
Characterization of Microbial Communities and Naturally Occurring Radionuclides in Soilless Growth Media Amended with Different Concentrations of Biochar

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Collection Site Description
2.2. Media Composition
2.3. Sample Analysis
2.3.1. pH Determination
2.3.2. Nuclear Magnetic Resonance Analysis (NMR)
2.3.3. Multi-Elemental Scanning Thermal Analysis (MESTA)
2.3.4. DNA Extraction, Quantification, and Purity, Metagenomics
2.3.5. Radionuclides Sample Preparation and Analysis
2.3.6. 235U, 226Ra, 232Th, 40K and 137Cs
2.3.7. Metagenomic Sequence Accession Numbers
2.3.8. Statistical Analysis
3. Results
3.1. Samples’ Basic Physicochemical Characteristics
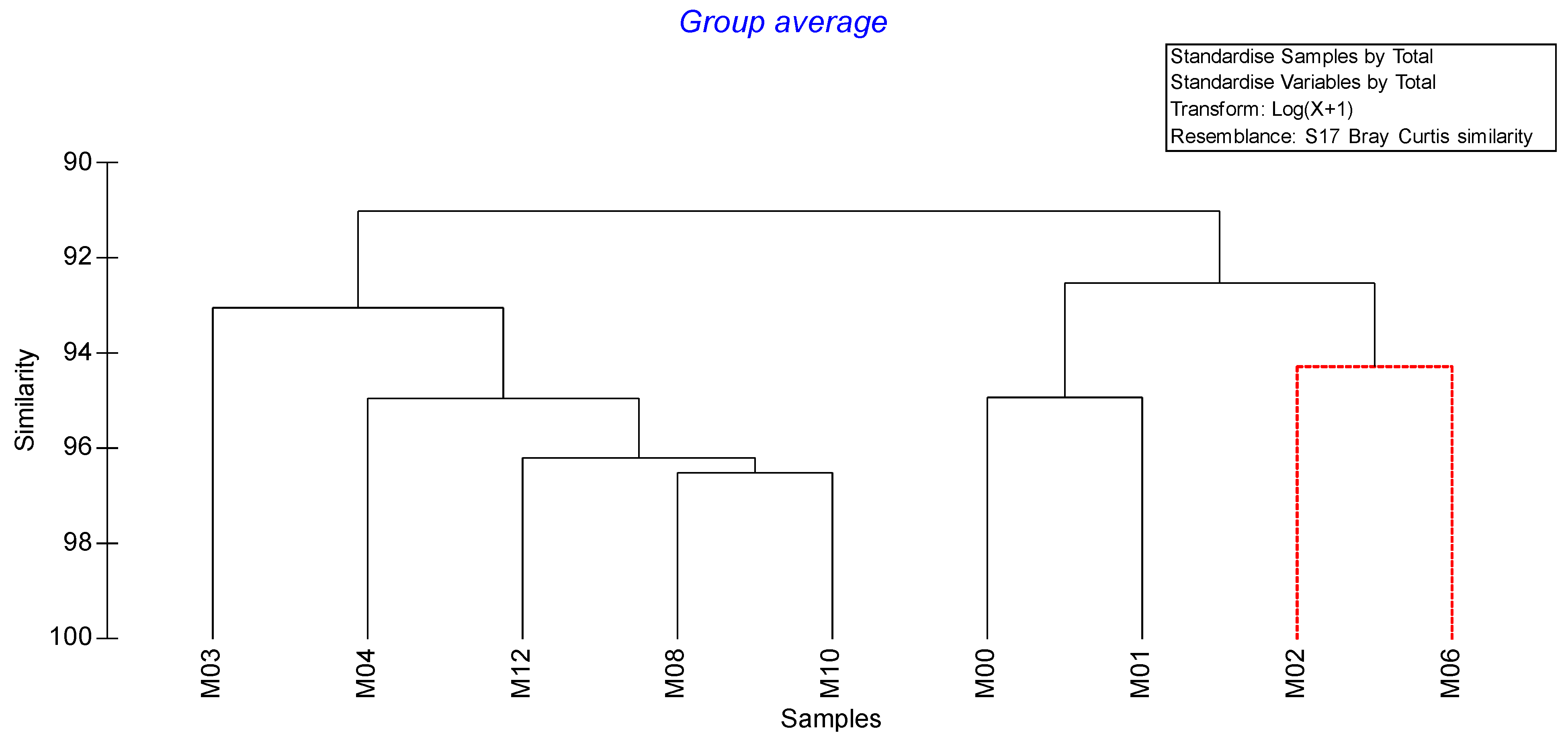
3.2. Media Microbial Composition
3.3. Radioactivity Measurements
3.4. Media Physicochemical Properties Relationship with Bacterial Composition and Radionuclides Activity Concentrations
3.5. Samples Bacteria Composition Relationship with Radionuclides Contents
4. Discussion
5. Conclusions and Future Research
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vaughn, S.F.; Kenar, J.A.; Thompson, A.R.; Peterson, S.C. Comparison of biochars derived from wood pellets and pelletized wheat straw as replacements for peat in potting substrates. Ind. Crop. Prod. 2013, 51, 437–443. [Google Scholar] [CrossRef]
- Gruda, N.S. Increasing Sustainability of Growing Media Constituents and Stand-Alone Substrates in Soilless Culture Systems. Agronomy 2019, 9, 298. [Google Scholar] [CrossRef]
- Barrett, G.E.; Alexander, P.D.; Robinson, J.S.; Bragg, N.C. Achieving environmentally sustainable growing media for soilless plant cultivation systems–A review. Sci. Hortic. 2016, 212, 220–234. [Google Scholar] [CrossRef]
- Jahromi, N.B.; Walker, F.; Fulcher, A.; Altland, J.; Wright, W.C. Growth response, mineral nutrition, and water utilization of con-tainer-grown woody ornamentals grown in biochar-amended pine bark. HortScience 2018, 53, 347–353. [Google Scholar]
- Graber, E.R.; Harel, Y.M.; Kolton, M.; Cytryn, E.; Silber, A.; David, D.R.; Tsechansky, L.; Borenshtein, M.; Elad, Y. Biochar impact on development and productivity of pepper and tomato grown in fertigated soilless media. Plant Soil 2010, 337, 481–496. [Google Scholar] [CrossRef]
- Ahmad, Z.; Mosa, A.; Zhan, L.; Gao, B. Biochar modulates mineral nitrogen dynamics in soil and terrestrial ecosystems: A critical review. Chemosphere 2021, 278, 130378. [Google Scholar] [CrossRef]
- Palansooriya, K.N.; Wong, J.T.F.; Hashimoto, Y.; Huang, L.; Rinklebe, J.; Chang, S.X.; Bolan, N.; Wang, H.; Ok, Y.S. Response of microbial communities to biochar-amended soils: A critical review. Biochar 2019, 1, 3–22. [Google Scholar] [CrossRef]
- Pérez-Guzmán, L.; Lower, B.H.; Dick, R.P. Corn and hardwood biochars affected soil microbial community and enzyme activities. Agrosystems Geosci. Environ. 2020, 3, e20082. [Google Scholar] [CrossRef]
