Potential Antidepressant Effects of Scutellaria baicalensis, Hericium erinaceus and Rhodiola rosea

 , , and
, , and
Abstract
:1. Introduction
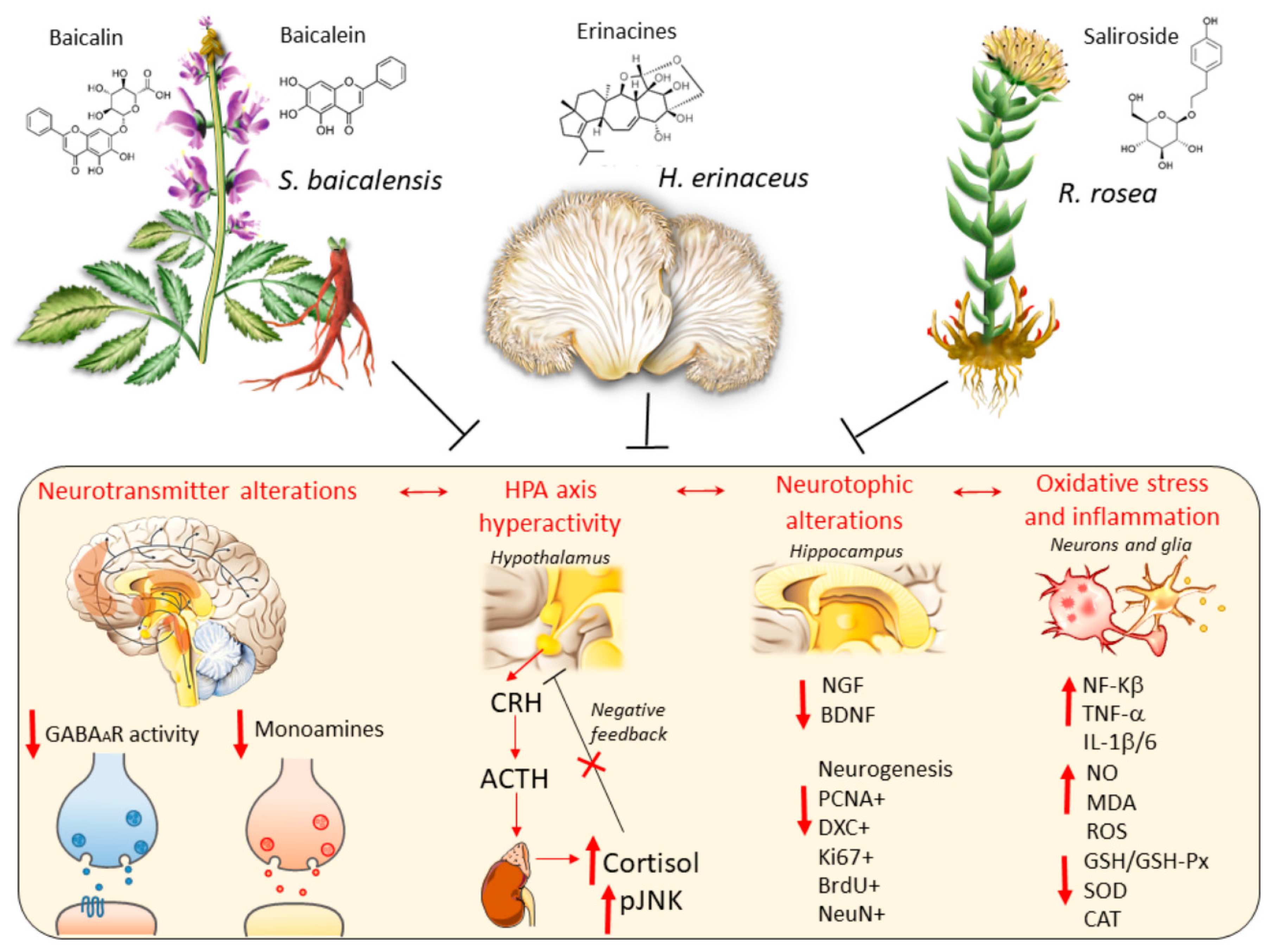
2. S. baicalensis, H. erinaceus and R. rosea: Toxicology and Neuroprotective Effects which may be Relevant for their Antidepressant Potential
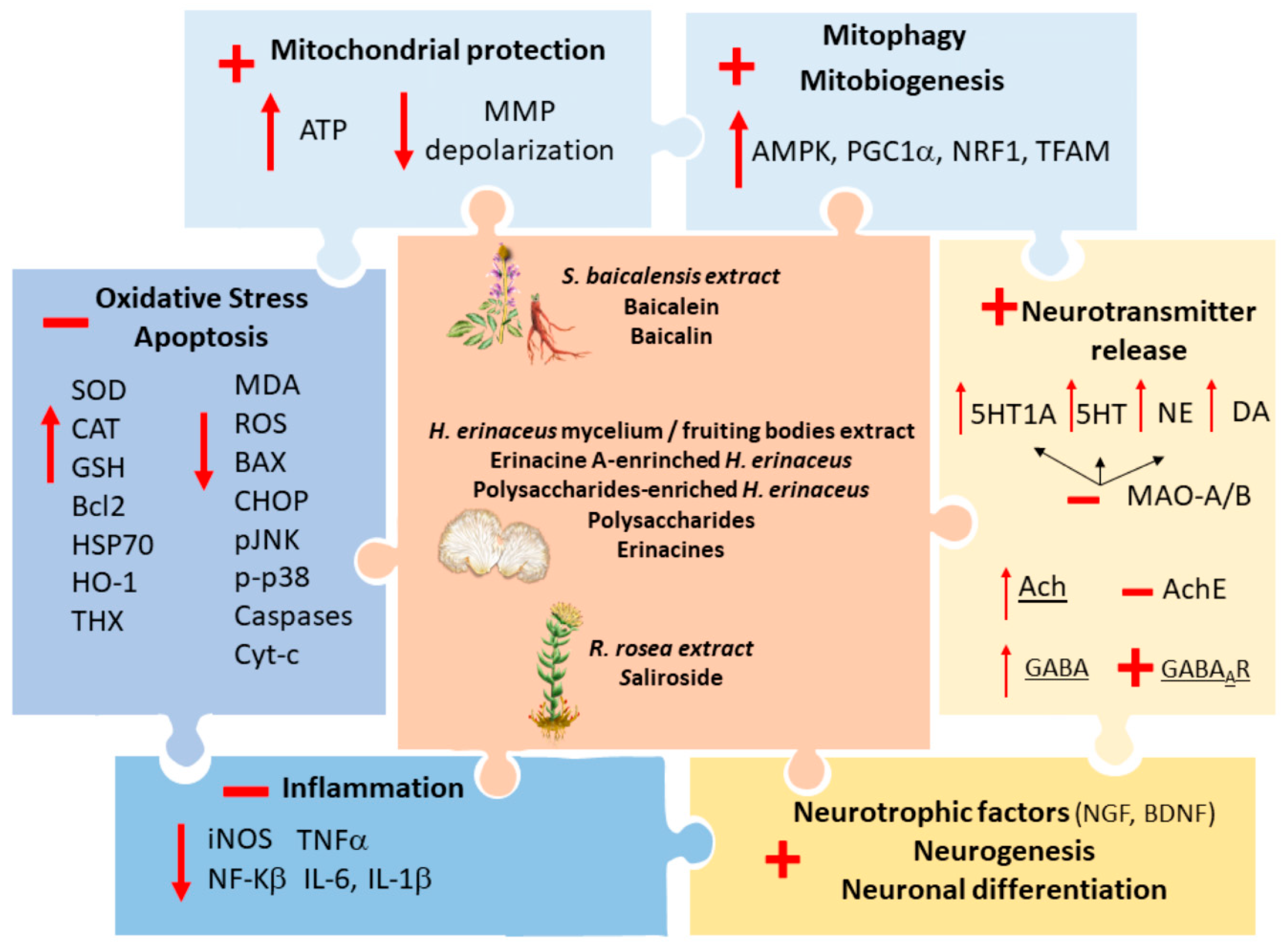
2.1. Common Neuroprotective Effects of S. baicalensis, H. erinaceus and R. rosea
2.2. Complementary Effects of S. baicalensis, H. erinaceus and R. rosea
3. S. baicalensis, H. erinaceus and R. rosea: Anxiolytic/Antidepressant Effects in Experimental and Clinical Studies
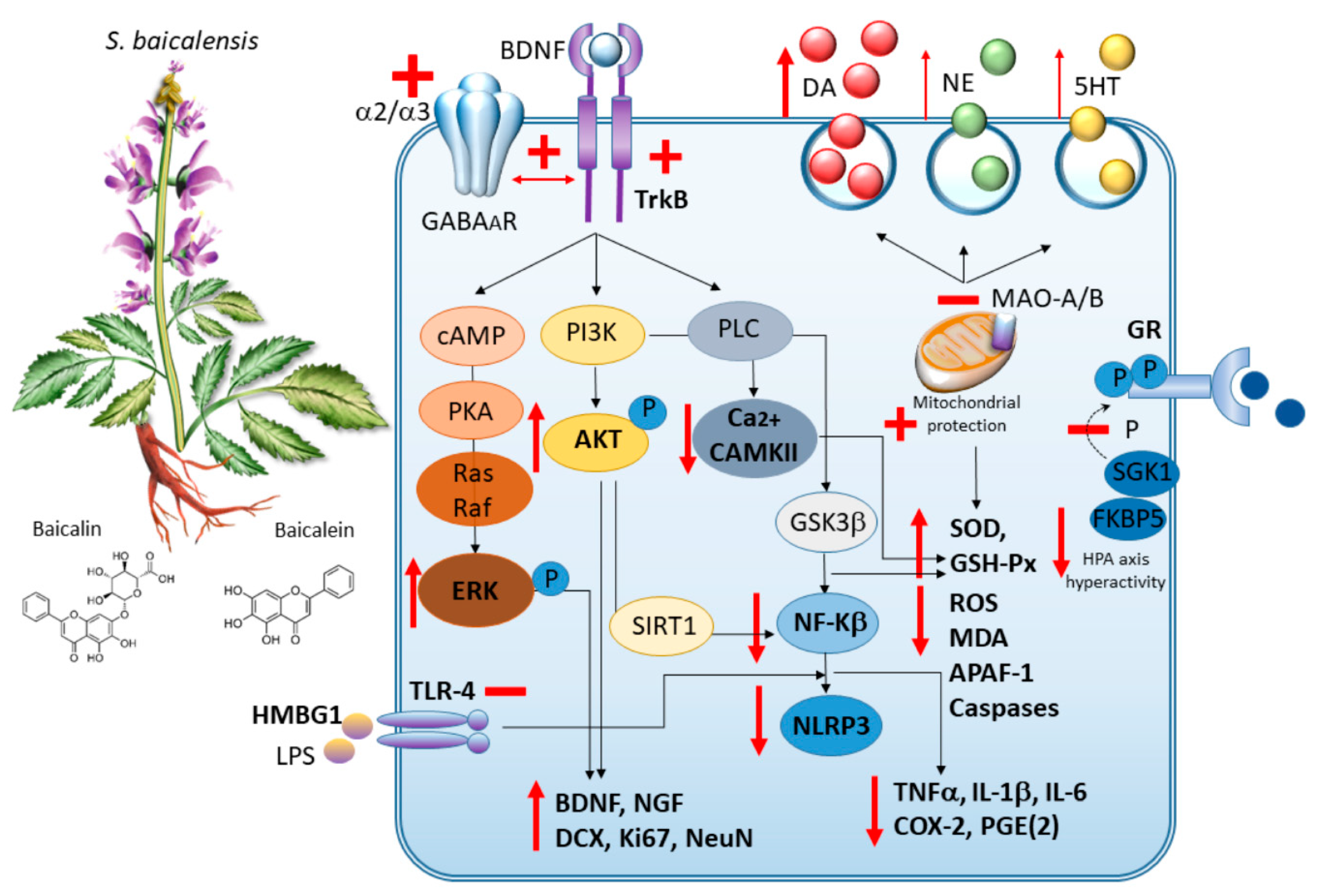
3.1. S. baicalensis in Experimental Models
3.1.1. Anxiety
3.1.2. Chronic Corticosterone-Induced Depression
3.1.3. Olfactory Bulbectomy-Induced Depression
3.1.4. Streptozotocin-Induced Depression
3.1.5. Chronic (unrestraint) Mild Stress-Induced Depression
3.2. S. baicalensis in clinical studies
3.3. H. erinaceus in Experimental Studies
3.3.1. Inflammation-Related Depression
3.3.2. Restraint Stress-Induced Depression
3.4. H. erinaceus in Clinical Studies
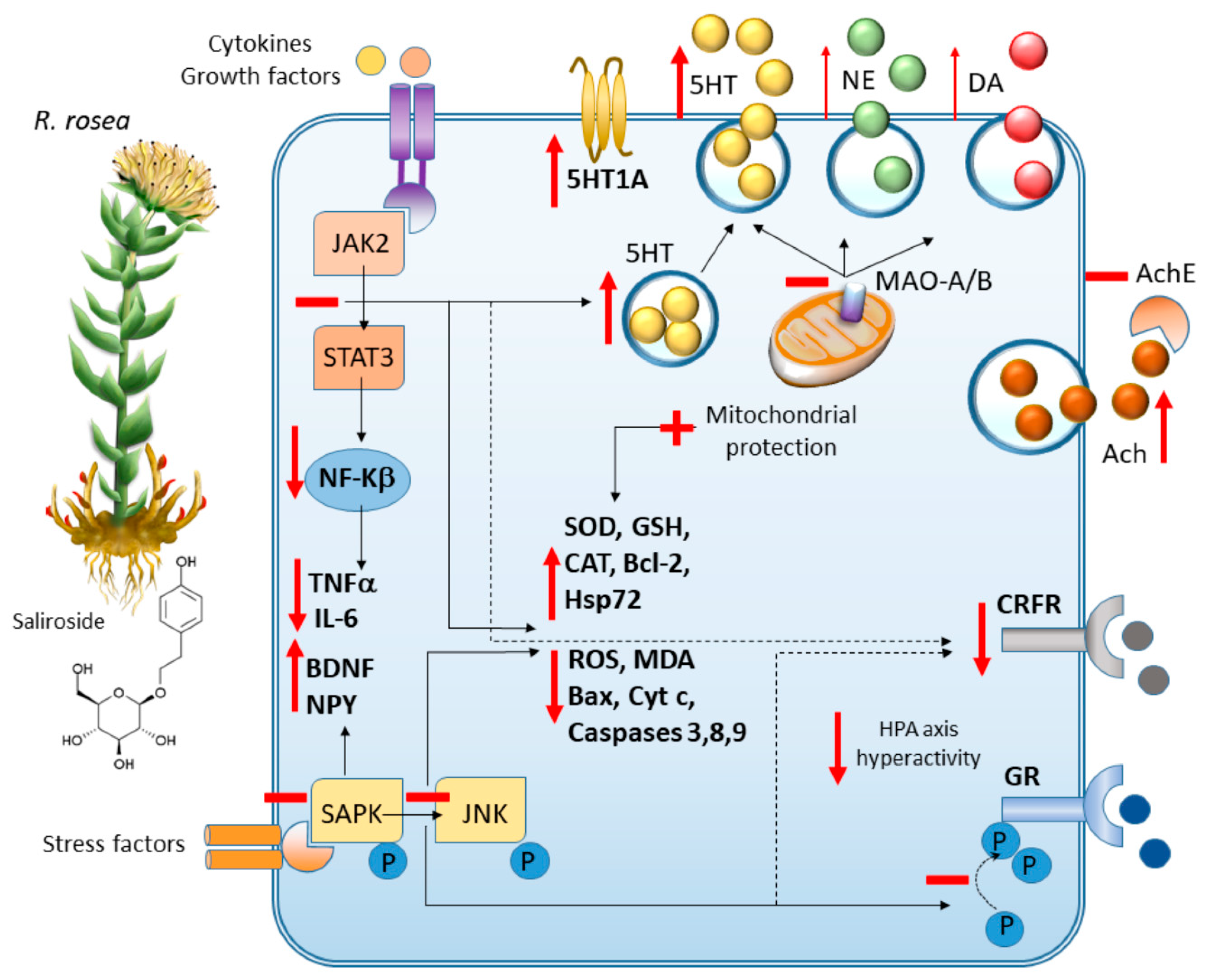
3.5. R. rosea in Experimental Studies
3.5.1. Behavioral Despair-Related Depression
3.5.2. Olfactory Bulbectomy-Induced Depression
3.5.3. Inflammation-Related Depression
3.5.4. Chronic Mild Stress-Induced Depression
3.5.5. Prepulse Inhibition-Related Depression and Psychosis
3.6. R. rosea in Clinical Studies
3.6.1. Anti-Fatigue and Anti-Stress Effects
3.6.2. Anxiolytic and Antidepressant Effects
3.6.3. Obstructive Sleep Apnea- and Menopause-Related Mood Alterations
4. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 6-OHDA | 6-hydroxydopamine |
| AChE | acetylcholinesterase |
| AD | Alzheimer’s disease |
| APPL2 | adaptor protein phosphotyrosine interacting with PH domain and leucine zipper 2 |
| BBB | blood-brain barrier |
| BDI | Beck Depression Inventory |
| BDNF | brain-derived neurotrophic factor |
| BZ | benzodiazepine |
| CAT | catalase |
| CES-D | Center for Epidemiologic Studies Depression Scale |
| CGI | Clinical Global Impression |
| ChAT | acetylcholine transporter |
| CORT | corticosterone |
| COX-2 | cyclooxygenase-2 |
| CRF | corticotrophin-releasing factor |
| CRH | corticotropin-releasing hormone |
| CUMS | chronic (unpredictable) mild stress |
| D1Rs | DA D1 receptors |
| DA | dopamine |
| DAT | DA transporter |
| DCX | doublecortin |
| DPPH | 1-diphenyl-2-picrylhydrazyl |
| DST | Direct Current Transcranial Stimulation |
| GABA | Gamma-Aminobutyric acid |
| GABAAR | GABA A subtype receptor |
| GAD | generalized anxiety disorder |
| GR | glucocorticoid receptor |
| GSH | glutathione |
| GSH-Px | glutathione peroxidase |
| GSK3β | glycogen synthase kinase-3 |
| H2O2 | hydrogen peroxide |
| HAMD | Hamilton Rating Scale for Depression |
| HARS | Hamilton Anxiety Rating Scale |
| HD | Huntington’s disease |
| HMBG1 | High Mobility Group Box 1 |
| HPA | hypothalamic-pituitary-adrenal axis |
| HSP70 | heat shock proteins 70 |
| ICI | Indefinite Complaints Index |
| IL-1β | interleukin 1 beta |
| KMI | Kupperman Menopausal Index |
| LC | Locus Coeruleus |
| LPS | Lipopolysaccharide |
| MAO A/B | monoamine oxidase A/B |
| MDA | malondialdehyde |
| METH | methamphetamine |
| MMP | mitochondrial membrane potential |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| NE | norepinephrine |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NGF | nerve growth factor |
| NLRP3 | nucleotide-binding domain, leucine-rich repeat, pyrin domain containing protein 3 |
| NMDAR/NR2B | N-methyl D-aspartate receptor subtype 2B |
| NPY | neuropeptide Y |
| OBX | olfactory bulbectomy |
| OSA | obstructive sleep apnea |
| PCNA | proliferating cell nuclear antigen |
| PD | Parkinson’s disease |
| pERK | phosphorylated Extracellular signal-regulated kinase |
| PGE(2) | prostaglandin E(2) |
| pJNK | phosphorylated c-Jun N-terminal kinases |
| PSD95 | postsynaptic density protein-95 |
| PSQI | Pittsburgh Sleep Quality Index |
| ROS | reactive oxygen species |
| RS | repeated restraint stress |
| SAS | self-rating anxiety scale |
| SDS | self-rating depression scale |
| SERM | selective oestrogen receptor modulator |
| SOD | superoxide dismutase |
| SYP | synaptophysin |
| TLR4 | toll-like receptor 4 |
| TNF-α | tumor necrosis factor alpha |
| TrkB | tyrosine kinase receptor B |
References
- Liu, L.; Liu, C.; Wang, Y.; Wang, P.; Li, Y.; Li, B. Herbal Medicine for Anxiety, Depression and Insomnia. Curr. Neuropharmacol. 2015, 13, 481–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, K.S.; Hernandez, M.; Mao, J.J.; Haviland, I.; Gubili, J. Herbal medicine for depression and anxiety: A systematic review with assessment of potential psycho-oncologic relevance. Phytother. Res. 2018, 32, 865–891. [Google Scholar] [CrossRef] [PubMed]
- Iovieno, N.; Dalton, E.D.; Fava, M.; Mischoulon, D. Second-tier natural antidepressants: Review and critique. J. Affect. Disord. 2011, 130, 343–357. [Google Scholar] [CrossRef] [PubMed]
- Rzymski, P.; Niedzielski, P.; Siwulski, M.; Mleczek, M.; Budzyńska, S.; Gąsecka, M.; Poniedziałek, B. Lithium biofortification of medicinal mushrooms Agrocybe cylindracea and Hericium erinaceus. J. Food Sci. Technol. 2017, 54, 2387–2393. [Google Scholar] [CrossRef] [PubMed]
- Sowndhararajan, K.; Deepa, P.; Kim, M.; Park, S.J.; Kim, S. Neuroprotective and Cognitive Enhancement Potentials of Baicalin: A Review. Brain Sci. 2018, 8, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, H.I.; Joo, W.S.; Nam, E.; Lee, E.S.; Hwang, Y.J.; Kim, Y.S. Baicalein prevents 6-hydroxydopamine-induced dopaminergic dysfunction and lipid peroxidation in mice. J. Pharmacol. Sci. 2005, 98, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Mu, X.; He, G.R.; Yuan, X.; Li, X.X.; Du, G.H. Baicalein protects the brain against neuron impairments induced by MPTP in C57BL/6 mice. Pharmacol. Biochem. Behav. 2011, 98, 286–291. [Google Scholar] [CrossRef]
- Ma, P.; Mao, X.Y.; Li, X.L.; Ma, Y.; Qiao, Y.D.; Liu, Z.Q.; Zhou, H.H.; Cao, Y.G. Baicalin alleviates diabetes-associated cognitive deficits via modulation of mitogen-activated protein kinase signaling, brain-derived neurotrophic factor and apoptosis. Mol. Med. Rep. 2015, 12, 6377–6383. [Google Scholar] [CrossRef]
