Bioactive Variability and In Vitro and In Vivo Antioxidant Activity of Unprocessed and Processed Flour of Nine Cultivars of Australian lupin Species: A Comprehensive Substantiation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Materials
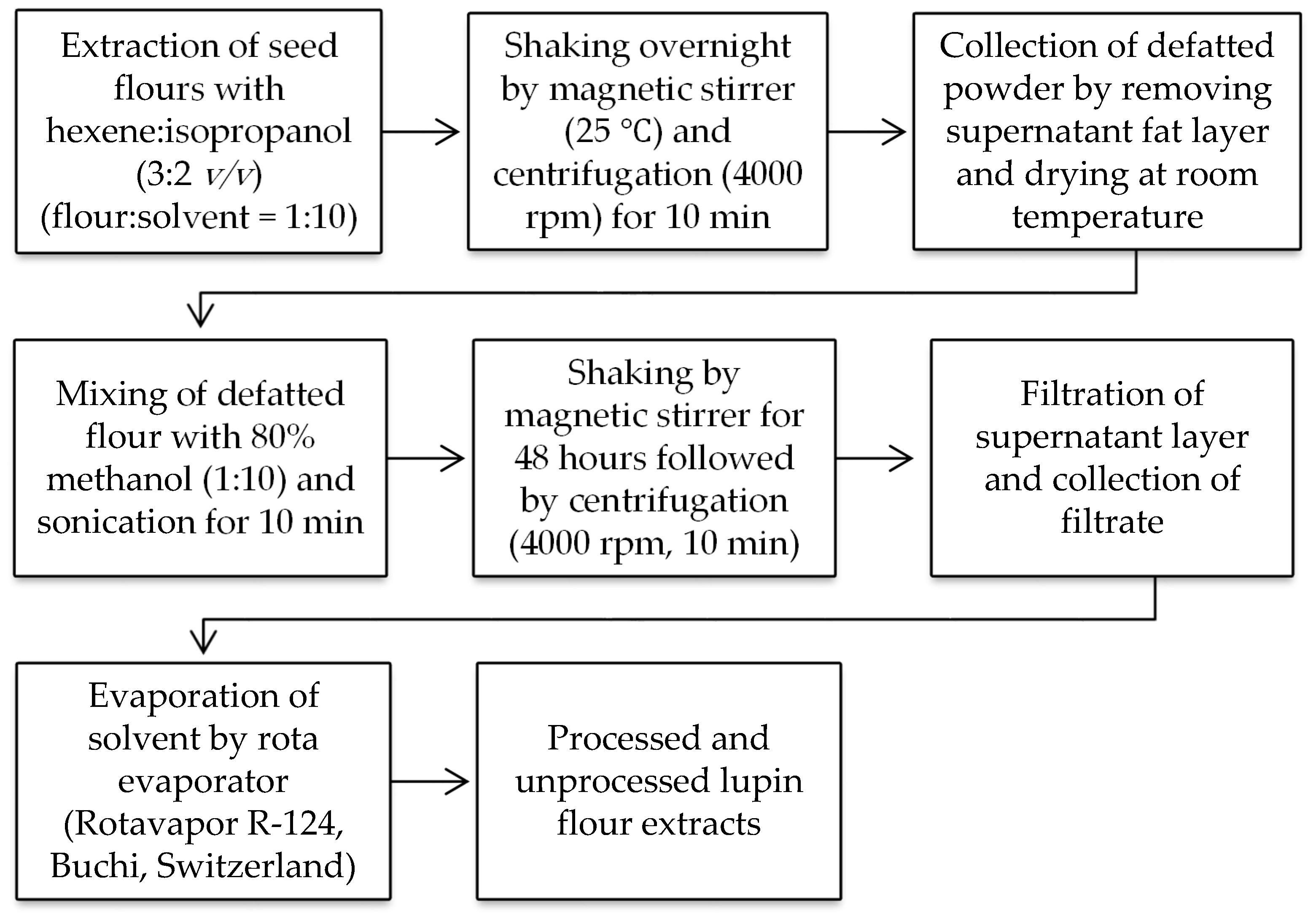
2.2.1. Preparation of Unprocessed and Processed Flour Samples
2.2.2. Extraction of Unprocessed and Processed Flours
2.3. GC-MS Analysis and Identification of Bioactive Compounds
2.4. Assessment of Antioxidant Activity
2.5. Assay of Non-Enzymatic Antioxidants
2.5.1. Total Phenolic Content (TPC)
2.5.2. Total Flavonoid Content
2.5.3. Total Flavonol Content
2.6. In-Vitro Evaluation of Antioxidant Activity
2.6.1. Evaluation of Superoxide Radical Scavenging Activity
2.6.2. Evaluation of DPPH• Radical Scavenging Activity
2.6.3. Evaluation of Inhibition of LPO Using TBARS Method
2.6.4. Evaluation of Hydroxyl Radical Scavenging Activity
2.6.5. Evaluation of ABTS+• Radical Scavenging Activity
2.6.6. Evaluation of Hydrogen Peroxide (H2O2) Scavenging Activity
2.6.7. Evaluation of Anti-Hemolytic Activity
2.7. In-Vivo Assay of Antioxidant Activity
2.7.1. Experimental Animals
2.7.2. Experimental Design
2.7.3. Biochemical Assay of CAT, SOD, MDA Activity and Protein Content
2.7.4. Assay of CAT Activity
2.7.5. Assay of LPO Activity by TBARS Method
2.7.6. Assay of SOD Activity
2.7.7. Estimation of Protein Content
2.8. Statistical Analysis
3. Results and Discussion
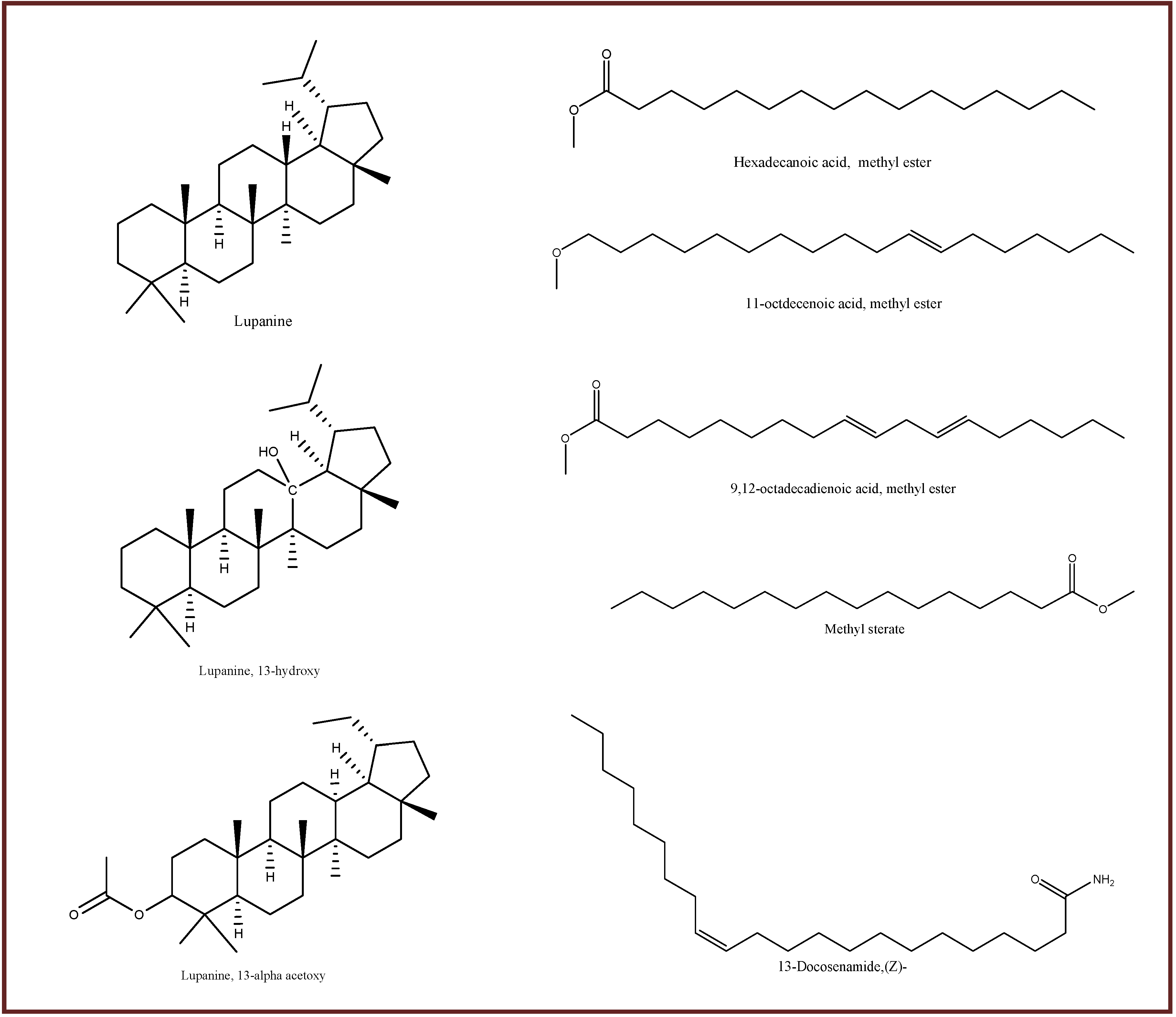
3.1. GC-MS Analysis and Identification of Bioactive Compounds
3.2. Assay of Non-Enzymatic Antioxidants
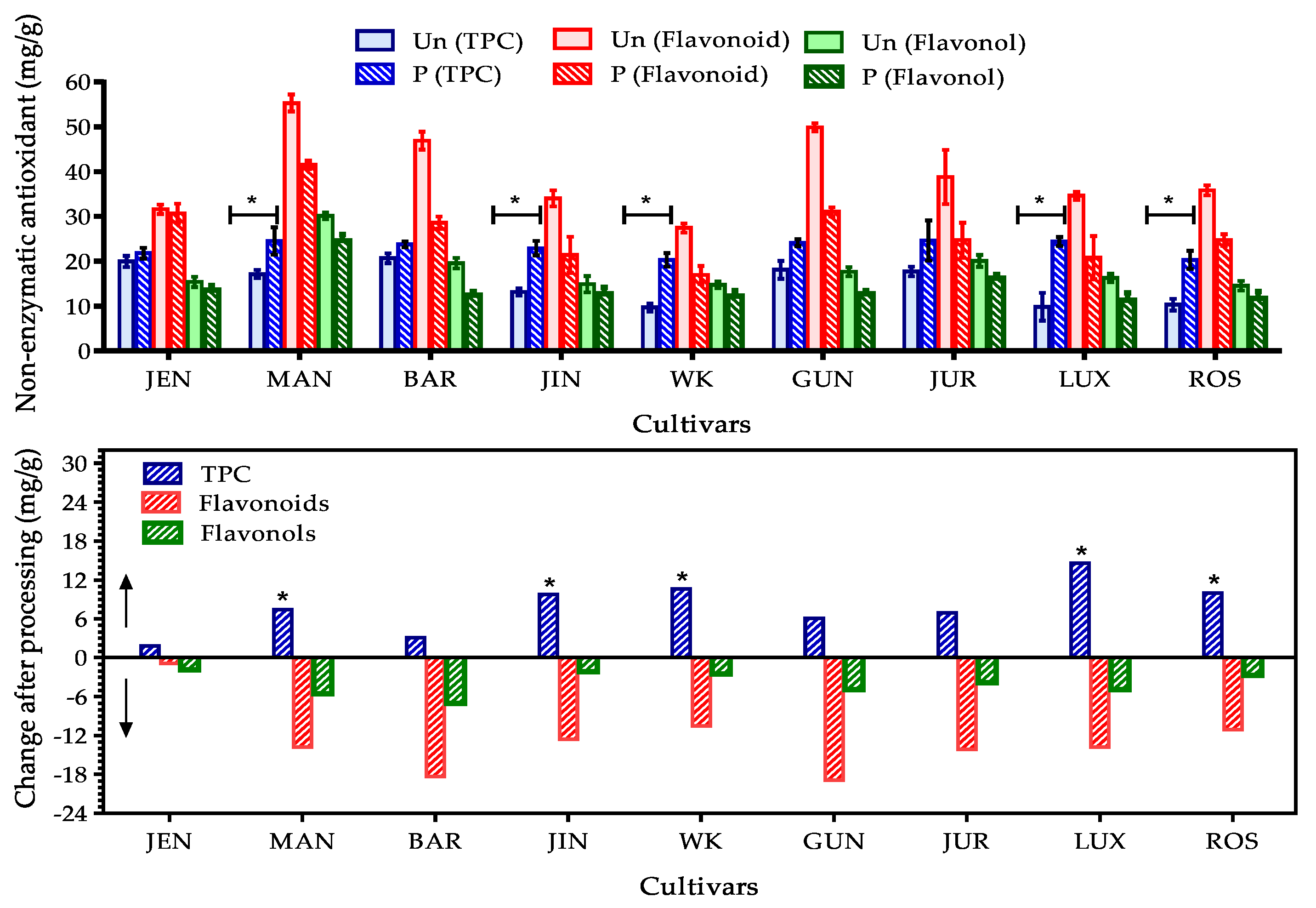
3.2.1. Total Phenolic, Flavonoid and Flavonol Content in Lupin Flours
3.2.2. Effect of Processing on Total Phenolic, Flavonoid and Flavonol Content
3.3. In-Vitro Evaluation of Antioxidant Activity
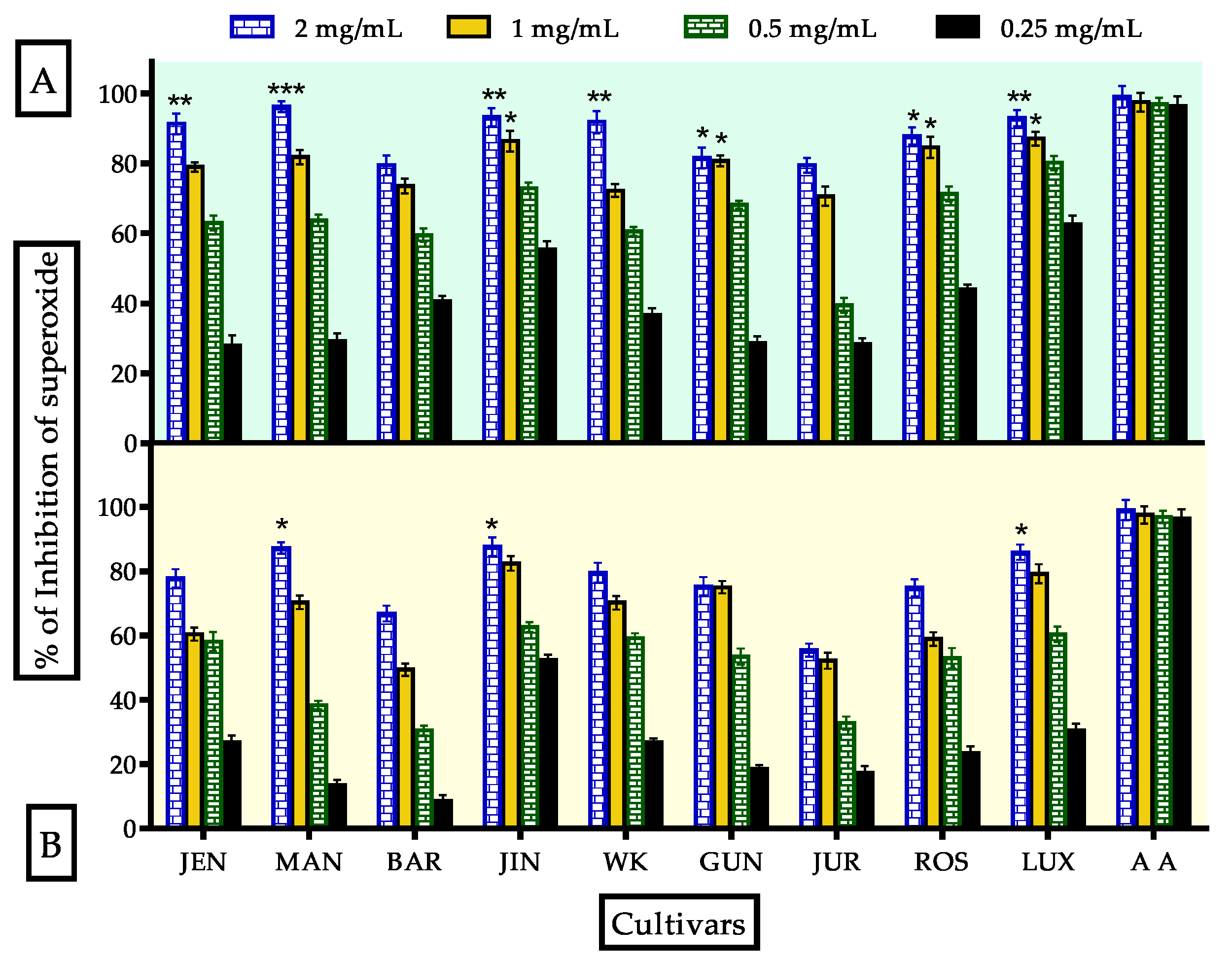
3.3.1. Evaluation of Superoxide Radical Scavenging Activity
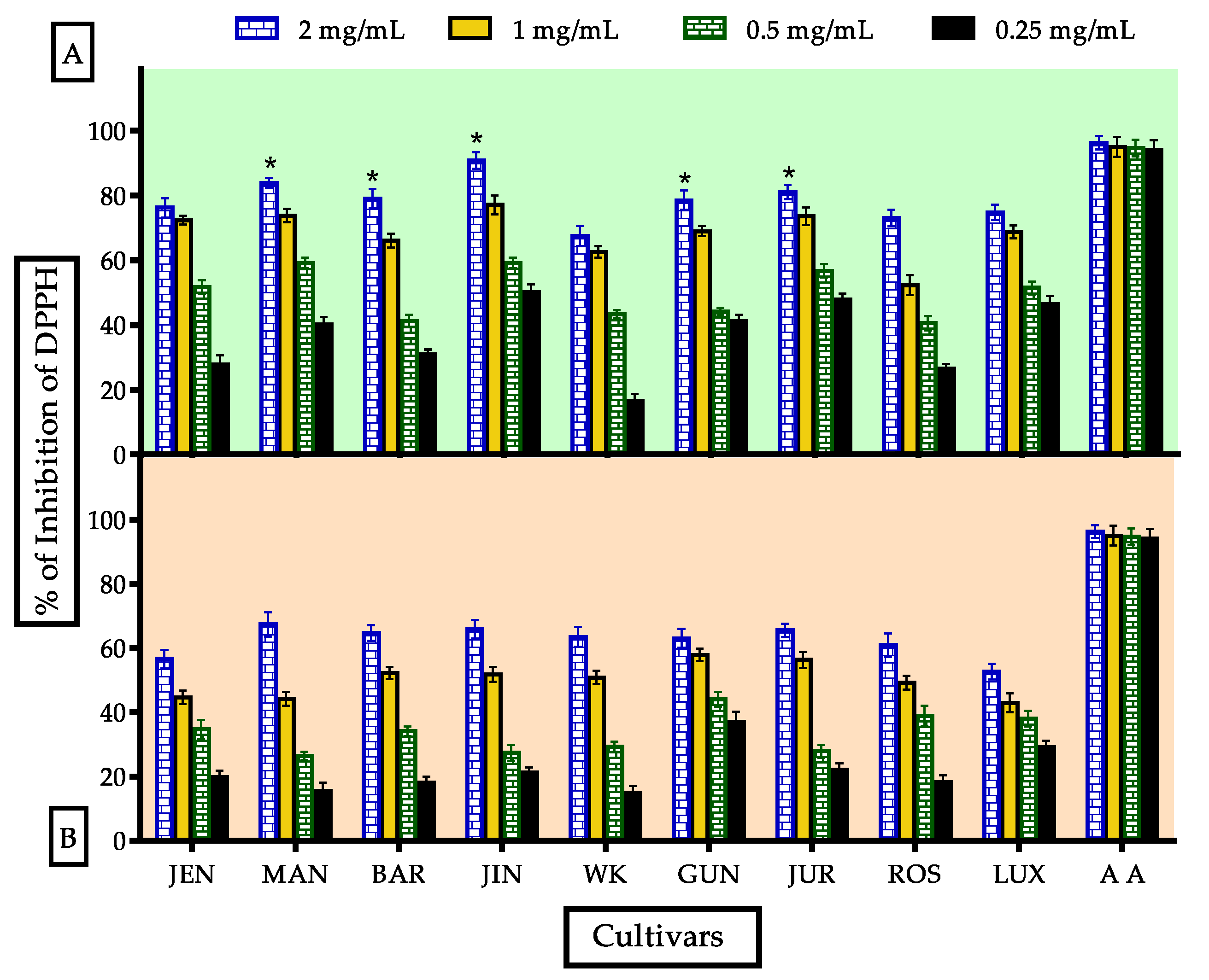
3.3.2. Evaluation of DPPH• Radical Scavenging Activity
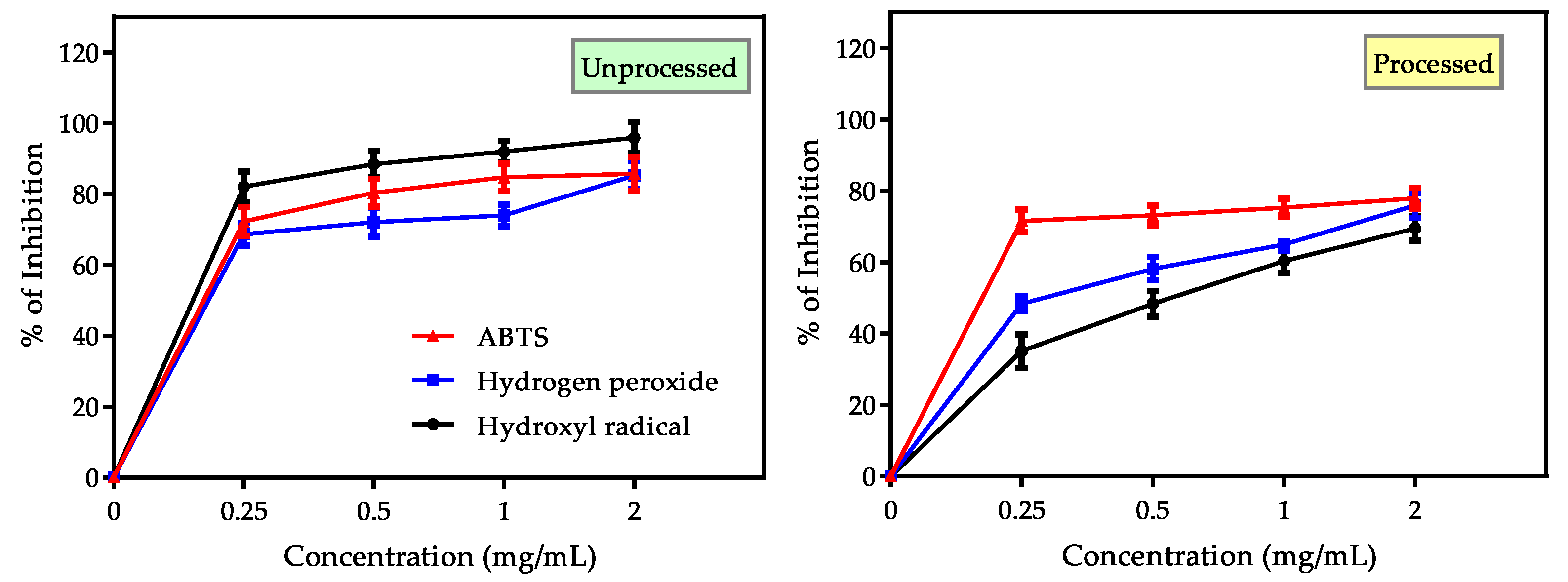
3.3.3. Evaluation of Hydrogen Peroxide, Hydroxyl and ABTS+• Radical Scavenging Activity
3.3.4. Evaluation of Inhibition of LPO Activity Using TBARS Method
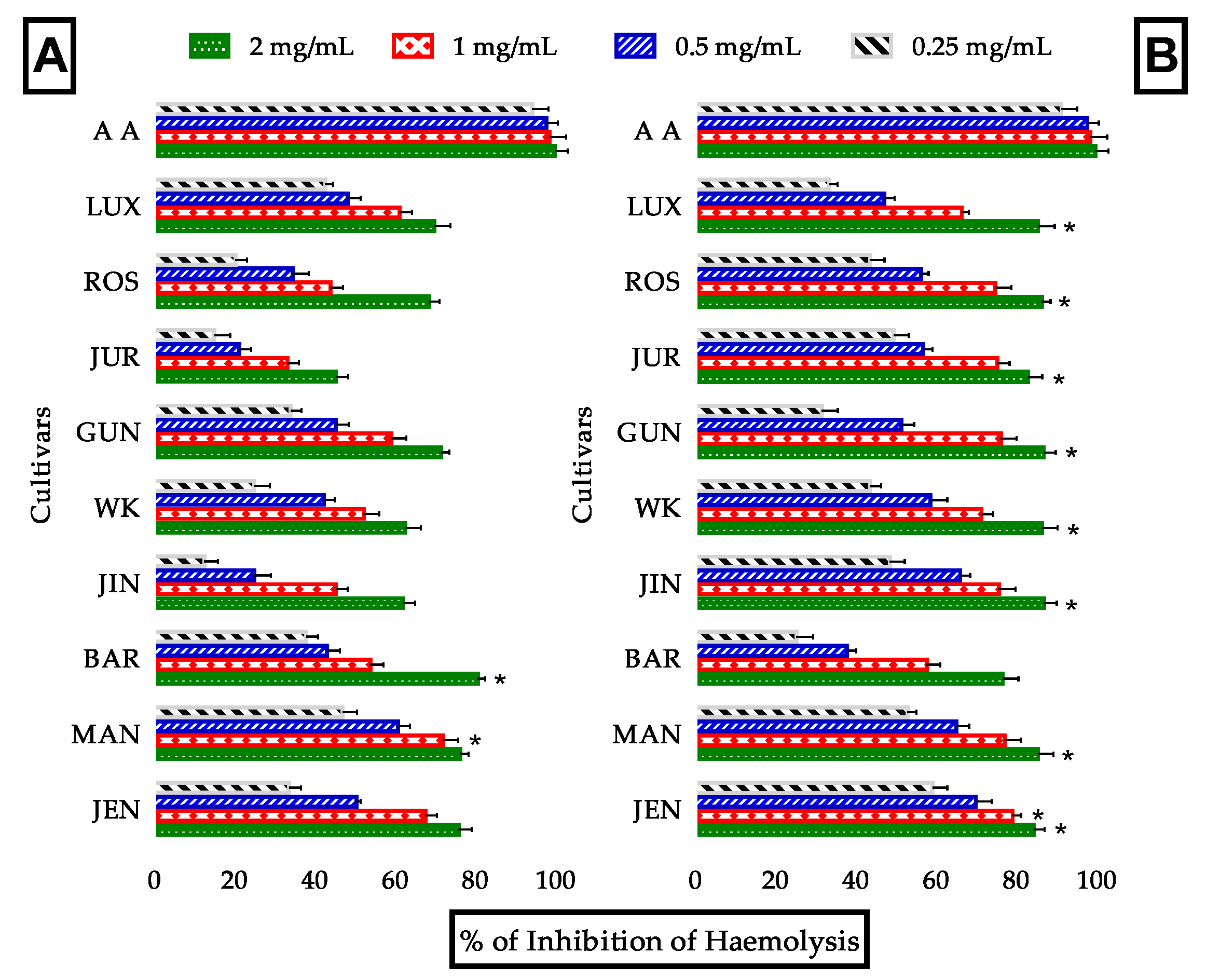
3.3.5. Evaluation of Anti-Hemolytic Activity
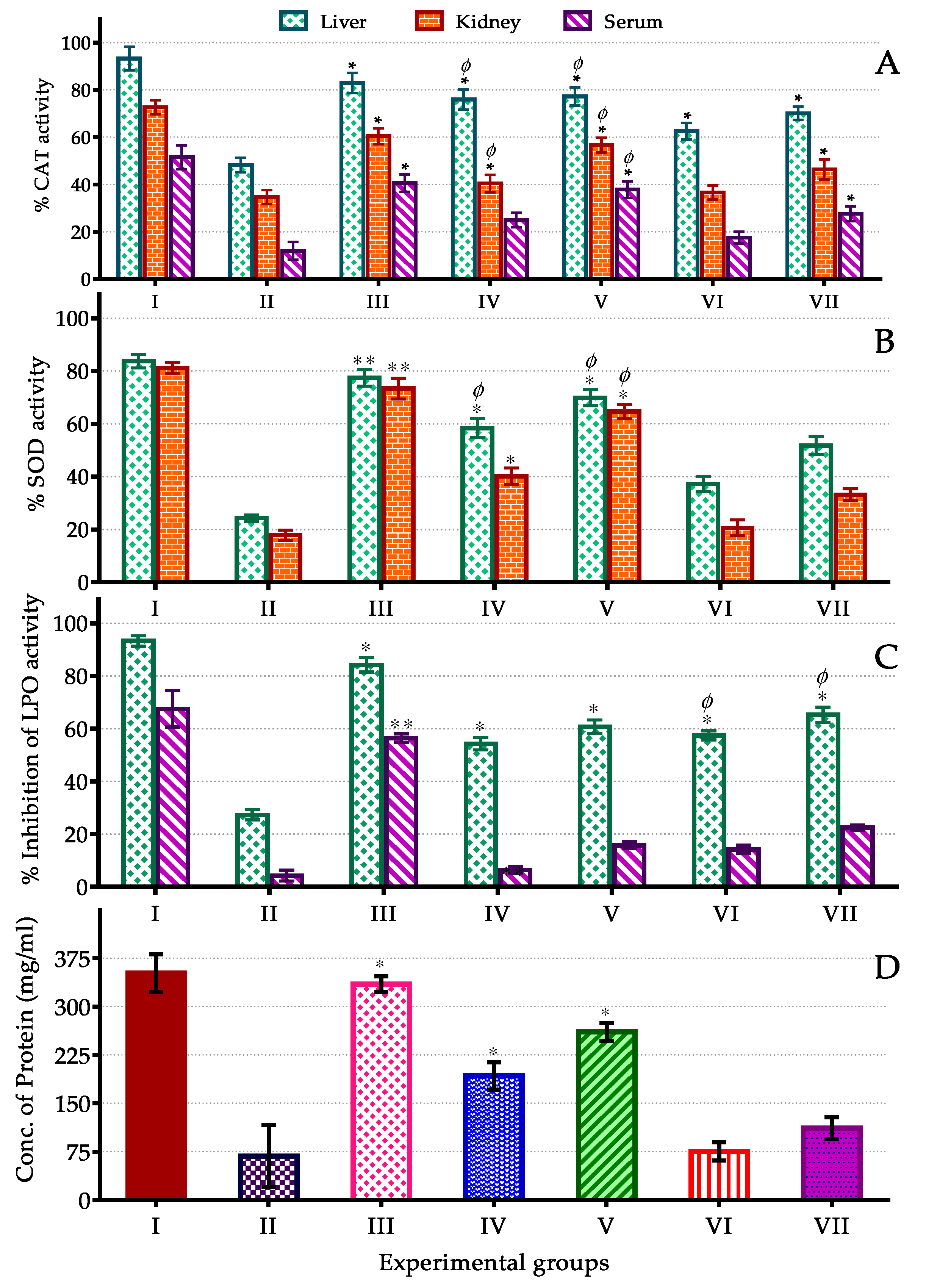
3.4. In Vivo Assay of CAT, SOD, LPO Activity and Protein Content
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Halliwell, B.; Gutteridge, J.M.; Cross, C.E. Free radicals, antioxidants, and human disease: Where are we now? J. Lab. Clin. Med. 1992, 119, 598–620. [Google Scholar] [PubMed]
- Young, I.S.; Woodside, J.V. Antioxidants in health and disease. J. Clin. Pathol. 2001, 54, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Dontha, S. A Review on Antioxidant Methods. Asian. J. Pharm. Clin. Res. 2016, 9, 14–32. [Google Scholar] [CrossRef]
- Scalbert, A.; Manach, C.; Morand, C.; Remesy, C.; Jimenez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef]
- Robertson, D. Metabonomics in Toxicology: A Review. Toxicol. Sci. 2005, 85, 809–822. [Google Scholar] [CrossRef]
- Fernie, A.R.; Trethewey, R.N.; Krotzky, A.J.; Willmitzer, L. Metabolite profiling: From diagnostics to systems biology. Nat. Rev. Mol. Cell Biol. 2004, 5, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Kell, D.B.; Brown, M.; Davey, H.M.; Dunn, W.B.; Spasic, I.; Oliver, S.G. Metabolic footprinting and systems biology: The medium is the message. Nat. Rev. Microbiol. 2005, 3, 557–565. [Google Scholar] [CrossRef]
