An Overview on Biogenic Amines in Wine


Consiglio per la ricerca in agricoltura e l’analisi dell’economia agraria-Viticoltura ed Enologia, via P. Micca 35, 14100 Asti, Italy
*
Author to whom correspondence should be addressed.
Beverages 2019, 5(1), 19; https://doi.org/10.3390/beverages5010019
Submission received: 30 December 2018
/
Revised: 30 January 2019
/
Accepted: 12 February 2019
/
Published: 1 March 2019
(This article belongs to the Special Issue Microbiological Safety of Beverages)
Abstract
:Biogenic amines (BAs) are low molecular weight compounds formed from precursor amino acids, mainly by microbial decarboxylation. The presence of these compounds is important in the food and beverage industry because, in high amounts, they can lead to negative effects on consumers. In this review, we illustrate the critical aspects needed to control the formation of BAs during winemaking and their presence in the final product. Recent biotechnological approaches related to microorganisms and their ability to reduce BAs are illustrated. The current methods used for BA detection and quantification are also presented. These methods are very important to consider, as BAs can serve as markers for the quality assessment of products. The information presented here offers an overview useful for identifying specific parameters and conditions which should be controlled to minimise BA content in wine; knowledge about BAs in foods and beverages has been accumulating in recent years, not only to ensure and improve quality (since BAs have been used as an indicator of spoilage) but especially to guarantee consumer safety due to the potential toxic effects of BAs on humans.
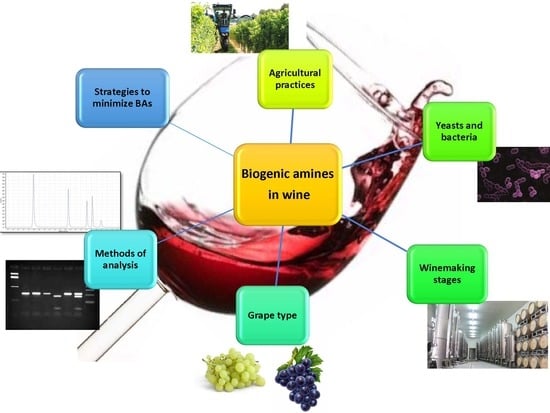
1. Introduction
Biogenic amines (BAs) are commonly found in many foods: high concentrations have been reported in fish, chocolate, cheese, soybean products, sausages and processed meat [1,2,3,4,5,6], as well as in fermented foods and beverages.
Several investigations have reported the presence of BAs in wine, the most important of which are histamine, tyramine and putrescine. BAs can be present in all types of wines. In white wines, the BA content is smaller than in red wines, i.e., 0–10 mg/L for white wines and 0–30 mg/L for red wines [7].
The literature has reported that different countries have established upper limits of histamine in wine: 2 mg/L in Germany, 5mg/L in Finland, 10 mg/L in Australia and Switzerland, 8 mg/L in France, 3.5 mg/L in Netherlands and 6 mg/L in Belgium [8]. However, to our knowledge, no legal limits for BA content in wine have been established.
A person’s sensitivity to BAs depends on his or her capacity for detoxification [9], which is also affected by the presence of ethanol in beverages and/or the assumption of determined medicines. It is difficult to establish maximum limits for these compounds in foodstuffs, because complex interactions occur between BAs and other substances.
Regarding the dose–response relationship, the European Food Safety Authority (EFSA) reported [10] that in healthy volunteers, no symptoms occurred after consuming 25–50 mg of histamine assumed from fishery products or non-alcoholic beverages; however, headache and flushing occurred after consuming 75–300 mg of histamine.
In a study of wine-intolerant subjects, Kanny et al. [11] did not find a relationship between the amine content of tested wines and the frequency and severity of symptoms caused by these wines. This finding suggests that wine intolerance and the generation of adverse reactions to wines may be due to the participation of a substance other than histamine or BAs.
Histamine has been detected in wines from different countries for example, in Portugal, 23.1 mg/L [12]; in Italy, 10.8 mg/L [13]; and in France, 14.05 mg/L [14]. Comuzzo et al. [15] also found 11.1 mg/L, 14.8 mg/L and 12.1 mg/L of histamine in Spanish, German and Austrian wines, respectively. Also, putrescine was found in wines at different concentrations in three surveys in Italy (31.8, 11.13 and 16.2 mg/L) [13,15,16] and in one survey in France (48.72 mg/L) [14].
Glória et al. [17] observed that in Cabernet Sauvignon wines from Oregon, USA, putrescine was the prevalent amine (63.5%), followed by histamine (16.8%) and spermidine (9.8%). The prevalence of these amines was also observed in Rioja wines [18] and red wines from Tarragona [19]. Bach et al. [14] reported concentrations of ethylamine and methylamine up to 10.46 mg/L and 36.64 mg/L, respectively, in French wines.
BA amounts depend on several factors. On the one hand, winemaking conditions, vinification techniques, ageing, agricultural practices and climatic conditions can affect BA amounts [20]; on the other, BA amounts are also affected by the species of microorganisms responsible for their production in wine with decarboxylase enzymes, the abundance of amino acid precursors in the medium and wine parameters, such as pH, alcohol and sulphur dioxide, that influence bacterial growth [8,21].
The presence of these compounds represents a critical aspect in oenology; recently, BA content has been considered as an important indicator of the quality and safety of wine [13]. In addition, some authors have reported regional differences in the quantities of BAs in wine [22].
The aim of this review is to summarise the state of the art regarding the main aspects involved in the presence of BAs in wine. A good knowledge of these aspects would be helpful for actively promoting the production of wine without BAs by correctly managing practices and processing methods.
2. Biogenic Amines and Microorganisms
In the winemaking process, both yeasts and bacteria may participate in amine production. There is general agreement that yeasts make a less significant contribution than lactic acid bacteria (LAB) to the final BA content in wine, a finding which has been supported by the greater quantity of data regarding the biochemistry, genetics and regulation of amine production by LAB compared with the data available for yeasts.
Few studies have been conducted on the formation of BAs by yeasts, and most of them only compared different yeast species and only quantified histamine [23].
According to Caruso et al. [24], Saccharomyces cerevisiae can produce significant amounts of ethanolamine and agmatine; these authors also tested non-Saccharomyces yeasts: Kloeckera apiculata, Candida stellata, Metschnikowia pulcherrima, Brettanomyces bruxellensis and the fungus Botrytis cinerea. They found that the highest concentration of total BAs was formed by B. bruxellensis, with an average value of 15 mg/L, followed by S. cerevisiae, with an average value of 12.14 mg/L. The other species formed less than 10 mg/L of total BAs. Two amines seemed to be species-specific, phenylethylamine and ethanolamine, and were produced in more considerable amounts by B. bruxellensis and S. cerevisiae, respectively. The same results were confirmed by Granchi et al. [25].
Torrea and Ancín-Azpilicueta [26] found slight BA production by S. cerevisiae depending on the strain, but the concentrations were very low. Tristezza et al. [27] demonstrated the ability of yeast species of oenological provenience to produce histamine during grape must fermentation. In particular, they found an isolate of Issatchenkia terricola, one strain of M. pulcherrima and two isolates of Pichia manshurica that were able to synthesise histamine.
Contrary to these authors, Landete et al. [28] did not find BA production in any of the 36 strains of different wine yeast genera screened: Aureobasidum, Candida, Hanseniaspora, Hansenula, Kloeckera, Metschnikowia, Pichia or strains of the species S. cerevisiae.
Marcobal et al. [29] demonstrated that some amines can be produced in grape or in grape must as putrescine, cadaverine or phenylethylamine, or they can be formed by yeasts during alcoholic fermentation (AF), as ethylamine and phenylethylamine, although only very low concentrations were reached during these early stages.
These results lead, therefore, to the conclusion that yeasts do not appear to be the main cause of most of the amines found in wines.
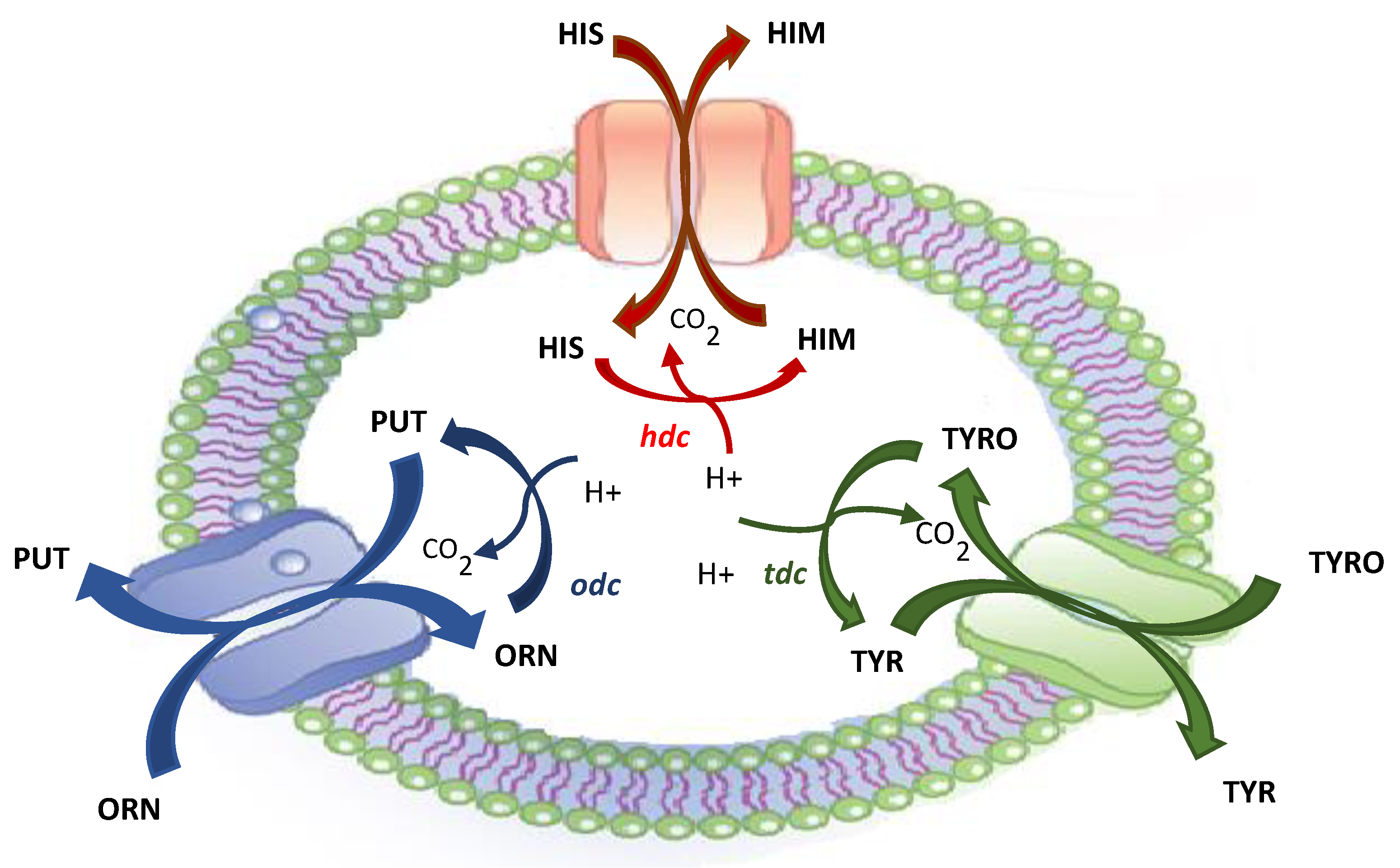
Usually, BA production results from the presence of bacteria that are capable of decarboxylating amino acids [30,31] (Figure 1); in particular, histamine is formed from histidine by histidine decarboxylase (hdc); ornithine decarboxylase (odc) catalyses the decarboxylation of ornithine to putrescine; and tyrosine is the precursor of tyramine produced by tyrosine decarboxylase (tdc).
