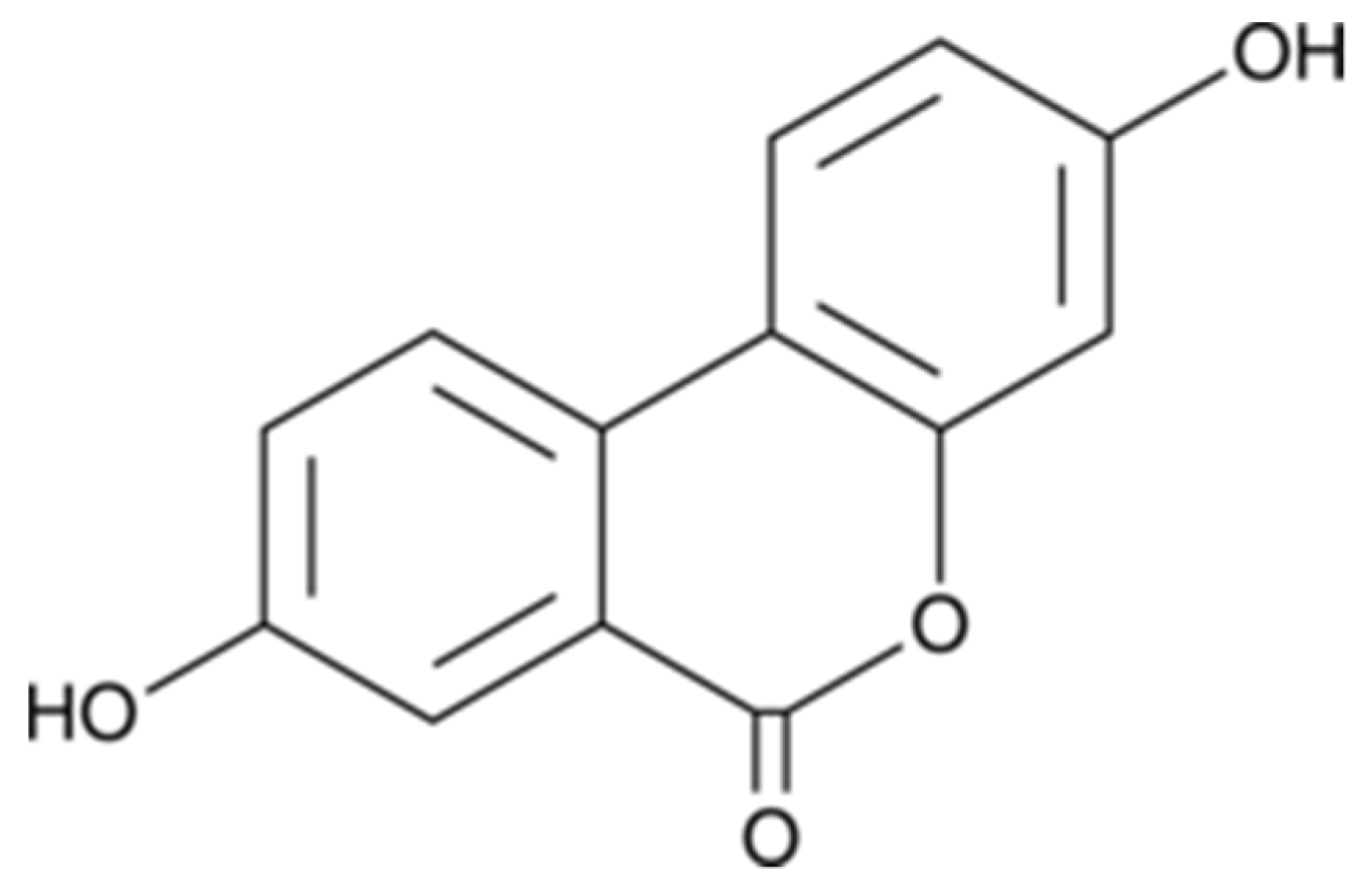
The Metabolite Urolithin-A Ameliorates Oxidative Stress in Neuro-2a Cells, Becoming a Potential Neuroprotective Agent


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Cytoprotective Properties of Urolithin A in Neuro-2a Cells
2.2.1. Neuro-2a Cell Culture and Treatments with Urolithin A and Hydrogen Peroxide
2.2.2. Mitochondrial Activity in Neuro-2a Cells Subjected to Oxidative Stress after Urolithin A Treatment
2.2.3. ROS Production in Neuro-2a Cells Subjected to Oxidative Stress after Exposition to Urolithin A
2.2.4. Lipid Peroxidation in Neuro-2a Cells Subjected to Oxidative Stress after Exposition to Urolithin A (TBARS assay)
2.2.5. Activity of Antioxidant Enzymes in Neuro-2a Cells Subjected to Oxidative Stress after Exposition to Urolithin A
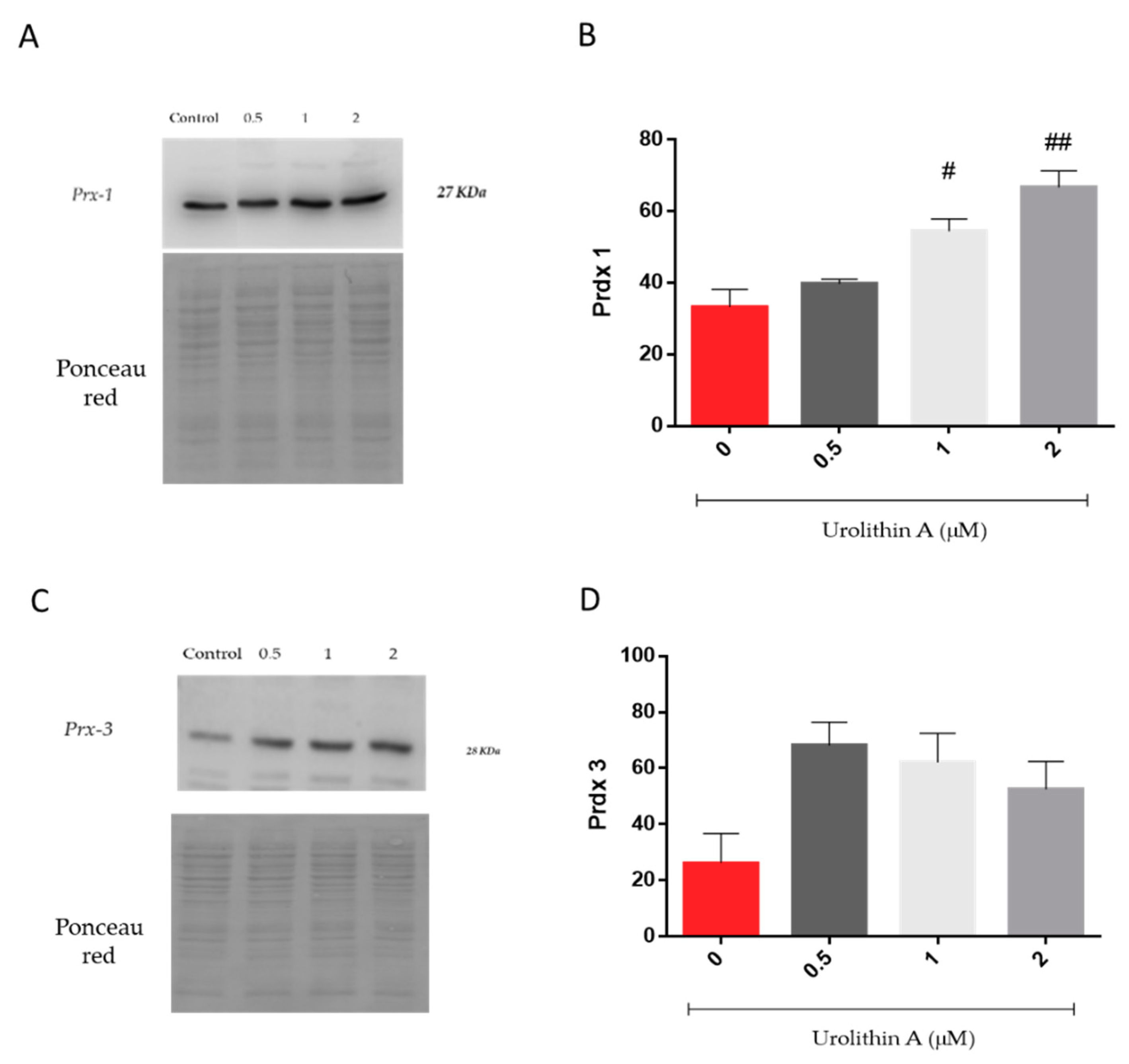
2.2.6. Peroxiredoxin Expression in Neuro-2a Cells by Immunoblotting
2.3. Urolithin A and Its Role as an Inhibitor of CNS Enzymatic Targets
2.3.1. Tyrosinase (TYR) Inhibition
2.3.2. Acetylcholinesterase (AChE) Inhibition
2.3.3. Monoamine Oxidase A (MAO-A) Inhibition
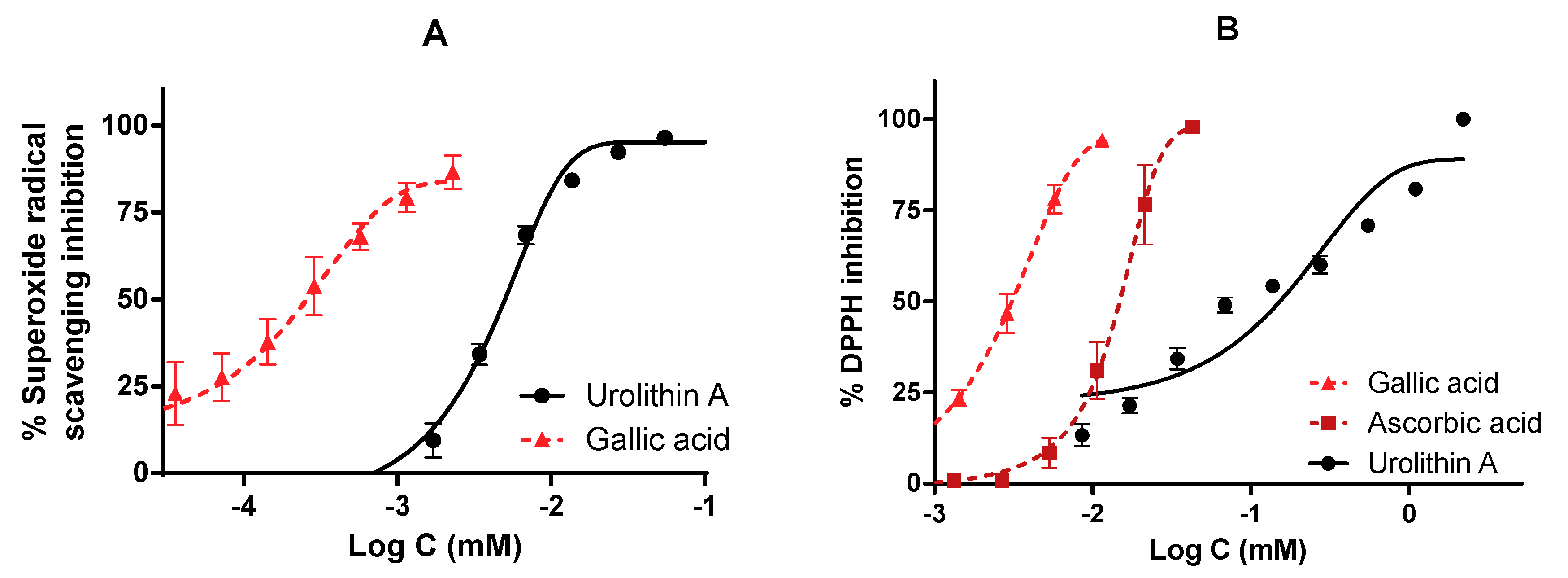
2.4. Urolithin A and Its Role as a Direct Free Radical Scavenger
2.4.1. Oxygen Radical Antioxidant Capacity ORAC Assay
2.4.2. Superoxide Radicals Generated by Xanthine/Xanthine Oxidase (X/XO) System
2.4.3. DPPH Radical Assay
2.5. Statistical Analysis
3. Results
3.1. Cytoprotective Properties of Urolithin A in Neuro-2a Cells
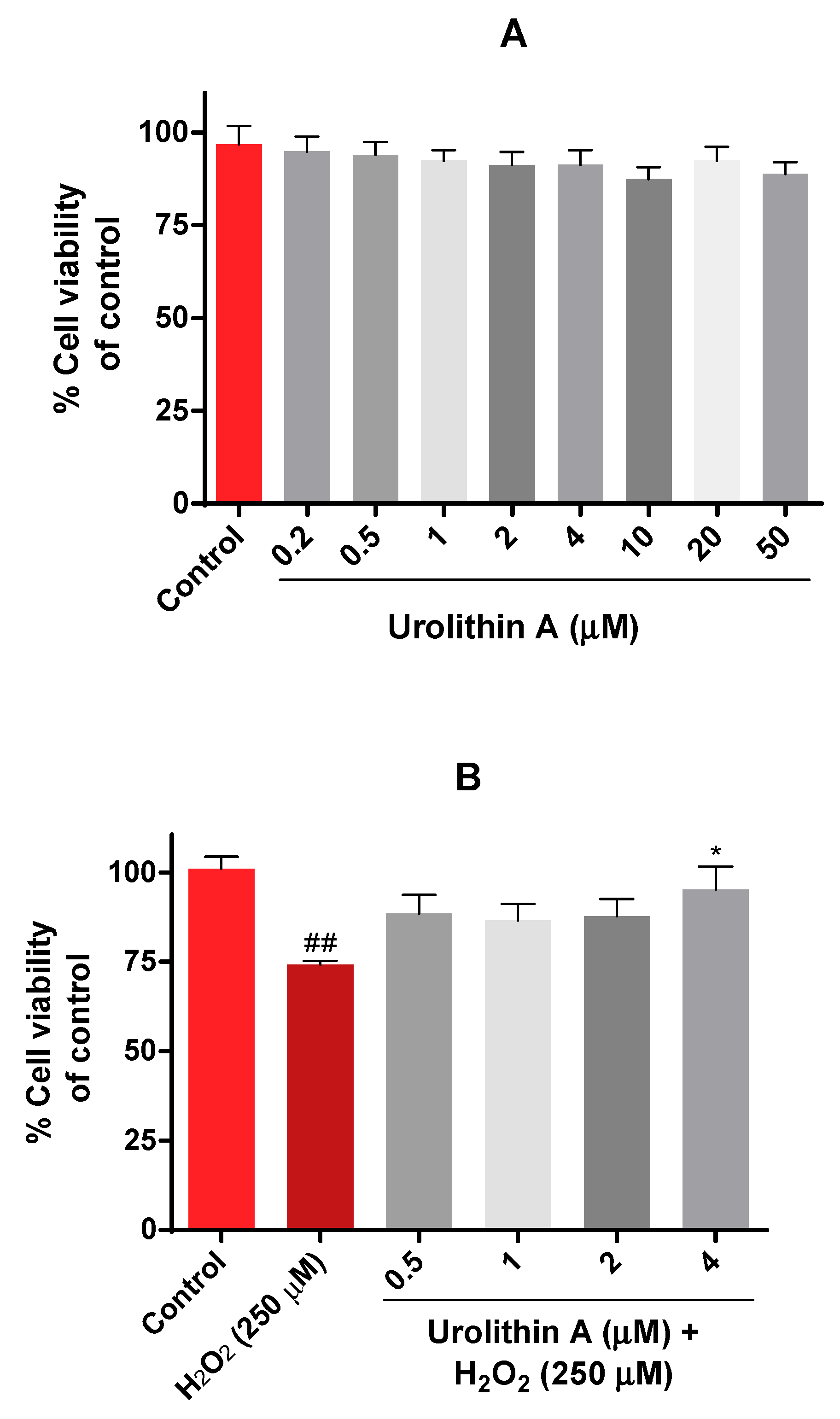
3.1.1. Urolithin A Improves Mitochondrial Activity in Neuro-2a Cells Subjected to Oxidative Stress (MTT Assay)
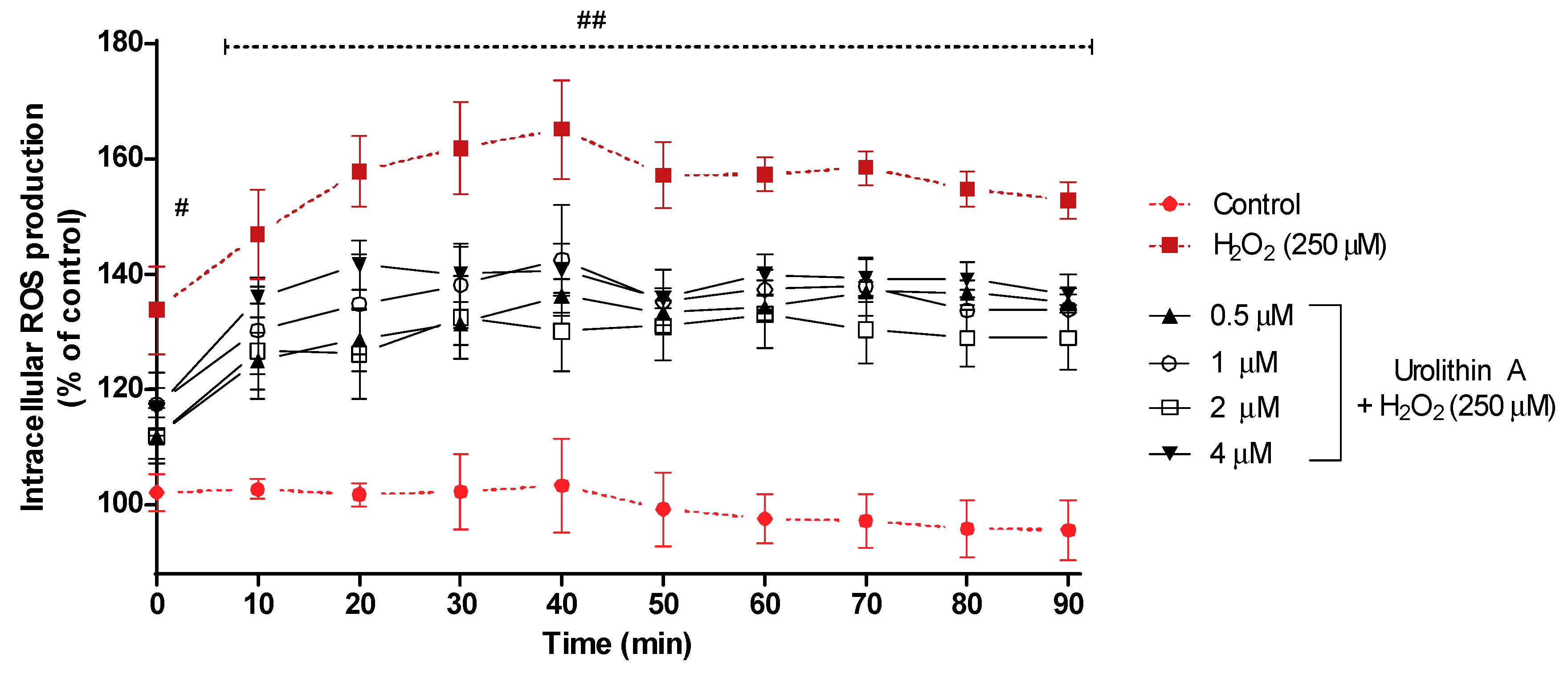
3.1.2. Urolithin A Decreases Intracellular ROS Production in Neuro-2a Cells Subjected to Oxidative Stress (DCFHA-DA Assay)
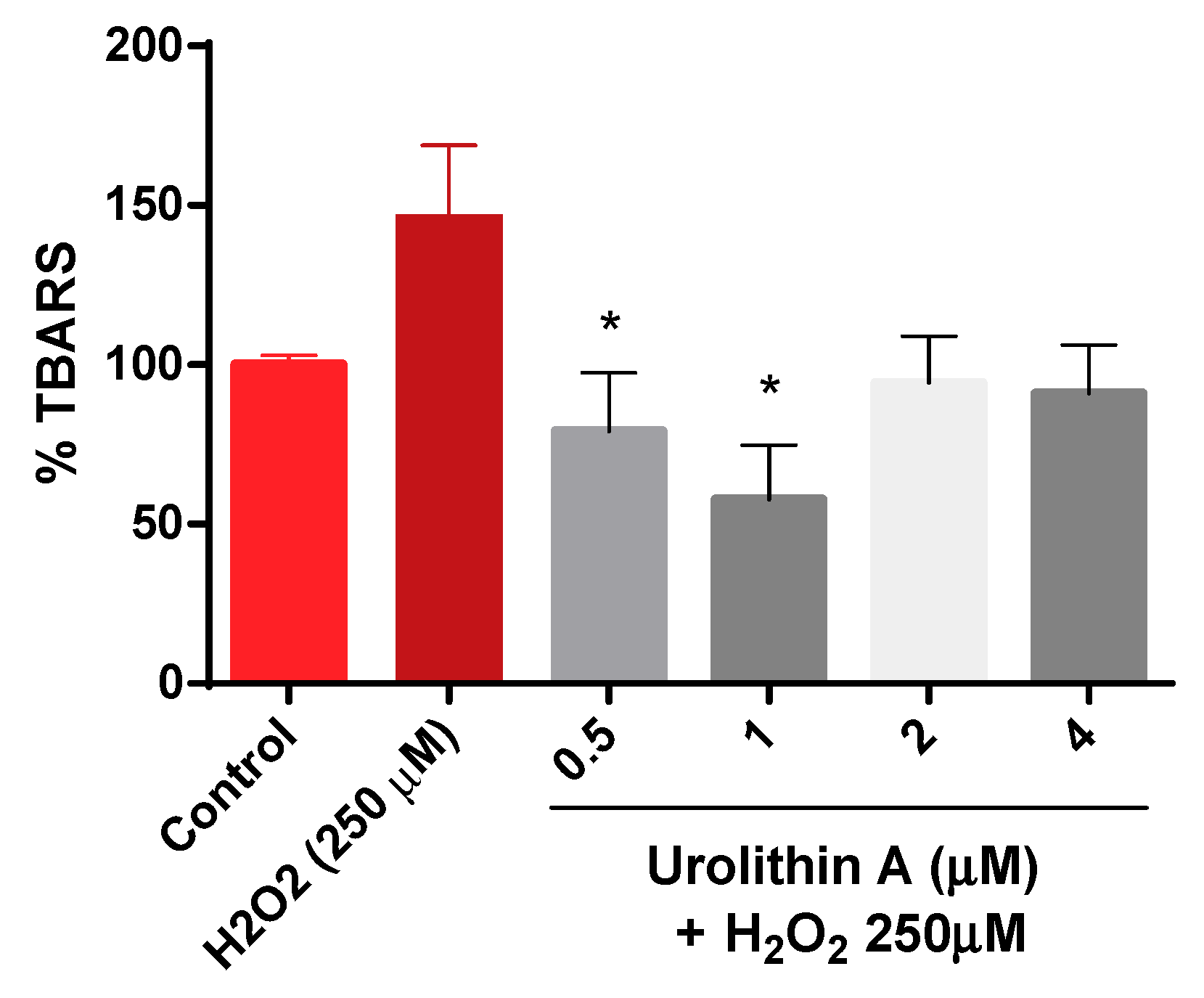
3.1.3. Urolithin A Decreases Lipid Peroxidation in Neuro-2a Cells Subjected to Oxidative Stress (Thiobarbituric Acid Reactive Species, TBARS)