- Wang, F.; Xu, Z.; Ren, L.; Tsang, S.Y.; Xue, H. GABA A receptor subtype selectivity underlying selective anxiolytic effect of baicalin. Neuropharmacology 2008, 55, 1231–1237. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, Nutrition, and Health-Promoting Properties of Hericium erinaceus (Lion’s Mane) Mushroom Fruiting Bodies and Mycelia and Their Bioactive Compounds. J. Agric. Food Chem. 2015, 63, 7108–7123. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, Q.; Cui, J.; Wang, L.; Xiong, L.; Wang, W.; Li, D.; Liu, N.; Wu, Y.; Mao, C. Systemic Screening of Strains of the Lion’s Mane Medicinal Mushroom Hericium erinaceus (Higher Basidiomycetes) and Its Protective Effects on Aβ-Triggered Neurotoxicity in PC12 Cells. Int. J. Med. Mushrooms 2015, 17, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.C.; Lu, C.C.; Shen, C.H.; Tung, S.Y.; Hsieh, M.C.; Lee, K.C.; Lee, L.Y.; Chen, C.C.; Teng, C.C.; Huang, W.S.; et al. Hericium erinaceus mycelium and its isolated erinacine A protection from MPTP-induced neurotoxicity through the ER stress, triggering an apoptosis cascade. J. Transl. Med. 2016, 14, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, W.; Zhang, J.C.; Dong, C.; Zhuang, C.; Hirota, S.; Inanaga, K.; Hashimoto, K. Effects of amycenone on serum levels of tumor necrosis factor-α, interleukin-10, and depression-like behavior in mice after lipopolysaccharide administration. Pharmacol. Biochem. Behav. 2015, 136, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.H.; Chyau, C.C.; Chen, C.C.; Lee, L.Y.; Chen, W.P.; Liu, J.L.; Lin, W.H.; Mong, M.C. Erinacine A-Enriched Hericium erinaceus Mycelium Produces Antidepressant-Like Effects through Modulating BDNF/PI3K/Akt/GSK-3β Signaling in Mice. Int. J. Mol. Sci. 2018, 19, 341. [Google Scholar] [CrossRef] [Green Version]
- Amsterdam, J.D.; Panossian, A.G. Rhodiola rosea, L as a putative botanical antidepressant. Phytomedicine 2016, 23, 770–783. [Google Scholar] [CrossRef]
- Lee, Y.; Jung, J.C.; Jang, S.; Kim, J.; Ali, Z.; Khan, I.A.; Oh, S. Anti-Inflammatory and Neuroprotective Effects of Constituents Isolated from Rhodiola rosea. Evid. Based Complement. Altern. Med. 2013, 2013, 514049. [Google Scholar] [CrossRef] [Green Version]
- Jacob, R.; Nalini, G.; Chidambaranathan, N. Neuroprotective effect of Rhodiola rosea Linn against MPTP induced cognitive impairment and oxidative stress. Ann. Neurosci. 2013, 20, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Li, H.; Zhang, J.; Yang, F.; Huang, A.; Deng, J.; Liang, M.; Ma, F.; Hu, M.; Huang, Z. Salidroside protects Caenorhabditis elegans neurons from polyglutamine-mediated toxicity by reducing oxidative stress. Molecules 2014, 19, 7757–7769. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.J.; Yu, H.Y.; Kang, D.Y.; Ma, Z.Q.; Qu, R.; Fu, Q.; Ma, S.P. Antidepressant-like effects of salidroside on olfactory bulbectomy-induced pro-inflammatory cytokine production and hyperactivity of HPA axis in rats. Pharmacol. Biochem. Behav. 2014, 124, 451–457. [Google Scholar] [CrossRef]
- Zhang, X.; Du, Q.; Liu, C.; Yang, Y.; Wang, J.; Duan, S.; Duan, J. Rhodioloside ameliorates depressive behavior via up-regulation of monoaminergic system activity and anti-inflammatory effect in olfactory bulbectomized rats. Int. Immunopharmacol. 2016, 36, 300–304. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, F.; Fan, L.; Zhang, W.; Wang, T.; Du, Y.; Bai, X. Baicalin alleviates atherosclerosis by relieving oxidative stress and inflammatory responses via inactivating the NF-κB and p38 MAPK signaling pathways. Biomed. Pharmacother. 2018, 97, 1673–1679. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, N.; Ismail, S.M.; Aminudin, N.; Shuib, A.S.; Lau, B.F. Evaluation of Selected Culinary-Medicinal Mushrooms for Antioxidant and ACE Inhibitory Activities. Evid. Based Complement. Altern. Med. 2012, 2012, 464238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Z.H.; Ye, J.M.; Wang, G.F. Evaluation of in vivo antioxidant activity of Hericium erinaceus polysaccharides. Int. J. Biol. Macromol. 2013, 52, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.Y.; Abdulla, M.A.; Raman, J.; Phan, C.W.; Kuppusamy, U.R.; Golbabapour, S.; Sabaratnam, V. Gastroprotective Effects of Lion’s Mane Mushroom Hericium erinaceus (Bull.:Fr.) Pers. (Aphyllophoromycetideae) Extract against Ethanol-Induced Ulcer in Rats. Evid. Based Complement. Altern. Med. 2013, 2013, 492976. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Lv, G.; Pan, H.; Pandey, A.; He, W.; Fan, L. Antioxidant and hepatoprotective potential of endo-polysaccharides from Hericium erinaceus grown on tofu whey. Int. J. Biol. Macromol. 2012, 51, 1140–1146. [Google Scholar] [CrossRef]
- Trovato, A.; Siracusa, R.; Di Paola, R.; Scuto, M.; Ontario, M.L.; Bua, O.; Di Mauro, P.; Toscano, M.A.; Petralia, C.C.T.; Maiolino, L.; et al. Redox modulation of cellular stress response and lipoxin A4 expression by Hericium Erinaceus in rat brain: Relevance to Alzheimer’s disease pathogenesis. Immun. Ageing 2016, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Wang, Y.; Zhang, X. Comparative studies on extracts from Hericium erinaceus by different polarity reagents to gain higher antioxidant activities. Exp. Ther. Med. 2016, 12, 513–517. [Google Scholar] [CrossRef] [Green Version]
- Kushairi, N.; Phan, C.W.; Sabaratnam, V.; David, P.; Naidu, M. Lion’s Mane Mushroom, Hericium erinaceus (Bull.: Fr.) Pers. Suppresses H(2)O(2)-Induced Oxidative Damage and LPS-Induced Inflammation in HT22 Hippocampal Neurons and BV2 Microglia. Antioxidants 2019, 8, 261. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Chen, T.; Chang, X.; Zhou, R.; Luo, F.; Liu, J.; Zhang, K.; Wang, Y.; Yang, Y.; Long, H.; et al. Salidroside ameliorates arthritis-induced brain cognition deficits by regulating Rho/ROCK/NF-κB pathway. Neuropharmacology 2016, 103, 134–142. [Google Scholar] [CrossRef]
- Cai, L.; Wang, H.; Li, Q.; Qian, Y.; Yao, W. Salidroside inhibits H2O2-induced apoptosis in PC12 cells by preventing cytochrome c release and inactivating of caspase cascade. Acta Biochim. Biophys. Sin. 2008, 40, 796–802. [Google Scholar] [CrossRef]
- Qu, Z.Q.; Zhou, Y.; Zeng, Y.S.; Li, Y.; Chung, P. Pretreatment with Rhodiola rosea extract reduces cognitive impairment induced by intracerebroventricular streptozotocin in rats: Implication of anti-oxidative and neuroprotective effects. Biomed. Environ. Sci. 2009, 22, 318–326. [Google Scholar] [CrossRef]
- Möhler, H. The GABA system in anxiety and depression and its therapeutic potential. Neuropharmacology 2012, 62, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Czarny, P.; Wigner, P.; Galecki, P.; Sliwinski, T. The interplay between inflammation, oxidative stress, DNA damage, DNA repair and mitochondrial dysfunction in depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 80, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Ma, S.; Qu, R.; Kang, D.; Liu, Y. Antidepressant effect of baicalin extracted from the root of Scutellaria baicalensis in mice and rats. Pharm. Biol. 2006, 44, 503–510. [Google Scholar] [CrossRef]
- Panossian, A.; Nikoyan, N.; Ohanyan, N.; Hovhannisyan, A.; Abrahamyan, H.; Gabrielyan, E.; Wikman, G. Comparative study of Rhodiola preparations on behavioral despair of rats. Phytomedicine 2008, 15, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Guo, Z.; Xie, F.; Zhao, A. Antihyperglycemic and antihyperlipidemic activities of aqueous extract of Hericium erinaceus in experimental diabetic rats. BMC Complement. Altern. Med. 2013, 13, 253. [Google Scholar] [CrossRef] [Green Version]