- Sohi, S.P.; Krull, E.; Lopez-Capel, E.; Bol, R. A review of biochar and its use and function in soil. Adv. Agron. 2010, 105, 47–82. [Google Scholar] [CrossRef]
- Saul-Tcherkas, V.; Steinberger, Y. Substrate utilization patterns of desert soil microbial communities in response to xeric and mesic conditions. Soil Biol. Biochem. 2009, 41, 1882–1893. [Google Scholar] [CrossRef]
- Waghmode, T.R.; Chen, S.; Li, J.; Sun, R.; Liu, B.; Hu, C. Response of Nitrifier and Denitrifier Abundance and Microbial Community Structure to Experimental Warming in an Agricultural Ecosystem. Front. Microbiol. 2018, 9, 474. [Google Scholar] [CrossRef] [PubMed]
- Ligi, T.; Oopkaup, K.; Truu, M.; Preem, J.-K.; Nõlvak, H.; Mitsch, W.J.; Mander, Ü.; Truu, J. Characterization of bacterial communities in soil and sediment of a created riverine wetland complex using high-throughput 16S rRNA amplicon sequencing. Ecol. Eng. 2014, 72, 56–66. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Chang, S.X.; Yang, Y.; Fu, S.; Jiang, P.; Luo, Y.; Yang, M.; Chen, Z.; Hu, S.; et al. Biochar reduces soil heterotrophic respi-ration in a subtropical plantation through increasing soil organic carbon recalcitrancy and decreasing carbon-degrading mi-crobial activity. Soil Biol. Biochem. 2018, 122, 173–185. [Google Scholar]
- Khandaker, M.U.; Jojo, P.J.; Abu Kassim, H. Determination of Primordial Radionuclides in Natural Samples Using HPGe Gamma-Ray Spectrometry. APCBEE Procedia 2012, 1, 187–192. [Google Scholar] [CrossRef]
- Shukla, A.; Parmar, P.; Saraf, M. Radiation, radionuclides and bacteria: An in-perspective review. J. Environ. Radioact. 2017, 180, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Theodorakopoulos, N.; Février, L.; Barakat, M.; Ortet, P.; Christen, R.; Piette, L.; Levchuk, S.; Beaugelin-Seiller, K.; Sergeant, C.; Berthomieu, C.; et al. Soil prokaryotic communities in Chernobyl waste disposal trench T22 are modulated by organic matter and radionuclide contamination. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef]
- Salbu, B.; Skipperud, L.; Lind, O.C. Sources contributing to radionuclides in the environment: With focus on radioactive particles. In Radionuclides in the Environment 2015; Springer: Cham, Switzerland, 2015; pp. 1–36. [Google Scholar]
- Hu, Q.-H.; Weng, J.-Q.; Wang, J.-S. Sources of anthropogenic radionuclides in the environment: A review. J. Environ. Radioact. 2010, 101, 426–437. [Google Scholar] [CrossRef]
- Correa, R.; Miranda, P.; Ortiz-Ramirez, P.; Wachter, J.; Camilla, S.; Mera, E.; Piñones, E. Activity concentration of NORM and 137Cs radionuclide in soil samples from the Andes Cordillera at latitude 33°56′ South. J. Physics Conf. Ser. 2018, 1043, 012028. [Google Scholar] [CrossRef]
- Lloyd, J.R.; Gadd, G.M. The geomicrobiology of radionuclides. Geomicrobiol. J. 2011, 28, 383–386. [Google Scholar]