3.1.4. Urolithin A Enhanced the Activity of Antioxidant Enzymes in Neuro-2a Cells Subjected to Oxidative Stress (CAT, SOD, GR, GPx)
3.1.5. Peroxiredoxins Expression
3.2. Urolithin A Inhibits Oxidases (Monoamine Oxidase A and Tyrosinase)
3.3. The Role of Urolithin A as a Direct Free Radical Scavenger
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mämmelä, P.; Savolainen, H.; Lindroos, L.; Kangas, J.; Vartiainen, T. Analysis of oak tannins by liquid chromatography-electrospray ionisation mass spectrometry. J. Chromatogr. A 2000, 891, 75–83. [Google Scholar] [CrossRef]
- Charrier -El Bouhtoury, F.; Charrier, B.; Zahri, S.; Belloncle, C.; Charrier, F.; Pardon, P.; Quideau, S.; Charrier, B. UV light impact on ellagitannins and wood surface color of European oak (Quercus petraea and Quercus robur). Appl. Surf. Sci. 2007, 253, 4985–4989. [Google Scholar]
- Moharram, F.A.; Marzouk, M.S.; El-Toumy, S.A.A.; Ahmed, A.A.E.; Aboutabl, E.A. Polyphenols of Melaleuca quinquenervia leaves – pharmacological studies of grandinin. Phyther. Res. 2003, 17, 767–773. [Google Scholar] [CrossRef]
- González-Sarrías, A.; García-Villalba, R.; Romo-Vaquero, M.; Alasalvar, C.; Örem, A.; Zafrilla, P.; Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. Clustering according to urolithin metabotype explains the interindividual variability in the improvement of cardiovascular risk biomarkers in overweight-obese individuals consuming pomegranate: A randomized clinical trial. Mol. Nutr. Food Res. 2017, 61, 1600830. [Google Scholar] [CrossRef]
- García-Villalba, R.; Vissenaekens, H.; Pitart, J.; Romo-Vaquero, M.; Espín, J.C.; Grootaert, C.; Selma, M.V.; Raes, K.; Smagghe, G.; Possemiers, S.; et al. Gastrointestinal Simulation Model TWIN-SHIME Shows Differences between Human Urolithin-Metabotypes in Gut Microbiota Composition, Pomegranate Polyphenol Metabolism, and Transport along the Intestinal Tract. J. Agric. Food Chem. 2017, 65, 5480–5493. [Google Scholar] [CrossRef] [Green Version]