- Lai, P.L.; Naidu, M.; Sabaratnam, V.; Wong, K.H.; David, R.P.; Kuppusamy, U.R.; Abdullah, N.; Malek, S.N. Neurotrophic properties of the Lion’s mane medicinal mushroom, Hericium erinaceus (Higher Basidiomycetes) from Malaysia. Int. J. Med. Mushrooms 2013, 15, 539–554. [Google Scholar] [CrossRef]
- He, X.; Wang, X.; Fang, J.; Chang, Y.; Ning, N.; Guo, H.; Huang, L.; Huang, X.; Zhao, Z. Structures, biological activities, and industrial applications of the polysaccharides from Hericium erinaceus (Lion’s Mane) mushroom: A review. Int. J. Biol. Macromol. 2017, 97, 228–237. [Google Scholar] [CrossRef]
- Li, I.C.; Lee, L.Y.; Tzeng, T.T.; Chen, W.P.; Chen, Y.P.; Shiao, Y.J.; Chen, C.C. Neurohealth Properties of Hericium erinaceus Mycelia Enriched with Erinacines. Behav. Neurol. 2018, 2018, 5802634. [Google Scholar] [CrossRef] [Green Version]
- Dimpfel, W.; Schombert, L.; Panossian, A.G. Assessing the Quality and Potential Efficacy of Commercial Extracts of Rhodiola rosea L. by Analyzing the Salidroside and Rosavin Content and the Electrophysiological Activity in Hippocampal Long-Term Potentiation, a Synaptic Model of Memory. Front. Pharmacol. 2018, 9, 425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Z.; Han, J.; Zhang, J.; Xiao, Q.; Hu, J.; Chen, L. Pharmacological activities, mechanisms of action, and safety of salidroside in the central nervous system. Drug Des. Dev. Ther. 2018, 12, 1479–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cifani, C.; Micioni Di, B.M.V.; Vitale, G.; Ruggieri, V.; Ciccocioppo, R.; Massi, M. Effect of salidroside, active principle of Rhodiola rosea extract, on binge eating. Physiol. Behav. 2010, 101, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ding, Y.; Zhou, J.; Sun, X.; Wang, S. The in vitro and in vivo antiviral effects of salidroside from Rhodiola rosea L. against coxsackievirus B3. Phytomedicine 2009, 16, 146–155. [Google Scholar] [CrossRef]
- Wu, D.; Yang, X.; Zheng, T.; Xing, S.; Wang, J.; Chi, J.; Bian, F.; Li, W.; Xu, G.; Bai, X.; et al. A novel mechanism of action for salidroside to alleviate diabetic albuminuria: Effects on albumin transcytosis across glomerular endothelial cells. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E225–E237. [Google Scholar] [CrossRef] [Green Version]
- Xing, S.; Yang, X.; Li, W.; Bian, F.; Wu, D.; Chi, J.; Xu, G.; Zhang, Y.; Jin, S. Salidroside stimulates mitochondrial biogenesis and protects against H2O2-induced endothelial dysfunction. Oxidative Med. Cell. Longev. 2014, 2014, 904834. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Pham, V.; Bui, M.; Song, L.; Wu, C.; Walia, A.; Uchio, E.; Smith-Liu, F.; Zi, X. Rhodiola rosea L.: An herb with anti-stress, anti-aging, and immunostimulating properties for cancer chemoprevention. Curr. Pharmacol. Rep. 2017, 3, 384–395. [Google Scholar] [CrossRef]
- Fong, S.Y.K.; Li, C.; Ho, Y.C.; Li, R.; Wang, Q.; Wong, Y.C.; Xue, H.; Zuo, Z. Brain Uptake of Bioactive Flavones in Scutellariae Radix and Its Relationship to Anxiolytic Effect in Mice. Mol. Pharm. 2017, 14, 2908–2916. [Google Scholar] [CrossRef]
- Hu, J.H.; Li, I.C.; Lin, T.W.; Chen, W.P.; Lee, L.Y.; Chen, C.C.; Kuo, C.F. Absolute Bioavailability, Tissue Distribution, and Excretion of Erinacine S in Hericium erinaceus Mycelia. Molecules 2019, 24, 1624. [Google Scholar] [CrossRef] [Green Version]
- Guo, N.; Hu, Z.; Fan, X.; Zheng, J.; Zhang, D.; Xu, T.; Yu, T.; Wang, Y.; Li, H. Simultaneous determination of salidroside and its aglycone metabolite p-tyrosol in rat plasma by liquid chromatography-tandem mass spectrometry. Molecules 2012, 17, 4733–4754. [Google Scholar] [CrossRef] [Green Version]
- Barhwal, K.; Das, S.K.; Kumar, A.; Hota, S.K.; Srivastava, R.B. Insulin receptor A and Sirtuin 1 synergistically improve learning and spatial memory following chronic salidroside treatment during hypoxia. J. Neurochem. 2015, 135, 332–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, T.; Tang, H.; Xie, L.; Zheng, Y.; Ma, Z.; Sun, Q.; Li, X. Scutellaria baicalensis Georgi. (Lamiaceae): a review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. J. Pharm. Pharmacol. 2019, 71, 1353–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, H.; Wu, X.; Cao, J.; Peng, Y.; Wang, A.; Pei, J.; Xiao, J.; Wang, S.; Wang, Y. Rhodiola species: A comprehensive review of traditional use, phytochemistry, pharmacology, toxicity, and clinical study. Med. Res. Rev. 2019, 39, 1779–1850. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.M.; But, P.P.H. (Eds.) Pharmacology and Applications of Chinese Materia Medica; Yeung, S.C.S.; Yao, S.C.; Wang, L.L., Translators; World Scientific: Singapore, 1987; Volume 2, pp. 1022–1028. [Google Scholar]
- Yi, Y.; Zhao, Y.; Li, C.; Zhang, Y.; Bin, Y.; Yuan, Y.; Pan, C.; Wang, L.; Liang, C. Potential chronic liver toxicity in rats orally administered an ethanol extract of Huangqin (Radix Scutellariae baicalensis). J. Tradit. Chin. Med. 2018, 38, 242–256. [Google Scholar]
- Zhang, W.; Song, D.R.; Wang, Y.N.; Zhu, Z. Evaluation of embryotoxicity of baicalin based on embryonic stem cell test system. Chin. J. Pharmacol. Toxicol. 2012, 26, 864–869. [Google Scholar]
- Pang, H.; Xue, W.; Shi, A.; Li, M.; Li, Y.; Cao, G.; Yan, B.; Dong, F.; Xiao, W.; He, G.; et al. Multiple-Ascending-Dose Pharmacokinetics and Safety Evaluation of Baicalein Chewable Tablets in Healthy Chinese Volunteers. Clin. Drug Investig. 2016, 36, 713–724. [Google Scholar] [CrossRef]
- Yimam, M.; Burnett, B.P.; Brownell, L.; Jia, Q. Clinical and Preclinical Cognitive Function Improvement after Oral Treatment of a Botanical Composition Composed of Extracts from Scutellaria baicalensis and Acacia catechu. Behav. Neurol. 2016, 2016, 7240802, Erratum in: Behav. Neurol. 2017, 2017, 3493086. [Google Scholar] [CrossRef]
- Li, I.C.; Chen, W.P.; Chen, Y.P.; Lee, L.Y.; Tsai, Y.T.; Chen, C.C. Acute and developmental toxicity assessment of erincine A-enriched Hericium erinaceus mycelia in Sprague-Dawley rats. Drug Chem. Toxicol. 2018, 1–6. [Google Scholar] [CrossRef]
- Li, I.C.; Chen, Y.L.; Lee, L.Y.; Chen, W.P.; Tsai, Y.T.; Chen, C.C.; Chen, C.S. Evaluation of the toxicological safety of erinacine A-enriched Hericium erinaceus in a 28-day oral feeding study in Sprague-Dawley rats. Food Chem. Toxicol. 2014, 70, 61–67. [Google Scholar] [CrossRef]
- Lee, L.Y.; Li, I.C.; Chen, W.P.; Tsai, Y.T.; Chen, C.C.; Tung, K.C. Thirteen-Week Oral Toxicity Evaluation of Erinacine AEnriched Lion’s Mane Medicinal Mushroom, Hericium erinaceus (Agaricomycetes), Mycelia in Sprague-Dawley Rats. Int. J. Med. Mushrooms 2019, 21, 401–411. [Google Scholar] [CrossRef]
