Expression of the Tyrosine Hydroxylase Gene from Rat Leads to Oxidative Stress in Potato Plants

 , , , and
, , , and
Abstract
:
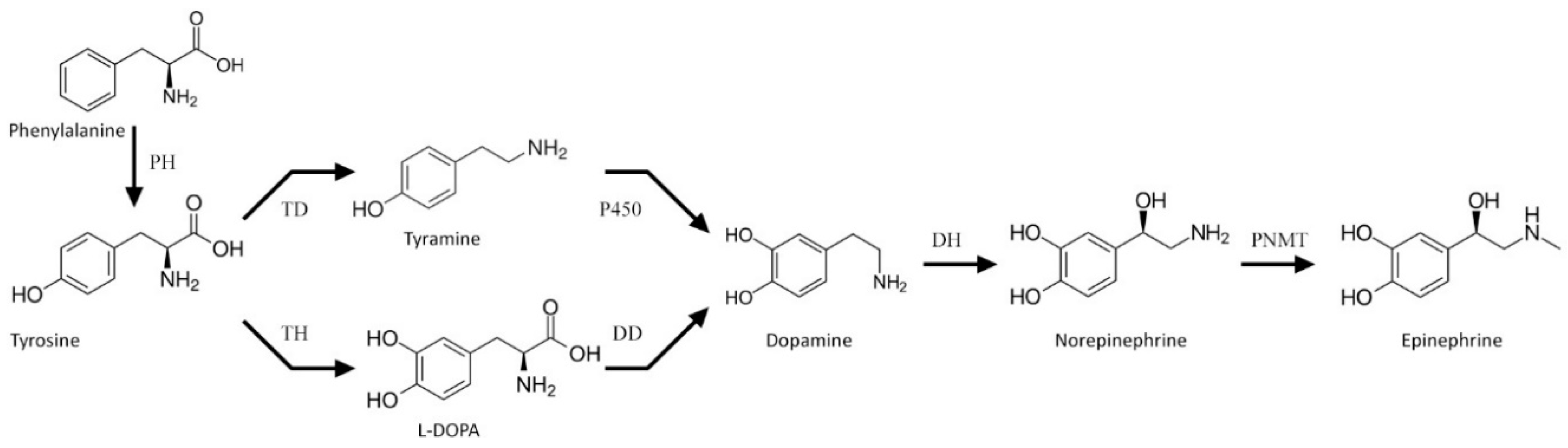
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Construction of Transgenic Potato Lines
2.3. Western Blot Analysis
2.4. Tyrosine Hydroxylase Activity Assay
2.5. Isolation and Analysis of Phenolics
2.6. Gas Chromatography–Mass Spectrometry (GC–MS) Analysis of Metabolome
2.7. Real-Time Polymerase Chain Reaction (RT-PCR)
2.8. Determination of Antioxidant Capacity
2.9. Hydrogen Peroxide Assay
2.10. Treatment of Plants with L-DOPA
2.11. Determination of Resistance to Pathogen
2.12. Statistical Analysis
3. Results
3.1. Construction of Transgenic Potato Plants
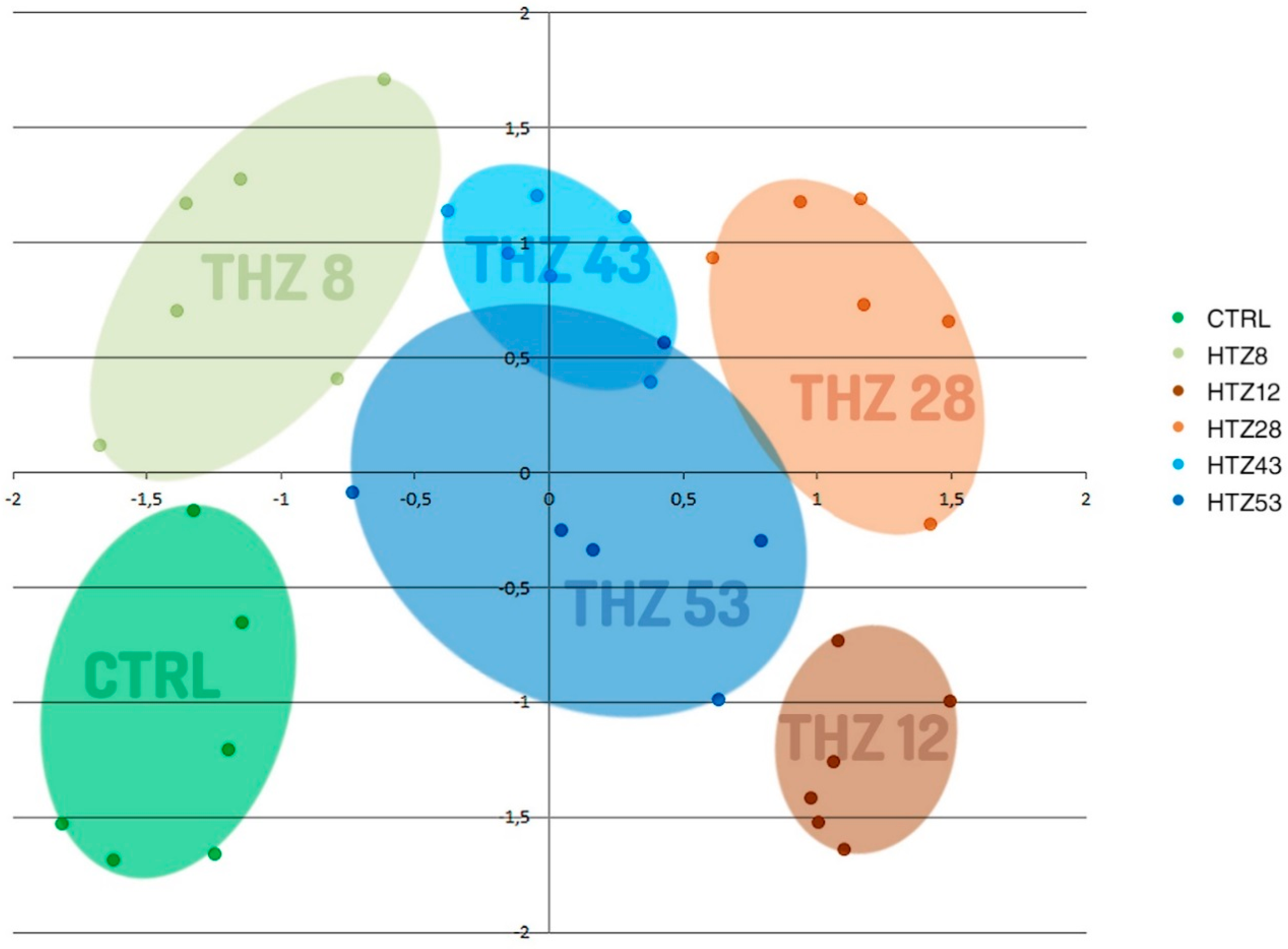
3.2. Metabolic Profile of the Transgenic Potato Plants
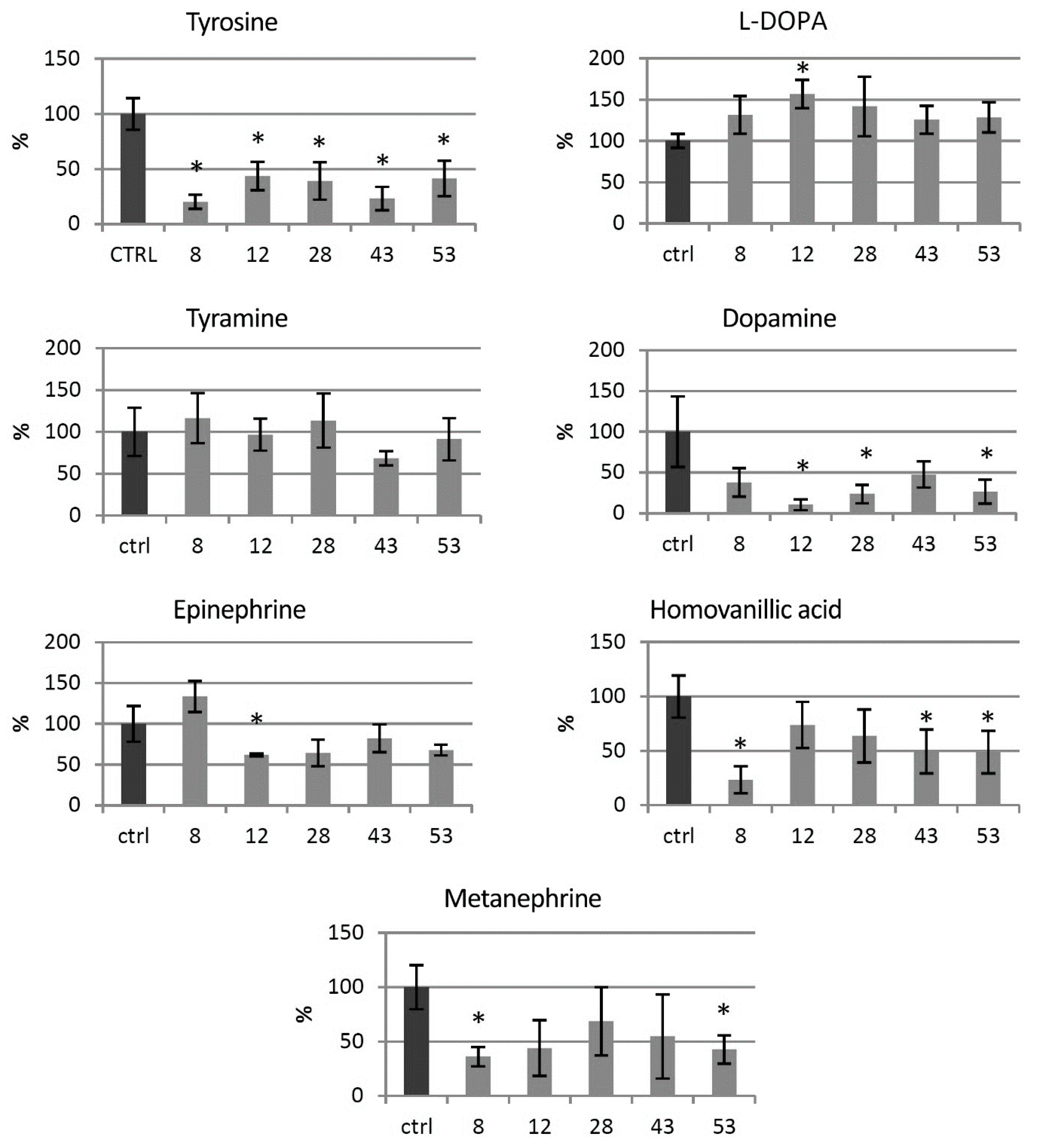
3.3. Catecholamine Level in the Transgenic HTZ Lines
3.4. Primary Metabolite Level in the Transgenic HTZ Lines
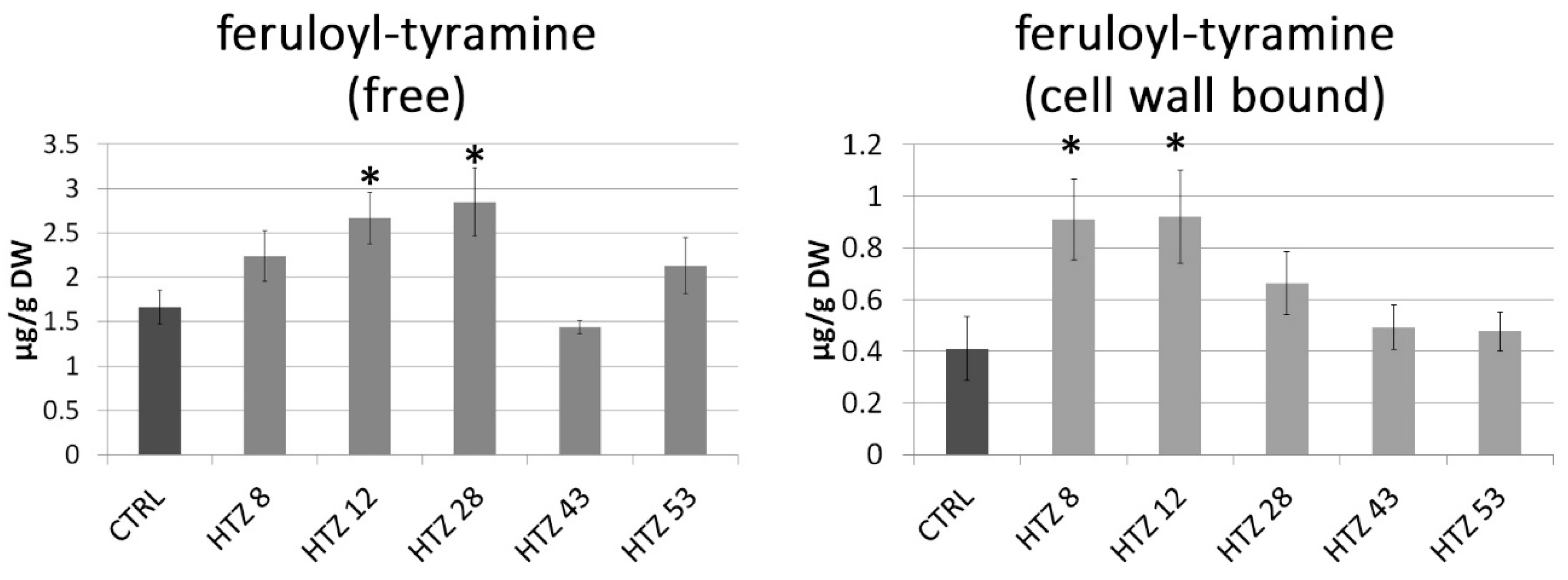
3.5. Phenolic Level in the Transgenic HTZ Lines
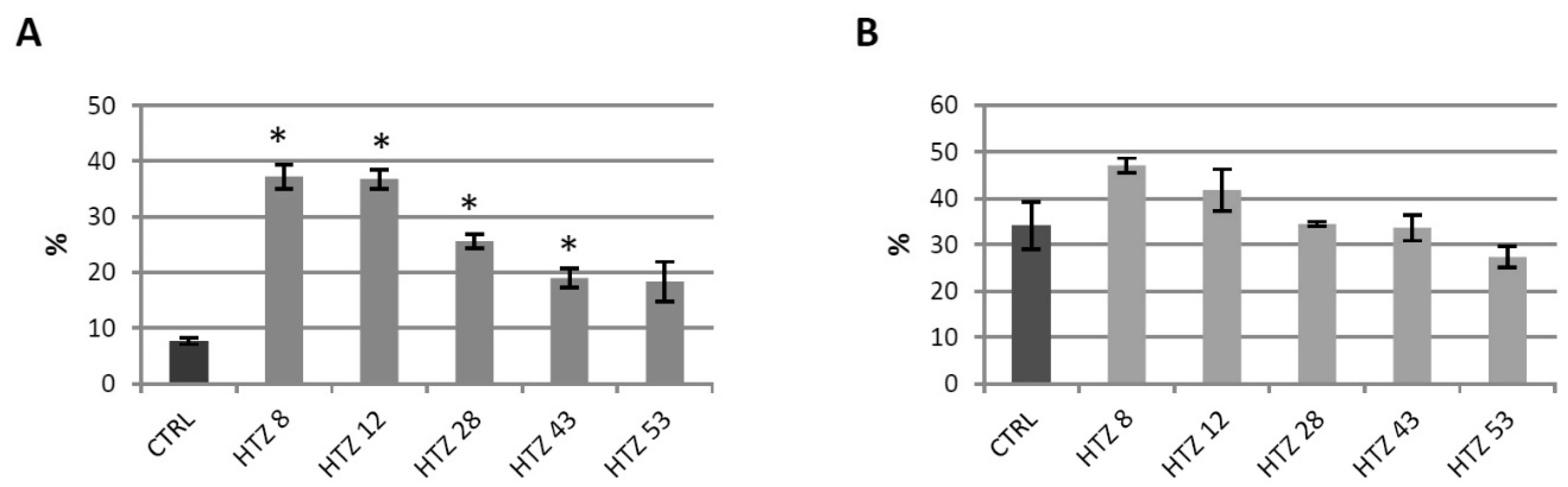
3.6. Antioxidant Potential of the Transgenic Potato Extracts
3.7. Red-ox Analysis in the Transgenic HTZ Lines
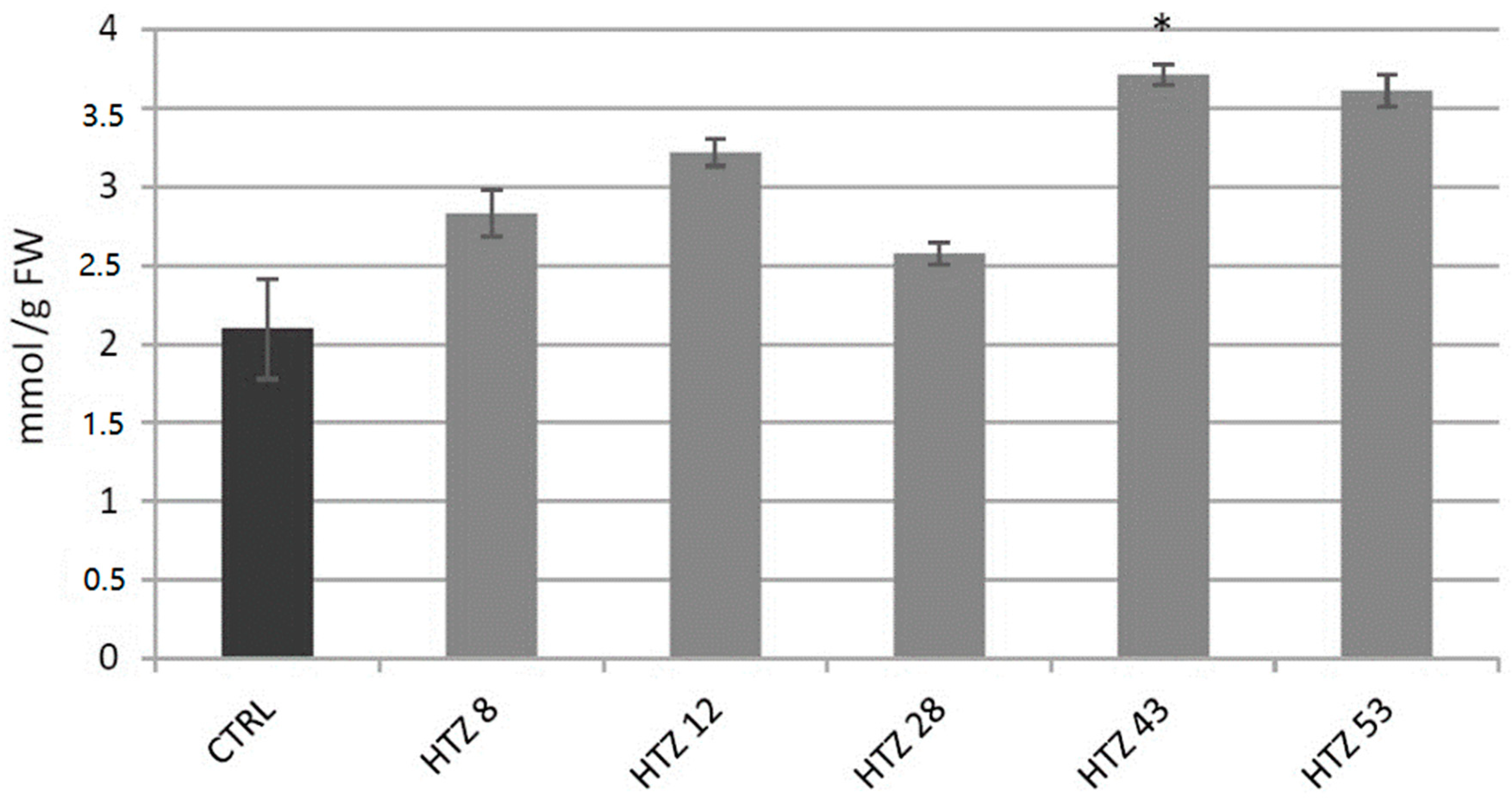
3.7.1. Analysis of H2O2 Level in Transgenic Potato Plants
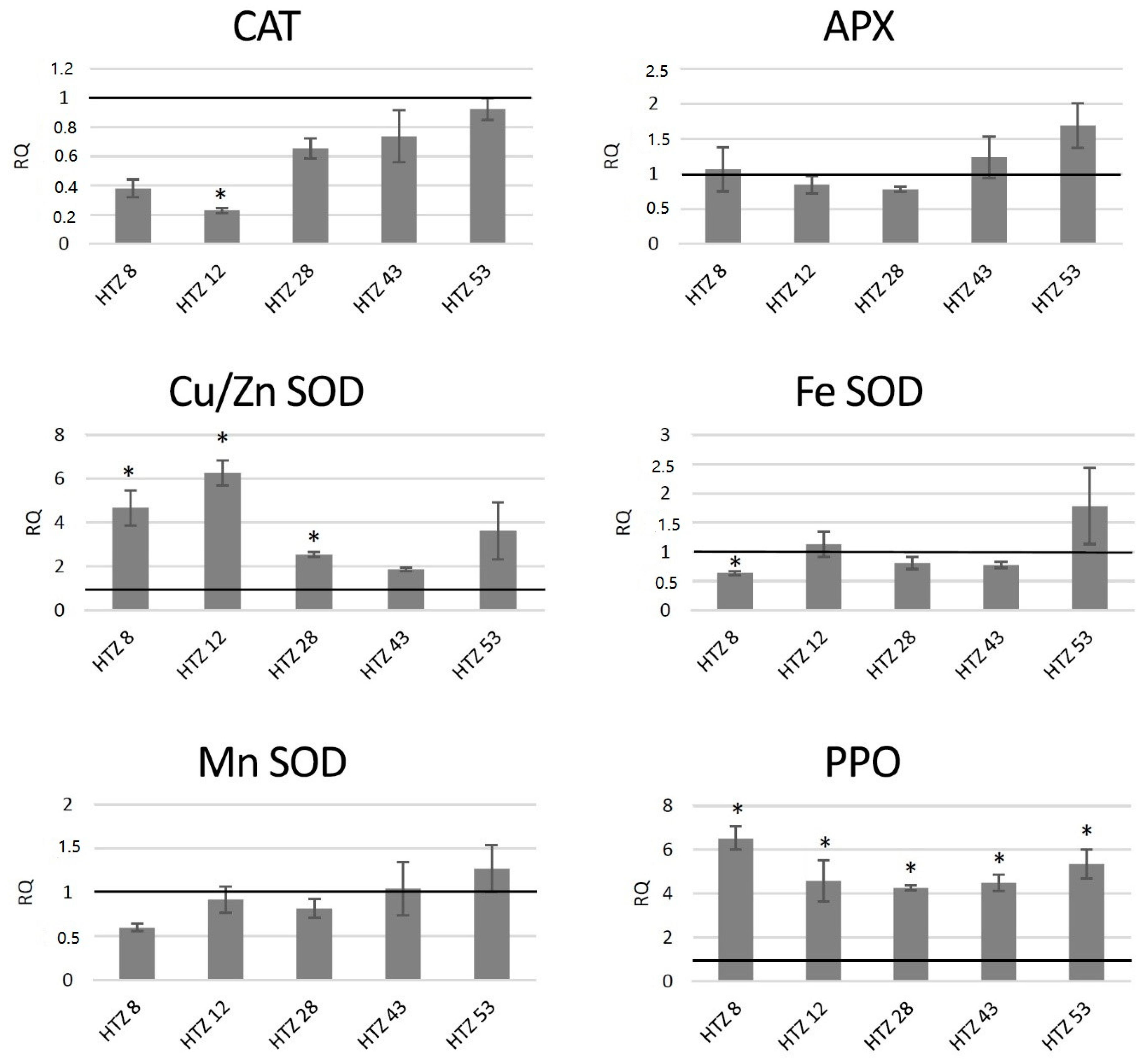
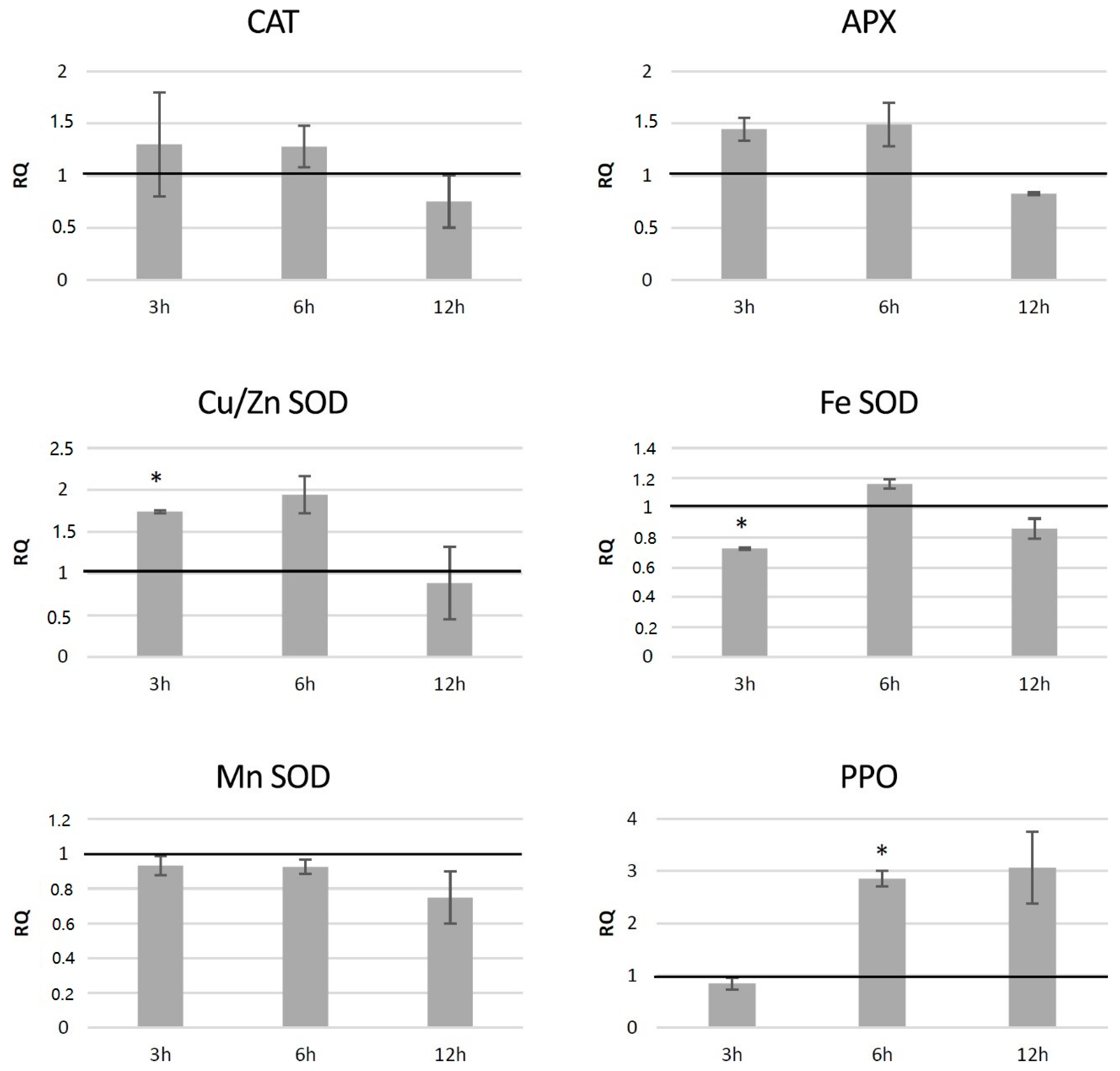
3.7.2. Analysis of Expression of Genes Involved in Free Radical Processing in Transgenic Potato Plants
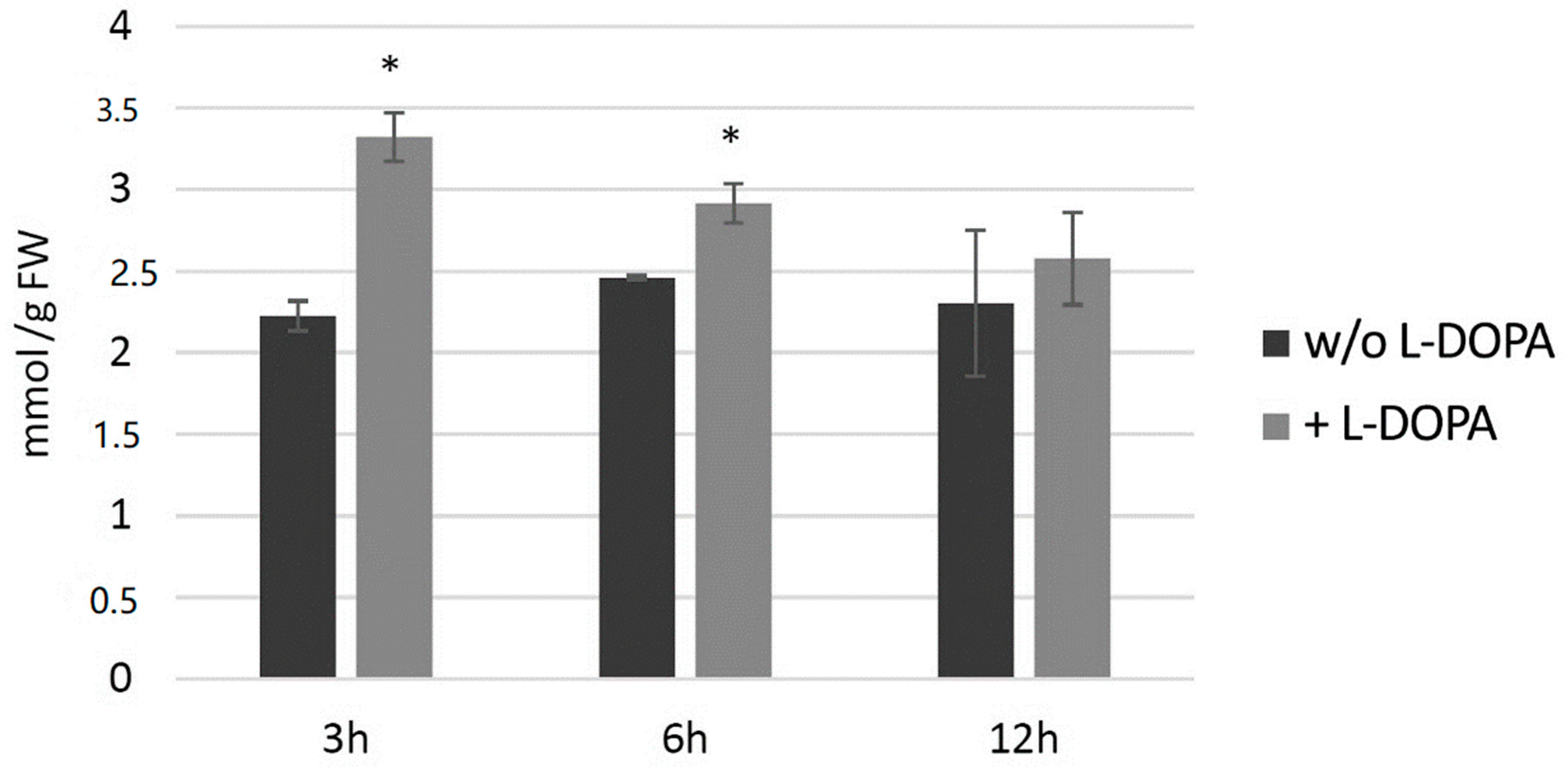
3.7.3. Analysis of H2O2 Level in Potato Plants after Exogenous l-DOPA Treatment
3.7.4. Analysis of Expression of Genes Involved in Free Radical Processing in Potato Plants after Exogenous l-DOPA Treatment
3.8. Resistance of the Transgenic Potatoes to Phytophtora Infestans Attack
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| APX | ascorbate peroxidase |
| CAD | cinnamyl alcohol dehydrogenase |
| CAT | catalase THT—n-hydroxycinnamoyl transferase |
| HCT | shikimate/quinate hydroxy-cinnamoyltransferase |
| PAL | phenylalanine ammonia lyase |
| PPO | polyphenol oxidase |
| SOD | superoxide dismutase |
| TD | tyrosine decarboxylase |
| TH | tyrosine hydroxylase |
References
- Wang, X.; Li, J.; Dong, G.; Yue, J. The endogenous substrates of brain CYP2D. Eur. J. Pharmacol. 2014, 724, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, D.S. Catecholamines 101. Clin. Auton. Res. Off. J. Clin. Auton. Res. Soc. 2010, 20, 331–352. [Google Scholar] [CrossRef] [PubMed]