- Heilman, J.; Andreux, P.; Tran, N.; Rinsch, C.; Blanco-Bose, W. Safety assessment of Urolithin A, a metabolite produced by the human gut microbiota upon dietary intake of plant derived ellagitannins and ellagic acid. Food Chem. Toxicol. 2017, 108, 289–297. [Google Scholar] [CrossRef]
- Les, F.; Arbonés-Mainar, J.M.; Valero, M.S.; López, V. Pomegranate polyphenols and urolithin A inhibit α-glucosidase, dipeptidyl peptidase-4, lipase, triglyceride accumulation and adipogenesis related genes in 3T3-L1 adipocyte-like cells. J. Ethnopharmacol. 2018, 220, 67–74. [Google Scholar] [CrossRef]
- Toney, A.; Chung, S. Urolithin A, a Gut Metabolite, Induces Metabolic Reprogramming of Adipose Tissue by Promoting M2 Macrophage Polarization and Mitochondrial Function (OR12-02-19). Curr. Dev. Nutr. 2019, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Haytowitz, D.; Pehrsson, P. Preliminary Analysis of Isoflavones in Processed Egg Products (P06-127-19). Curr. Dev. Nutr. 2019, 3. [Google Scholar] [CrossRef] [Green Version]
- Mohammed Saleem, Y.I.; Albassam, H.; Selim, M. Urolithin A induces prostate cancer cell death in p53-dependent and in p53-independent manner. Eur. J. Nutr. 2019. [Google Scholar] [CrossRef]
- Liu, F.; Cui, Y.; Yang, F.; Xu, Z.; Da, L.-T.; Zhang, Y. Inhibition of polypeptide N-acetyl-α-galactosaminyltransferases is an underlying mechanism of dietary polyphenols preventing colorectal tumorigenesis. Bioorg. Med. Chem. 2019, 27, 3372–3382. [Google Scholar] [CrossRef]
- Ryu, D.; Mouchiroud, L.; Andreux, P.A.; Katsyuba, E.; Moullan, N.; Nicolet-dit-Félix, A.A.; Williams, E.G.; Jha, P.; Lo Sasso, G.; Huzard, D.; et al. Urolithin A induces mitophagy and prolongs lifespan in C. elegans and increases muscle function in rodents. Nat. Med. 2016, 22, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Andreux, P.; Blanco-Bose, W.; Ryu, D.; Aebischer, P.; Auwerx, J.; Rinsch, C. Orally administered urolithin a is safe and modulates muscle and mitochondrial biomarkers in elderly. Innov. Aging 2017, 1, 1223–1224. [Google Scholar] [CrossRef] [Green Version]