- Belimov, A.A.; Kunakova, A.M.; Vasilyeva, N.D.; Kovatcheva, T.S.; Dritchko, V.F.; Kuzovatov, S.N.; Trushkina, I.R.; Alekseyev, Y.U. Accumulation of radionuclides by associative bacteria and the uptake of 134 Cs by the inoculated barley plants. In Nitrogen Fixation with Non-Legumes 1998; Springer: Dordrecht, The Netherlands, 1998; pp. 275–280. [Google Scholar]
- Yan, X.; Luo, X. Radionuclides distribution, properties, and microbial diversity of soils in uranium mill tailings from southeastern China. J. Environ. Radioact. 2015, 139, 85–90. [Google Scholar] [CrossRef]
- Newsome, L.; Morris, K.; Lloyd, J.R. The biogeochemistry and bioremediation of uranium and other priority radionuclides. Chem. Geol. 2013, 363, 164–184. [Google Scholar] [CrossRef]
- Merroun, M.L.; Selenska-Pobell, S. Bacterial interactions with uranium: An environmental perspective. J. Contam. Hydrol. 2008, 102, 285–295. [Google Scholar] [CrossRef]
- Cáliz, J.; Montserrat, G.; Martí, E.; Sierra, J.; Chung, A.P.; Morais, P.V.; Vila, X. Emerging resistant microbiota from an acidic soil exposed to toxicity of Cr, Cd and Pb is mainly influenced by the bioavailability of these metals. J. Soils Sediments 2012, 13, 413–428. [Google Scholar] [CrossRef]
- Wolman, M.G. Population, land use, and environment: A long history. In Population and Land Use in Developing Countries; The National Academies Press: Washington, DC, USA, 1993. [Google Scholar]
- Camill, P. Global change. Nat. Educ. Knowl. 2010, 3, 49. [Google Scholar]
- Assessment, G.F. Main Report; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010. [Google Scholar]
- Huang, L.; Gu, M. Effects of Biochar on Container Substrate Properties and Growth of Plants—A Review. Horticulturae 2019, 5, 14. [Google Scholar] [CrossRef]
- Méndez, A.; Cárdenas-Aguiar, E.; Paz-Ferreiro, J.; Plaza, C.; Gascó, G. The effect of sewage sludge biochar on peat-based growing media. Biol. Agric. Hortic. 2016, 33, 40–51. [Google Scholar] [CrossRef]
- Ngatia, L.; Hsieh, Y.; Nemours, D.; Fu, R.; Taylor, R. Potential phosphorus eutrophication mitigation strategy: Biochar carbon composition, thermal stability and pH influence phosphorus sorption. Chemosphere 2017, 180, 201–211. [Google Scholar] [CrossRef]
- Rajkovich, S.; Enders, A.; Hanley, K.; Hyland, C.; Zimmerman, A.R.; Lehmann, J. Corn growth and nitrogen nutrition after additions of biochars with varying properties to a temperate soil. Biol. Fertil. Soils 2011, 48, 271–284. [Google Scholar] [CrossRef]
- Yao, Y.; Gao, B.; Zhang, M.; Inyang, M.; Zimmerman, A.R. Effect of biochar amendment on sorption and leaching of nitrate, ammonium, and phosphate in a sandy soil. Chemosphere 2012, 89, 1467–1471. [Google Scholar] [CrossRef]
- Knicker, H. Solid state CPMAS 13C and 15N NMR spectroscopy in organic geochemistry and how spin dynamics can either aggravate or improve spectra interpretation. Org. Geochem. 2011, 42, 867–890. [Google Scholar]
- Hsieh, Y.P.; Bugna, G.C. Analysis of black carbon in sediments and soils using multi-element scanning thermal analysis (MESTA). Org. Geochem. 2008, 39, 1562–1571. [Google Scholar] [CrossRef]