- Udenfriend, S.; Lovenberg, W.; Sjoerdsma, A. Physiologically Active Amines in Common Fruits and Vegetables. Arch. Biochem. Biophys. 1959, 85, 487–490. [Google Scholar] [CrossRef]
- Kulma, A.; Szopa, J. Catecholamines are Active Compounds in Plants. Plant Sci. 2007, 172, 433–440. [Google Scholar] [CrossRef]
- Gandia-Herrero, F.; Garcia-Carmona, F. Biosynthesis of Betalains: Yellow and Violet Plant Pigments. Trends Plant Sci. 2013, 18, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Aniszewski, T. Alkaloids: Chemistry, Biology, Ecology, and Applications; Elsevier Science: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Newman, M.-A.; von Roepenack-Lahaye, E.; Parr, A.; Daniels, M.J.; Dow, J.M. Induction of Hydroxycinnamoyl-Tyramine Conjugates in Pepper by Xanthomonas campestris, a Plant Defense Response Activated by hrp Gene-Dependent and hrp Gene-Independent Mechanisms. Mol. Plant-Microbe Interact. 2001, 14, 785–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostyn, K.; Czemplik, M.; Kulma, A.; Bortniczuk, M.; Skała, J.; Szopa, J. Genes of Phenylpropanoid Pathway are Activated in Early Response to Fusarium Attack in Flax Plants. Plant Sci. 2012, 190, 103–115. [Google Scholar] [CrossRef]
- Kanazawa, K.; Sakakibara, H. High Content of Dopamine, a Strong Antioxidant, in Cavendish Banana. J. Agric. Food Chem. 2000, 48, 844–848. [Google Scholar] [CrossRef]
- Rahman, M.M.; Ichiyanagi, T.; Komiyama, T.; Hatano, Y.; Konishi, T. Superoxide Radical- and Peroxynitrite-Scavenging Activity of Anthocyanins; Structure-Activity Relationship and their Synergism. Free Radic. Res. 2006, 40, 993–1002. [Google Scholar] [CrossRef]
- Shimizu, T.; Nakanishi, Y.; Nakahara, M.; Wada, N.; Moro-Oka, Y.; Hirano, T.; Konishi, T.; Matsugo, S. Structure Effect on Antioxidant Activity of Catecholamines toward Singlet Oxygen and Other Reactive Oxygen Species in vitro. J. Clin. Biochem. Nutr. 2010, 47, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Ullah, F.; Zhou, D.-X.; Yi, M.; Zhao, Y. Mechanisms of ROS Regulation of Plant Development and Stress Responses. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Swiedrych, A.; Stachowiak, J.; Szopa, J. The Catecholamine Potentates Starch Mobilization in Transgenic Potato Tubers. Plant Physiol. Biochem. 