- Cásedas, G.; González-burgos, E.; Smith, C.; López, V.; Pilar, M. Regulation of redox status in neuronal SH-SY5Y cells by blueberry (Vaccinium myrtillus L.) juice, cranberry (Vaccinium macrocarpon A.) juice and cyanidin. Food Chem. Toxicol. 2018, 118, 572–580. [Google Scholar] [CrossRef]
- Mitsuru Uchiyama, M.M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef]
- González-Burgos, E.; Carretero, M.E.; Gómez-Serranillos, M.P. Kaurane diterpenes from Sideritis spp. exert a cytoprotective effect against oxidative injury that is associated with modulation of the Nrf2 system. Phytochemistry 2013, 93, 116–123. [Google Scholar] [CrossRef]
- Cásedas, G.; González-Burgos, E.; Smith, C.; López, V.; Gómez-Serranillos, M.P. Sour cherry (Prunus cerasus L.) juice protects against hydrogen peroxide-induced neurotoxicity by modulating the antioxidant response. J. Funct. Foods 2018, 46, 243–249. [Google Scholar] [CrossRef]
- Staal, G.E.; Helleman, P.W.; De Wael, J.; Veeger, C. Purification and properties of an abnormal glutathione reductase from human erythrocytes. Biochim. Biophys. Acta 1969, 185, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W.G.; Siggins, G.R.; Battenberg, E.F.; Tioffer, B.J.; Bloom, F.E.; et al. Selenium: Biochemical Role as a Component of Glutathione Peroxidase. Source Sci. New Ser. J. Biol. Chem. 1973, 179, 588–590. [Google Scholar] [CrossRef]
- Wilson, S.R.; Zucker, P.A.; Huang, C.R.; Spector, A. Development of Synthetic Compounds with Glutathione Peroxidase Activity. J. Am. Chem. Soc. 1989, 111, 5936–5939. [Google Scholar] [CrossRef]