Decarboxylase genes are contained in cassettes constituted by four genes, one of which encodes for a transmembrane amino acid/amine antiporter; this has been demonstrated for the tyrosine/tyramine antiporter by Lucas et al. [32] in Lactobacillus brevis; for the ornithine/putrescine antiporter by Romano et al. [33] in L. brevis; and for the histidine/histamine antiporter in Lactobacillus buchneri by Molenaar et al. [34] and Cruz Martin et al. [35].
This operon has particular importance since it generates a proton motive force and regulates cellular pH, thereby contributing to energy production; therefore, the production of BAs can be considered a defence mechanism used by bacteria to better survive in an acidic medium [36].
Amino acid decarboxylase activity depends on several factors: pH, temperature, sugars and amino acid precursors. Pessione et al. [37] reported that in LAB, pH can regulate decarboxylase enzymes by catalytic activation and biosynthetic overexpression. In a different work, it was demonstrated that decarboxylase biosynthesis is maximal during the stationary phase when the pH is low due to lactic acid accumulation [38]; this aspect is consistent with the hypothesis that BAs constitute a protective mechanism against a low pH. For histidine decarboxylase (HDC), the optimum pH is 4.8 [39]; for tyrosine decarboxylase (TDC), the optimum pH is 5.0 [40]; while for ornithine decarboxylase, (ODC), the optimum pH is 5.8 [41].
Temperature is also an important factor. Moderate temperatures increase enzyme activity, as shown by Marcobal et al. [42], who found that in Enterococcus faecium and L. brevis, the optimum temperature for tyramine production under aerobic conditions was 32 °C; whereas under anaerobic conditions, the maximum tyramine production was obtained at 22.0–24.5 °C.
The sugar concentration of the medium can influence decarboxylase activity; in particular, the lack of sugars has often been associated with higher BA production. This association can be explained by the fact that the decarboxylative pathway transport system provides metabolic energy [43]. Landete et al. [44] observed that in the oenological LAB (Lactobacillus hilgardii, Pediococcus parvulus and Oenococcus oeni), an increasing concentration of fructose and glucose progressively inhibited histamine formation; moreover, hdc gene expression was also reduced by malic acid and citric acid, whereas ethanol enhanced HDC enzyme activity.
Finally, HDC and ODC biosynthesis were shown to be closely dependent on the presence of high concentrations of free amino acids in the growth medium [38].
Among LAB, O. oeni is the main species present in wine and is the best adapted one for carrying out malolactic fermentation (MLF) to the stressful conditions of wine [45]. If BA formation is associated with MLF, then O. oeni would be expected to be mainly responsible.
Some authors have found that O. oeni significantly contributes to the overall content of histamine in wines, but that the ability of the species to produce this amine varies among strains [46,47]. Contrarily, Costantini et al. [48] and Moreno Arribas et al. [30] did not find O. oeni to be capable of producing histamine, a finding also confirmed by Garcia-Moruno and Munoz [49].
Marcobal et al. [50] isolated and identified a strain of O. oeni that produces putrescine, and they further studied this ability in another 42 strains of this species by molecular methods (odc gene detection). Additionally, they found that the odc gene was not present in any of the strains.
Regarding Lactobacillus, different strains have been found to be capable of producing BAs: Lactobacillus hilgardii, Lactobacillus buchneri, Lactobacillus rossiae, Lactobacillus sakei and Lactobacillus mali have all been found capable of producing histamine [48,51,52,53]
Tyramine-producing LAB in wine that has undergone MLF were identified and isolated by Moreno-Arribas et al. [54], all of which belong to the Lactobacillus genera; in particular, they found L. brevis and one strain of L. hilgardii. Costantini et al. [48] also found that L. brevis was able to produce tyramine. This finding was confirmed by Sebastian et al. [55], who tested 57 strains of L. brevis and observed the formation of BAs in all of them. Lastly, the ability of wine Lactobacillus plantarum strains to form tyramine was analysed by Arena et al. [56]. They found one L. plantarum strain harbouring the tdc gene, but tyramine production was occasional because tyrosine decarboxylase activity was negatively affected by sugars, such as glucose and fructose, and by L-malic acid. As the literature suggests, only one strain (O. oeni DSM 2025) was shown capable of producing tyramine in a defined growth medium [57].
From this description, we could conclude that the ability to produce BAs is mostly distributed among species that usually do not perform MLF, which is mainly carried out by O. oeni. These species are contaminants that can spoil wine and are therefore indicative of poor winemaking and bad sanitisation practices.
3. Biogenic Amines and the Type of Grape/Wine and Winemaking Stages
Cultivar-related differences in BA content have been observed in many studies of wines worldwide—for instance, in Italian, Spanish, Greek, Chinese and Chilean grapes and wines—and are summarised in Table 1.
The sanitary state of the grape also influences the amount of BAs, as demonstrated by Cecchini et al. [63] and in agreement with Leitao et al. [64], who associated putrescine and cadaverine in musts with poor sanitary conditions of grapes.
Moreover, during winemaking, AF and MLF may influence BA accumulation.
Del Prete et al. [58] conducted a study on different grape varieties—Merlot, Syrah, Sangiovese, Cesanese d’Affile, Carmenere, Montepulciano and Cabernet Franc—and monitored BA content during the different stages of vinification. The cultivars were grown in the same pedoclimatic conditions and with the same training system in order to reduce variability. In Table 2, the results obtained from analysing BAs in different winemaking phases are shown. Ethanolamine, ethylamine and putrescine were present in the grapes, while tyramine, cadaverine and agmatine were not found. These findings are in agreement with those of other authors: Ough et al. [65] found ethanolamine and ethylamine in grapes, while Broquedis et al. [66] detected putrescine in Cabernet Sauvignon and Ugni blanc.
During AF [58], ethanolamine and tyramine content increased, while the quantity of ethylamine and putrescine decreased. During MLF, agmatine increased, while tyramine decreased to nearly zero. The authors [58] explained that these differences were due to the normal metabolic processes of yeast and bacteria: ethanolamine is an intermediate in phospholipid synthesis that is released in wine by S. cerevisiae; agmatine and tyramine may likely be formed as a consequence of the hydrolysis of hydroxycinnamic amide compounds in grapes by the action of yeast and bacteria.
Generally, MLF is considered one of the most crucial factors for BA production: several authors have presented evidence that, in winemaking, BAs are mainly formed during the MLF phase [29,60,67,68].
Landete et al. [60] reported that MLF was the main source of tyramine, histamine and putrescine for red wines, while cadaverine and tryptamine concentrations were not affected. Conversely, Bauza et al. [69] observed an increase in putrescine levels during fermentation from must to AF to MLF, in agreement with Soufleros et al. [70].
About this matter, the data available in the literature are contradictory, since Soufleros et al. [70] found a significant correlation between the levels of tyramine and histamine during fermentation processes, while Herbert et al. [68] reported no significant changes in the levels of histamine during either alcoholic or malolactic fermentative processes, yet both putrescine and tyramine increased in red wines immediately after MLF. On the other hand, Del Prete et al. [58] reported that during MLF, tyramine disappeared, whereas the concentration of putrescine remained nearly unchanged; in addition, the level of agmatine increased during MLF.
Garcia-Marino et al. [71] observed that in high-quality wines, the greatest increase in BAs corresponded to AF—no new amines were observed after MLF. In contrast, in organic wines, MLF increased BA levels, particularly histamine, tyramine, tryptamine and phenylethylamine. Ethanolamine and putrescine had a different evolution: They reached their highest levels in both high-quality wines and organic wines; after AF and during MLF, putrescine further increased in organic wines.
During wine ageing and storage, concentrations of BAs have been reported to increase [60,72]. In the case of histamine, a consistent increase was noticed only several months after the end of MLF. Conversely, putrescine and tyramine concentrations increased immediately after MLF had finished [68].
Henriquez-Aedo et al. [73] analysed BAs in five different wineries during Cabernet Sauvignon vinification, as shown in Table 3. They observed that only one winery (B) showed the highest formation of BAs during MLF, whereas the others showed the highest formation during AF and in bottled wines. Therefore, it seems that the critical point for BA formation is bottled wine, which could be related to a lack of sanitation during bottling or to an insufficient SO2 concentration to microbiologically stabilise the wine.
A similar study was conducted on Portuguese wines by Herbert et al. [68]. They showed that histamine, tyramine, tryptamine, phenylethylamine, putrescine and cadaverine did not vary during AF—particularly histamine, whose levels were close to the values found in the musts (average 1.2 mg/L for red musts). Nevertheless, samples taken about 18 months after the end of MLF in red wines presented higher levels of histamine (11.1 mg/L for red and 12.5 mg/L for white wines) than in the corresponding musts. Therefore, they identified the storage period as the critical stage.
Ordonez et al. [74] studied the effect of storage conditions on the BA profile in open bottles. They did not find significant changes in BA content, which remained similar to the initial concentration. However, after Principal component analysis (PCA), the authors observed that BA content could be useful for separating the different types of wines, particularly high-quality wines from standard wines and young white wines: Sandard wines were placed in the PC quadrant dominated by putrescine, while high-quality wines were placed in the quadrant described by agmatine and phenylethylamine.
These data lead to the conclusion that, in order to reduce BA levels, it is necessary to control every stage of the vinification. By identifying the critical point of BA formation, it is possible to carry out corrective actions to reduce or prevent their presence, developing or modifying some oenological and/or technological practices.
4. Agricultural and Oenological Practices
BA content can also be influenced by agricultural and oenological factors.
Regarding agricultural practices, Smit et al. [75] demonstrated that the use of nitrogen fertilisation can increase BAs in musts and wines. In particular, the total amine content increased significantly in musts; whereas in wines, the fertilisation effect had a lower impact on amine content with respect to the activity of microorganisms.