2004, 42, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Skirycz, A.; Swiedrych, A.; Szopa, J. Expression of Human Dopamine Receptor in Potato (Solanum Tuberosum) Results in Altered Tuber Carbon Metabolism. BMC Plant Biol. 2005, 5, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.-R.; Michaels, P.J.; Flores, H.E. Stimulation of Ethylene Production by Catecholamines and Phenylethylamine in Potato Cell Suspension Cultures. Plant Growth Regul. 1993, 12, 219–222. [Google Scholar] [CrossRef]
- Yokoyama, M.; Yamaguchi, S.; Inomata, S.; Komatsu, K.; Yoshida, S.; Iida, T.; Yokokawa, Y.; Yamaguchi, M.; Kaihara, S.; Takimoto, A. Stress-Induced Factor Involved in Flower Formation of Lemna is an Alpha-Ketol Derivative of Linolenic Acid. Plant Cell Physiol. 2000, 41, 110–113. [Google Scholar] [CrossRef] [Green Version]
- Kamisaka, S. Catecholamine Stimulation of the Gibberellin Action that Induces Lettuce Hypocotyl Elongation. Plant Cell Physiol. 1979, 20, 1199–1207. [Google Scholar] [CrossRef]
- Protacio, C.M.; Dai, Y.-R.; Lewis, E.F.; Flores, H.E. Growth Stimulation by Catecholamines in Plant Tissue/Organ Cultures. Plant Physiol. 1992, 98, 89. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Xu, G.; Zhang, K.; Kong, X.; Han, R.; Zhou, J.; Ni, Y. Crystal Structure of Tyrosine Decarboxylase and Identification of Key Residues Involved in Conformational Swing and Substrate Binding. Sci. Rep. 2016, 6, 27779. [Google Scholar] [CrossRef]
- Lehmann, T.; Pollmann, S. Gene Expression and Characterization of a Stress-Induced Tyrosine Decarboxylase from Arabidopsis Thaliana. FEBS Lett. 2009, 583, 1895–1900. [Google Scholar] [CrossRef]
- Świędrych, A.; Lorenc-Kukuła, K.; Skirycz, A.; Szopa, J. The catecholamine Biosynthesis Route in Potato is Affected by Stress. Plant Physiol. Biochem. 2004, 42, 593–600. [Google Scholar] [CrossRef]
- Soares, A.R.; Marchiosi, R.; Siqueira-Soares Rde, C.; Barbosa de Lima, R.; Dantas dos Santos, W.; Ferrarese-Filho, O. The Role of L-DOPA in Plants. Plant Signal Behav. 2014, 9, e28275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.H.; Schwinn, K.E.; Ngo, H.M.; Lewis, D.H.; Massey, B.; Calcott, K.E.; Crowhurst, R.; Joyce, D.C.; Gould, K.S.; Davies, K.M.; et al. Characterisation of Betalain Biosynthesis in Parakeelya Flowers Identifies the Key Biosynthetic Gene Dod as Belonging to an Expanded Ligb Gene Family that is Conserved in Betalain-Producing Species. Front. Plant Sci. 2015, 6, 499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, K.; Kobayashi, N.; Yoshitama, K.; Teramoto, S.; Komamine, A. Isolation and Purification of Tyrosine Hydroxylase from Callus Cultures of Portulaca Grandiflora. Plant Cell Physiol. 