- Sezer Senol, F.; Orhan, I.E.; Ozgen, U.; Renda, G.; Bulut, G.; Guven, L.; Karaoglan, E.S.; Sevindik, H.G.; Skalicka-Wozniak, K.; Koca Caliskan, U.; et al. Memory-vitalizing effect of twenty-five medicinal and edible plants and their isolated compounds. S. Afr. J. Bot. 2015, 102, 102–109. [Google Scholar] [CrossRef]
- Les, F.; Prieto, J.M.; Arbonés-Mainar, J.M.; Valero, M.S.; López, V. Bioactive properties of commercialised pomegranate (Punica granatum) juice: Antioxidant, antiproliferative and enzyme inhibiting activities. Food Funct. 2015, 6, 2049–2057. [Google Scholar] [CrossRef] [PubMed]
- Olsen, H.T.; Stafford, G.I.; van Staden, J.; Christensen, S.B.; Jäger, A.K. Isolation of the MAO-inhibitor naringenin from Mentha aquatica L. J. Ethnopharmacol. 2008, 117, 500–502. [Google Scholar] [CrossRef] [PubMed]
- Dávalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Extending Applicability of the Oxygen Radical Absorbance Capacity (ORAC-Fluorescein) Assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef]
- Rodríguez-Chávez, J.L.; Coballase-Urrutia, E.; Nieto-Camacho, A.; Delgado-Lamas, G. Antioxidant capacity of “mexican arnica” Heterotheca inuloides cass natural products and some derivatives: Their anti-inflammatory evaluation and effect on C. elegans life span. Oxidative Med. Cell. Longev. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [Green Version]
- López, V.; Akerreta, S.; Casanova, E.; García-Mina, J.M.; Cavero, R.Y.; Calvo, M.I. In vitro antioxidant and anti-rhizopus activities of lamiaceae herbal extracts. Plant Foods Hum. Nutr. 2007, 62, 151–155. [Google Scholar] [CrossRef]
- Manta, B.; Hugo, M.; Ortiz, C.; Ferrer-Sueta, G.; Trujillo, M.; Denicola, A. The peroxidase and peroxynitrite reductase activity of human erythrocyte peroxiredoxin 2. Arch. Biochem. Biophys. 2009, 484, 146–154. [Google Scholar] [CrossRef]
- Randall, L.M.; Ferrer-Sueta, G.; Denicola, A. Peroxiredoxins as preferential targets in H2O2-induced signaling. Methods Enzymol. 2013, 527, 41–63. [Google Scholar]