- Hsieh, Y.-P. A novel multielemental scanning thermal analysis (MESTA) method for the identification and characterization of solid substances. J. AOAC Int. 2007, 90. [Google Scholar]
- Disnar, J.R.; Jacob, J.; Morched-Issa, M.; Lottier, N.; Arnaud, F. Assessment of peat quality by molecular and bulk geochemical analysis: Application to the Holocene record of the Chautagne marsh (Haute Savoie, France). Chem. Geol. 2008, 254, 101–112. [Google Scholar] [CrossRef]
- Ning, J.; Liebich, J.; Kästner, M.; Zhou, J.; Schäffer, A.; Burauel, P. Different influences of DNA purity indices and quantity on PCR-based DGGE and functional gene microarray in soil microbial community study. Appl. Microbiol. Biotechnol. 2009, 82, 983–993. [Google Scholar] [CrossRef]
- Jaswal, R.; Pathak, A.; Chauhan, A. Metagenomic Evaluation of Bacterial and Fungal Assemblages Enriched within Diffusion Chambers and Microbial Traps Containing Uraniferous Soils. Microorganisms 2019, 7, 324. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2013, 30, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Glöckner, F.O.; Yilmaz, P.; Quast, C.; Gerken, J.; Beccati, A.; Ciuprina, A.; Bruns, G.; Yarza, P.; Peplies, J.; Westram, R.; et al. 25 years of serving the community with ribosomal RNA gene reference databases and tools. J. Biotechnol. 2017, 261, 169–176. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Tikhonov, M.; Leach, R.W.; Wingreen, N.S. Interpreting 16S metagenomic data without clustering to achieve sub-OTU resolution. ISME J. 2014, 9, 68–80. [Google Scholar] [CrossRef]
- Jabbar, A.; Arshed, W.; Bhatti, A.S.; Ahmad, S.S.; Akhter, P.; Rehman, S.-U.; Anjum, M.I. Measurement of soil radioactivity levels and radiation hazard assessment in southern Rechna interfluvial region, Pakistan. Environ. Monit. Assess. 2009, 169, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Hilal, M.; Attallah, M.; Mohamed, G.Y.; Fayez-Hassan, M. Evaluation of radiation hazard potential of TENORM waste from oil and natural gas production. J. Environ. Radioact. 2014, 136, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Völgyesi, P.; Kis, Z.; Szabo, Z.; Szabó, C. Using the 186-keV peak for 226Ra activity concentration determination in Hungarian coal-slag samples by gamma-ray spectroscopy. J. Radioanal. Nucl. Chem. Artic. 2014, 302, 375–383. [Google Scholar] [CrossRef]
- Bikit, I.; Forkapic, S.; Nikolov, J.; Todorovic, N.; Mrdja, D. Radioactivity of the agricultural soil in northern province of Serbia, Vojvodina. Int. J. Environ. Ecol. Eng. 2011, 5, 232–237. [Google Scholar]
- Alnour, I.A.; Ibrahim, N.; Hossain, I. Concentrations of 214 Pb, 214 Bi in 238 U series and 208 Tl, 228 Ac in 232 Th series in granite rock in (Kadugli) Sudan. Available online: http://nopr.niscpr.res.in/bitstream/123456789/13996/1/IJPAP%2050%285%29%20285-288.pdf (accessed on 25 June 2022).