2001, 42, 969–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luthra, P.M.; Singh, S. Identification and optimization Of Tyrosine Hydroxylase Activity in Mucuna Pruriens DC. Var. utilis. Planta 2010, 231, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Kahn, V.; Shalom, N.B.; Zakin, V. Effect of Kojic Acid on the Hydroxylation of L-Tyrosine and Tyramine by Mushroom Tyrosinase1. J. Food Biochem. 1998, 22, 455–474. [Google Scholar] [CrossRef]
- Fürstenberg-Hägg, J.; Zagrobelny, M.; Bak, S. Plant Defense against Insect Herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar] [CrossRef] [Green Version]
- Fujii, Y.; Shibuya, T.; Yasuda, T. l-3,4-Dihydroxyphenylalanine as an Allelochemical Candidate from Mucuna Pruriens (L.) DC. Var. utilis. Agric. Biol. Chem. 1991, 55, 617–618. [Google Scholar] [CrossRef]
- Mushtaq, M.N.; Sunohara, Y.; Matsumoto, H. Bioactive l-DOPA Induced Quinoprotein Formation to Inhibit Root Growth of Cucumber Seedlings. J. Pestic. Sci. 2013, 38, 68–73. [Google Scholar] [CrossRef] [Green Version]
- Jana, S.; Sinha, M.; Chanda, D.; Roy, T.; Banerjee, K.; Munshi, S.; Patro, B.S.; Chakrabarti, S. Mitochondrial Dysfunction Mediated by Quinone Oxidation Products of Dopamine: Implications in Dopamine Cytotoxicity and Pathogenesis of Parkinson’s Disease. Biochim. Biophys. Acta (Bba)—Mol. Basis Dis. 2011, 1812, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, K.J.; Shiozawa, N. Misincorporation of Amino Acid Analogues into Proteins by Biosynthesis. Int. J. Biochem. Cell Biol. 2008, 40, 1452–1466. [Google Scholar] [CrossRef]
- Herrera-Vásquez, A.; Salinas, P.; Holuigue, L. Salicylic Acid and Reactive Oxygen Species Interplay in the Transcriptional Control of Defense Genes Expression. Front. Plant Sci. 2015, 6, 171. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Aksamit-Stachurska, A.; Korobczak-Sosna, A.; Kulma, A.; Szopa, J. Glycosyltransferase efficiently Controls Phenylpropanoid Pathway. BMC Biotechnol. 2008, 8, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szopa, J.; Wilczyński, G.; Fiehn, O.; Wenczel, A.; Willmitzer, L. Identification and Quantification of Catecholamines in Potato Plants (Solanum Tuberosum) by GC–MS. Phytochemistry 2001, 58, 315–320. [Google Scholar] [CrossRef]