- Rohloff, J. Analysis of phenolic and cyclic compounds in plants using derivatization techniques in combination with GC-MS-based metabolite profiling. Molecules 2015, 20, 3431–3462. [Google Scholar] [CrossRef] [Green Version]
- Siger, A. Antioxidant activity and phenolic content in three lupin species. Subtrop. Plant Sci. 2012, 25, 190–197. [Google Scholar] [CrossRef]
- Khan, M.K.; Karnpanit, W.; Nasar-Abbas, S.M.; Huma, Z.-e.; Jayasena, V. Phytochemical composition and bioactivities of lupin: A review. Int. J. Food Sci. Tech. 2015, 50, 2004–2012. [Google Scholar] [CrossRef]
- French, R.J.; Buirchell, B.J. Lupin: The largest grain legume crop in Western Australia, its adaptation and improvement through plant breeding. Aust. J. Agric. Res. 2005, 56, 1169–1180. [Google Scholar] [CrossRef]
- Kalogeropoulos, N.; Chiou, A.; Ioannou, M.; Karathanos, V.; Hassapidou, M.; Andrikopoulos, N. Nutritional evaluation and bioactive microconstituents (phytosterols, tocopherols, polyphenols, triterpenic acids) in cooked dry legumes usually consumed in the Mediterranean countries. Food Chem. 2010, 121, 682–690. [Google Scholar] [CrossRef]
- Turkmen, N.; Sari, F.; Velioglu, Y.S. The effect of cooking methods on total phenolics and antioxidant activity of selected green vegetables. Food Chem. 2005, 93, 713–718. [Google Scholar] [CrossRef]
- Davies, N.W. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicon and Carbowax 20M phases. J. Chromatogr. A 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Song, F.-L.; Gan, R.-Y.; Zhang, Y.; Xiao, Q.; Kuang, L.; Li, H.-B. Total phenolic contents and antioxidant capacities of selected chinese medicinal plants. Int. J. Mol. Sci. 2010, 11, 2362–2372. [Google Scholar] [CrossRef] [PubMed]
- Fernando, C.D.; Soysa, P. Total phenolic, flavonoid contents, in-vitro antioxidant activities and hepatoprotective effect of aqueous leaf extract of Atalantia ceylanica. BMC Complementary Altern. Med. 2014, 14, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almaraz, N.; Campos, M.; Ávila-Reyes, J.; Naranjo-Jiménez, N.; Corral, J.; Gonzalez, L. Antioxidant activity of polyphenolic extract of monofloral honeybee-collected pollen from mesquite (Prosopis juliflora, Leguminosae). J. Food Compos. Anal. 2007, 20, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Sahoo, S.; Ghosh, G.; Das, D.; Nayak, S. Phytochemical investigation and in vitro antioxidant activity of an indigenous medicinal plant Alpinia nigra BL Burtt. Asian Pac. J. Trop. Biomed. 2013, 3, 871–876. [Google Scholar] [CrossRef] [Green Version]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Hill, S.; Lamberson, C.R.; Xu, L.; To, R.; Tsui, H.S.; Shmanai, V.V.; Bekish, A.V.; Awad, A.M.; Marbois, B.N.; Cantor, C.R.; et al. Small amounts of isotope-reinforced polyunsaturated fatty acids suppress lipid autoxidation. Free Radic. Biol. Med. 2012, 53, 893–906. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, R.; Chandrasekar, M.J.; Nanjan, M.J.; Suresh, B. Antioxidant activity of Caesalpinia digyna root. J. Ethnopharmacol. 2007, 113, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Shirwaikar, A.; Rajendran, K.; Kumar, C.D. In vitro antioxidant studies of Annona squamosa Linn. leaves. Indian J. Exp. Biol. 2004, 42, 803–807. [Google Scholar] [PubMed]