- Papp, Z.; Dezső, Z.; Daroczy, S. Measurement of the radioactivity of238U, 232Th, 226Ra, 137Cs and40K in soil using direct Ge (Li) γ-ray spectrometry. J. Radioanal. Nucl. Chem. 1997, 222, 171–176. [Google Scholar] [CrossRef]
- Thies, J.E.; Rillig, M.C.; Graber, E.R. Biochar effects on the abundance, activity and diversity of the soil biota. Biochar Environ. Manag. Sci. Technol. Implement. 2015, 2, 327–389. [Google Scholar]
- Husson, O. Redox potential (Eh) and pH as drivers of soil/plant/microorganism systems: A transdisciplinary overview pointing to integrative opportunities for agronomy. Plant Soil 2012, 362, 389–417. [Google Scholar] [CrossRef]
- Chen, J.; Liu, X.; Zheng, J.; Zhang, B.; Lu, H.; Chi, Z.; Pan, G.; Li, L.; Zheng, J.; Zhang, X.; et al. Biochar soil amendment increased bacterial but decreased fungal gene abundance with shifts in community structure in a slightly acid rice paddy from Southwest China. Appl. Soil Ecol. 2013, 71, 33–44. [Google Scholar] [CrossRef]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef]
- Rampelotto, P.H.; Ferreira, A.D.S.; Barboza, A.D.M.; Roesch, L.F.W. Changes in Diversity, Abundance, and Structure of Soil Bacterial Communities in Brazilian Savanna Under Different Land Use Systems. Microb. Ecol. 2013, 66, 593–607. [Google Scholar] [CrossRef]
- Acosta-Martínez, V.; Dowd, S.; Sun, Y.; Allen, V. Tag-encoded pyrosequencing analysis of bacterial diversity in a single soil type as affected by management and land use. Soil Biol. Biochem. 2008, 40, 2762–2770. [Google Scholar] [CrossRef]
- Naether, A.; Foesel, B.U.; Naegele, V.; Wüst, P.K.; Weinert, J.; Bonkowski, M.; Alt, F.; Oelmann, Y.; Polle, A.; Lohaus, G.; et al. Environmental factors affect acidobacterial communities below the subgroup level in grassland and forest soils. Appl. Environ. Microbiol. 2012, 78, 7398–7406. [Google Scholar] [CrossRef]
- Nacke, H.; Thürmer, A.; Wollherr, A.; Will, C.; HoDac, L.; Herold, N.; Schöning, I.; Schrumpf, M.; Daniel, R. Pyrosequencing-Based Assessment of Bacterial Community Structure Along Different Management Types in German Forest and Grassland Soils. PLoS ONE 2011, 6, e17000. [Google Scholar] [CrossRef]
- Zheng, J.; Chen, J.; Pan, G.; Liu, X.; Zhang, X.; Li, L.; Bian, R.; Cheng, K.; Jinwei, Z. Biochar decreased microbial metabolic quotient and shifted community composition four years after a single incorporation in a slightly acid rice paddy from southwest China. Sci. Total Environ. 2016, 571, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Delgado-Baquerizo, M.; Anderson, I.C.; Singh, B.K. Response of Soil Properties and Microbial Communities to Agriculture: Implications for Primary Productivity and Soil Health Indicators. Front. Plant Sci. 2016, 7, 990. [Google Scholar] [CrossRef]
- Nielsen, S.; Minchin, T.; Kimber, S.; Van Zwieten, L.; Gilbert, J.; Munroe, P.; Joseph, S.; Thomas, T. Comparative analysis of the microbial communities in agricultural soil amended with enhanced biochars or traditional fertilisers. Agric. Ecosyst. Environ. 2014, 191, 73–82. [Google Scholar] [CrossRef]