- Ahsan, A.; Zheng, Y.; Wu, X.; Tang, W.; Liu, M.; Ma, S.; Jiang, L.; Hu, W.; Zhang, X.; Chen, Z. Urolithin A-activated autophagy but not mitophagy protects against ischemic neuronal injury by inhibiting ER stress in vitro and in vivo. CNS Neurosci. Ther. 2019, 25, 976–986. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Qiu, Z.; Zhou, B.; Liu, C.; Ruan, J.; Yan, Q.; Liao, J.; Zhu, F. In vitro antiproliferative and antioxidant effects of urolithin A, the colonic metabolite of ellagic acid, on hepatocellular carcinomas HepG2 cells. Toxicol. In Vitro 2015, 29, 1107–1115. [Google Scholar] [CrossRef]
- Dobroslawa, B.; Kasimsetty, S.G.; Khan, S.I.; Daneel, F. Urolithins, intestinal microbial metabolites of pomegranate ellagitannins, exhibit potent antioxidant activity in a cell-based assay. J. Agric. Food Chem. 2009, 57, 10181–10186. [Google Scholar]
- Qiu, Z.; Zhou, B.; Jin, L.; Yu, H.; Liu, L.; Liu, Y.; Qin, C.; Xie, S.; Zhu, F. In vitro antioxidant and antiproliferative effects of ellagic acid and its colonic metabolite, urolithins, on human bladder cancer T24 cells. Food Chem. Toxicol. 2013, 59, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Jing, T.; Liao, J.; Shen, K.; Chen, X.; Xu, Z.; Tian, W.; Wang, Y.; Jin, B.; Pan, H. Protective effect of urolithin a on cisplatin-induced nephrotoxicity in mice via modulation of inflammation and oxidative stress. Food Chem. Toxicol. 2019, 129, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, M.K.; Choudhury, S.; Borah, A. An in silico investigation on the inhibitory potential of the constituents of Pomegranate juice on antioxidant defense mechanism: Relevance to neurodegenerative diseases. IBRO Rep. 2019, 6, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Khodaie, N.; Tajuddin, N.; Mitchell, R.M.; Neafsey, E.J.; Collins, M.A. Combinatorial Preconditioning of Rat Brain Cultures with Subprotective Ethanol and Resveratrol Concentrations