- Jayaprakasha, G.K.; Jaganmohan Rao, L.; Sakariah, K.K. Antioxidant activities of flavidin in different in vitro model systems. Bioorg. Med. Chem. 2004, 12, 5141–5146. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimzadeh, M.A.; Nabavi, S.F.; Nabavi, S.M.; Eslami, B. Antihemolytic and antioxidant activities of Allium paradoxum. Cent. Eur. J. Biol. 2010, 5, 338–345. [Google Scholar] [CrossRef]
- Sinha, A.K. Colorimetric assay of catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [CrossRef]
- McCord, J.M.; Fridovich, I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Jahan, N.; Parvin, M.S.; Das, N.; Islam, M.S.; Islam, M.E. Studies on the antioxidant activity of ethanol extract and its fractions from Pterygota alata leaves. J. Acute Med. 2014, 4, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Frick, K.; Foley, R.; Kamphuis, L.; Siddique, K.; Garg, G.; Singh, K. Characterisation of the genetic factors affecting quinolizidine alkaloid biosynthesis and its response to abiotic stress in narrow-leafed lupin (Lupinus angustifolius L.): Lupin alkaloid biosynthesis and response to stress. Plant Cell Environ. 2018, 41, 2155–2168. [Google Scholar] [CrossRef] [Green Version]
- Giallourou, N.; Oruna-Concha, M.J.; Harbourne, N. Effects of domestic processing methods on the phytochemical content of watercress (Nasturtium officinale). Food Chem. 2016, 212, 411–419. [Google Scholar] [CrossRef]
- Palermo, M.; Pellegrini, N.; Fogliano, V. The effect of cooking on the phytochemical content of vegetables. J. Sci. Food Agr. 2014, 94, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Lalitha, P.; Veena, V.; Vidhyapriya, P.; Lakshmi, P.; Krishna, R.; Sakthivel, N. Anticancer potential of pyrrole (1, 2, a) pyrazine 1, 4, dione, hexahydro 3-(2-methyl propyl) (PPDHMP) extracted from a new marine bacterium, Staphylococcus sp. strain MB30. Apoptosis 2016, 21, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Kalpana, D.; Shanmugasundaram, R.; Mohan, V. GC–MS analysis of ethanol extract of Entada pursaetha DC seed. Biosci Disc. 2012, 3, 30–33. [Google Scholar]
- Tyagi, T.; Mala, A. Phytochemical and GC-MS analysis of bioactive constituents in the ethanolic of Pistia stratiotes L. and Eichhornia crassipes (Mart.) solms. J. Pharm. Phytochem. 2017, 6, 195–206. [Google Scholar]
- Freire, R.; Morais, S.; Pinheiro, D. Synthesis and antioxidant, anti-inflammatory and gastroprotector activities of anethole and related compounds. Bioorg. Med. Chem. 2005, 13, 4353–4358. [Google Scholar] [CrossRef] [PubMed]
- Saravana Kumar, P.; Al-Dhabi, N.A.; Duraipandiyan, V.; Balachandran, C.; Praveen Kumar, P.; Ignacimuthu, S. In vitro antimicrobial, antioxidant and cytotoxic properties of Streptomyces lavendulae strain SCA5. BMC Microbiol. 2014, 14, 291. [Google Scholar] [CrossRef] [Green Version]
- Erdemoglu, N.; Ozkan, S.; Tosun, F. Alkaloid profile and antimicrobial activity of Lupinus angustifolius L. alkaloid extract. Phytochem. Rev. 2007, 6, 197–201. [Google Scholar] [CrossRef]
- Ahmad, M.; Baba, W.N.; Gani, A.; Wani, T.A.; Gani, A.; Masoodi, F.A. Effect of extraction time on antioxidants and bioactive volatile components of green tea (Camellia sinensis), using GC/MS. Cogent Food Agric. 2015, 1, 1106387. [Google Scholar] [CrossRef]
- Caboni, P.; Ntalli, N.; Aissani, N.; Cavoski, I.; Angioni, A. Nematicidal Activity of (E,E)-2,4-Decadienal and (E)-2-Decenal from Ailanthus altissima against Meloidogyne javanica. J. Agr. Food Chem. 2012, 60, 1146–1151. [Google Scholar] [CrossRef]
- Georgieva, N.; Kosev, V.; Genov, N.; Butnariu, M. Morphological and biological characteristics of white lupine cultivars (Lupinus albus L.). Rom. Agric. Res. 2018, 35, 109–119. [Google Scholar]