- Lee, S.H.; Ka, J.O.; Cho, J.C. Members of the phylum Acidobacteria are dominant and metabolically active in rhizosphere soil. FEMS Microbiol. Lett. 2008, 285, 263–269. [Google Scholar] [CrossRef]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota–a review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Kirby, R. Actinomycetes and Lignin Degradation. Adv. Appl. Microbiol. 2005, 58, 125–168. [Google Scholar] [CrossRef]
- Xu, N.; Tan, G.; Wang, H.; Gai, X. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Leff, J.W.; Jones, S.E.; Prober, S.M.; Barberán, A.; Borer, E.T.; Firn, J.L.; Harpole, W.S.; Hobbie, S.E.; Hofmockel, K.S.; Knops, J.M.H.; et al. Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proc. Natl. Acad. Sci. USA 2015, 112, 10967–10972. [Google Scholar] [CrossRef] [PubMed]
- Yadav, A.N.; Verma, P.; Kumar, S.; Kumar, V.; Kumar, M.; Sugitha, T.C.; Singh, B.P.; Saxena, A.K.; Dhaliwal, H.S. Actinobacteria from rhizosphere: Molecular diversity, distributions, and potential biotechnological applications. In New and Future Developments in Microbial Biotechnology and Bioengineering 2018 Jan 1; Elsevier: Amsterdam, The Netherlands, 2018; pp. 13–41. [Google Scholar]
- Ventura, M.; Canchaya, C.; Tauch, A.; Chandra, G.; Fitzgerald, G.F.; Chater, K.F.; van Sinderen, D. Genomics of Actinobacteria: Tracing the Evolutionary History of an Ancient Phylum. Microbiol. Mol. Biol. Rev. 2007, 71, 495–548. [Google Scholar] [CrossRef]
- Huang, L.N.; Zhu, S.; Zhou, H.; Qu, L.H. Molecular phylogenetic diversity of bacteria associated with the leachate of a closed municipal solid waste landfill. FEMS Microbiol. Lett. 2005, 242, 297–303. [Google Scholar] [PubMed]
- Singh, H.; Khattar, J.S.; Ahluwalia, A.S. Cyanobacteria and agricultural crops. Vegetos 2014, 27, 37. [Google Scholar] [CrossRef]
- Rana, A.; Joshi, M.; Prasanna, R.; Shivay, Y.S.; Nain, L. Biofortification of wheat through inoculation of plant growth promoting rhizobacteria and cyanobacteria. Eur. J. Soil Biol. 2012, 50, 118–126. [Google Scholar] [CrossRef]
- Strauss, G.; Fuchs, G. Enzymes of a novel autotrophic CO2 fixation pathway in the phototrophic bacterium Chloroflexus aurantiacus, the 3-hydroxypropionate cycle. Eur. J. Biochem. 1993, 215, 633–643. [Google Scholar]
- Kolton, M.; Harel, Y.M.; Pasternak, Z.; Graber, E.R.; Elad, Y.; Cytryn, E. Impact of Biochar Application to Soil on the Root-Associated Bacterial Community Structure of Fully Developed Greenhouse Pepper Plants. Appl. Environ. Microbiol. 2011, 77, 4924–4930. [Google Scholar] [CrossRef]
- McBride, M.J.; Xie, G.; Martens, E.C.; Lapidus, A.; Henrissat, B.; Rhodes, R.G.; Goltsman, E.; Wang, W.; Xu, J.; Hunnicutt, D.W.; et al. Novel Features of the Polysaccharide-Digesting Gliding Bacterium Flavobacterium johnsoniae as Revealed by Genome Sequence Analysis. Appl. Environ. Microbiol. 2009, 75, 6864–6875. [Google Scholar] [CrossRef]