Other factors which can influence BA content are the degrees of vintage and maturity of grapes as well as irrigation. Martínez-Pinilla et al. [76] showed that in red wines from cv Monastel, Tempranillo and Maturana Tinta de Navarrete, the BA content varied between vintages. They observed that wines of vintage 2009 showed more total BA content than wines of vintage 2010, leading to a change in their amine profile; these results agree with those of other studies [58,77]. The authors explained that differences in amine content may be due to the diversity of wine microorganisms, which are naturally differently selected each year, probably due to climatic conditions.
Ortega-Heras et al. [20] observed that the amino acid content in grapes was affected by climatic conditions and vintages. Amino acid content also depended on the degree of maturity; in fact, in grapes or musts, the content increased from the veraison to the harvest. During ripening, a deceleration of berry growth and reduced protein synthesis occurs, and this may explain the accumulation of free amino acids [78]. The same authors observed that irrigation did not affect the evolution of nitrogen compounds during the AF process, but the degree of maturity in some of the amino acids tested was affected. No direct relationship could be established between irrigation or maturity degree and BAs.
Conflicting results regarding the correlation between grape skin maceration practices and the BA levels in wines have been reported in the literature. Soleas et al. [79] affirmed that skin contact time and BA concentration are not correlated. However, other authors have stated that the duration of skin maceration can affect the BA content in wine: a long skin contact time can potentially increase the production of BAs [80,81]. With regard to the influence of commercial enzymes on BAs, Martín-Álvarez et al. [81] concluded that the addition of pectolytic enzymes to grapes did not promote BA accumulation in their wine.
Pogorzelski [82], Ancín-Azpilicueta et al. [83] and Smit et al. [75] observed that in Cabernet Sauvignon and Syrah, the absence of skin contact resulted in a higher content of BAs; this can be explained by the absence of a phenolic compound, which can affect BA content. Galgano et al. [84] reported that phenolic compounds seem to be a natural mechanism for reducing putrescine formation because they can protect the cell against oxidative stress.
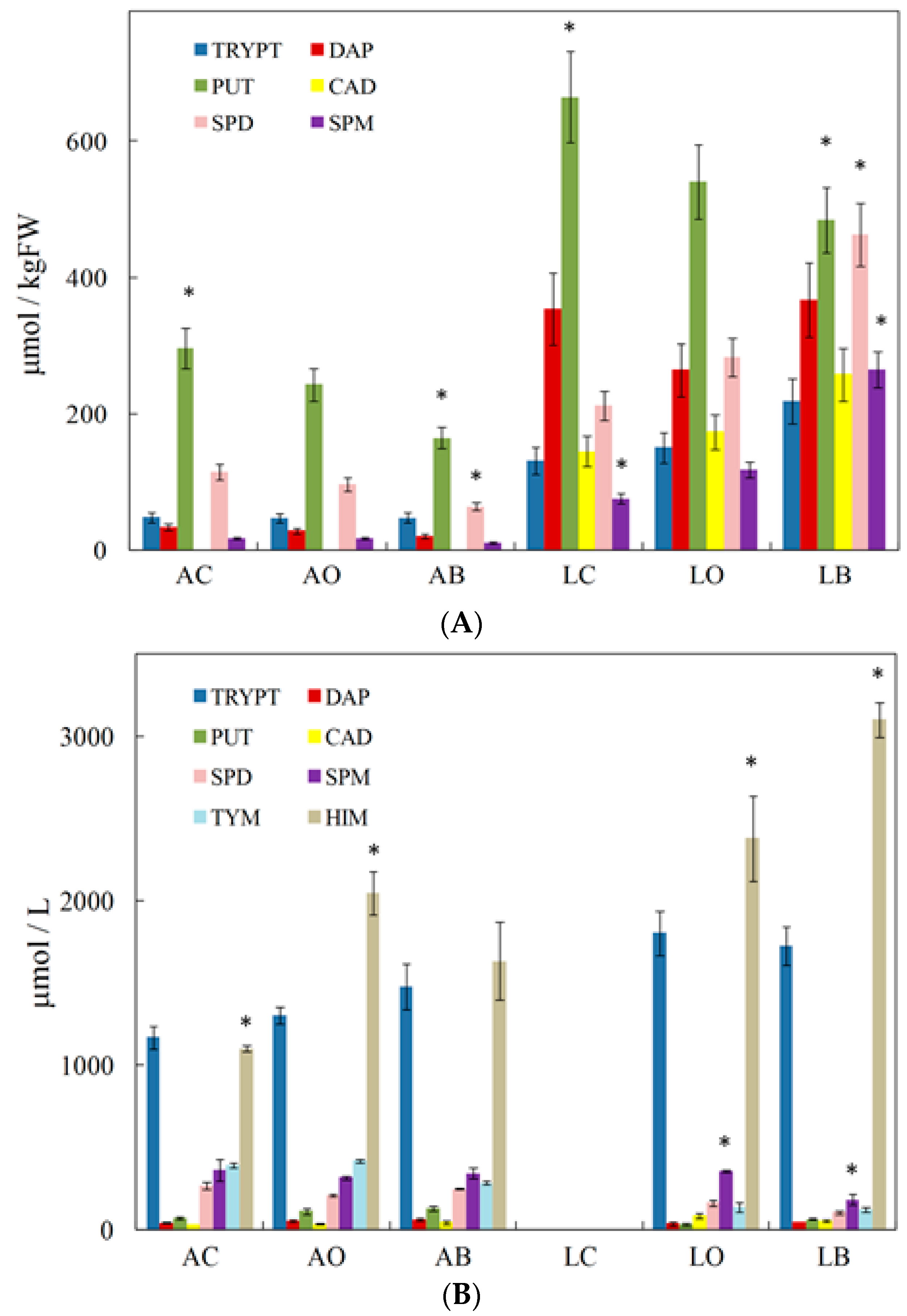
Finally, some studies have focused on the effect of organic or conventional agriculture on the BA content of wines. Tassoni et al. [85] analysed BA content in Lambrusco (red wine) and Albana (white wine), and they compared conventional, organic and biodynamic agricultural and oenological practices. In Figure 2A, the BA levels determined by HPLC are shown. In all the samples, putrescine was the most abundant polyamine, but its content was lower in biodynamic samples than in conventional samples. Cadaverine was totally absent in the Albana samples, while tryptamine was present in both grapes. Spermidine and spermine were higher in the biodynamic samples. In general, the total amount of BAs was on average 4.2-fold higher in Lambrusco than in Albana (Figure 2A). Figure 2B shows the BA levels found in wines. Samples AO and LB contained the highest BA amounts, respectively, for white and red wines. Histamine and tryptamine were the most abundant amines in both white and red wines, with average quantities 1.7- and 1.3-fold higher in red than in white wines. On the contrary, tyramine was on average 2.6-times more abundant in Albana than in Lambrusco wines. In Lambrusco, spermidine was present in organic and biodynamic samples, but it was absent in conventional samples; in Albana, this amine was present in the same amount in all the samples (Figure 2B).
Tassoni et al. [86] conducted the same study on Pignoletto and Sangiovese wines and observed a major concentration of BAs in Italian Pignoletto white wines compared to Sangiovese red wines; the authors hypothesised that the higher levels of BAs in Pignoletto may be due to the presence of higher levels of amino acids. Their data seem to indicate no significant differences between conventional, organic and biodynamic practices.
5. Detection and Quantification of BAs
5.1. Enzymatic Methods
Enzymatic methods for the quantification of histamine were first developed for fish; subsequently, they have been introduced for wine.
Marcobal et al. [87] applied the ELISA (enzyme-linked immunosorbent assay) method for histamine determination in wines. The proposed method was validated by comparison with HPLC, and a good correlation with HPLC analysis was found. The advantages of using ELISA include its simplicity, rapidity and low cost, and it can therefore be proposed as a tool for histamine determination in wines instead of HPLC.
There is also an enzymatic test used to quantify histamine in foods and beverages. It is based on the reaction of histamine-dehydrogenase, which catalyses the oxidation of histamine to imidacetaldehyde in the presence of an electron carrier and a dye. The formation of the dye is measured at 450 nm by a microplate reader and is proportional to the histamine concentration.
The presence of histamine in samples, including wine, can be accomplished using specific strips; this technology allows the visual detection of histamine in only four minutes. The strips contain an enzyme that reacts specifically with histamine and reduces a dye indicator, producing a colour change for easy visual interpretation of the results. When the strips are dipped into samples containing histamine, the pad rapidly changes from white to a colour designating the histamine concentration present in the food or drink. This technique does not require laboratory equipment and can thus be used anywhere.
5.2. Chromatographic Methods
Basic information on BAs in foods is very important for protecting consumer health and meeting industry standards. Therefore, it is necessary to have sensitive and effective methods for BA quantification. The most popular analytical technique applied to quantify amines is HPLC with C18 reverse-phase (RP) columns. The determination of BAs is a bit difficult due in part to the physicochemical properties of these compounds. Numerous analytical methods have been proposed, and most analyses have been performed with a previous derivatization of amines, although some researchers have also determined amines without derivatization.
Typically, wine samples are analysed either directly or after a simple treatment with polyvinylpolypyrolidone (PVPP) to remove some phenolic compounds [62,88]. BAs do not have enough absorption in the UV-Vis or FLD (fluorescence detector) wavelength ranges. Therefore, a pre-column or post-column derivatization is often needed for their detection. The derivatization step improves separation in the RP columns by decreasing the polarity of the original compounds [62,88,89,90].
Different derivatization reagents can be used for HPLC analysis; the main goal is to quickly obtain amino derivatives that do not require a high reaction temperature and yield derivatives that remain stable over time. The most commonly used derivatizing reagents are dansyl chloride (DNS-Cl) and o-phthalaldeyde (OPA). The determination of dansylated amines in wines has been the target of several studies [91,92]. Both DAD (diode array detector) and FLD can be used, but FLD has demonstrated better sensitivity for detecting dansylated amines [62,88]. For the quantification of BAs in wine, DNS-Cl has been largely used because it produces stable derivatized compounds. The disadvantage of using this reagent is the time required for the reaction together with the application of a high external temperature: 10 to 60 min at 40 to 70 °C [92]. Jiang et al. [93] improved the dansylation reaction by using ionic liquids as media for the derivatization at room temperature and reducing the time to around 20 min.
OPA is also often employed in HPLC analysis. OPA derivatives are less stable, but the reaction can occur at room temperature in a short amount of time [94,95]. Pereira et al. [90] proposed a pre-column derivatization with OPA without using any preliminary separation or clean-up; derivatization was performed in three minutes into the sample injection loop of the HPLC and allowed the determination of both BAs and amino acids in wine using an FLD detector.