- Mazur, M.; Polakowski, P.; Szadowska, A. ”Pharmacologic studies on lupanine and 13-hydroxylupanine”. Acta Phys. Pol. 1966, 17, 299–309. [Google Scholar]
- Mazur, M.; Polakowski, P.; Szadowska, A. “Pharmacologic studies on 17-oxolupanine, lupanine aminooxide and 17-hydroxylupanine”. Acta Phys. Pol. 1966, 17, 311–320. [Google Scholar]
- Garcia Lopez, P.M.; de la Mora, P.G.; Wysocka, W.; Maiztegui, B.; Alzugaray, M.E.; Del Zotto, H.; Borelli, M.I. Quinolizidine alkaloids isolated from Lupinus species enhance insulin secretion. Eur. J. Pharmacol. 2004, 504, 139–142. [Google Scholar] [CrossRef]
- Wiedemann, M.; Gurrola-Diaz, C.M.; Vargas-Guerrero, B.; Wink, M.; Garcia-Lopez, P.M.; Dufer, M. Lupanine Improves Glucose Homeostasis by Influencing KATP Channels and Insulin Gene Expression. Molecules 2015, 20, 19085–19100. [Google Scholar] [CrossRef]
- Kolak, U.; Gazioğlu, I.; Boga, M.; Özgökçe, F.; Ünal, M.; Choudhary, M.; Ulubelen, A. Phytochemical Investigation of Leontice leontopetalum L. subsp. ewersmannii with Antioxidant and Anticholinesterase Activities. Rec. Nat. Prod. 2011, 5, 309–313. [Google Scholar]
- Rahman, M.M.; Ahmad, S.H.; Mohamed, M.T.M.; Ab Rahman, M.Z. Antimicrobial compounds from leaf extracts of Jatropha curcas, Psidium guajava, and Andrographis paniculata. Sci. World J. 2014, 2014, 635240. [Google Scholar] [CrossRef] [Green Version]
- Adnan, M.; Nazim Uddin Chy, M.; Mostafa Kamal, A.T.M.; Azad, M.O.K.; Paul, A.; Uddin, S.B.; Barlow, J.W.; Faruque, M.O.; Park, C.H.; Cho, D.H. Investigation of the Biological Activities and Characterization of Bioactive Constituents of Ophiorrhiza rugosa var. prostrata (D.Don) & Mondal Leaves through In Vivo, In Vitro, and In Silico Approaches. Molecules 2019, 24, 1367. [Google Scholar] [CrossRef] [Green Version]
- Pinto, M.E.A.; Araújo, S.G.; Morais, M.I.; Sá, N.P.; Lima, C.M.; Rosa, C.A.; Siqueira, E.P.; Johann, S.; Lima, L.A.R.S. Antifungal and antioxidant activity of fatty acid methyl esters from vegetable oils. An. Acad. Bras. Ciênc. 2017, 89, 1671–1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andor, B.; Danciu, C.; Alexa, E.; Zupko, I.; Hogea, E.; Cioca, A.; Coricovac, D.; Pinzaru, I.; Pătrașcu, J.M.; Mioc, M.; et al. Germinated and Ungerminated Seeds Extract from Two Lupinus Species: Biological Compounds Characterization and In Vitro and In Vivo Evaluations. Evid. Based Complementary Altern. Med. 2016, 2016, 8. [Google Scholar] [CrossRef] [Green Version]
- Lampart-Szczapa, E.; Siger, A.; Trojanowska, K.; Nogala-Kalucka, M.; Malecka, M.; Pacholek, B. Chemical composition and antibacterial activities of lupin seeds extracts. Nahrung 2003, 47, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Oomah, B.D.; Tiger, N.; Olson, M.; Balasubramanian, P. Phenolics and antioxidative activities in narrow-leafed lupins (Lupinus angustifolius L.). Plant Food Hum. Nutr. 2006, 61, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Kalam Azad, M.O.; Jeong, D.I.; Adnan, M.; Salitxay, T.; Heo, J.W.; Naznin, M.T.; Lim, J.D.; Cho, D.H.; Park, B.J.; Park, C.H. Effect of Different Processing Methods on the Accumulation of the Phenolic Compounds and Antioxidant Profile of Broomcorn Millet (Panicum miliaceum L.) Flour. Foods 2019, 8, 230. [Google Scholar] [CrossRef] [Green Version]
- Kruawan, K.; Tongyonk, L.; Kangsadalampai, K. Antimutagenic and co-mutagenic activities of some legume seeds and their seed coats. J. Med. Plant Res. 2012, 6, 3845–3851. [Google Scholar]