- Pérez-Jaramillo, J.E.; De Hollander, M.; Ramírez, C.A.; Mendes, R.; Raaijmakers, J.M.; Carrión, V.J. Deciphering rhizosphere microbiome assembly of wild and modern common bean (Phaseolus vulgaris) in native and agricultural soils from Colombia. Microbiome 2019, 7, 114. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, A.K.; Elad, Y.; Paudel, I.; Graber, E.R.; Cytryn, E.; Frenkel, O. Linking the Belowground Microbial Composition, Diversity and Activity to Soilborne Disease Suppression and Growth Promotion of Tomato Amended with Biochar. Sci. Rep. 2017, 7, srep44382. [Google Scholar] [CrossRef]
- Vinson, D.S.; Vengosh, A.; Hirschfeld, D.; Dwyer, G.S. Relationships between radium and radon occurrence and hydrochemistry in fresh groundwater from fractured crystalline rocks, North Carolina (USA). Chem. Geol. 2008, 260, 159–171. [Google Scholar] [CrossRef]
- Szabo, Z.; DePaul, V.T.; Kraemer, T.F.; Parsa, B. Occurrence of Radium-224, Radium-226, and Radium-228 in Water of the Unconfined Kirkwood-Cohansey Aquifer System, Southern New Jersey; U. S. Geological Survey: Reston, VA, USA, 2005. [Google Scholar]
- Gaspar, L.; Lizaga, I.; Navas, A. Spatial distribution of fallout and lithogenic radionuclides controlled by soil carbon and water erosion in an agroforestry South-Pyrenean catchment. Geoderma 2021, 391, 114941. [Google Scholar] [CrossRef]
- Lopez-Fernandez, M.; Jroundi, F.; Ruiz-Fresneda, M.A.; Merroun, M.L. Microbial interaction with and tolerance of radionuclides: Underlying mechanisms and biotechnological applications. Microb. Biotechnol. 2020, 14, 810–828. [Google Scholar] [CrossRef]
- Kumar, V.; Chandra, R. Bacteria-assisted phytoremediation of industrial waste pollutants and ecorestoration. In Phytoremediation of Environmental Pollutants 2017 Dec 14; CRC Press: Boca Raton, FL, USA, 2017; pp. 159–200. [Google Scholar]
- Suriya, J.; Shekar, M.C.; Nathani, N.M.; Suganya, T.; Bharathiraja, S.; Krishnan, M. Assessment of bacterial community composition in response to uranium levels in sediment samples of sacred Cauvery River. Appl. Microbiol. Biotechnol. 2016, 101, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Hoyos-Hernandez, C.; Courbert, C.; Simonucci, C.; David, S.; Vogel, T.M.; Larose, C. Community structure and functional genes in radionuclide contaminated soils in Chernobyl and Fukushima. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef]
- Barns, S.M.; Cain, E.C.; Sommerville, L.; Kuske, C.R. Acidobacteria phylum sequences in uranium-contaminated subsurface sediments greatly expand the known diversity within the phylum. Appl. Environ. Microbiol. 2007, 73, 3113–3116. [Google Scholar]
- Mondani, L.; Benzerara, K.; Carrière, M.; Christen, R.; Mamindy-Pajany, Y.; Février, L.; Marmier, N.; Achouak, W.; Nardoux, P.; Berthomieu, C.; et al. Influence of Uranium on Bacterial Communities: A Comparison of Natural Uranium-Rich Soils with Controls. PLoS ONE 2011, 6, e25771. [Google Scholar] [CrossRef]
- Hug, L.A.; Castelle, C.J.; Wrighton, K.C.; Thomas, B.C.; Sharon, I.; Frischkorn, K.R.; Williams, K.H.; Tringe, S.G.; Banfield, J.F. Community genomic analyses constrain the distribution of metabolic traits across the Chloroflexi phylum and indicate roles in sediment carbon cycling. Microbiome 2013, 1, 22. [Google Scholar] [CrossRef]