- Vigna, L.; Morelli, F.; Agnelli, G.M.; Napolitano, F.; Ratto, D.; Occhinegro, A.; Di Iorio, C.; Savino, E.; Girometta, C.; Brandalise, F.; et al. Hericium erinaceus Improves Mood and Sleep Disorders in Patients Affected by Overweight or Obesity: Could Circulating Pro-BDNF and BDNF Be Potential Biomarkers? Evid. Based Complement. Altern. Med. 2019, 2019, 7861297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurkin, V.A.; Zapesochnaya, G.G.; Shchavlinskii, A.N.; Nukhi-movskii, E.L.; Vandyshev, V.V. Method of analysis of identity and quality of Rhodiola rosea rhizome. Khim. Farm. Zh. 1985, 19, 185–190. [Google Scholar]
- Montiel-Ruiz, R.M.; Roa-Coria, J.E.; Patiño-Camacho, S.I.; Flores-Murrieta, F.J.; Déciga-Campos, M. Neuropharmacological and toxicity evaluations of ethanol extract from Rhodiola rosea. Drug Dev. Res. 2012, 73, 106–113. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhu, J.; Ma, X.; Tian, Y.; Wan, X.; Zhang, T. Evaluation for developmental toxicity of rhodioside injection in rats. Chin. J. New Drugs 2009, 18, 2068–2071. [Google Scholar]
- Zhu, J.; Wan, X.; Zhu, Y.; Ma, X.; Zheng, Y.; Zhang, T. Evaluation of salidroside in vitro and in vivo genotoxicity. Drug Chem. Toxicol. 2010, 33, 220–226. [Google Scholar] [CrossRef]
- Darbinyan, V.; Aslanyan, G.; Amroyan, E.; Gabrielyan, E.; Malmström, C.; Panossian, A. Clinical trial of Rhodiola rosea L. extract SHR-5 in the treatment of mild to moderate depression. Nord. J. Psychiatry 2007, 61, 343–348, Erratum in: Nord. J. Psychiatry 2007, 61, 503. [Google Scholar] [CrossRef]
- Zhang, X.; Du, L.; Zhang, W.; Yang, Y.; Zhou, Q.; Du, G. Therapeutic effects of baicalein on rotenone-induced Parkinson’s disease through protecting mitochondrial function and biogenesis. Sci. Rep. 2017, 7, 9968. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; He, H.; Chen, L.; Zhang, W.; Zhang, X.; Chen, J. Protective effects of salidroside in the MPTP/MPP(+)-induced model of Parkinson’s disease through ROS-NO-related mitochondrion pathway. Mol. Neurobiol. 2015, 51, 718–728. [Google Scholar] [CrossRef]
- Lebeau, A.; Esclaire, F.; Rostène, W.; Pélaprat, D. Baicalein protects cortical neurons from beta-amyloid (25–35) induced toxicity. Neuroreport 2001, 12, 2199–2202. [Google Scholar] [CrossRef]
- Wang, R.; Shen, X.; Xing, E.; Guan, L.; Xin, L. Scutellaria baicalensis stem-leaf total flavonoid reduces neuronal apoptosis induced by amyloid beta-peptide (25–35). Neural Regen. Res. 2013, 8, 1081–1090. [Google Scholar] [CrossRef]
- Zhao, J.; Lu, S.; Yu, H.; Duan, S.; Zhao, J. Baicalin and ginsenoside Rb1 promote the proliferation and differentiation of neural stem cells in Alzheimer’s disease model rats. Brain Res. 2018, 1678, 187–194. [Google Scholar] [CrossRef]
- Cheng, J.H.; Tsai, C.L.; Lien, Y.Y.; Lee, M.S.; Sheu, S.C. High molecular weight of polysaccharides from Hericium erinaceus against amyloid beta-induced neurotoxicity. BMC Complement. Altern. Med. 2016, 16, 170. [Google Scholar] [CrossRef] [Green Version]
- Tsai-Teng, T.; Chin-Chu, C.; Li-Ya, L.; Wan-Ping, C.; Chung-Kuang, L.; Chien-Chang, S.; Chi-Ying, H.F.; Chien-Chih, C.; Shiao, Y.J. Erinacine A-enriched Hericium erinaceus mycelium ameliorates Alzheimer’s disease-related pathologies in APPswe/PS1dE9 transgenic mice. J. Biomed. Sci. 2016, 23, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; An, S.; Hu, W.; Teng, M.; Wang, X.; Qu, Y.; Liu, Y.; Yuan, Y.; Wang, D. The Neuroprotective Properties of Hericium erinaceus in Glutamate-Damaged Differentiated PC12 Cells and an Alzheimer’s Disease Mouse Model. Int. J. Mol. Sci. 2016, 17, 1810. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.F.; Braidy, N.; Orhan, I.E.; Badiee, A.; Daglia, M.; Nabavi, S.M. Rhodiola rosea L. and Alzheimer’s disease: From farm to pharmacy. Phytother. Res. 2016, 30, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.L.; Su, H.; Tan, Y.F.; Qiu, Y.J.; Zhu, J.P.; Chen, Y.; Lin, S.S.; Wu, M.H.; Mao, Y.P.; Hu, J.J.; et al. Salidroside protects PC-12 cells against amyloid β-induced apoptosis by activation of the ERK1/2 and AKT signaling pathways. Int. J. Mol. Med. 2019, 43, 1769–1777. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yu, H.; Zhao, X.; Lin, X.; Tan, C.; Cao, G.; Wang, Z. Neuroprotective effects of salidroside against beta-amyloid-induced oxidative stress in SH-SY5Y human neuroblastoma cells. Neurochem. Int. 2010, 57, 547–555. [Google Scholar] [CrossRef]
- Li, Q.; Wang, J.; Li, Y.; Xu, X. Neuroprotective effects of salidroside administration in a mouse model of Alzheimer’s disease. Mol. Med. Rep. 2018, 17, 7287–7292. [Google Scholar] [CrossRef]
- Dai, J.; Qiu, Y.M.; Ma, Z.W.; Yan, G.F.; Zhou, J.; Li, S.Q.; Wu, H.; Jin, Y.C.; Zhang, X.H. Neuroprotective effect of baicalin on focal cerebral ischemia in rats. Neural Regen. Res. 2018, 13, 2129–2133. [Google Scholar] [CrossRef]
- Lee, K.F.; Chen, J.H.; Teng, C.C.; Shen, C.H.; Hsieh, M.C.; Lu, C.C.; Lee, K.C.; Lee, L.Y.; Chen, W.P.; Chen, C.C.; et al. Protective effects of Hericium erinaceus mycelium and its isolated erinacine A against ischemia-injury-induced neuronal cell death via the inhibition of iNOS/p38 MAPK and nitrotyrosine. Int. J. Mol. Sci. 2014, 15, 15073–15089. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.H.; Naidu, M.; David, R.P.; Abdulla, M.A.; Abdullah, N.; Kuppusamy, U.R.; Sabaratnam, V. Functional recovery enhancement following injury to rodent peroneal nerve by Lion’s Mane mushroom, Hericium erinaceus (Bull.: Fr.) Pers. (Aphyllophoromycetideae). Int. J. Med. Mushrooms 2009, 11, 225–236. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.F.; Tsai, H.J.; Hung, T.H.; Chen, C.C.; Lee, C.Y.; Wu, C.H.; Wang, P.Y.; Liao, N.C. Salidroside improves behavioral and histological outcomes and reduces apoptosis via PI3K/Akt signaling after experimental traumatic brain injury. PLoS ONE 2012, 7, e45763. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; Chen, Y.; Yew, X.X.; Chen, H.X.; Kim, J.X.; Chang, C.C.; Peng, C.C.; Peng, R.Y. Improvement of erinacine A productivity in Hericium erinaceus mycelia and its neuroprotective bioactivity against the glutamate-insulted apoptosis. LWT Food Sci. Technol. 2016, 65, 1100–1108. [Google Scholar] [CrossRef]
- Li, Q.; Li, Q.Q.; Jia, J.N.; Sun, Q.Y.; Zhou, H.H.; Jin, W.L.; Mao, X.Y. Baicalein Exerts Neuroprotective Effects in FeCl(3)-Induced Posttraumatic Epileptic Seizures via Suppressing Ferroptosis. Front. Pharmacol. 2019, 10, 638. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Gao, F.; Li, X.W.; Jia, R.H.; Meng, X.D.; Zhao, R.; Jing, Y.Y.; Wang, Y.; Jiang, W. The anticonvulsant and neuroprotective effects of baicalin on pilocarpine-induced epileptic model in rats. Neurochem. Res. 2012, 37, 1670–1680. [Google Scholar] [CrossRef] [PubMed]