Wang et al. [96] used diethyl ethoxymethylenemalonate (DEEM) as a pre-column derivatization method for the quantification of 10 BAs and 23 amino acids, with RP-HPLC-DAD: DEEM shortened the analysis time to 30 min. This pre-column derivatizing compound enables the quantification of secondary and primary amines, and its derivatives are stable at room temperature for several days and can be detected by a UV detector. In the official OIV (Organization International de la Vigne et du Vin) method, DEEM is also proposed as a derivatizing reagent.
The labelling agent 1,2-naphthoquinone-4-sulfonate (NQS) has also been described in the literature. Its derivatization is conducted, in general, in a basic medium (pH 8.5–10.5) at temperatures from 50 to 80 °C with reaction times from 2 to 20 min. NQS has been used previously for the pre- and post-column derivatization of amino acids [97,98,99] in HPLC and in capillary electrophoresis (CE) methods.
Recently, Jastrzebska et al. [100] employed a new derivatization reagent, namely 1-fluoro-2-nitro-4-(trifluoromethyl)benzene (FNBT), for the determination of histamine, tyramine, tryptamine and phenylethylamine in wines using RP-HPLC-DAD analysis. This compound was shown to be simple and less time-consuming when compared to other reagents.
Other than HPLC, other methods can be used to analyse BAs. Thin Layer Chromatography (TLC) is an economical method employed for BA semi-quantification, but it requires time for analysis. Samples must be derivatized with dansyl chloride, and amines must be visualised under a UV lamp. TLC was described by Garcia-Moruno et al. [101] and employed by Costantini et al. [48] and Sebastian et al. [55] to detect histamine, putrescine and tyramine produced by LAB. A TLC method coupled to a densitometer to quantify dansylated BAs was also developed [102], and it can be used for routine analysis of histamine, tyramine, putrescine and cadaverine in wine.
5.3. Capillary Electrophoresis
Capillary electrophoresis (CE) is an alternative separation technique for the analysis of BAs that offers some advantages: It is simple, has a high resolving power, and allows for the screening of a large number of samples with a small sample requirement [103]. However, its LOD (limit of detection) is higher when compared with HPLC and ion chromatography, mainly because of lower amounts of the injected sample and a shorter optical path length. Ginterova et al. [104] reported that the reproducibility of CE is usually not as high as that of HPLC. For this reason, its suitability depends on the expected level of BAs in the samples. However, Daniel et al. [105] reached an LOD of 1–2 µg/L, and Uzasci et al. [106] used non-ionic micellar electrokinetic chromatography (a specific type of CE) to obtain an LOD of 0.06–0.11 µg/L.
5.4. Biosensors
In recent years, the development of biosensors has attracted much interest in the scientific community, because these low-cost devices can give results in a few minutes without the need for any kind of sample pre-treatment and with the option to be used outside the laboratory [107]. Biosensors for BAs comprise various combinations of different enzymes for selective biorecognition and signal transduction systems and are based on different signal mechanisms. Several works have focused on the development and application of biosensors. In 2010, Alonso-Lomillo et al. [108] reported, for the first time, monoamine oxidase (MAO)/horseradish peroxidase (HRP) and diamine oxidase (DAO)/horseradish peroxidase (HRP)-based biosensors using screen-printed carbon electrodes for the determination of BAs, which were successfully applied for the quantification of BAs in fish. A DAO-based biosensor was applied by Di Fusco et al. [109]. In 2016, Henao-Escobar et al. [110] described a biosensor for the simultaneous detection of histamine and putrescine: This system consisted of two working electrodes connected in array mode. Histamine dehydrogenase and putrescine oxidase enzymes were respectively immobilised by crosslinking on each working screen-printed electrode and were simultaneously determined by measuring the oxidation current with an amperometer.
The fast response, minimal sample treatment and high sensitivity of the apparatus indicate that biosensors may conveniently be employed within production plants or wineries to provide a reliable estimate of the overall BA content—a parameter that is increasingly being required by food quality commissions worldwide [109].
5.5. Molecular Methods
Molecular methods are also employed to detect BAs that produce bacteria. The PCR technique, using specific primers, can detect the presence of the hdc, odc and tdc genes that code for the decarboxylase enzymes [46,48,50,111,112].
Landete et al. [28], in their review, listed all the primers described in the literature for hdc, tdc and odc, which are reported here in Table 4. Assays can be conducted as single or multiplex reactions. The multiplex PCR assay can be successfully used for the routine detection of strains that are potential producers of histamine, tyramine, putrescine and cadaverine in foods. The simultaneous amplification of all the genes targeting amines in the same PCR reaction reduces both reagent quantities and labour costs. A multiplex PCR assay for the detection of histamine-, tyramine- and putrescine-producing LAB was developed by Coton and Coton, Marcobal et al., de Las Rivas et al. and Costantini et al. [51,113,114,115].
Finally, qPCR protocols have been set up for the quantification of hdc, odc and tdc genes and consequently for the enumeration of bacteria producing histamine, tyramine and putrescine in wine [52,116].
The ability of microorganisms to decarboxylate amino acids is highly variable. It depends not only on the species but also on the strain and on environmental conditions. PCR methods can be useful for detecting BA-producing bacteria early, but the presence of the decarboxylase gene is not always related to effective production. Therefore, in our opinion, both molecular and chemical methods should be applied for amine detection to obtain complete information from each wine sample.
6. BA Degradation
Present knowledge indicates that the control of microbiota is a good strategy for reducing BA production. Consequently, the use of selected non-BA-producing starter cultures can be useful for reducing the growth of contaminating microorganisms [81]. Reduction can also be enhanced by using bacteria capable of degrading BAs.
A current topic concerning BAs is the use of BA-degrading microorganisms [121]. García-Ruiz et al. [122] examined the ability of 85 LAB strains isolated from wines and other related oenological sources to degrade histamine, tyramine and putrescine; they found that the greatest BA-degrading ability was exhibited by nine strains belonging to the Lactobacillus and Pediococcus groups, the best of which was L. casei IFI-CA 52.
Two strains of L. plantarum were found by Capozzi et al. [123] to degrade putrescine and tyramine. One interesting aspect of their research was that L. plantarum survived in stressful wine conditions and improved wine aroma compounds, which are useful factors for competing with spoilage LAB. These strains show promising technological properties, suggesting that the ability to degrade BAs could also serve as a criterion for selecting a new generation of starter cultures [123]. Recently, Callejon et al. [124] demonstrated that two enzymes isolated and purified from the L. plantarum J16 and P. acidilactici CECT 5930 strains, and identified as multicopper oxidases, were able to degrade histamine, tyramine and putrescine, relevant BAs in wine [125].
Some yeasts have also been found capable of degrading BAs. Recently, Bäumlisberger et al. [126] observed that some strains of Debaryomyces hansenii and Yarrowia lipolytica were also able to reduce such compounds. The most effective strain, D. hanseniii H525, was able to metabolise a broad spectrum of BAs. This property is likely due to peroxisomal amine oxidase activity.
Such a finding introduces a new perspective on the possibility of employing microorganisms or purified microbial enzymes to deal with the problem of high amine concentrations in wine [124], and further research should be performed to find new strains capable of degrading BA.
7. Conclusions
In conclusion, the uncontrolled growth of indigenous microorganisms, especially bacteria with decarboxylase capacity, is among the main causes of the increase of amines in wine, together with the high quantity of amino acid precursors.
Although BAs are generally present in wine in low concentrations, it should be noted that, in the daily diet, their negative effects on health could be enhanced by their simultaneous presence in many other fermented foods, such as cheese, fish and meat, and in the presence of ethanol.
It is therefore essential to reduce the risks of BA production in the wine industry by applying appropriate procedures in the cellar. From a microbiological point of view, it is important to correctly manage wine fermentation to prevent the growth of contaminating microorganisms. A reduction in BA formation might also be achievable by optimising winemaking parameters and manufacturing practices to prevent wine contamination and the growth of BA-producer microorganisms.