Promotes Synergistic Neuroprotection. Neurotox. Res. 2018, 34, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Kallio, T.; Kallio, J.; Jaakkola, M.; Mäki, M.; Kilpeläinen, P.; Virtanen, V. Urolithins display both antioxidant and pro-oxidant activities depending on assay system and conditions. J. Agric. Food Chem. 2013, 61, 10720–10729. [Google Scholar] [CrossRef]
- Ishimoto, H.; Tai, A.; Yoshimura, M.; Amakura, Y.; Yoshida, T.; Hatano, T.; Ito, H. Antioxidative Properties of Functional Polyphenols and Their Metabolites Assessed by an ORAC Assay. Biosci. Biotechnol. Biochem. 2012, 76, 395–399. [Google Scholar] [CrossRef] [Green Version]
- Tomás-Barberán, F.A.; González-Sarrías, A.; García-Villalba, R.; Núñez-Sánchez, M.A.; Selma, M.V.; García-Conesa, M.T.; Espín, J.C. Urolithins, the rescue of “old” metabolites to understand a “new” concept: Metabotypes as a nexus among phenolic metabolism, microbiota dysbiosis, and host health status. Mol. Nutr. Food Res. 2017, 61, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Nealmongkol, P.; Tangdenpaisal, K.; Sitthimonchai, S.; Ruchirawat, S.; Thasana, N. Cu(I)-mediated lactone formation in subcritical water: A benign synthesis of benzopyranones and urolithins A–C. Tetrahedron 2013, 69, 9277–9283. [Google Scholar] [CrossRef]
- Hideyuki Ito, A. Metabolites of the Ellagitannin Geraniin and Their Antioxidant Activities Reviews. Planta Med. 2011, 77, 1110–1115. [Google Scholar]
- Cerda, B.; Espin, J.C.; Parra, S.; Martinez, P.; Tomas-Barberan, F.A. The potent in vitro antioxidant ellagitannins from pomegranate juice are metabolised into bioavailable but poor antioxidant hydroxy-6H-dibenzopyran-6-one derivatives by the colonic microflora of healthy humans. Eur. J. Nutr. 2004, 43, 205–220. [Google Scholar] [CrossRef]