- Tekin, I.; Roskoski, R.; Carkaci-Salli, N.; Vrana, K.E. Complex molecular Regulation of Tyrosine Hydroxylase. J. Neural Transm. 2014, 121, 1451–1481. [Google Scholar] [CrossRef] [PubMed]
- Roessner, U.; Wagner, C.; Kopka, J.; Trethewey, R.N.; Willmitzer, L. Simultaneous Analysis of Metabolites in Potato Tuber by Gas Chromatography–Mass Spectrometry. Plant J. 2000, 23, 131–142. [Google Scholar] [CrossRef]
- Luedemann, A.; Strassburg, K.; Erban, A.; Kopka, J. TagFinder for the Quantitative Analysis Of Gas Chromatography—Mass Spectrometry (GC-MS)-Based Metabolite Profiling Experiments. Bioinformatics 2008, 24, 732–737. [Google Scholar] [CrossRef]
- Kopka, J.; Schauer, N.; Krueger, S.; Birkemeyer, C.; Usadel, B.; Bergmüller, E.; Dörmann, P.; Weckwerth, W.; Gibon, Y.; Stitt, M.; et al. [email protected]: The Golm Metabolome Database. Bioinformatics 2004, 21, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Nicot, N.; Hausman, J.F.; Hoffmann, L.; Evers, D. Housekeeping Gene Selection for Real-Time RT-PCR Normalization in Potato during Biotic and Abiotic Stress. J. Exp. Bot. 2005, 56, 2907–2914. [Google Scholar] [CrossRef]
- Kedare, S.B.; Singh, R.P. Genesis and Development of DPPH Method of Antioxidant Assay. J. Food Sci. Technol. 2011, 48, 412–422. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Miller, C.J.; Wang, P.; Waite, T.D. Quantitative Determination of Trace Hydrogen Peroxide in the Presence of Sulfide Using the Amplex Red/Horseradish Peroxidase Assay. Anal. Chim. Acta 2017, 963, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient Requirements of Suspension Cultures of Soybean Root Cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Scholz, M.; Kaplan, F.; Guy, C.L.; Kopka, J.; Selbig, J. Non-linear PCA: A Missing Data Approach. Bioinformatics 2005, 21, 3887–3895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of Antioxidant Potential of Plants and its Relevance to Therapeutic Applications. Int. J. Biol. Sci. 2015, 11, 982–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, M.; Levitt, M.; Udenfriend, S. Phenylalanine as Substrate and Inhibitor of Tyrosine Hydroxylase. Arch. Biochem. Biophys. 1967, 120, 420–427. [Google Scholar] [CrossRef]
- DePietro, F.R.; Fernstrom, J.D. The Effect of Phenylalanine on DOPA Synthesis in PC12 Cells. Neurochem. Res. 1998, 23, 1011–1020. [Google Scholar] [CrossRef]