Author Contributions
All authors contributed to the writing of the manuscript and read and approved the final version.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Him | Histamine |
| Tyr | Tyramine |
| Put | Putrescine |
| Cad | Cadaverine |
| Met | Methylamine |
| Agm | Agmatine |
| Phe | Phenylethylamine |
| Spm | Spermine |
| Spd | Spermidine |
| Try | Tryptamine |
| Ety | Ethylamine |
| Eth | Ethanolamine |
| Tea | Triethylamine |
| Tma | Trimethylamine |
| Dap | Diamine-propane |
| Ism | Isoamylamine |
References
- Al Bulushi, I.; Poole, S.; Deeth, H.C.; Dykes, G.A. Biogenic amines in fish: Roles in intoxication, spoilage, and nitrosamine formation—A review. Crit. Rev. Food Sci. Nutr. 2009, 49, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Pastore, P.; Favaro, G.; Badocco, D.; Tapparo, A.; Cavalli, S.; Saccani, G. Determination of biogenic amines in chocolate by ion chromatographic separation and pulsed integrated amperometric detection with implemented wave-form at Au disposable electrode. J. Chromatogr. A 2005, 1098, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Innocente, N.; D’Agostin, P. Formation of biogenic amines in a typical semihard Italian cheese. J. Food Prot. 2002, 65, 1498–1501. [Google Scholar] [CrossRef] [PubMed]
- Toro-Funes, N.; Bosch-Fuste, J.; Latorre-Moratalla, M.L.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Biologically active amines in fermented and non-fermented commercial soybean products from the Spanish market. Food Chem. 2015, 173, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Suzzi, G.; Gardini, F. Biogenic amines in dry fermented sausages: A review. Int. J. Food Microbiol. 2003, 88, 41–54. [Google Scholar] [CrossRef]
- Favaro, G.; Pastore, P.; Saccani, G.; Cavalli, S. Determination of biogenic amines in fresh and processed meat by ion chromatography and integrated pulsed amperometric detection on Au electrode. Food Chem. 2007, 105, 1652–1658. [Google Scholar] [CrossRef]
- Maintz, L.; Novak, N. Histamine and histamine intolerance. Am. J. Clin. Nutr. 2007, 85, 1185–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, A.Y.Y.; Du Toit, W.J.J.; Du Toit, M. Biogenic amines in wine: Understanding the headache. S. Afr. J. Enol. Vitic. 2008, 29, 109–127. [Google Scholar] [CrossRef]
- Bauza, T.; Blaise, A.; Teissedre, P.; Cabanis, J.; Kanny, G.; Moneret-Vautrin, D.A.; Daumas, F. Les amines biogènes du vin. Métabolisme et toxicité. Bulletin de l’O.I.V. 1995, 68, 42–67. [Google Scholar]
- EFSA Panel on Biological Hazards. Scientific opinion on risk based control of biogenic amine formation in fermented foods. EFSA J. 2011, 9, 2393. [Google Scholar] [CrossRef]
- Kanny, G.; Gerbaux, V.; Olszewski, A.; Frémont, S.; Empereur, F.; Nabet, F.; Cabanis, J.-C.C.; Moneret-Vautrin, D.-A.A. No correlation between wine intolerance and histamine content of wine. J. Allergy Clin. Immunol. 2001, 107, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Ramos, R.M.; Valente, I.M.; Rodrigues, J.A. Analysis of biogenic amines in wines by salting-out assisted liquid-liquid extraction and high-performance liquid chromatography with fluorimetric detection. Talanta 2014, 124, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Martuscelli, M.; Arfelli, G.; Manetta, A.C.; Suzzi, G. Biogenic amines content as a measure of the quality of wines of Abruzzo (Italy). Food Chem. 2013, 140, 590–597. [Google Scholar] [CrossRef] [PubMed]
- Bach, B.; Le Quere, S.; Vuchot, P.; Grinbaum, M.; Barnavon, L. Validation of a method for the analysis of biogenic amines: Histamine instability during wine sample storage. Anal. Chim. Acta 2012, 732, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Comuzzo, P.; Rauhut, D.; Werner, M.; Lagazio, C.; Zironi, R. A survey on wines from organic viticulture from different European countries. Food Control 2013, 34, 274–282. [Google Scholar] [CrossRef]
- Preti, R.; Antonelli, M.L.; Bernacchia, R.; Vinci, G. Fast determination of biogenic amines in beverages by a core-shell particle column. Food Chem. 2015, 187, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Glória, M.B.A.; Watson, B.T.; Simon-Sarkadi, L.; Daeschel, M.A. A survey of biogenic amines in Oregon Pinot Noir and Cabernet Sauvignon wines. Am. J. Enol. Vitic. 1998, 49, 279–282. [Google Scholar]
- Vazquez-Lasa, M.B.; Iniguez-Crespo, M.; González-Larraina, M.; González-Guerrero, A. Biogenic amines in Rioja wines. Am. J. Enol. Vitic. 1998, 49, 229. [Google Scholar]
- Busto, O.; Mestres, M.; Guasch, J.; Borrull, F. Determination of biogenic amines in wine after clean-up by solid-phase extraction. Chromatographia 1995, 40, 404–410. [Google Scholar] [CrossRef]
- Ortega-Heras, M.; Pérez-Magariño, S.; Del-Villar-Garrachón, V.; González-Huerta, C.; Moro Gonzalez, L.C.; Guadarrama Rodríguez, A.; Villanueva Sanchez, S.; Gallo González, R.; Martín de la Helguera, S. Study of the effect of vintage, maturity degree, and irrigation on the amino acid and biogenic amine content of a white wine from the Verdejo variety. J. Sci. Food Agric. 2014, 94, 2073–2082. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Arribas, M.V.; Smit, A.Y.; du Toit, M. Biogenic amines and the winemaking process. In Understanding and Managing Wine Quality and Safety; Reynolds, A.G., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2010. [Google Scholar]
- Preti, R.; Vinci, G. Biogenic amine content in red wines from different protected designations of origin of Southern Italy: Chemometric characterization and classification. Food Anal. Methods 2016, 9, 2280–2287. [Google Scholar] [CrossRef]
- Torrea, D.; Ancín, C. Content of biogenic amines in a Chardonnay wine obtained through spontaneous and inoculated fermentations. J. Agric. Food Chem. 2002, 50, 4895–4899. [Google Scholar] [CrossRef] [PubMed]
- Caruso, M.; Fiore, C.; Contursi, M.; Salzano, G.; Paparella, A.; Romano, P. Formation of biogenic amines as criteria for the selection of wine yeasts. World J. Microbiol. Biotechnol. 2002, 18, 159–163. [Google Scholar] [CrossRef]
- Granchi, L.; Romano, P.; Mangani, S.; Guerrini, S.; Vincenzini, M. Production of biogenic amines by wine microorganisms. Bulletin de l’O.I.V. 2005, 78, 595–609. [Google Scholar]
- Torrea, D.; Azpilicueta, C.A. Influence of yeast strain on biogenic amines content in wines: Relationship with the utilization of amino acids during fermentation. Am. J. Enol. Vitic. 2001, 52, 185–190. [Google Scholar]
- Tristezza, M.; Vetrano, C.; Bleve, G.; Spano, G.; Capozzi, V.; Logrieco, A.; Mita, G.; Grieco, F. Biodiversity and safety aspects of yeast strains characterized from vineyards and spontaneous fermentations in the Apulia Region, Italy. Food Microbiol. 2013, 36, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; de las Rivas, B.; Marcobal, A.; Muñoz, R. Molecular methods for the detection of biogenic amine-producing bacteria on foods. Int. J. Food Microbiol. 2007, 117, 258–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcobal, A.; Martin-Alvarez, P.J.; Polo, M.C.; Munoz, R.; Moreno-Arribas, M.V. Formation of Biogenic Amines throughout the Industrial Manufacture of Red Wine. J. Food Prot. 2006, 69, 397–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Arribas, M.V.; Polo, M.C.; Jorganes, F.; Muñoz, R. Screening of biogenic amine production by lactic acid bacteria isolated from grape must and wine. Int. J. Food Microbiol. 2003, 84, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Henríquez-Aedo, K.; Durán, D.; Garcia, A.; Hengst, M.B.; Aranda, M. Identification of biogenic amines-producing lactic acid bacteria isolated from spontaneous malolactic fermentation of chilean red wines. LWT Food Sci. Technol. 2016, 68, 183–189. [Google Scholar] [CrossRef]
- Lucas, P.; Landete, J.; Coton, M.; Coton, E.; Lonvaud-Funel, A. The tyrosine decarboxylase operon of Lactobacillus brevis IOEB 9809: Characterization and conservation in tyramine-producing bacteria. FEMS Microbiol. Lett. 2003, 229, 65–71. [Google Scholar] [CrossRef]
- Romano, A.; Trip, H.; Lonvaud-Funel, A.; Lolkema, J.S.; Lucas, P.M. Evidence of two functionally distinct ornithine decarboxylation systems in lactic acid bacteria. Appl. Environ. Microbiol. 2012, 78, 1953–1961. [Google Scholar] [CrossRef] [PubMed]
- Molenaar, D.; Bosscher, J.S.; Ten Brink, B.; Driessen, A.J.M.; Konings, W.N. Generation of a proton motive force by histidine decarboxylation and electrogenic histidine/histamine antiport in Lactobacillus buchneri. J. Bacteriol. 1993, 175, 2864–2870. [Google Scholar] [CrossRef] [PubMed]
- Cruz Martín, M.; Fernández, M.; Linares, D.M.; Alvarez, M.A. Sequencing, characterization and transcriptional analysis of the histidine decarboxylase operon of Lactobacillus buchneri. Microbiology 2005, 151, 1219–1228. [Google Scholar] [CrossRef] [PubMed]
- Spano, G.; Russo, P.; Lonvaud-Funel, A.; Lucas, P.; Alexandre, H.; Grandvalet, C.; Coton, E.; Coton, M.; Barnavon, L.; Bach, B.; et al. Biogenic amines in fermented foods. Eur. J. Clin. Nutr. 2010, 64, S95–S100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pessione, A.; Lamberti, C.; Pessione, E. Proteomics as a tool for studying energy metabolism in lactic acid bacteria. Mol. Biosyst. 2010, 6, 1419–1430. [Google Scholar] [CrossRef] [PubMed]
- Pessione, E.; Mazzoli, R.; Giuffrida, M.G.; Lamberti, C.; Garcia-Moruno, E.; Barello, C.; Conti, A.; Giunta, C. A proteomic approach to studying biogenic amine producing lactic acid bacteria. Proteomics 2005, 5, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Lonvaud-Funel, A. Lactic acid bacteria in the quality improvement and depreciation of wine. Antonie Van Leeuwenhoek 1999, 76, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Arribas, V.; Lonvaud-Funel, A. Purification and characterization of tyrosine decarboxylase of Lactobacillus brevis IOEB 9809 isolated from wine. FEMS Microbiol. Lett. 2001, 195, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Guirard, B.; Snell, E. Purification and Properties of Ornithine Decarboxylase from Lactobacillus sp. 30a. J. Biol. Chem. 1980, 255, 5960–5964. [Google Scholar] [PubMed]