- Si, P.P.; Zhen, J.L.; Cai, Y.L.; Wang, W.J.; Wang, W.P. Salidroside protects against kainic acid-induced status epilepticus via suppressing oxidative stress. Neurosci. Lett. 2016, 618, 19–24. [Google Scholar] [CrossRef]
- Zhang, L.; Xing, D.; Wang, W.; Wang, R.; Du, L. Kinetic difference of baicalin in rat blood and cerebral nuclei after intravenous administration of Scutellariae Radix extract. J. Ethnopharmacol. 2006, 103, 120–125. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, L.; Wang, X.; Lin, H.; Du, L. Determination of dopamine and its relativity of baicalin in rat nuclei after intravenous administration of flavonoids from Scutellariae radix. Biomed. Chromatogr. 2007, 21, 84–88. [Google Scholar] [CrossRef]
- Shimbo, M.; Kawagishi, H.; Yokogoshi, H. Erinacine A increases catecholamine and nerve growth factor content in the central nervous system of rats. Nutr. Res. 2005, 25, 617–623. [Google Scholar] [CrossRef]
- van Diermen, D.; Marston, A.; Bravo, J.; Reist, M.; Carrupt, P.A.; Hostettmann, K. Monoamine oxidase inhibition by Rhodiola rosea L. roots. J. Ethnopharmacol. 2009, 122, 397–401. [Google Scholar] [CrossRef]
- Zhou, R.; Han, X.; Wang, J.; Sun, J. Baicalin may have a therapeutic effect in attention deficit hyperactivity disorder. Med. Hypotheses 2015, 85, 761–764. [Google Scholar] [CrossRef]
- Zhou, R.; Wang, J.; Han, X.; Ma, B.; Yuan, H.; Song, Y. Baicalin regulates the dopamine system to control the core symptoms of ADHD. Mol. Brain 2019, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.H.; Shen, Y.C.; Wang, Y.H.; Chi, C.W.; Yen, J.C. Baicalein attenuates methamphetamine-induced loss of dopamine transporter in mouse striatum. Toxicology 2006, 226, 238–245. [Google Scholar] [CrossRef]
- Ding, S.; Zhuge, W.; Hu, J.; Yang, J.; Wang, X.; Wen, F.; Wang, C.; Zhuge, Q. Baicalin reverses the impairment of synaptogenesis induced by dopamine burden via the stimulation of GABA(A)R-TrkB interaction in minimal hepatic encephalopathy. Psychopharmacology 2018, 235, 1163–1178. [Google Scholar] [CrossRef] [Green Version]
- Su, G.Y.; Yang, J.Y.; Wang, F.; Ma, J.; Zhang, K.; Dong, Y.X.; Song, S.J.; Lu, X.M.; Wu, C.F. Antidepressant-like effects of Xiaochaihutang in a rat model of chronic unpredictable mild stress. J. Ethnopharmacol. 2014, 152, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Kawagishi, H.; Ando, M.; Sakamoto, H.; Yoshida, S.; Ojima, F.; Ishiguro, Y.; Ukai, N.; Furukawa, S. Hericenones C, D and E, stimulators of nerve growth factor (NGF)-synthesis, from the mushroom Hericium erinaceum. Tetrahedron Lett. 1991, 32, 4561–4564. [Google Scholar] [CrossRef]
- Kawagishi, H.; Ishiyama, D.; Mori, H.; Sakamoto, H.; Ishiguro, Y.; Furukawa, S.; Jingxuan, L. Dictyophorines A and B, two stimulators of NGF-synthesis from the mushroom Dictyophora indusiata. Phytochemistry 1997, 45, 1203–1205. [Google Scholar] [CrossRef]
- Kawagishi, H.; Shimada, A.; Hosokawa, S.; Mori, H.; Sakamoto, H.; Ishiguro, Y.; Sakemi, S.; Bordner, J.; Kojima, N.; Furukawa, S. Erinacines E, F, and G, stimulators of nerve growth factor (NGF)-synthesis, from the mycelia of Hericium erinaceum. Tetrahedron Lett. 1996, 37, 7399–7402. [Google Scholar] [CrossRef]
- Kawagishi, H.; Shimada, A.; Shirai, R.; Okamoto, K.; Ojima, F.; Sakamoto, H.; Ishiguro, Y.; Furukawa, S. Erinacines A, B and C, strong stimulators of nerve growth factor (NGF)-synthesis, from the mycelia of Hericium erinaceum. Tetrahedron Lett. 1994, 35, 1569–1572. [Google Scholar] [CrossRef]
- Kawagishi, H.; Simada, A.; Shizuki, K.; Mori, H.; Okamoto, K.; Sakamoto, H.; Furukawa, S. Erinacine D, a stimulator of NGF-synthesis, from the mycelia of Hericium erinaceum. Heterocycl. Commun. 1996, 2, 51–54. [Google Scholar] [CrossRef]
- Lee, E.W.; Shizuki, K.; Hosokawa, S.; Suzuki, M.; Suganuma, H.; Inakuma, T.; Li, J.; Ohnishi-Kameyama, M.; Nagata, T.; Furukawa, S.; et al. Two novel diterpenoids, erinacines H and I from the Mycelia of Hericium erinaceum. Biosci. Biotechnol. Biochem. 2000, 64, 2402–2405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, B.J.; Shen, J.W.; Yu, H.Y.; Ruan, Y.; Wu, T.T.; Zhao, X. Hericenones and erinacines: Stimulators of nerve growth factor (NGF) biosynthesis in Hericium erinaceus. Mycology 2010, 1, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Hao-Yu, T.; Xia, Y.; Cheng-Chen, Z.; Qian, J.; Jin-Ming, G. Structure Diversity, Synthesis, and Biological Activity of Cyathane Diterpenoids in Higher Fungi. Curr. Med. Chem. 2015, 22, 2375–2391. [Google Scholar] [CrossRef]
- Zhang, C.C.; Cao, C.Y.; Kubo, M.; Harada, K.; Yan, X.T.; Fukuyama, Y.; Gao, J.M. Chemical Constituents from Hericium erinaceus Promote Neuronal Survival and Potentiate Neurite Outgrowth via the TrkA/Erk1/2 Pathway. Int. J. Mol. Sci. 2017, 18, 1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phan, C.W.; Lee, G.S.; Hong, S.L.; Wong, Y.T.; Brkljača, R.; Urban, S.; Abd Malek, S.N.; Sabaratnam, V. Hericium erinaceus (Bull.: Fr) Pers. Cultivated under tropical conditions: Isolation of hericenones and demonstration of NGF-mediated neurite outgrowth in PC12 cells via MEK/ERK and PI3K-Akt signaling pathways. Food Funct. 2014, 5, 3160–3169. [Google Scholar] [CrossRef] [PubMed]
- Phan, C.W.; David, P.; Naidu, M.; Wong, K.H.; Sabaratnam, V. Neurite outgrowth stimulatory effects of culinary-medicinal mushrooms and their toxicity assessment using differentiating Neuro-2a and embryonic fibroblast BALB/3T3. BMC Complement. Altern. Med. 2013, 13, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samberkar, S.; Gandhi, S.; Naidu, M.; Wong, K.H.; Raman, J.; Sabaratnam, V. Lion’s mane, hericium erinaceus and tiger milk, lignosus rhinocerotis (Higher basidiomycetes) medicinal mushrooms stimulate neurite outgrowth in dissociated cells of brain, spinal cord, and retina: An in vitro study. Int. J. Med. Mushrooms 2015, 17, 1047–1054. [Google Scholar] [CrossRef] [Green Version]
- Rupcic, Z.; Rascher, M.; Kanaki, S.; Köster, R.W.; Stadler, M.; Wittstein, K. Two new cyathane diterpenoids from mycelial cultures of the medicinal mushroom Hericium erinaceus and the rare species, Hericium flagellum. Int. J. Mol. Sci. 2018, 19, 740. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Stormmesand, J.; Fang, Z.; Zhu, Q.; Balesar, R.; van Heerikhuize, J.; Sluiter, A.; Swaab, D.; Bao, A.M. Quantification of Tyrosine Hydroxylase and ErbB4 in the Locus Coeruleus of Mood Disorder Patients Using a Multispectral Method to Prevent Interference with Immunocytochemical Signals by Neuromelanin. Neurosci. Bull. 2019, 35, 205–215. [Google Scholar] [CrossRef]