- Fujii, Y. Allelopathy in the Natural and Agricultural Ecosystems and Isolation of Potent Allelochemicals from Velvet bean (Mucuna pruriens) and Hairy vetch (Vicia villosa). Biol. Sci. Space 2003, 17, 6–13. [Google Scholar] [CrossRef]
- Nishihara, E.; Parvez, M.M.; Araya, H.; Fujii, Y. Germination Growth Response of Different Plant Species to the Allelochemical L-3,4-Dihydroxyphenylalanine (L-DOPA). Plant Growth Regul. 2004, 42, 181–189. [Google Scholar] [CrossRef]
- Golisz, A.; Sugano, M.; Hiradate, S.; Fujii, Y. Microarray Analysis of Arabidopsis Plants in Response to Allelochemical L-DOPA. Planta 2010, 233, 231–240. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Hossain, M.A.; da Silva, J.A.T.; Fujita, M. Plant Response and Tolerance to Abiotic Oxidative Stress: Antioxidant Defense is a Key Factor. In Crop Stress and Its Management: Perspectives and Strategies; Venkateswarlu, B., Shanker, A.K., Shanker, C., Maheswari, M., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 261–315. [Google Scholar] [CrossRef]
- Yi, S.Y.; Yu, S.H.; Choi, D. Involvement of Hydrogen Peroxide in Repression of Catalase in TMV-Infected Resistant Tobacco. Mol. Cells 2003, 15, 364–369. [Google Scholar]
- Shim, I.-S.; Momose, Y.; Yamamoto, A.; Kim, D.-W.; Usui, K. Inhibition of Catalase Activity by Oxidative Stress and its Relationship to Salicylic Acid Accumulation in Plants. Plant Growth Regul. 2003, 39, 285–292. [Google Scholar] [CrossRef]
- Lamb, C.; Dixon, R.A. The Oxidative Burst in Plant Disease Resistance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 251–275. [Google Scholar] [CrossRef] [PubMed]
- Averyanov, A. Oxidative Burst and Plant Disease Resistance. Front. Biosci. (Elite Ed.) 2009, 1, 142–152. [Google Scholar] [PubMed]
- Anna, J.-Ż.; Monika, K.; Katarzyna, N. Phenylalanine Ammonia Lyase under Combined Effects of Enhanced Uv-B Radiation and Allelopathy Stress. Acta Biol. Crac. Ser. Bot. 2011, 53, 73–78. [Google Scholar] [CrossRef]
- Wada, K.C.; Mizuuchi, K.; Koshio, A.; Kaneko, K.; Mitsui, T.; Takeno, K. Stress Enhances the Gene Expression and Enzyme Activity of Phenylalanine Ammonia-Lyase and the Endogenous Content of Salicylic Acid to Induce Flowering in Pharbitis. J. Plant Physiol. 2014, 171, 895–902. [Google Scholar] [CrossRef]
- Darmanti, S.; Santosa, L.H.; Dewi, K. Reactive Oxygen Species Accumulations, Phenylalanine Ammonia-Lyase Activity and Phenolic Acid Composition of Soybean [Glycine max (L.) Merr.] cv. Grobogan that Exposed to Multiple Stress of Purple Nutsedge (Cyperus rotundus L.) Interference and Drought. J. Anim. Plant Sci. 2018, 28, 244–251. [Google Scholar]