- Marcobal, Á.; Martín-Álvarez, P.J.; Moreno-Arribas, M.V.; Muñoz, R. A multifactorial design for studying factors influencing growth and tyramine production of the lactic acid bacteria Lactobacillus brevis CECT 4669 and Enterococcus faecium BIFI-58. Res. Microbiol. 2006, 157, 417–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardini, F.; Özogul, Y.; Suzzi, G.; Tabanelli, G.; Özogul, F. Technological factors affecting biogenic amine content in foods: A review. Front. Microbiol. 2016, 7, 1218. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; Pardo, I.; Ferrer, S. Regulation of hdc expression and HDC activity by enological factors in lactic acid bacteria. J. Appl. Microbiol. 2008, 105, 1544–1551. [Google Scholar] [CrossRef] [PubMed]
- Wibowo, D.; Eschenbruch, R.; Davis, C.R.; Fleet, G.H.; Lee, T.H. Occurrence and growth of lactic acid bacteria in wine: A review. Am. J. Enol. Vitic. 1985, 36, 302–313. [Google Scholar]
- Coton, E.; Rollan, G.; Bertrand, A.; Lonvaud-Funel, A. Histamine-producing lactic acid bacteria in wines: Early detection, frequency and distribution. Am. J. Enol. Vitic. 1998, 49, 199–204. [Google Scholar]
- Guerrini, S.; Mangani, S.; Granchi, L.; Vincenzini, M. Biogenic Amine Production by Oenococcus oeni. Curr. Microbiol. 2002, 44, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Costantini, A.; Cersosimo, M.; Del Prete, V.; Garcia-Moruno, E. Production of biogenic amines by lactic acid bacteria: Screening by PCR, thin-layer chromatography, and high-performance liquid chromatography of strains isolated from wine and must. J. Food Prot. 2006, 69, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Moruno, E.; Muñoz, R. Does Oenococcus oeni produce histamine? Int. J. Food Microbiol. 2012, 157, 121–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcobal, A.; de Las Rivas, B.; Moreno-Arribas, M.V.; Muñoz, R. Identification of the ornithine decarboxylase gene in the putrescine-producer Oenococcus oeni BIFI-83. FEMS Microbiol. Lett. 2004, 239, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Coton, E.; Coton, M. Multiplex PCR for colony direct detection of Gram-positive histamine- and tyramine-producing bacteria. J. Microbiol. Methods 2005, 63, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Lucas, P.M.; Claisse, O.; Lonvaud-Funel, A. High Frequency of Histamine-Producing Bacteria in the Enological Environment and Instability of the Histidine Decarboxylase Production Phenotype. Appl. Environ. Microbiol. 2008, 74, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Costantini, A.; García-Moruno, E.; Moreno-Arribas, M.V. Biochemical Transformations Produced by Malolactic Fermentation; Springer: New York, NY, USA, 2009; ISBN 9780387741161. [Google Scholar]
- Moreno-Arribas, V.; Torlois, S.; Joyeux, A.; Bertrand, A.; Lonvaud-Funel, A. Isolation, properties and behaviour of tyramine-producing lactic acid bacteria from wine. J. Appl. Microbiol. 2000, 88, 584–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebastian, P.; Herr, P.; Fischer, U.; König, H. Molecular identification of lactic acid bacteria occurring in must and wine. S. Afr. J. Enol. Vitic. 2011, 32, 300–309. [Google Scholar] [CrossRef]
- Arena, M.E.; Fiocco, D.; Manca de Nadra, M.C.; Pardo, I.; Spano, G. Characterization of a Lactobacillus plantarum Strain Able to Produce Tyramine and Partial Cloning of a Putative Tyrosine Decarboxylase Gene. Curr. Microbiol. 2007, 55, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, N.; Hansen, W.; Engesser, D.; Hammes, W.P.; Holzapfel, W.H. Formation of histamine and tyramine by lactic acid bacteria in decarboxylase assay medium. Lett. Appl. Microbiol. 1990, 11, 278–281. [Google Scholar] [CrossRef]
- Del Prete, V.; Costantini, A.; Cecchini, F.; Morassut, M.; Garcia-Moruno, E. Occurrence of biogenic amines in wine: The role of grapes. Food Chem. 2009, 112, 474–481. [Google Scholar] [CrossRef]
- Zhijun, L.; Yongning, W.; Gong, Z.; Yunfeng, Z.; Changhu, X. A survey of biogenic amines in chinese red wines. Food Chem. 2007, 105, 1530–1535. [Google Scholar] [CrossRef]
- Landete, J.M.; Ferrer, S.; Polo, L.; Pardo, I. Biogenic amines in wines from three Spanish regions. J. Agric. Food Chem. 2005, 53, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Soufleros, E.H.; Bouloumpasi, E.; Zotou, A.; Loukou, Z. Determination of biogenic amines in Greek wines by HPLC and ultraviolet detection after dansylation and examination of factors affecting their presence and concentration. Food Chem. 2007, 101, 704–716. [Google Scholar] [CrossRef]
- Pineda, A.; Carrasco, J.; Peña-Farfal, C.; Henríquez-Aedo, K.; Aranda, M. Preliminary evaluation of biogenic amines content in Chilean young varietal wines by HPLC. Food Control 2012, 23, 251–257. [Google Scholar] [CrossRef]
- Cecchini, F.; Morassut, M. Effect of grape storage time on biogenic amines content in must. Food Chem. 2010, 123, 263–268. [Google Scholar] [CrossRef]
- Leitão, M.C.; Marques, A.P.; San Romão, M.V. A survey of biogenic amines in commercial Portuguese wines. Food Control 2005, 16, 199–204. [Google Scholar] [CrossRef]
- Ough, C.S.; Daudt, C.E.; Crowell, E.A. Identification of new volatile amines in grapes and wines. J. Agric. Food Chem. 1981, 29, 938–941. [Google Scholar] [CrossRef] [PubMed]
- Broquedis, M.; Dumery, B.; Boucard, J. Mise en evidence de polyamines (putrescine, cadaverine, norspermidine, spermidine et spermine) dans les feuilles et les grappes de Vitis vinifera L. Connaiss. la Vigne du Vin 1989, 23, 1–6. [Google Scholar]
- Vidal-Carou, M.C.; Codony-Salcedo, R.; Mariné-Font, A. Histamine and tyramine in spanish wines: Relationships with total sulfur dioxide level, volatile acidity and malo-lactic fermentation intensity. Food Chem. 1990, 35, 217–227. [Google Scholar] [CrossRef]
- Herbert, P.; Cabrita, M.J.; Ratola, N.; Laureano, O.; Alves, A. Free amino acids and biogenic amines in wines and musts from the Alentejo region. Evolution of amines during alcoholic fermentation and relationship with variety, sub-region and vintage. J. Food Eng. 2005, 66, 315–322. [Google Scholar] [CrossRef]
- Bauza, T.; Kelly, M.T.; Blaise, A. Study of polyamines and their precursor amino acids in Grenache noir and Syrah grapes and wine of the Rhone Valley. Food Chem. 2007, 105, 405–413. [Google Scholar] [CrossRef]
- Soufleros, E.; Barrios, M.-L.; Bertrand, A. Correlation between the content of biogenic amines and other wine compounds. Am. J. Enol. Vitic. 1998, 49, 266–278. [Google Scholar]
- García-Marino, M.; Trigueros, Á.; Escribano-Bailón, T. Influence of oenological practices on the formation of biogenic amines in quality red wines. J. Food Compos. Anal. 2010, 23, 455–462. [Google Scholar] [CrossRef] [Green Version]
- Gerbaux, V.; Money, C. Biogenic amines in Burgundy wines. Contents and origin in wines. Rev. Fr. Oenol. 2000, 183, 25–28. [Google Scholar]
- Henríquez-Aedo, K.; Galarce-Bustos, O.; Aqueveque, P.; García, A.; Aranda, M. Dynamic of biogenic amines and precursor amino acids during cabernet sauvignon vinification. LWT 2018, 97, 238–244. [Google Scholar] [CrossRef]
- Ordóñez, J.L.; Callejón, R.M.; Troncoso, A.M.; García–Parrilla, M.C. Evaluation of biogenic amines profile in opened wine bottles: Effect of storage conditions. J. Food Compos. Anal. 2017, 63, 139–147. [Google Scholar] [CrossRef]
- Smit, A.Y.; du Toit, W.J.; Stander, M.; du Toit, M. Evaluating the influence of maceration practices on biogenic amine formation in wine. LWT Food Sci. Technol. 2013, 53, 297–307. [Google Scholar] [CrossRef]
- Martínez-Pinilla, O.; Guadalupe, Z.; Hernández, Z.; Ayestarán, B. Amino acids and biogenic amines in red varietal wines: The role of grape variety, malolactic fermentation and vintage. Eur. Food Res. Technol. 2013, 237, 887–895. [Google Scholar] [CrossRef]
- Marques, A.P.; Leitão, M.C.; San Romão, M.V. Biogenic amines in wines: Influence of oenological factors. Food Chem. 2008, 107, 853–860. [Google Scholar] [CrossRef]
- Hernandez-Orte, P.; Guitart, A.; Cacho, J. Changes in the Concentration of Amino Acids During the Ripening of Vitis vinifera Tempranillo Variety from the Denomination d’Origine Somontano (Spain). Am. J. Enol. Vitic. 1999, 50, 144–154. [Google Scholar]
- Soleas, G.J.; Carey, M.; Goldberg, D.M. Method development and cultivar-related differences of nine biogenic amines in Ontario wines. Food Chem. 1999, 64, 49–58. [Google Scholar] [CrossRef]
- Bauza, T.; Blaise, A.; Daumas, F.; Cabanis, J.C. Determination of biogenic amines and their precursor amino acids in wines of the Vallée du Rhône by high-performance liquid chromatography with precolumn derivatization and fluorimetric detection. J. Chromatogr. A 1995, 707, 373–379. [Google Scholar] [CrossRef]
- Martín-Álvarez, P.J.; Marcobal, A.; Polo, C.; Moreno-Arribas, M.V. Influence of technological practices on biogenic amine contents in red wines. Eur. Food Res. Technol. 2006, 222, 420–424. [Google Scholar] [CrossRef]
- Pogorzelski, E. Studies on the formation of histamine in must and wines from elderberry fruit. J. Sci. Food Agric. 1992, 60, 239–244. [Google Scholar] [CrossRef]
- Ancín-Azpilicueta, C.; González-Marco, A.; Jiménez-Moreno, N. Comparative study of the amine concentration in wines obtained from the traditional fermentation and from a more anaerobic fermentation method. LWT Food Sci. Technol. 2010, 43, 771–776. [Google Scholar] [CrossRef]
- Galgano, F.; Caruso, M.; Condelli, N.; Favati, F. Focused review: Agmatine in fermented foods. Front. Microbiol. 2012, 3, 199. [Google Scholar] [CrossRef] [PubMed]
- Tassoni, A.; Tango, N.; Ferri, M. Polyphenol and Biogenic Amine Profiles of Albana and Lambrusco Grape Berries and Wines Obtained Following Different Agricultural and Oenological Practices. Food Nutr. Sci. 2014, 5, 8–16. [Google Scholar] [CrossRef]
- Tassoni, A.; Tango, N.; Ferri, M. Comparison of biogenic amine and polyphenol profiles of grape berries and wines obtained following conventional, organic and biodynamic agricultural and oenological practices. Food Chem. 2013, 139, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Marcobal, A.; Polo, M.C.; Martín-Álvarez, P.J.; Moreno-Arribas, M.V. Biogenic amine content of red Spanish wines: Comparison of a direct ELISA and an HPLC method for the determination of histamine in wines. Food Res. Int. 2005, 38, 387–394. [Google Scholar] [CrossRef]