- Zhang, Q.; Hu, D.X.; He, F.; Li, C.Y.; Qi, G.J.; Cai, H.W.; Li, T.X.; Ming, J.; Zhang, P.; Chen, X.Q.; et al. Locus coeruleus-CA1 projections are involved in chronic depressive stress-induced hippocampal vulnerability to transient global ischaemia. Nat. Commun. 2019, 10, 2942. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, F.S.; Ryskalin, L.; Ruffoli, R.; Biagioni, F.; Limanaqi, F.; Ferrucci, M.; Busceti, C.L.; Bonuccelli, U.; Fornai, F. The Neuroanatomy of the Reticular Nucleus Locus Coeruleus in Alzheimer’s Disease. Front. Neuroanat. 2017, 11, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szot, P.; Franklin, A.; Miguelez, C.; Wang, Y.; Vidaurrazaga, I.; Ugedo, L.; Sikkema, C.; Wilkinson, C.W.; Raskind, M.A. Depressive-like behavior observed with a minimal loss of locus coeruleus (LC) neurons following administration of 6-hydroxydopamine is associated with electrophysiological changes and reversed with precursors of norepinephrine. Neuropharmacology 2016, 101, 76–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, K.; Obara, Y.; Moriya, T.; Inatomi, S.; Nakahata, N. Effects of Hericium erinaceus on amyloid β(25–35) peptide-induced learning and memory deficits in mice. Biomed. Res. 2011, 32, 67–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panossian, A.; Wikman, G.; Sarris, J. Rosenroot (Rhodiola rosea): Traditional use, chemical composition, pharmacology and clinical efficacy. Phytomedicine 2010, 17, 481–493. [Google Scholar] [CrossRef]
- Panossian, A.; Hamm, R.; Wikman, G.; Efferth, T. Mechanism of action of Rhodiola, salidroside, tyrosol and triandrin in isolated neuroglial cells: An interactive pathway analysis of the downstream effects using RNA microarray data. Phytomedicine 2014, 21, 1325–1348. [Google Scholar] [CrossRef]
- Mattioli, L.; Titomanlio, F.; Perfumi, M. Effects of a Rhodiola rosea L. extract on the acquisition, expression, extinction, and reinstatement of morphine-induced conditioned place preference in mice. Psychopharmacology 2012, 221, 183–193. [Google Scholar] [CrossRef]
- Mattioli, L.; Funari, C.; Perfumi, M. Effects of Rhodiola rosea L. extract on behavioural and physiological alterations induced by chronic mild stress in female rats. J. Psychopharmacol. 2009, 23, 130–142. [Google Scholar] [CrossRef]
- Vasileva, L.V.; Getova, D.P.; Doncheva, N.D.; Marchev, A.S.; Georgiev, M.I. Beneficial effect of commercial Rhodiola extract in rats with scopolamine-induced memory impairment on active avoidance. J. Ethnopharmacol. 2016, 193, 586–591. [Google Scholar] [CrossRef]
- Hillhouse, B.; Ming, D.S.; French, C.; Towers, G.H. Acetylcholine esterase inhibitors in Rhodiola rosea. Pharm. Biol. 2004, 42, 68–72. [Google Scholar] [CrossRef]
- Chen, Q.G.; Zeng, Y.S.; Qu, Z.Q.; Tang, J.Y.; Qin, Y.J.; Chung, P.; Wong, R.; Hägg, U. The effects of Rhodiola rosea extract on 5-HT level, cell proliferation and quantity of neurons at cerebral hippocampus of depressive rats. Phytomedicine 2009, 16, 830–838. [Google Scholar] [CrossRef]
- Mannucci, C.; Navarra, M.; Calzavara, E.; Caputi, A.P.; Calapai, G. Serotonin involvement in Rhodiola rosea attenuation of nicotine withdrawal signs in rats. Phytomedicine 2012, 19, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Panossian, A.; Hambardzumyan, M.; Hovhanissyan, A.; Wikman, G. The adaptogens rhodiola and schizandra modify the response to immobilization stress in rabbits by suppressing the increase of phosphorylated stress-activated protein kinase, nitric oxide and cortisol. Drug Target Insights 2007, 2, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Panossian, A.; Wikman, G.; Kaur, P.; Asea, A. Adaptogens stimulate neuropeptide y and hsp72 expression and release in neuroglia cells. Front. Neurosci. 2012, 6, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Titomanlio, F.; Perfumi, M.; Mattioli, L. Rhodiola rosea L. extract and its active compound salidroside antagonized both induction and reinstatement of nicotine place preference in mice. Psychopharmacology 2014, 231, 2077–2086. [Google Scholar] [CrossRef] [PubMed]
- Palmeri, A.; Mammana, L.; Tropea, M.R.; Gulisano, W.; Puzzo, D. Salidroside, a bioactive compound of rhodiola rosea, ameliorates memory and emotional behavior in adult mice. J. Alzheimers Dis. 2016, 52, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Pei, L.; Shu, X.; Yang, X.; Yan, T.; Wu, Y.; Wei, N.; Yan, H.; Wang, S.; Yao, C.; et al. Therapeutic Intervention of Learning and Memory Decays by Salidroside Stimulation of Neurogenesis in Aging. Mol. Neurobiol. 2016, 53, 851–866. [Google Scholar] [CrossRef]
- Udintsev, S.N.; Krylova, S.G.; Konovalova, O.N. Correction by natural adaptogens of hormonal-metabolic status disorders in rats during the development of adaptation syndrome using functional tests with dexamethasone and ACTH. Biull. Eksp. Biol. Med. 1991, 112, 599–601. [Google Scholar] [CrossRef]
- Hui, K.M.; Wang, X.H.; Xue, H. Interaction of flavones from the roots of Scutellaria baicalensis with the benzodiazepine site. Planta Med. 2000, 66, 91–93. [Google Scholar] [CrossRef]
- Liao, J.F.; Hung, W.Y.; Chen, C.F. Anxiolytic-like effects of baicalein and baicalin in the Vogel conflict test in mice. Eur. J. Pharmacol. 2003, 464, 141–146. [Google Scholar] [CrossRef]
- Xu, Z.; Wang, F.; Tsang, S.Y.; Ho, K.H.; Zheng, H.; Yuen, C.T.; Chow, C.Y.; Xue, H. Anxiolytic-like effect of baicalin and its additivity with other anxiolytics. Planta Med. 2006, 72, 189–192. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho, R.S.; Duarte, F.S.; de Lima, T.C. Involvement of GABAergic non-benzodiazepine sites in the anxiolytic-like and sedative effects of the flavonoid baicalein in mice. Behav. Brain Res. 2011, 221, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; He, M.; Wang, F.; Zhang, H.; Li, Y.; Yang, J.; Wu, C. Revealing Antidepressant Mechanisms of Baicalin in Hypothalamus Through Systems Approaches in Corticosterone- Induced Depressed Mice. Front. Neurosci. 2019, 13, 834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Pan, X.; Wang, F.; Ma, J.; Su, G.; Dong, Y.; Yang, J.; Wu, C. Baicalin promotes hippocampal neurogenesis via SGK1- and FKBP5-mediated glucocorticoid receptor phosphorylation in a neuroendocrine mouse model of anxiety/depression. Sci. Rep. 2016, 6, 30951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Zhang, F.; Guan, X. Baicalin reverse depressive-like behaviors through regulation SIRT1-NF-kB signalling pathway in olfactory bulbectomized rats. Phytother. Res. 2019, 33, 1480–1489. [Google Scholar] [CrossRef]
- Yu, H.Y.; Yin, Z.J.; Yang, S.J.; Ma, S.P.; Qu, R. Baicalin Reverses Depressive-Like Behaviours and Regulates Apoptotic Signalling Induced by Olfactory Bulbectomy. Phytother. Res. 2016, 30, 469–475. [Google Scholar] [CrossRef]
- Gao, C.; Du, Q.; Li, W.; Deng, R.; Wang, Q.; Xu, A.; Shen, J. Baicalin Modulates APPL2/Glucocorticoid Receptor Signaling Cascade, Promotes Neurogenesis, and Attenuates Emotional and Olfactory Dysfunctions in Chronic Corticosterone-Induced Depression. Mol. Neurobiol. 2018, 55, 9334–9348. [Google Scholar] [CrossRef]
