M.; Dorsey, J.E.; Lowers, R.H.; Guillette, L.J., Jr.; Long, S.E.; Bowden, J.A.; Schock, T.B. Evaluating mercury concentrations and body condition in American alligators (Alligator mississippiensis) at Merritt Island National Wildlife Refuge (MINWR), Florida. Sci. Total Environ. 2017, 607–608, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Bangma, J.T.; Bowden, J.A.; Brunell, A.M.; Christie, I.; Finnell, B.; Guillette, M.P.; Jones, M.; Lowers, R.H.; Rainwater, T.R.; Reiner, J.L.; et al. Perfluorinated alkyl acids in plasma of American alligators (Alligator mississippiensis) from Florida and South Carolina. Environ. Toxicol. Chem. 2017, 36, 917–925. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, P.M.; Rainwater, T.R.; Woodward, A.R.; Leone, E.H.; Carter, C. Determinate Growth and Reproductive Lifespan in the American Alligator (Alligator mississippiensis): Evidence from Long-term Recaptures. Copeia 2016, 104, 843–852. [Google Scholar] [CrossRef]
- Work, T.M.; Dagenais, J.; Breeden, R.; Schneemann, A.; Sung, J.; Hew, B.; Balazs, G.H.; Berestecky, J.M. Green Turtles (Chelonia mydas) Have Novel Asymmetrical Antibodies. J. Immunol. 2015, 195, 5452–5460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, M.L.; Rice, C.D. Monoclonal antibodies against loggerhead sea turtle, Caretta caretta, IgY isoforms reveal differential contributions to antibody titers and relatedness among other sea turtles. Dev. Comp. Immunol. 2018, 87, 12–15. [Google Scholar] [CrossRef]
- Rice, C.D.; Schlenk, D.; Ainsworth, J.; Goksøyr, A. Cross-reactivity of monoclonal antibodies against peptide 277–294 of rainbow trout CYP1A1 with hepatic CYP1A among fish. Mar. Environ. Res. 1998, 46, 87–91. [Google Scholar] [CrossRef]
- Margiotta, A.L.; Bain, L.J.; Rice, C.D. Expression of the Major Vault Protein (MVP) and Cellular Vault Particles in Fish. Anat. Rec. 2017, 300, 1981–1992. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, M.L.; Toline, C.A.; Rice, C.D. Humoral Immune Responses to Select Marine Bacteria in Loggerhead Sea Turtles Caretta caretta and Kemp’s Ridley Sea Turtles Lepidochelys kempii from the Southeastern United States. J. Aquat. Anim. Health 2018, 1, 20–30. [Google Scholar] [CrossRef]
- Beck, B.M.; Rice, C.D. Serum antibody levels against select bacterial pathogens in Atlantic bottlenose dolphins, Tursiops truncatus, from Beaufort NC USA and Charleston Harbor, Charleston, SC, USA. Mar. Environ. Res. 2003, 55, 161–179. [Google Scholar] [CrossRef]
- Karsten, A.H.; Rice, C.D. Serum IgM levels against select marine bacteria in the Atlantic sharpnose shark (Rhizoprionodon terraenovae) from three estuaries. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 143, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Waterstrat, P.R.; Brazil, J.; Ainsworth, A.J. Use of an ELISA-based assay for the detection of antibody-secreting cells in channel catfish, Ictalurus punctatus (Rafinesque). J. Fish Dis. 1991, 14, 669–675. [Google Scholar] [CrossRef]
- Warr, G.W.; Magor, K.E.; Higgins, D.A. IgY: Clues to the origins of modern antibodies. Immunol. Today 1995, 16, 392–398. [Google Scholar] [CrossRef]
- Miller, D.L.; Mauel, M.J.; Baldwin, C.; Burtle, G.; Ingram, D.; Hines, M.E., 2nd; Frazier, K.S. West Nile virus in farmed alligators. Emerg. Infect. Dis. 2003, 9, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Huchzermeyer, F.W. Crocodiles: Biology, Husbandry and Diseases; CABI: Wallingford, UK, 2003. [Google Scholar]