- Soares, A.R.; de Cássia Siqueira-Soares, R.; Salvador, V.H.; de Lourdes Lucio Ferrarese, M.; Ferrarese-Filho, O. The Effects of l-DOPA on Root Growth, Lignification and Enzyme Activity in Soybean Seedlings. Acta Physiol. Plant. 2012, 34, 1811–1817. [Google Scholar] [CrossRef]
- Jang, S.-M.; Ishihara, A.; Back, K. Production of Coumaroylserotonin and Feruloylserotonin in Transgenic Rice Expressing Pepper Hydroxycinnamoyl-Coenzyme A:Serotonin N-(Hydroxycinnamoyl)transferase. Plant Physiol. 2004, 135, 346. [Google Scholar] [CrossRef] [Green Version]
- Pearce, G.; Marchand, P.; Griswold, J.; Lewis, N.; Ryan, C. Accumulation of Feruloyltyramine and P-Coumaroyltyramine in Tomato Leaves in Response to Wounding. Phytochemistry 1998, 47, 659. [Google Scholar] [CrossRef]
- Sprenger, H.; Kurowsky, C.; Horn, R.; Erban, A.; Seddig, S.; Rudack, K.; Fischer, A.; Walther, D.; Zuther, E.; Köhl, K.; et al. The Drought Response of Potato Reference Cultivars with Contrasting Tolerance. Plant Cell Environ. 2016, 39, 2370–2389. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.; Flors, V.; Garcia-Agustin, P.; Jakab, G.; Mauch, F.; Newman, M.A.; Pieterse, C.M.; Poinssot, B.; Pozo, M.J.; et al. Priming: Getting Ready for Battle. Mol. Plant Microbe Interact. 2006, 19, 1062–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalenahalli, Y.; Kumar, A.; Sarkar, K.; Li, Y.; Doddaraju, P.; Mosa, K.; Duggavathi, R.; Kushalappa, A. Transcription Factor StWRKY1 regulates Phenylpropanoid Metabolites Conferring Late Blight Resistance in Potato. J. Exp. Bot. 2015. [Google Scholar] [CrossRef] [Green Version]
- Supriya, A.; Sharma, P.; Yadav, N.; Yadav, R. MYB transcription Factor Genes as Regulators for Plant Responses: An Overview. Physiol. Mol. Biol. Plants 2013, 19. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.; Constabel, C.P. MYB Repressors as Regulators of Phenylpropanoid Metabolism in Plants. Trends Plant Sci. 2019, 24, 275–289. [Google Scholar] [CrossRef]
- Etalo, D.W.; Stulemeijer, I.J.E.; Peter van Esse, H.; de Vos, R.C.H.; Bouwmeester, H.J.; Joosten, M.H.A.J. System-Wide Hypersensitive Response-Associated Transcriptome and Metabolome Reprogramming in Tomato. Plant Physiol. 2013, 162, 1599. [Google Scholar] [CrossRef] [Green Version]