- Yu, L.; Nanguet, A.L.; Beta, T. Comparison of Antioxidant Properties of Refined and Whole Wheat Flour and Bread. Antioxidants 2013, 2, 370–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaaban, H.; Ioannou, I.; Chebil, L.; Slimane, M.; Gérardin, C.; Paris, C.; Charbonnel, C.; Chekir, L.; Ghoul, M. Effect of heat processing on thermal stability and antioxidant activity of six flavonoids. J. Food Process. Preserv. 2017, 41, e13203. [Google Scholar] [CrossRef]
- Aoyama, S.; Yamamoto, Y. Antioxidant Activity and Flavonoid Content of Welsh Onion (Allium fistulosum) and the Effect of Thermal Treatment. Food Sci. Technol. Res. 2007, 13, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Viña, S.Z.; Chaves, A.R. Effect of heat treatment and refrigerated storage on antioxidant properties of pre-cut celery (Apium graveolens L.). Int. J. Food Sci. Tech. 2008, 43, 44–51. [Google Scholar] [CrossRef]
- Serpen, A.; Gökmen, V.; Fogliano, V. Total antioxidant capacities of raw and cooked meats. Meat Sci. 2012, 90, 60–65. [Google Scholar] [CrossRef]
- Sharma, K.; Ko, E.Y.; Assefa, A.D.; Ha, S.; Nile, S.H.; Lee, E.T.; Park, S.W. Temperature-dependent studies on the total phenolics, flavonoids, antioxidant activities, and sugar content in six onion varieties. J. Food Drug Anal. 2015, 23, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Monreal, A.M.; Garcia-Diz, L.; Martinez-Tome, M.; Mariscal, M.; Murcia, M.A. Influence of cooking methods on antioxidant activity of vegetables. J. Food Sci. 2009, 74, H97–H103. [Google Scholar] [CrossRef]
- Nicoli, M.C.; Anese, M.; Parpinel, M. Influence of processing on the antioxidant properties of fruit and vegetables. Trends Food Sci. Technol. 1999, 10, 94–100. [Google Scholar] [CrossRef]
- Terpinc, P.; Polak, T.; Šegatin, N.; Hanzlowsky, A.; Ulrih, N.; Abramovi, H. Antioxidant properties of 4-vinyl derivatives of hydroxycinnamic acid. Food Chem. 2011, 128, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Stepanić, V.; Matijašić, M.; Horvat, T.; Verbanac, D.; Kučerová-Chlupáčová, M.; Saso, L.; Žarković, N. Antioxidant Activities of Alkyl Substituted Pyrazine Derivatives of Chalcones-In Vitro and In Silico Study. Antioxidants 2019, 8, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puntel, G.; de Carvalho, N.; Gubert, P.; Schwertner Palma, A.; Dalla Corte, C.; Avila, D.; Pereira, M.; Gervini, V.; Bresolin, L.; Rocha, J.B.; et al. Butane-2,3-dionethiosemicarbazone: An oxime with antioxidant properties. Chem. Biol. Interact. 2008, 177, 153–160. [Google Scholar] [CrossRef]
- Van Acker, S.A.; van den Berg, D.J.; Tromp, M.N.; Griffioen, D.H.; van Bennekom, W.P.; van der Vijgh, W.J.; Bast, A. Structural aspects of antioxidant activity of flavonoids. Free Radic. Biol. Med. 1996, 20, 331–342. [Google Scholar] [CrossRef]
- Punithavathi, V.R.; Prince, P.S.; Kumar, R.; Selvakumari, J. Antihyperglycaemic, antilipid peroxidative and antioxidant effects of gallic acid on streptozotocin induced diabetic Wistar rats. Eur. J. Pharmacol. 2011, 650, 465–471. [Google Scholar] [CrossRef]
- Li, L.; Ng, T.B.; Gao, W.; Li, W.; Fu, M.; Niu, S.M.; Zhao, L.; Chen, R.R.; Liu, F. Antioxidant activity of gallic acid from rose flowers in senescence accelerated mice. Life Sci. 2005, 77, 230–240. [Google Scholar] [CrossRef]
- Kim, Y.J. Antimelanogenic and antioxidant properties of gallic acid. Biol. Pharm. Bull. 2007, 30, 1052–1055. [Google Scholar] [CrossRef] [Green Version]