- Rossetti, C.; Uhart, M.; Romero, G.; Prado, W. Detection of leptospiral antibodies in caimans from the Argentinian Chaco. Vet. Rec. 2003, 153, 632–633. [Google Scholar] [CrossRef] [PubMed]
- Pérez Flores, J.; Charruau, P.; Cedeño-Vázquez, R.; Atilano, D. Evidence for Wild Crocodiles as a Risk for Human Leptospirosis, Mexico. EcoHealth 2016, 14, 58–68. [Google Scholar] [CrossRef]
- Motes, M.L.; DePaola, A.; Cook, D.W.; Veazey, J.E.; Hunsucker, J.C.; Garthright, W.E.; Blodgett, R.J.; Chirtel, S.J. Influence of water temperature and salinity on Vibrio vulnificus in Northern Gulf and Atlantic Coast oysters (Crassostrea virginica). Appl. Environ. Microbiol. 1998, 64, 1459–1465. [Google Scholar] [CrossRef] [Green Version]
- Froelich, B.A.; Daines, D.A. In hot water: Effects of climate change on Vibrio–human interactions. Environ. Microbiol. 2020, 22, 4101–4111. [Google Scholar] [CrossRef] [Green Version]
- Bock, S.L.; Lowers, R.H.; Rainwater, T.R.; Stolen, E.; Drake, J.M.; Wilkinson, P.M.; Weiss, S.; Back, B.; Guillette, L.; Parrott, B.B. Spatial and temporal variation in nest temperatures forecasts sex ratio skews in a crocodilian with environmental sex determination. Proc. Royal Soc. B Biol. Sci. 2020, 287, 20200210. [Google Scholar] [CrossRef]
- Ogden, L.E. Climate change, pathogens, and people: The challenges of monitoring a moving target. BioScience 2018, 68, 733–739. [Google Scholar] [CrossRef]
- LaVere, A.A.; Hamlin, H.J.; Lowers, R.H.; Parrott, B.B.; Ezenwa, V.O. Associations between testosterone and immune activity in alligators depend on bacteria species and temperature. Funct. Ecol. 2021, 3, 1018–1027. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M. fortuitum | E. rhusiopthiae | V. vulnificus | E. coli |
|---|---|---|---|
| March–July * | January–July * | February–June * | January–June * |
| May–January 2011 * | March–June **** | January–July *** | |
| June–January 2011 ** | March–July **** | January–August ** | |
| July–January 2011 *** | May–June * | January–September * | |
| August–January 2011 * | June–January 2011 * | January–October ** | |
| February–July *** | |||
| February–August * | |||
| February–September * | |||
| February–October ** | |||
| March–June * | |||
| March–July *** | |||
| March–August ** | |||
| March–September * | |||
| March–October ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alston, B.M.; Rainwater, T.R.; Parrott, B.B.; Wilkinson, P.M.; Bowden, J.A.; Rice, C.D. Quantifying Circulating IgY Antibody Responses against Select Opportunistic Bacterial Pathogens and Correlations with Body Condition Factors in Wild American Alligators, Alligator mississippiensis. Biology 2022, 11, 269. https://doi.org/10.3390/biology11020269
Alston BM, Rainwater TR, Parrott BB, Wilkinson PM, Bowden JA, Rice CD. Quantifying Circulating IgY Antibody Responses against Select Opportunistic Bacterial Pathogens and Correlations with Body Condition Factors in Wild American Alligators, Alligator mississippiensis. Biology. 2022; 11(2):269. https://doi.org/10.3390/biology11020269
Chicago/Turabian StyleAlston, Bailey M., Thomas R. Rainwater, Benjamin B. Parrott, Philip M. Wilkinson, John A. Bowden, and Charles D. Rice. 2022. "Quantifying Circulating IgY Antibody Responses against Select Opportunistic Bacterial Pathogens and Correlations with Body Condition Factors in Wild American Alligators, Alligator mississippiensis" Biology 11, no. 2: 269. https://doi.org/10.3390/biology11020269