- Kage, U.; Yogendra, K.N.; Kushalappa, A.C. TaWRKY70 Transcription Factor in Wheat QTL-2DL Regulates Downstream Metabolite Biosynthetic Genes to Resist Fusarium Graminearum Infection Spread within Spike. Sci. Rep. 2017, 7, 42596. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Das, A.; Khandagale, P.; Maiti, I.B.; Chattopadhyay, S.; Dey, N. Interaction of Arabidopsis TGA3 and WRKY53 Transcription Factors on Cestrum Yellow Leaf Curling Virus (CmYLCV) promoter Mediates Salicylic Acid-Dependent Gene Expression in Planta. Planta 2018, 247, 181–199. [Google Scholar] [CrossRef]
- Kuhar, M.J.; Couceyro, P.R.; Lambert, P.D. Biosynthesis of Catecholamines. In Basic Neurochemistry: Molecular, Cellular and Medical Aspects, 6th ed.; Lippincott-Raven: Philadelphia, PA, USA, 1999. [Google Scholar]
- Wang, J.; Lou, H.; Pedersen, C.; Smith, A.; Perez, R. 14-3-3zeta Contributes to Tyrosine Hydroxylase Activity in Mn9d Cells: Localization of Dopamine Regulatory Proteins to Mitochondria. J. Biol. Chem. 2009, 284, 14011–14019. [Google Scholar] [CrossRef] [Green Version]
- Cartier, E.A.; Parra, L.A.; Baust, T.B.; Quiroz, M.; Salazar, G.; Faundez, V.; Egaña, L.; Torres, G.E. A Biochemical and Functional Protein Complex Involving Dopamine Synthesis and Transport into Synaptic Vesicles. J. Biol. Chem. 2010, 285, 1957–1966. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Potato Line | Free Phenolics [%] | Cell Wall-Bound Phenolics [%] |
|---|---|---|
| CTRL | 100 | 100 |
| HTZ8 | 212.3 | 148.5 |
| HTZ12 | 194.6 | 145.3 |
| HTZ28 | 249.7 | 169.4 |
| HTZ43 | 198.5 | 137.4 |
| HTZ53 | 132.5 | 107.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostyn, K.; Boba, A.; Kostyn, A.; Kozak, B.; Starzycki, M.; Kulma, A.; Szopa, J. Expression of the Tyrosine Hydroxylase Gene from Rat Leads to Oxidative Stress in Potato Plants. Antioxidants 2020, 9, 717. https://doi.org/10.3390/antiox9080717
Kostyn K, Boba A, Kostyn A, Kozak B, Starzycki M, Kulma A, Szopa J. Expression of the Tyrosine Hydroxylase Gene from Rat Leads to Oxidative Stress in Potato Plants. Antioxidants. 2020; 9(8):717. https://doi.org/10.3390/antiox9080717
Chicago/Turabian StyleKostyn, Kamil, Aleksandra Boba, Anna Kostyn, Bartosz Kozak, Michał Starzycki, Anna Kulma, and Jan Szopa. 2020. "Expression of the Tyrosine Hydroxylase Gene from Rat Leads to Oxidative Stress in Potato Plants" Antioxidants 9, no. 8: 717. https://doi.org/10.3390/antiox9080717