- Zhong, J.; Li, G.; Xu, H.; Wang, Y.; Shi, M. Baicalin ameliorates chronic mild stress-induced depression-like behaviors in mice and attenuates inflammatory cytokines and oxidative stress. Braz. J. Med. Biol. Res. 2019, 52, e8434. [Google Scholar] [CrossRef]
- Liu, L.; Dong, Y.; Shan, X.; Li, L.; Xia, B.; Wang, H. Anti-Depressive Effectiveness of Baicalin In Vitro and In Vivo. Molecules 2019, 24, 326. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.T.; Wang, S.Q.; Su, J.; Xu, L.X.; Ji, Z.Y.; Zhang, R.Y.; Zhao, Q.W.; Ma, Z.Q.; Deng, X.Y.; Ma, S.P. Baicalin ameliorates neuroinflammation-induced depressive-like behavior through inhibition of toll-like receptor 4 expression via the PI3K/AKT/FoxO1 pathway. J. Neuroinflammation 2019, 16, 95. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.Y.; Zeng, M.J.; Zhou, L.P.; Li, Y.Q.; Zhao, F.; Shang, Z.Y.; Deng, X.Y.; Ma, Z.Q.; Fu, Q.; Ma, S.P.; et al. Baicalin exerts neuroprotective effects via inhibiting activation of GSK3β/NF-κB/NLRP3 signal pathway in a rat model of depression. Int. Immunopharmacol. 2018, 64, 175–182. [Google Scholar] [CrossRef]
- Liu, X.; Liu, C. Baicalin ameliorates chronic unpredictable mild stress-induced depressive behavior: Involving the inhibition of NLRP3 inflammasome activation in rat prefrontal cortex. Int. Immunopharmacol. 2017, 48, 30–34. [Google Scholar] [CrossRef]
- Lu, Y.; Sun, G.; Yang, F.; Guan, Z.; Zhang, Z.; Zhao, J.; Liu, Y.; Chu, L.; Pei, L. Baicalin regulates depression behavior in mice exposed to chronic mild stress via the Rac/LIMK/cofilin pathway. Biomed. Pharmacother. 2019, 116, 109054. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Shen, J.D.; Li, J.; Wang, R.; Jiao, S.; Yi, L.T. Chronic treatment with baicalin prevents the chronic mild stress-induced depressive-like behavior: Involving the inhibition of cyclooxygenase-2 in rat brain. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 40, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Ma, Z.; Liu, K.; Li, Y.; Liu, D.; Xu, L.; Deng, X.; Qu, R.; Ma, Z.; Ma, S. Baicalin exerts antidepressant effects through Akt/FOXG1 pathway promoting neuronal differentiation and survival. Life Sci. 2019, 221, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Guo, L.; Ji, Z.; Li, X.; Zhang, C.; Ma, Z.; Fu, Q.; Qu, R.; Ma, S. Radix Scutellariae Attenuates CUMS-Induced Depressive-Like Behavior by Promoting Neurogenesis via cAMP/PKA Pathway. Neurochem. Res. 2018, 43, 2111–2120. [Google Scholar] [CrossRef]
- Mori, K.; Inatomi, S.; Ouchi, K.; Azumi, Y.; Tuchida, T. Improving effects of the mushroom Yamabushitake (Hericium erinaceus) on mild cognitive impairment: A double-blind placebo-controlled clinical trial. Phytother. Res. 2009, 23, 367–372. [Google Scholar] [CrossRef]
- Chong, P.S.; Fung, M.L.; Wong, K.H.; Lim, L.W. Therapeutic Potential of Hericium erinaceus for Depressive Disorder. Int. J. Mol. Sci. 2019, 21, 163. [Google Scholar] [CrossRef] [Green Version]
- Ryu, S.; Kim, H.G.; Kim, J.Y.; Kim, S.Y.; Cho, K.O. Hericium erinaceus Extract Reduces Anxiety and Depressive Behaviors by Promoting Hippocampal Neurogenesis in the Adult Mouse Brain. J. Med. Food. 2018, 21, 174–180. [Google Scholar] [CrossRef]
- Nagano, M.; Shimizu, K.; Kondo, R.; Hayashi, C.; Sato, D.; Kitagawa, K.; Ohnuki, K. Reduction of depression and anxiety by 4 weeks Hericium erinaceus intake. BioMed Res. 2010, 31, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Inanaga, K. Marked improvement of neurocognitive impairment after treatment with compounds from Hericium erinaceum: A case study of recurrent depressive disorder. Pers. Med. Universe 2014, 3, 46–48. [Google Scholar] [CrossRef]
- Okamura, H.; Anno, N.; Tsuda, A.; Inokuchi, T.; Uchimura, N.; Inanaga, K. The effects of Hericium erinaceus (Amyloban® 3399) on sleep quality and subjective well-being among female undergraduate students: A pilot study. Pers. Med. Universe 2015, 4, 76–78. [Google Scholar] [CrossRef]
- Zhu, L.; Wei, T.; Gao, J.; Chang, X.; He, H.; Miao, M.; Yan, T. Salidroside attenuates lipopolysaccharide (LPS) induced serum cytokines and depressive-like behavior in mice. Neurosci. Lett. 2015, 606, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Vasileva, L.V.; Saracheva, K.E.; Ivanovska, M.V.; Petrova, A.P.; Sucouglu, E.; Murdjeva, M.A.; Getova-Spasova, D.P. Beneficial Effect of Chronic Treatment with Extracts from Rhodiola Rosea L. and Curcuma Longa L. on the Immunoreactivity of Animals Subjected to a Chronic Mild Stress Model. Folia Med. 2017, 59, 443–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamel Ismail, Z.M.; Morcos, M.A.; Eldin Mohammad, M.D.; Gamal Aboulkhair, A. Enhancement of Neural Stem Cells after Induction of Depression in Male Albino Rats (A histological & Immunohistochemical Study). Int. J. Stem Cells 2014, 7, 70–78. [Google Scholar] [PubMed] [Green Version]
- Coors, A.; Brosch, M.; Kahl, E.; Khalil, R.; Michels, B.; Laub, A.; Franke, K.; Gerber, B.; Fendt, M. Rhodiola rosea root extract has antipsychotic-like effects in rodent models of sensorimotor gating. J. Ethnopharmacol. 2019, 235, 320–328. [Google Scholar] [CrossRef]
- Concerto, C.; Infortuna, C.; Muscatello, M.R.A.; Bruno, A.; Zoccali, R.; Chusid, E.; Aguglia, E.; Battaglia, F. Exploring the effect of adaptogenic Rhodiola Rosea extract on neuroplasticity in humans. Complement. Ther. Med. 2018, 41, 141–146. [Google Scholar] [CrossRef]
- Darbinyan, V.; Kteyan, A.; Panossian, A.; Gabrielian, E.; Wikman, G.; Wagner, H. Rhodiola rosea in stress induced fatigue – a double blind cross-over study of a standardized extract SHR-5 with a repeated low-dose regimen on the mental performance of healthy physicians during night duty. Phytomedicine 2000, 7, 365–371. [Google Scholar] [CrossRef]
- Spasov, A.A.; Wikman, G.K.; Mandrikov, V.B.; Mironova, I.A.; Neumoin, V.V. A double-blind, placebo-controlled pilot study of the stimulating and adaptogenic effect of Rhodiola rosea SHR-5 extract on the fatigue of students caused by stress during an examination period with a repeated low-dose regimen. Phytomedicine 2000, 7, 85–89. [Google Scholar] [CrossRef]
- Shevtsov, V.A.; Zholus, B.I.; Shervarly, V.I.; Vol’skij, V.B.; Korovin, Y.P.; Khristich, M.P.; Roslyakova, N.A.; Wikman, G. A randomized trial of two different doses of a SHR-5 Rhodiola rosea extract versus placebo and control of capacity for mental work. Phytomedicine 2003, 10, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Bystritsky, A.; Kerwin, L.; Feusner, J.D. A pilot study of Rhodiola rosea (Rhodax) for generalized anxiety disorder (GAD). J. Altern. Complement. Med. 2008, 14, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Brichenko, V.S.; Kupriyanova, I.E.; Skorokhodova, T.F. The use of herbal adaptogens together with tricyclic antidepressants in patients with psychogenic depressions. In Modern Problems of Pharmacology and Search for New Medicines; Goldsberg ED, Tomsk, Russia, 1986; Volume 2, pp. 58–60.