- Nabavi, S.M.; Habtemariam, S.; Nabavi, S.F.; Sureda, A.; Daglia, M.; Moghaddam, A.H.; Amani, M.A. Protective effect of gallic acid isolated from Peltiphyllum peltatum against sodium fluoride-induced oxidative stress in rat’s kidney. Mol. Cell Biochem. 2013, 372, 233–239. [Google Scholar] [CrossRef]
- Kerry, N.L.; Abbey, M. Red wine and fractionated phenolic compounds prepared from red wine inhibit low density lipoprotein oxidation in vitro. Atherosclerosis 1997, 135, 93–102. [Google Scholar] [CrossRef]
- Sekher Pannala, A.; Chan, T.S.; O’Brien, P.J.; Rice-Evans, C.A. Flavonoid B-ring chemistry and antioxidant activity: Fast reaction kinetics. Biochem. Biophys. Res. Commun. 2001, 282, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Gawlik-Dziki, U.; Bryda, J.; Dziki, D.; Swieca, M.; Habza-Kowalska, E.; Złotek, U. Impact of Interactions between Ferulic and Chlorogenic Acids on Enzymatic and Non-Enzymatic Lipids Oxidation: An Example of Bread Enriched with Green Coffee Flour. Appl. Sci. 2019, 9, 568. [Google Scholar] [CrossRef] [Green Version]
- Shenoy, A.; Somayaji, S.; Bairy, L. Hepatoprotective effects of Ginko biloba against carbon tetrachloride induced hepatic injury in rats. Indian J. Pharmacol. 2001, 33, 260–266. [Google Scholar]
- Ahr, H.J.; King, L.J.; Nastainczyk, W.; Ullrich, V. The mechanism of reductive dehalogenation of halothane by liver cytochrome P450. Biochem. Pharmacol. 1982, 31, 383–390. [Google Scholar] [CrossRef]
- Maitland, D.J.; Awaad, A.S.; Soliman, G.A. Hepatoprotective activity of Schouwia thebica Webb. Bioorg. Med. Chem. Lett. 2006, 16, 4624–4628. [Google Scholar] [CrossRef]
- Eliza, J.; Daisy, P.; Ignacimuthu, S. Antioxidant activity of costunolide and eremanthin isolated from Costus speciosus (Koen ex. Retz) Sm. Chem. Biol. Interact. 2010, 188, 467–472. [Google Scholar] [CrossRef]
- Venkatraman, J.T.; Angkeow, P.; Satsangi, N.; Fernandes, G. Effects of Dietary n-6 and n-3 Lipids on Antioxidant Defense System in Livers of Exercised Rats. J. Am. Coll. Nutr. 1998, 17, 586–594. [Google Scholar] [CrossRef]
- Pompella, A.; Visvikis, A.; Paolicchi, A.; De Tata, V.; Casini, A.F. The changing faces of glutathione, a cellular protagonist. Biochem. Pharmacol. 2003, 66, 1499–1503. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wang, F.; Zheng, Q.; Lu, L.; Yao, H.; Zhou, C.; Wu, X.; Zhao, Y. Hepatoprotective effect of total flavonoids from Laggera alata against carbon tetrachloride-induced injury in primary cultured neonatal rat hepatocytes and in rats with hepatic damage. J. Biomed. Sci. 2006, 13, 569–578. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types of Method | Adopted Test Methods | Method Summary | Ref. | |
|---|---|---|---|---|
| Assay of non-enzymatic antioxidant compounds | Total phenolic content | Folin Ciocalteu method | [15] | |
| Total flavonoid content | Colorimetric assay based on intensity of colour change of the mixture | [16,17] | ||
| Total flavonol content | ||||
| In-vitro assay | Electron Transfer (ET) based assay | Superoxide radical scavenging assay | Measurement of reducing capacity of antioxidant (αH) in sample based on redox reaction (indicated by colour change) in presence of free radicals (R•) R• + αH + H2O RH + α• + H2O | [3,18,19,20] |
| DPPH• scavenging assay | ||||
| TBARS assay | ||||
| Hydrogen Atom Transfer (HAT) based assay | Hydroxyl radical scavenging assay | Quantitation of hydrogen atom donating capacity of antioxidant present in sample based on proton-coupled reaction (indicated by colour change) in presence of free radicals R• + αH R- + αH+• | [3,21,22,23] | |