- Banach, A.M.; Kuźniar, A.; Grządziel, J.; Wolińska, A. Azolla filiculoides L. as a source of metal-tolerant microorganisms. PLoS ONE 2020, 15, e0232699. [Google Scholar] [CrossRef]
- Shilpi, G.; Shilpi, S.; Sunita, S. Tolerance against heavy metal toxicity in cyanobacteria: Role of antioxidant defense system. Int. J. Pharm. Pharm. Sci. 2015, 7, 1–8. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biochar | pH | TC | Aromatic | Phenolic | Carboxyl | R400 |
|---|---|---|---|---|---|---|
| 0% | 6.15 | 443.67 | 67.76 | 32.17 | 14.89 | 0.61 |
| 1% | 6.02 | 447.80 | 67.96 | 39.82 | 17.48 | 0.70 |
| 2% | 6.03 | 443.03 | 65.66 | 34.72 | 14.58 | 0.65 |
| 3% | 6.23 | 454.60 | 73.98 | 42.32 | 27.72 | 0.67 |
| 4% | 6.47 | 470.17 | 78.04 | 42.40 | 23.06 | 0.65 |
| 6% | 6.46 | 459.97 | 73.88 | 42.24 | 20.59 | 0.62 |
| 8% | 6.68 | 478.40 | 82.16 | 48.34 | 25.40 | 0.60 |
| 10% | 6.76 | 492.43 | 92.60 | 53.34 | 26.38 | 0.59 |
| 12% | 6.84 | 514.47 | 95.59 | 56.11 | 38.29 | 0.50 |
| Biochar | 235U | 226Ra | 232Th | 40K | 137Cs |
|---|---|---|---|---|---|
| 0% | 0.92 ± 0.05 | 2.93 ± 0.12 | 1.01 ± 0.04 | 0.00 | 0.21 ± 0.02 |
| 1% | 0.92 ± 0.05 | 2.54 ± 0.11 | 0.68 ± 0.08 | 0.00 | 0.19 ± 0.02 |
| 2% | 0.99 ± 0.05 | 2.94 ± 0.12 | 1.02 ± 0.11 | 22.54 ± 0.54 | 0.22 ± 0.02 |
| 3% | 0.94 ± 0.05 | 3.10 ± 0.13 | 0.98 ± 0.18 | 21.10 ± 0.52 | 0.39 ± 0.03 |
| 4% | 0.92 ± 0.05 | 3.78 ± 0.14 | 0.57 ± 0.09 | 21.37 ± 0.53 | 0.37 ± 0.03 |
| 6% | 0.81 ± 0.04 | 2.89 ± 0.13 | 0.90 ± 0.04 | 19.57 ± 0.51 | 0.39 ± 0.03 |
| 8% | 0.90 ± 0.05 | 3.63 ± 0.14 | 0.88 ± 0.10 | 22.18 ± 0.54 | 0.48 ± 0.03 |
| 10% | 0.96 ± 0.05 | 3.93 ± 0.14 | 1.05 ± 0.10 | 24.23 ± 0.57 | 0.45 ± 0.02 |
| 12% | 0.94 ± 0.10 | 4.14 ± 0.29 | 1.02 ± 0.22 | 23.70 ± 1.13 | 0.55 ± 0.06 |
| 235U | 226Ra | 232Th | 40K | 137Cs | |
|---|---|---|---|---|---|
| 235U | 1.00 | 0.16 | 0.13 | −0.15 | |
| 226Ra | 0.23 | 1.00 | −0.13 | 0.67 | 0.83 |
| 232Th | 0.16 | −0.13 | 1.00 | 0.17 | 0.11 |
| 40K | 0.13 | 0.67 | 0.17 | 1.00 | 0.74 |
| 137Cs | −0.15 | 0.83 | 0.11 | 0.74 | 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osei, G.K.; Abazinge, M.; Ngatia, L.; Chauhan, A.; Bolques, A.; Jagoe, C.; Pathak, A. Characterization of Microbial Communities and Naturally Occurring Radionuclides in Soilless Growth Media Amended with Different Concentrations of Biochar. Appl. Microbiol. 2022, 2, 662-679. https://doi.org/10.3390/applmicrobiol2030051
Osei GK, Abazinge M, Ngatia L, Chauhan A, Bolques A, Jagoe C, Pathak A. Characterization of Microbial Communities and Naturally Occurring Radionuclides in Soilless Growth Media Amended with Different Concentrations of Biochar. Applied Microbiology. 2022; 2(3):662-679. https://doi.org/10.3390/applmicrobiol2030051
Chicago/Turabian StyleOsei, George K., Michael Abazinge, Lucy Ngatia, Ashvini Chauhan, Alejandro Bolques, Charles Jagoe, and Ashish Pathak. 2022. "Characterization of Microbial Communities and Naturally Occurring Radionuclides in Soilless Growth Media Amended with Different Concentrations of Biochar" Applied Microbiology 2, no. 3: 662-679. https://doi.org/10.3390/applmicrobiol2030051