- Brichenko, V.S.; Skorokhodova, T.F. Herbal Adaptogens in Rehabilitation of Patients with depression. In Clinical and Organisational Aspects of early Manifestations of Nervous and Mental diseases; Altajskoe Kniže: Barnaul, Russia, 1987; p. 15. [Google Scholar]
- Mao, J.J.; Li, Q.S.; Soeller, I.; Xie, S.X.; Amsterdam, J.D. Rhodiola rosea therapy for major depressive disorder: A study protocol for a randomized, double-blind, placebo-controlled trial. J. Clin. Trials 2014, 4, 170. [Google Scholar] [PubMed] [Green Version]
- Mao, J.J.; Xie, S.X.; Zee, J.; Soeller, I.; Li, Q.S.; Rockwell, K.; Amsterdam, J.D. Rhodiola rosea versus sertraline for major depressive disorder: A randomized placebo-controlled trial. Phytomedicine 2015, 22, 394–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cropley, M.; Banks, A.P.; Boyle, J. The Effects of Rhodiola rosea L. Extract on Anxiety, Stress, Cognition and Other Mood Symptoms. Phytother. Res. 2015, 29, 1934–1939. [Google Scholar] [CrossRef] [PubMed]
- Bangratz, M.; Ait Abdellah, S.; Berlin, A.; Blondeau, C.; Guilbot, A.; Dubourdeaux, M.; Lemoine, P. A preliminary assessment of a combination of rhodiola and saffron in the management of mild-moderate depression. Neuropsychiatr. Dis. Treat. 2018, 14, 1821–1829. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.L.; Zhang, P.P.; Zhang, C.; Zhang, X.; Li, Z.Z.; Li, W.Q.; Fu, A.S. Effects of rhodiola rosea on oxidative stress and negative emotional states in patients with obstructive sleep apnea. Lin Chung Er Bi Yan Hou Tou Jing Wai Ke Za Zhi 2019, 33, 954–957. [Google Scholar] [CrossRef] [PubMed]
- Gerbarg, P.L.; Brown, R.P. Pause menopause with Rhodiola rosea, a natural selective estrogen receptor modulator. Phytomedicine 2016, 23, 763–769. [Google Scholar] [CrossRef]
- Limanaqi, F.; Biagioni, F.; Ryskalin, L.; Busceti, C.L.; Fornai, F. Molecular Mechanisms Linking ALS/FTD and Psychiatric Disorders, the Potential Effects of Lithium. Front. Cell. Neurosci. 2019, 13, 450. [Google Scholar] [CrossRef]
- Galts, C.P.C.; Bettio, L.E.B.; Jewett, D.C.; Yang, C.C.; Brocardo, P.S.; Rodrigues, A.L.S.; Thacker, J.S.; Gil-Mohapel, J. Depression in neurodegenerative diseases: Common mechanisms and current treatment options. Neurosci. Biobehav. Rev. 2019, 102, 56–84. [Google Scholar] [CrossRef]
- Hargreaves, I.P.; Al Shahrani, M.; Wainwright, L.; Heales, S.J. Drug-Induced Mitochondrial Toxicity. Drug Saf. 2016, 39, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharmacol. 2014, 4, 177. [Google Scholar] [CrossRef] [Green Version]
- Limanaqi, F.; Biagioni, F.; Busceti, C.L.; Ryskalin, L.; Polzella, M.; Frati, A.; Fornai, F. Phytochemicals Bridging Autophagy Induction and Alpha-Synuclein Degradation in Parkinsonism. Int. J. Mol. Sci. 2019, 20, 3274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Zhang, J.; Qian, J.; Wu, G.; Ma, Z. Baicalin attenuates liver hypoxia/reoxygenation injury by inducing autophagy. Exp. Ther. Med. 2018, 16, 657–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, J.; Zhu, Y.; Wang, H.; Cao, B.; Fei, M.; Niu, W.; Zhou, Y.; Wang, X.; Li, X.; Zhou, M. Baicalin Protects Mice Brain From Apoptosis in Traumatic Brain Injury Model Through Activation of Autophagy. Front. Neurosci. 2019, 12, 1006. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Cai, F.; Wang, J.; Yang, Z.; Gu, C.; Wang, G.; Mao, G.; Yan, J. Salidroside protects SH-SY5Y from pathogenic α-synuclein by promoting cell autophagy via mediation of mTOR/p70S6K signaling. Mol. Med. Rep. 2019, 20, 529–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gassen, N.C.; Rein, T. Is There a Role of Autophagy in Depression and Antidepressant Action? Front. Psychiatry 2019, 10, 337. [Google Scholar] [CrossRef] [PubMed]
- Ryskalin, L.; Limanaqi, F.; Frati, A.; Busceti, C.L.; Fornai, F. mTOR-Related Brain Dysfunctions in Neuropsychiatric Disorders. Int. J. Mol. Sci. 2018, 19, 2226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limanaqi, F.; Biagioni, F.; Gambardella, S.; Ryskalin, L.; Fornai, F. Interdependency Between Autophagy and Synaptic Vesicle Trafficking: Implications for Dopamine Release. Front. Mol. Neurosci. 2018, 11, 299. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Models of Depression | S. baicalensis | H. erinaceus | R. rosea |
|---|---|---|---|
| Chronic corticosterone-induced depression (CORT) | Baicalin (40, 80, and 160 mg/kg) [133,134] ↑cytoplasmic GR levels ↓nuclear GR levels ↓GR phosphorylation ↑negative feedback of HPA axis [133] ↑neurogenesis (Ki67- DCX-positive cells) in the dentate gyrus of the hippocampus [134]. | ||
| Olfactory bulbectomy-induced depression (OBX) | Baicalin (20 and 40 mg/kg) [135,136,137] ↓serum corticosterone [135] ↓IL-1β, IL-6, and TNF-α in the brain via inhibition of the SIRT1-NF-kB [135] ↓oxidative stress and apoptosis [136] ↑GSH-Px and ↓MDA, APAF-1 and caspases ↑neurogenesis and olfactory function via APPL2/GR signaling pathway [137]. | Saliroside (20 and 40 mg/kg) [19,20] ↓TNF-α, IL-1β, IL-6 and NF-κB in the hippocampus and prefrontal cortex ↑BDNF expression in the hippocampus [19,20] ↑5-HT and NE levels in the prefrontal cortex [20] ↓CRF expression in hypothalamus and ↓serum corticosterone levels [19]. | |
| Chronic (unrestraint) mild stress-induced depression (CUMS) and Chronic mild stress + inflammation (CUMS+LPS) | Baicalin (10, 20, 25, 40, 50, 60, and 100 mg/kg) [35,138,139,140,141,142,143,144,145] ↓monoamine oxidase A and B (MAO A/B) activity [35] ↓IL-1β, IL-6, and TNF-α in serum and in the hippocampus [138,139] ↓COX-2 and PGE(2) in the brain [144] ↓TLR4 via the HMBG1/NF-kb and PI3K/AKT/FoxO1 pathways [139,140]. ↑SOD and ↓ROS, MDA and caspase 1 [138,141] via ↓NMDAR/NR2B, ↓Ca2+/CaMPK-II and ↑pERK [139], and ↓GSK3β/ NF-κB / NLRP3 [141,142] ↓ultrastructural hippocampal alterations ↑synaptic proteins SYP PSD95, TrkB, Rac1, cofilin ↑BDNF [143] ↑neurogenesis (DCX-positive cells) ↑neuronal maturation, differentiation and survival [145]. Radix Scuellariae extract (500 and 1000 mg/kg) [146] ↑cAMP/PKA-dependent neurogenesis, ↑BrdU, DCX and NeuN in the mice hippocampi [146]. | R. rosea extract (1.5 g/kg/day) [155] ↑neuronal stem cell proliferation ↑PCNA-positive cells [155] ↓Caspase 3-positive neuronal cells [155]. R. rosea extract (1.5, 3 and 6 g/kg) [121] ↑5-HT hippocampal levels already at the lowest dose. ↑neural stem cell proliferation ↑BrdU-positive cells [121]. R. rosea extract (250 and 500 mg/kg) [154] ↓TNF-α and IL-6 in CMS exposed and LPS-challenged mice [154] Potentiated effects when administered with curcuma longa (500 mg/kg) [154]. | |
| Streptozotocin (diabetes)-induced depression | Baicalin (50, 100 and 200 mg/kg) [8] ↑hippocampal acetylcholine transporter (ChAT) ↓acetylcholinesterase (AChE) ↑pERK) ↑Bcl 2 ↑BDNF ↓pJNK) ↓p-p38, ↓caspase 3 and ↓Bax [8]. | ||
| Inflammation (LPS)-induced depression | Baicalin (25, 50 and 60 mg/kg) [139,140] ↓TLR4 via the HMBG1/NF-kb and PI3K/AKT/FoxO1 pathways [139,140]. | H. erinaceus fruit body extract amycenone (200mg/kg) [13] ↓serum TNF-α ↑IL-10 [13]. | Saliroside (12 and 24 mg/kg) [153] ↓serum IL-6 and TNF-α ↓NF-κB in the brain ↑TrkB/BDNF levels ↑5-HT and NE levels in the prefrontal cortex [153]. |
| Restraint stress-induced depression | H. erinaceus mycelium ethanolic extract (100, 200 and 400 mg7kg) [14] ↑monoamines levels ↓IL-6, TNF-α and NF-κB ↑BDNF [14]. | ||
| Clinical studies | S. baicalensis | H. erinaceus | R.rosea |
| Baicalein chewable tablets (200, 400, and 800 mg once daily on days 1 and 10, and twice daily on days 3–9). Safe and well tolerated. Related mood effects were not analysed/reported [57]. S. baicalensis extract (300 mg daily for 30 days). Safe and well tolerated. Marked improvement in speed and accuracy of processing complex information in computer tasks [58]. | H. erinaceus cookies (0.5 g of fruit bodies powder for 4 weeks). Lower scores associated with insensitivity, agitation, irritation, palpitation and anxiety in H. erinaceus-receiving women compared with placebo group [155]. H. erinaceus (1.650 g/day, 80% mycelium extract and 20% fruiting body extract for 8 weeks). Safe and well tolerated. Coupled with a low calorie diet improves depression, anxiety, sleep, and binge eating compared with subjects undergoing low calorie diet only [91]. Increases circulating pro-BDNF levels and pro-BDNF/BDNF ratio [91]. H. erinaceus extract (AmylobanⓇ) daily for 6 months combined with Mirtazapine. Combats depression, and improves cognitive function and body weight in the absence of adverse reactions [151]. H. erinaceus extract (AmylobanⓇ3399) intake for 4 weeks counteracts sleep disturbances in a pilot study on female undergraduate students. It increases the levels of salivary free- 3-methoxy-4-hydroxyphenylglycol, an index of chronic stress and depressive symptoms reflecting sympathetic nervous system activity [152]. | R. rosea extract (340 mg/day for 10 weeks). Improvement of general anxiety disorder symptoms evaluated by HARS scores. Generally mild adverse effects, the most common being dizziness and dry mouth [161]. R. rosea extract (SHR-5, 340 or 680 mg/day for six weeks). Safe and well tolerated. Improvement of depressive symptoms, insomnia, emotional instability and somatization compared with placebo group [69]. R. rosea powdered extract (SHR-5 capsule, 340 mg, one capsule/day for the first week, two capsules/day for the following two weeks, up to up to 4 capsules/day for the last six weeks). Improves depressive symptoms compared with placebo and produces antidepressant effects which are comparable with sertraline (50 mg). Fewer adverse effects were reported for R. rosea compared with sertraline [164,165]. R. rosea extract (Vitano®, 200 mg twice a day for 14 days). Safe and well tolerated. Reduces self-reported anxiety, stress, anger, confusion and depression, and overall improvement in mood [166] R. rosea extract (one tablet, 154mg, combined with saffron tablet 15 mg, twice a day for 6 weeks). Excellent safety, no serious adverse effects. Improvements in HARS scores and depression symptoms reported by both general practitioners and patients [167]. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Limanaqi, F.; Biagioni, F.; Busceti, C.L.; Polzella, M.; Fabrizi, C.; Fornai, F. Potential Antidepressant Effects of Scutellaria baicalensis, Hericium erinaceus and Rhodiola rosea. Antioxidants 2020, 9, 234. https://doi.org/10.3390/antiox9030234
Limanaqi F, Biagioni F, Busceti CL, Polzella M, Fabrizi C, Fornai F. Potential Antidepressant Effects of Scutellaria baicalensis, Hericium erinaceus and Rhodiola rosea. Antioxidants. 2020; 9(3):234. https://doi.org/10.3390/antiox9030234
Chicago/Turabian StyleLimanaqi, Fiona, Francesca Biagioni, Carla Letizia Busceti, Maico Polzella, Cinzia Fabrizi, and Francesco Fornai. 2020. "Potential Antidepressant Effects of Scutellaria baicalensis, Hericium erinaceus and Rhodiola rosea" Antioxidants 9, no. 3: 234. https://doi.org/10.3390/antiox9030234