- Henríquez-Aedo, K.; Vega, M.; Prieto-Rodríguez, S.; Aranda, M. Evaluation of biogenic amines content in chilean reserve varietal wines. Food Chem. Toxicol. 2012, 50, 2742–2750. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Alonso, S.; Hermosín-Gutiérrez, I.; García-Romero, E. Simultaneous HPLC analysis of biogenic amines, amino acids, and ammonium ion as aminoenone derivatives in wine and beer samples. J. Agric. Food Chem. 2007, 55, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Pereira, V.; Pontes, M.; Câmara, J.S.; Marques, J.C. Simultaneous analysis of free amino acids and biogenic amines in honey and wine samples using in loop orthophthalaldeyde derivatization procedure. J Chromatogr A 2008, 1189, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Önal, A.; Tekkeli, S.E.K.; Önal, C. A review of the liquid chromatographic methods for the determination of biogenic amines in foods. Food Chem. 2013, 138, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Bedia Erim, F. Recent analytical approaches to the analysis of biogenic amines in food samples. TrAC Trends Anal. Chem. 2013, 52, 239–247. [Google Scholar] [CrossRef]
- Jiang, H.-L.; Ying, L.-Y.; Zhou, S.-C.; Ying, M.; Shen, W.; Qiu, D.-H. Chromatographic determination of biogenic amines in wines after treatment with ionic liquids as novel media. J. Sep. Sci. 2011, 34, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Arrieta, M.P.; Prats-Moya, M.S. Free amino acids and biogenic amines in Alicante Monastrell wines. Food Chem. 2012, 135, 1511–1519. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.T.; Blaise, A.; Larroque, M. Rapid automated high performance liquid chromatography method for simultaneous determination of amino acids and biogenic amines in wine, fruit and honey. J. Chromatogr. A 2010, 1217, 7385–7392. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Q.; Ye, D.-Q.; Zhu, B.-Q.; Wu, G.-F.; Duan, C.-Q. Rapid HPLC analysis of amino acids and biogenic amines in wines during fermentation and evaluation of matrix effect. Food Chem. 2014, 163, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Saurina, J.; Hernández-Cassou, S. Chromatographic determination of amino acids by pre-column derivatization using 1,2-naphthoquinone-4-sulfonate as reagent. J. Chromatogr. A 1996, 740, 21–30. [Google Scholar] [CrossRef]
- Latorre, R.M.; Hernández-Cassou, S.; Saurina, J. Strategies for in-capillary derivatization of amino acids in capillary electrophoresis using 1,2-naphthoquinone-4-sulfonate as a labeling reagent. J. Chromatogr. A 2001, 934, 105–112. [Google Scholar] [CrossRef]
- Hlabangana, L.; Hernández-Cassou, S.; Saurina, J. Determination of biogenic amines in wines by ion-pair liquid chromatography and post-column derivatization with 1,2-naphthoquinone-4-sulphonate. J. Chromatogr. A 2006, 1130, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Jastrzębska, A.; Piasta, A.; Kowalska, S.; Krzemiński, M.; Szłyk, E. A new derivatization reagent for determination of biogenic amines in wines. J. Food Compos. Anal. 2016, 48, 111–119. [Google Scholar] [CrossRef]
- García-Moruno, E.; Carrascosa, A.V.; Muñoz, R. A rapid and inexpensive method for the determination of biogenic amines from bacterial cultures by thin-layer chromatography. J. Food Prot. 2005, 68, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Romano, A.; Klebanowski, H.; La Guerche, S.; Beneduce, L.; Spano, G.; Murat, M.-L.; Lucas, P. Determination of biogenic amines in wine by thin-layer chromatography/densitometry. Food Chem. 2012, 135, 1392–1396. [Google Scholar] [CrossRef] [PubMed]
- Papageorgiou, M.; Lambropoulou, D.; Morrison, C.; Kłodzińska, E.; Namieśnik, J.; Płotka-Wasylka, J. Literature update of analytical methods for biogenic amines determination in food and beverages. TrAC Trends Anal. Chem. 2018, 98, 128–142. [Google Scholar] [CrossRef]
- Ginterová, P.; Marák, J.; Staňová, A.; Maier, V.; Ševčík, J.; Kaniansky, D. Determination of selected biogenic amines in red wines by automated on-line combination of capillary isotachophoresis–capillary zone electrophoresis. J. Chromatogr. B 2012, 904, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Daniel, D.; dos Santos, V.B.; Vidal, D.T.R.; do Lago, C.L. Determination of biogenic amines in beer and wine by capillary electrophoresis–tandem mass spectrometry. J. Chromatogr. A 2015, 1416, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Uzaşçı, S.; Başkan, S.; Erim, F.B. Biogenic Amines in Wines and Pomegranate Molasses—A Non-Ionic Micellar Electrokinetic Chromatography Assay with Laser-Induced Fluorescence Detection. Food Anal. Methods 2012, 5, 104–108. [Google Scholar] [CrossRef]
- Pereira, V.; Andreia, M.; Leca, J.M.; Marques, J.C. Analytical methodologies for the determination of biogenic amines in wines: An overview of the recent trends. J. Anal. Bioanal. Sep. Tech. 2017, 2, 52–57. [Google Scholar]
- Alonso-Lomillo, M.A.; Domínguez-Renedo, O.; Matos, P.; Arcos-Martínez, M.J. Disposable biosensors for determination of biogenic amines. Anal. Chim. Acta 2010, 665, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Di Fusco, M.; Federico, R.; Boffi, A.; Macone, A.; Favero, G.; Mazzei, F.; Di Fusco, M.; Federico, R.; Boffi, A.; Macone, A.; et al. Characterization and application of a diamine oxidase from Lathyrus sativus as component of an electrochemical biosensor for the determination of biogenic amines in wine and beer. Anal. Bioanal. Chem. 2011, 401, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Henao-Escobar, W.; del Torno-de Román, L.; Domínguez-Renedo, O.; Alonso-Lomillo, M.A.; Arcos-Martínez, M.J. Dual enzymatic biosensor for simultaneous amperometric determination of histamine and putrescine. Food Chem. 2016, 190, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Le Jeune, C.; Lonvaud-Funel, A.; ten Brink, B.; Hofstra, H.; van der Vossen, J.M. Development of a detection system for histidine decarboxylating lactic acid bacteria based on DNA probes, PCR and activity test. J. Appl. Bacteriol. 1995, 78, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Lucas, P.; Lonvaud-Funel, A. Purification and partial gene sequence of the tyrosine decarboxylase of Lactobacillus brevis IOEB 9809. FEMS Microbiol. Lett. 2002, 211, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Marcobal, A.; de las Rivas, B.; Moreno-Arribas, M.V.; Munoz, R. Multiplex PCR Method for the Simultaneous Detection of Histamine-, Tyramine-, and Putrescine-Producing Lactic Acid Bacteria in Foods. J. Food Prot. 2005, 68, 874–878. [Google Scholar] [CrossRef] [PubMed]
- De las Rivas, B.; Marcobal, A.; Carrascosa, A.V.; Munoz, R. PCR Detection of Foodborne Bacteria Producing the Biogenic Amines Histamine, Tyramine, Putrescine, and Cadaverine. J. Food Prot. 2006, 69, 2509–2514. [Google Scholar] [CrossRef] [PubMed]
- Costantini, A.; Doria, F.; Vaudano, E.; Garcia-Moruno, E. Chemical and molecular methods for the control of biogenic amine production by microorganisms. Ann. Microbiol. 2011, 61, 173–178. [Google Scholar] [CrossRef]
- Nannelli, F.; Claisse, O.; Gindreau, E.; De Revel, G.; Lonvaud-Funel, A.; Lucas, P.M. Determination of lactic acid bacteria producing biogenic amines in wine by quantitative PCR methods. Lett. Appl. Microbiol. 2008, 47, 594–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, M.; del Río, B.; Linares, D.M.; Martín, M.C.; Alvarez, M.A. Real-Time Polymerase Chain Reaction for Quantitative Detection of Histamine-Producing Bacteria: Use in Cheese Production. J. Dairy Sci. 2006, 89, 3763–3769. [Google Scholar] [CrossRef] [Green Version]
- Coton, M.; Coton, E.; Lucas, P.; Lonvaud, A. Identification of the gene encoding a putative tyrosine decarboxylase of Carnobacterium divergens 508. Development of molecular tools for the detection of tyramine-producing bacteria. Food Microbiol. 2004, 21, 125–130. [Google Scholar] [CrossRef]
- Marcobal, Á.; de las Rivas, B.; García-Moruno, E.; Muñoz, R. The Tyrosine Decarboxylation Test Does Not Differentiate Enterococcus faecalis from Enterococcus faecium. Syst. Appl. Microbiol. 2004, 27, 423–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández, M.; Linares, D.M.; Alvarez, M.A. Sequencing of the Tyrosine Decarboxylase Cluster of Lactococcus lactis IPLA 655 and the Development of a PCR Method for Detecting Tyrosine Decarboxylating Lactic Acid Bacteria. J. Food Prot. 2004, 67, 2521–2529. [Google Scholar] [CrossRef] [PubMed]
- Mohedano, M.L.; Spano, G.; López, P.; Russo, P. Controlling the formation of biogenic amines in fermented foods. In Advances in Fermented Foods and Beverages. Improving Quality, Technologies and Health Benefits; Holzapfel, W., Ed.; Woodhead Publishing: Cambridge, UK, 2014; pp. 273–310. [Google Scholar]
- García-Ruiz, A.; González-Rompinelli, E.M.; Bartolomé, B.; Moreno-Arribas, M.V. Potential of wine-associated lactic acid bacteria to degrade biogenic amines. Int. J. Food Microbiol. 2011, 148, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capozzi, V.; Russo, P.; Ladero, V.; Fernández, M.; Fiocco, D.; Alvarez, M.A.; Grieco, F.; Spano, G. Biogenic Amines Degradation by Lactobacillus plantarum: Toward a Potential Application in Wine. Front. Microbiol. 2012, 3, 122. [Google Scholar] [CrossRef] [PubMed]
- Callejón, S.; Sendra, R.; Ferrer, S.; Pardo, I. Identification of a novel enzymatic activity from lactic acid bacteria able to degrade biogenic amines in wine. Appl. Microbiol. Biotechnol. 2014, 98, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Beneduce, L.; Romano, A.; Capozzi, V.; Lucas, P.; Barnavon, L.; Bach, B.; Vuchot, P.; Grieco, F.; Spano, G. Biogenic amine in wines. Ann. Microbiol. 2010, 60, 573–578. [Google Scholar] [CrossRef]
- Bäumlisberger, M.; Moellecken, U.; König, H.; Claus, H.; Bäumlisberger, M.; Moellecken, U.; König, H.; Claus, H. The Potential of the Yeast Debaryomyces hansenii H525 to Degrade Biogenic Amines in Food. Microorganisms 2015, 3, 839–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Biogenic amine formation from their precursor amino acids and their intake/uptake by the transmembrane antiporter.
Figure 1.
Biogenic amine formation from their precursor amino acids and their intake/uptake by the transmembrane antiporter.

Figure 2.
(A) Biogenic amine levels (μmol/kg) measured in Albana and Lambrusco berries grown following conventional (AC and LC), organic (AO and LO) and biodynamic (AB and LB) agricultural practices. (B) Biogenic amine levels (μmol/L) measured in Albana and Lambrusco wines obtained following conventional (AC), organic (AO and LO) and biodynamic (AB and LB) oenological practices. The star symbol indicates the statistically significant differences (Student’s t-test, p < 0.05) among single compounds of the same group. Data are the mean ± SE (n = 4). (from Tassoni et al. [85]).
Figure 2.