| Hydrogen peroxide scavenging assay | ||||
| ABTS+• scavenging assay | ||||
| Others | Anti-hemolytic activity assay | Measurement of inhibiting capacity of RBCs hemolysis by ROS inducing oxidative stress | [24] | |
| In-vivo assay | CAT activity assay | Quantifying the activity of enzymes responsible for suppressing ROS generation such as- catalase (CAT) and superoxide dismutase (SOD) in animal model treated with sample | [25,26] | |
| SOD activity assay | ||||
| LPO activity assay | Measurement of the effect of sample on lipid peroxidation (auto-oxidation) and generation of malondialdehyde (MDA) in mice model | [19] | ||
| Estimation of protein | Assessment of total protein in oxidative stressed animal model treated with sample containing antioxidant | [27] | ||
| Cultivars | Bioactive Compounds Concentration (%) | ||||||
|---|---|---|---|---|---|---|---|
| A | B | C | D | E | F | ||
| BAR | Un | 31.06 ± 6.29 | 4.64 ± 2.10 | 1.07 ± 0.09 | 3.95 ± 2.11 | 1.83 ± 1.01 | 20.04 ± 8.07 |
| P | 5.73 ± 3.38 | 2.22 ± 1.07 | 0.33 ± 0.13 | 4.39 ± 1.92 | 3.58 ± 2.31 | 48.75 ± 19.04 | |
| GUN | Un | 8.52 ± 2.11 | 4.01 ± 2.22 | 0.95 ± 0.77 | 1.12 ± 1.02 | 1.47 ± 0.91 | 16.87 ± 3.92 |
| P | 6.28 ± 2.09 | 1.45 ± 0.77 | 0.35 ± 0.13 | 1.42 ± 1.11 | 1.91 ± 0.22 | 21.73 ± 6.22 | |
| JEN | Un | 25.92 ± 7.20 | 7.52 ± 3.10 | 0.78 ± 0.27 | 2.93 ± 2.00 | 2.73 ± 0.92 | 31.74 ± 6.31 |
| P | 2.87 ± 0.91 | 1.58 ± 1.07 | 0.35 ± 0.03 | 4.09 ± 3.81 | 3.30 ± 1.00 | 37.42 ± 9.28 | |
| JIN | Un | 2.29 ± 1.88 | 0.59 ± 0.23 | 1.21 ± 1.01 | 3.39 ± 0.87 | 3.68 ± 2.13 | 13.60 ± 4.08 |
| P | 1.32 ± 1.01 | 0.35 ± 0.13 | 0.65 ± 0.92 | 4.29 ± 0.02 | 4.70 ± 2.01 | 12.84 ± 4.98 | |
| JUR | Un | 2.91 ± 1.88 | 2.09 ± 1.11 | 2.31 ± 0.21 | 1.44 ± 0.22 | 3.59 ± 3.00 | 18.82 ± 7.30 |
| P | 20.70 ± 2.03 | 3.91 ± 2.08 | 0.25 ± 0.03 | 2.79 ± 1.99 | 4.91 ± 1.98 | 26.44 ± 3.99 | |
| MAN | Un | 18.30 ± 5.31 | 11.73 ± 3.40 | 0.84 ± 0.65 | 3.97 ± 1.00 | 0.72 ± 0.02 | 23.21 ± 6.31 |
| P | 2.89 ± 0.98 | 4.82 ± 2.19 | 1.22 ± 1.13 | 0.73 ± 0.55 | 3.28 ± 2.03 | 27.98 ± 13.21 | |
| LUX | Un | 18.22 ± 3.99 | 0.23 ± 0.16 | 0.74 ± 1.31 | 10.47 ± 3.21 | 0.24 ± 0.11 | 13.18 ± 2.99 |
| P | 2.67 ± 2.02 | 0.59 ± 0.03 | 1.76 ± 1.33 | 1.28 ± 1.10 | 7.08 ± 0.63 | 16.43 ± 1.31 | |
| ROS | Un | 0.23 ± 0.41 | 2.12 ± 2.00 | 2.13 ± 2.09 | 5.22 ± 2.03 | 2.30 ± 1.00 | 1.66 ± 0.51 |
| P | 0.11 ± 0.39 | 0.22 ± 0.17 | 0.94 ± 0.88 | 2.39 ± 0.81 | 5.12 ± 2.62 | 3.22 ± 1.07 | |
| WK | Un | 10.08 ± 1.49 | 3.96 ± 2.18 | 0.54 ± 0.41 | 8.91 ± 2.22 | 6.36 ± 3.19 | 18.33 ± 3.88 |
| P | 3.29 ± 1.00 | 1.24 ± 0.91 | 3.88 ± 2.14 | 0.53 ± 0.44 | 8.75 ± 5.66 | 38.25 ± 14.23 | |
| IC50 Values (mg/mL) | Cultivars/Compounds | ||||||
|---|---|---|---|---|---|---|---|
| MAN | JUR | BAR | GUN | A A | Trolox | ||
| DPPH | Un | 193.8 ± 7.8 | 676.6 ± 16.1 | 591.9 ± 21.9 | 660.9 ± 70.1 | 56.6 ± 2.33 | -- |
| P | 1898 ± 72.4 | 751.3 ± 25.3 | 746.5 ± 18.0 | 811 ± 22.0 | |||
| ABTS | Un | 366 ± 10.3 | -- | 156.2 ± 7.3 | 349.6 ± 8.2 | -- | 137 ± 4.28 |
| P | 511.2 ± 22.2 | -- | 223.2 ± 10.2 | 455.6 ± 14.4 | |||
| LPO | Un | 487.7 ± 34.2 | 323.9 ± 8.4 | 530.8 ± 19.0 | 584 ± 13.8 | 91.3 ± 3.89 | -- |
| P | 369.8 ± 9.3 | 157.7 ± 7.6 | 368 ± 12.9 | 345.3 ± 7.3 | |||
| Hydroxyl | Un | 553.2 ± 29.2 | 723.3 ± 14.9 | 1040 ± 47.5 | 519.5 ± 16.4 | 126.2 ± 4.22 | -- |
| P | 673.1 ± 18.2 | 819 ± 17.2 | 1198 ± 90.9 | 553.2 ± 31.5 | |||
| Anti-hemolytic | Un | 426.2 ± 12.2 | 475.5 ± 13.3 | 580.2 ± 31.2 | 572.1 ± 19.7 | 237.1 ± 2.39 | -- |