- Bookheimer, S.Y.; Renner, B.A.; Ekstrom, A.; Li, Z.; Henning, S.M.; Brown, J.A.; Jones, M.; Moody, T.; Small, G.W. Pomegranate Juice Augments Memory and fMRI Activity in Middle-Aged and Older Adults with Mild Memory Complaints. Evid. Complement. Altern. Med. 2013, 2013, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gulcan, H.O.; Unlu, S.; Esiringu, I.; Ercetin, T.; Sahin, Y.; Oz, D.; Sahin, M.F. Design, synthesis and biological evaluation of novel 6H-benzo[c]chromen-6-one, and 7,8,9,10-tetrahydro-benzo[c]chromen-6-one derivatives as potential cholinesterase inhibitors. Bioorg. Med. Chem. 2014, 22, 5141–5154. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.; Ma, H.; Liu, W.; Niesen, D.B.; Shah, N.; Crews, R.; Rose, K.N.; Vattem, D.A.; Seeram, N.P. Pomegranate’s Neuroprotective Effects against Alzheimer’s Disease Are Mediated by Urolithins, Its Ellagitannin-Gut Microbial Derived Metabolites. ACS Chem. Neurosci. 2016, 7, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-T.; Chang, W.-C.; Hsu, C.; Su, N.-W. Antimelanogenic Effect of Urolithin A and Urolithin B, the Colonic Metabolites of Ellagic Acid, in B16 Melanoma Cells. J. Agric. Food Chem. 2017, 65, 6870–6876. [Google Scholar] [CrossRef] [PubMed]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cásedas, G.; Les, F.; Choya-Foces, C.; Hugo, M.; López, V. The Metabolite Urolithin-A Ameliorates Oxidative Stress in Neuro-2a Cells, Becoming a Potential Neuroprotective Agent. Antioxidants 2020, 9, 177. https://doi.org/10.3390/antiox9020177
Cásedas G, Les F, Choya-Foces C, Hugo M, López V. The Metabolite Urolithin-A Ameliorates Oxidative Stress in Neuro-2a Cells, Becoming a Potential Neuroprotective Agent. Antioxidants. 2020; 9(2):177. https://doi.org/10.3390/antiox9020177
Chicago/Turabian StyleCásedas, Guillermo, Francisco Les, Carmen Choya-Foces, Martín Hugo, and Víctor López. 2020. "The Metabolite Urolithin-A Ameliorates Oxidative Stress in Neuro-2a Cells, Becoming a Potential Neuroprotective Agent" Antioxidants 9, no. 2: 177. https://doi.org/10.3390/antiox9020177