(A) Biogenic amine levels (μmol/kg) measured in Albana and Lambrusco berries grown following conventional (AC and LC), organic (AO and LO) and biodynamic (AB and LB) agricultural practices. (B) Biogenic amine levels (μmol/L) measured in Albana and Lambrusco wines obtained following conventional (AC), organic (AO and LO) and biodynamic (AB and LB) oenological practices. The star symbol indicates the statistically significant differences (Student’s t-test, p < 0.05) among single compounds of the same group. Data are the mean ± SE (n = 4). (from Tassoni et al. [85]).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Biogenic amines found in different grape and wine.
| Wine | Type of Wine | Biogenic Amine | Reference |
|---|---|---|---|
| Cabernet Franc | red | Eth,Ety, Put, Agm | [58] |
| Carmenere | red | Eth,Ety, Put, Agm | |
| Cesanese d’Affile | red | Eth,Ety, Put, Agm | |
| Merlot | red | Eth,Ety, Put, Agm | |
| Montepulciano | red | Eth,Ety, Put, Agm | |
| Sangiovese | red | Eth,Ety, Put, Agm | |
| Syrah | red | Eth,Ety, Put, Agm | |
| Aglianico (13) | red | Met, Eth, Agm, Pea, Put, Cad, Him, Spd, Tym | [22] |
| Primitivo di Manduria (15) | red | Eth, Agm, Pea, Put, Cad, Him, Spd, Tym | |
| Syrah (15) | red | Met, Eth, Agm, Pea, Put, Cad, Him, Spd, Tym | |
| Etna Rosso (13) | red | Met, Eth, Agm, Pea, Put, Cad, Him, Spd, Tym | |
| Chinese red wines from Shandong | red | Phe, Put, Cad, Him, Tyr, Spd and Spm | [59] |
| Chinese red wines from Beijing | red | Phe, Put, Cad, Him, Tyr, Spd and Spm | |
| Chinese red wines from Tianjin | red | Phe, Put, Cad, Him, Tyr, Spd and Spm | |
| Chinese red wines from Hebei | red | Phe, Put, Cad, Him, Tyr, Spd and Spm | |
| Chinese red wines from Xinjiang | red | Phe, Put, Cad, Him, Tyr | |
| Tempranillo from La Rioja, | red | Him, Tyr, Put, Phe | [60] |
| Tempranillo from Utiel-Requena | red | Him, Tyr, Put, Phe | |
| Tempranillo from Tarragona | red | Him, Tyr, Put, Phe | |
| Bobal | red | Him, Tyr, Put, Phe | |
| Garnacha | red | Him, Tyr, Put, Phe | |
| Xinomavro | red | [61] Note: amines >1 mg/L are indicated | |
| Roditis | white | Phe | |
| Agiorgitiko | red | Phe, Put | |
| Cabernet Sauvignon | red | ||
| Mantilaria | red | Ism, Him, Tym, Spd, Put | |
| Syrah | red | Put | |
| Merlot | red | Ism | |
| Debina | white | Cad | |
| Moshofilero | white | Put | |
| Malagouzia | white | ||
| Asyrtiko | red | Put, Cad | |
| Grenache rouge | red | Put | |
| Chardonnay | white | ||
| Muscat white | white | Ism, Put | |
| Muscat Hamburg | red | ||
| Muscat d’Alexandrie | white | Put, Cad, Him | |
| Limnio | red | Cad | |
| Chilean young wines: | Put, Him, Tyr, Spd | [62] | |
| Cabernet Sauvignon (9) | red | ||
| Merlot (8) | red | ||
| Carménère (10) | red |
Table 2.
Individual biogenic amines during winemaking (from Del Prete et al. [58]).
Table 2.
Individual biogenic amines during winemaking (from Del Prete et al. [58]).
| Biogenic Amines (mg/L) | Must | Must 72h | Wine/AF | Wine/MLF |
|---|---|---|---|---|
| Ethanolamine | 7.91 a | 8.73 a | 11.54 b | 11.65 b |
| Agmatime | nd a | nd a | 0.51 a | 6.4 b |
| Ethylamine | 9.83 b | 10.07 b | 1.87 a | 1.96 a |
| Tyramine | nd a | nd a | 0.36 b | 0.069 a |
| Putrescine | 11.23 d | 3.76 c | 1.76 a | 2.26 ab |
Mean concentration of individual BA in all cultivars in years 2004–2005; Mean values with the same superscript letters in the same line, do not significantly differ (p < 0.05, LSD, Least Significant Difference, test); Letters a, b indicates the statistically significant differences; AF: Alcoholic fermentation; MLF: Malolactic fermentation; nd: not detected.
Table 3.
Quantification of biogenic amines (BAs) levels (mg/L) during vinification (from Henriquez-Aedo et al. [73]).
Table 3.
Quantification of biogenic amines (BAs) levels (mg/L) during vinification (from Henriquez-Aedo et al. [73]).
| Winery | Sample | Phe | Put | Cad | Him | Tyr | Spd | Spm | Total |
|---|---|---|---|---|---|---|---|---|---|
| A | grapes | nd | nd | nd | nd | nd | tr | tr | tr |
| AF | nd | 29.2 | tr | nd | nd | tr | tr | 29.2 | |
| MLF | tr | 11 | tr | nd | nd | tr | tr | 11 | |
| Wine | nd | 36.7 | tr | nd | nd | 1.6 | tr | 38.3 | |
| B | grapes | nd | nd | nd | nd | tr | tr | tr | tr |
| AF | nd | 4 | tr | nd | tr | tr | tr | 5 | |
| MLF | tr | 9.2 | tr | nd | tr | tr | tr | 9.2 | |
| Wine | nd | 5.5 | tr | nd | tr | tr | tr | 5.5 | |
| C | grapes | tr | 1.2 | tr | 1.9 | nd | 2.2 | 2.5 | 7.8 |
| AF | nd | 9.3 | tr | nd | tr | 1.3 | tr | 10.7 | |
| MLF | nd | 14.1 | tr | nd | nd | tr | tr | 14.2 | |
| Wine | 10 | 142.1 | tr | 8.4 | 8.1 | tr | tr | 168.6 | |
| D | grapes | nd | 8.9 | nd | nd | tr | 2.9 | tr | 11.8 |
| AF | nd | 38.6 | tr | tr | tr | 1.6 | tr | 40.2 | |
| MLF | nd | 11.7 | 1.3 | 1.3 | tr | 2.2 | tr | 13.9 | |
| Wine | tr | 8.5 | 3.5 | 3.5 | 1.5 | 2.7 | tr | 16.2 | |
| E | grapes | nd | 9.8 | tr | nd | tr | 4.1 | 1.2 | 15.1 |
| AF | nd | 14.7 | tr | nd | tr | 1.1 | tr | 15.8 | |
| MLF | nd | 12.9 | tr | nd | tr | 1.3 | tr | 14.2 | |
| Wine | nd | 14.3 | tr | nd | tr | 1.5 | tr | 15.8 |
nd: not detected; tr: traces, amount below calibration range; AF: Alcoholic fermentation; MLF: malolactic fermentation.
Table 4.
List of the primers described in literature for the detection of decarboxylase genes (hdc, odc, tdc).
Table 4.
List of the primers described in literature for the detection of decarboxylase genes (hdc, odc, tdc).
| Primer | 5′ → 3′Sequence | Coding for | Reference |
|---|---|---|---|
| CL1 | CCWGGWAAWATWGGWAATGGWTA | hdc | [111] |
| CL2 | GAWGCWGTWGTCATATTWATTTGWCC | hdc | [111] |
| JV16HC | AGATGGTATTGTTTCTTATG | hdc | [111] |
| JV17HC | AGACCATACACCATAACCTT | hdc | [111] |
| JV17 | AGACCATACACCATAACCTTG | hdc | [46] |
| CL1mod | CCAGGWAACATTGGTAATGGATA | hdc | [60] |
| HDC3 | GATGGTATTGTTTCKTATGA | hdc | [51] |
| HDC4 | CAAACACCAGCATCTTC | hdc | [51] |
| PHDC1 | CCGTGCGGAAACAAAGAAT | hdc | [48] |
| PHDC2 | CCAAACACCAGCATCTTCA | hdc | [48] |
| HIS1-F | GGNATNGTNWSNTAYGAYMGNGCNGA | hdc | [114] |
| HIS1-R | ATNGCDATNGCNSWCCANACNCCRTA | hdc | [114] |
| Hdc1 | TTGACCGTATCTCAGTGAGTCCAT | hdc | [117] |
| Hdc2 | ACGGTCATACGAAACAATACCATC | hdc | [117] |
| 3 | GTNTTYAAYGCNGAYAARACNTAYTTYGT | odc | [50] |
| 16 | TACRCARAATACTCCNGGNGGRTANGG | odc | [50] |
| 4 | ATNGARTTNAGTTCRCAYTTYTCNGG | odc | [113] |
| 15 | GGTAYTGTTYGAYCGGAAWAAWCAYAA | odc | [113] |
| AODC1 | GMTCGTGAAATYAARCKG | odc | [48] |
| AODC2 | KGRGTTCMGCYGGRGTAT | odc | [48] |
| Put1-F | TWYMAYGCNGAYAARACNTAYYYTGT | odc | [114] |
| Put1-R | ACRCANAGNACNCCNGNGGRTANGG | odc | [114] |
| Put2-F | ATHWGNTWYGGNAAYACNATHAARAA | odc | [114] |
| Put2-R | GCNARNCCNCCRAAYTTNCCDARTC | odc | [114] |
| P2-for | GAYATIATIGGIATIGGIYTIGAYCARG | tdc | [112] |
| P1-rev | CCRTARTCIGGIATIGCRAARTCIGTRTG | tdc | [112] |
| 41 | CAYGTNGAYGCNGCNTAYGGNGG | tdc | [113] |
| 42 | AYRTANCCCATYTTRTGNGGRTC | tdc | [113] |
| Pt3 | TACACGTAGATGCTGCATATG | tdc | [48] |
| Pt4 | ATGGTTGACTATGTTTTAAAAGAA | tdc | [48] |
| p0303 | CCACTGCTGCATCTGTTTG | tdc | [118] |
| TD5 | CAAATGGAAGAAGAAGTAGG | tdc | [118] |
| TD2 | ACATAGTCAACCATRTTGAA | tdc | [118] |
| 57 | ATGAGTGAATCATTGTCG | tdc | [119] |
| 58 | TTATTTTGCTTCGCTTGCC | tdc | [119] |
| TDC1 | AACTATCGTATGGATATCAACG | tdc | [120] |
| TDC2 | TAGTCAACCATATTGAAATCTGG | tdc | [120] |
| TDC-F | TGGYTNGTNCCNCARACNAARCAYTA | tdc | [114] |
| TDC-R | ACRTARTCNACCATRTTRAARTCNGG | tdc | [114] |
K = G or T; R = A or G; W = A or T; Y = C or T; S = C or G; M = A or C; D = A, G, or T; N = A, G, C, or T.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Costantini, A.; Vaudano, E.; Pulcini, L.; Carafa, T.; Garcia-Moruno, E. An Overview on Biogenic Amines in Wine. Beverages 2019, 5, 19. https://doi.org/10.3390/beverages5010019
AMA Style
Costantini A, Vaudano E, Pulcini L, Carafa T, Garcia-Moruno E. An Overview on Biogenic Amines in Wine. Beverages. 2019; 5(1):19. https://doi.org/10.3390/beverages5010019
Chicago/Turabian StyleCostantini, Antonella, Enrico Vaudano, Laura Pulcini, Tommaso Carafa, and Emilia Garcia-Moruno. 2019. "An Overview on Biogenic Amines in Wine" Beverages 5, no. 1: 19. https://doi.org/10.3390/beverages5010019
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.