| P | 500 ± 20.0 | 716.7 ± 11.8 | 937 ± 35.9 | 802.2 ± 41.0 | |||
| Superoxide | Un | 552.2 ± 28.2 | 535.8 ± 19.3 | 376.2 ± 12.2 | 366.2 ± 11.0 | 276 ± 3.37 | -- |
| P | 604.9 ± 13.3 | 679.8 ± 16.1 | 759 ± 18.3 | 465.1 ± 10.6 | |||
| H2O2 | Un | 376.7 ± 8.2 | 340 ± 31.0 | 521.8 ± 27.7 | 532.4 ± 23.8 | 23.8 ± 1.97 | -- |
| P | 690 ± 12.7 | 561 ± 7.9 | 677.4 ± 22.9 | 883.5 ± 32.3 | |||
| Cultivars | TBARS Assay (Egg-Yolk and Bovine Brain Homogenate) | |||||||
|---|---|---|---|---|---|---|---|---|
| 2 mg/mL | 1 mg/mL | 0.5 mg/mL | 0.25 mg/mL | |||||
| Un | P | Un | P | Un | P | Un | P | |
| JEN 1 | 76.12 ± 2.49 | 93.66 ± 4.01 b | 62.62 ± 1.34 | 91.58 ± 3.02 bd | 58.46 ± 2.11 | 90.18 ± 1.9 bd | 47.76 ± 2.93 | 87.17 ± 3.19 ac |
| MAN 1 | 70.82 ± 3.16 | 93.25 ± 3.98 bc | 69.26 ± 2.02 | 87.22 ± 4.56 a | 45.06 ± 2.62 | 84.47± 3.67 ac | 17.96 ± 2.22 | 59.29 ± 2.09 d |
| MAN 2 | 82.97 ± 2.99 | 88.01 ± 4.12 a | 76.85 ± 2.76 | 83.57 ± 3.76 | 72.78 ± 4.01 | 80.69 ± 2.69 | 70.50 ± 2.67 | 75.06 ± 3.43 |
| BAR 1 | 56.65 ± 2.17 | 88.31 ± 2.88 ad | 44.54 ± 2.09 | 73.05 ± 2.98 | 37.90 ± 3.92 | 67.65 ± 2.93 | 16.77 ± 1.56 | 43.45 ± 4.12 |
| JIN 1 | 61.01 ± 2.26 | 95.53 ± 3.06 bd | 42.93 ± 2.96 | 92.52 ± 2.88 bd | 40.39 ± 2.61 | 87.85 ± 2.64 c | 26.01 ± 2.34 | 68.48 ± 4.67 |
| JIN 2 | 87.65 ± 3.12 a | 88.37 ± 3.09 a | 84.77 ± 1.97 a | 86.45 ± 2.67 a | 78.65 ± 3.96 | 84.4 ± 3.79 a | 62.58 ± 2.83 | 66.43 ± 3.93 |
| WK 1 | 88.73 ± 3.21 a | 90.60 ± 1.99 a | 70.19 ± 3.81 | 84.42 ± 3.71 a | 46.41 ± 1.23 | 70.87 ± 4.04 | 21.65 ± 3.99 | 51.55 ± 3.97 |
| GUN 1 | 87.64 ± 2.38 a | 94.39 ± 2.78 b | 70.9 8 ± 3.52 | 86.55 ± 1.97 a | 54.10 ± 1.12 | 65.93 ± 3.98 | 32.29 ± 2.01 | 50.67 ± 2.78 |
| JUR 1 | 85.72 ± 2.92 | 95.01 ± 4.28 b | 81.88 ± 2.73 | 87.79 ± 1.29 a | 72.89 ± 2.09 | 83.59 ± 3.42 | 11.37 ± 3.73 | 77.62 ± 3.29 c |
| JUR 2 | 88.24 ± 3.34 a | 89.33 ± 2.67 a | 84.05 ± 2.85 a | 87.53 ± 3.43 a | 82.97 ± 3.19 | 85.97± 1.88 a | 79.61 ± 3.09 | 84.53 ± 4.13 a |
| LUX 1 | 86.34 ± 2.71 | 95.11 ± 2.44 b | 51.77 ± 3.02 | 85.61 ± 3.43 ac | 40.18 ± 2.13 | 72.48 ± 2.98 | 23.52 ± 1.46 | 57.89 ± 1.09 |
| ROS 1 | 76.64 ± 2.92 | 95.84 ± 3.78 b | 69.26 ± 4.33 | 92.10 ± 3.91 c | 52.44 ± 2.01 | 88.52 ±1.87 ac | 50.13 ± 2.60 | 70.82 ± 2.09 |
| A A 1 | 92.89 ± 3.04 | 92.88 ± 3.04 | 91.92 ± 2.67 | 91.92 ± 2.67 | 90.52 ± 3.11 | 90.52 ± 3.11 | 89.23 ± 2.87 | 89.23 ± 2.87 |
| A A 2 | 92.88 ± 3.04 | 92.89 ± 3.04 | 91.89 ± 2.67 | 91.85 ± 2.67 | 90.52 ± 3.11 | 90.52 ± 3.11 | 89.23 ± 2.87 | 89.23 ± 2.87 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazumder, K.; Nabila, A.; Aktar, A.; Farahnaky, A. Bioactive Variability and In Vitro and In Vivo Antioxidant Activity of Unprocessed and Processed Flour of Nine Cultivars of Australian lupin Species: A Comprehensive Substantiation. Antioxidants 2020, 9, 282. https://doi.org/10.3390/antiox9040282
Mazumder K, Nabila A, Aktar A, Farahnaky A. Bioactive Variability and In Vitro and In Vivo Antioxidant Activity of Unprocessed and Processed Flour of Nine Cultivars of Australian lupin Species: A Comprehensive Substantiation. Antioxidants. 2020; 9(4):282. https://doi.org/10.3390/antiox9040282
Chicago/Turabian StyleMazumder, Kishor, Afia Nabila, Asma Aktar, and Asgar Farahnaky. 2020. "Bioactive Variability and In Vitro and In Vivo Antioxidant Activity of Unprocessed and Processed Flour of Nine Cultivars of Australian lupin Species: A Comprehensive Substantiation" Antioxidants 9, no. 4: 282. https://doi.org/10.3390/antiox9040282