
The Role of Adipokines and Myokines in the Pathogenesis of Different Obesity Phenotypes—New Perspectives
 , ,
, ,
Abstract
:1. Introduction
2. Phenotypes of Obesity
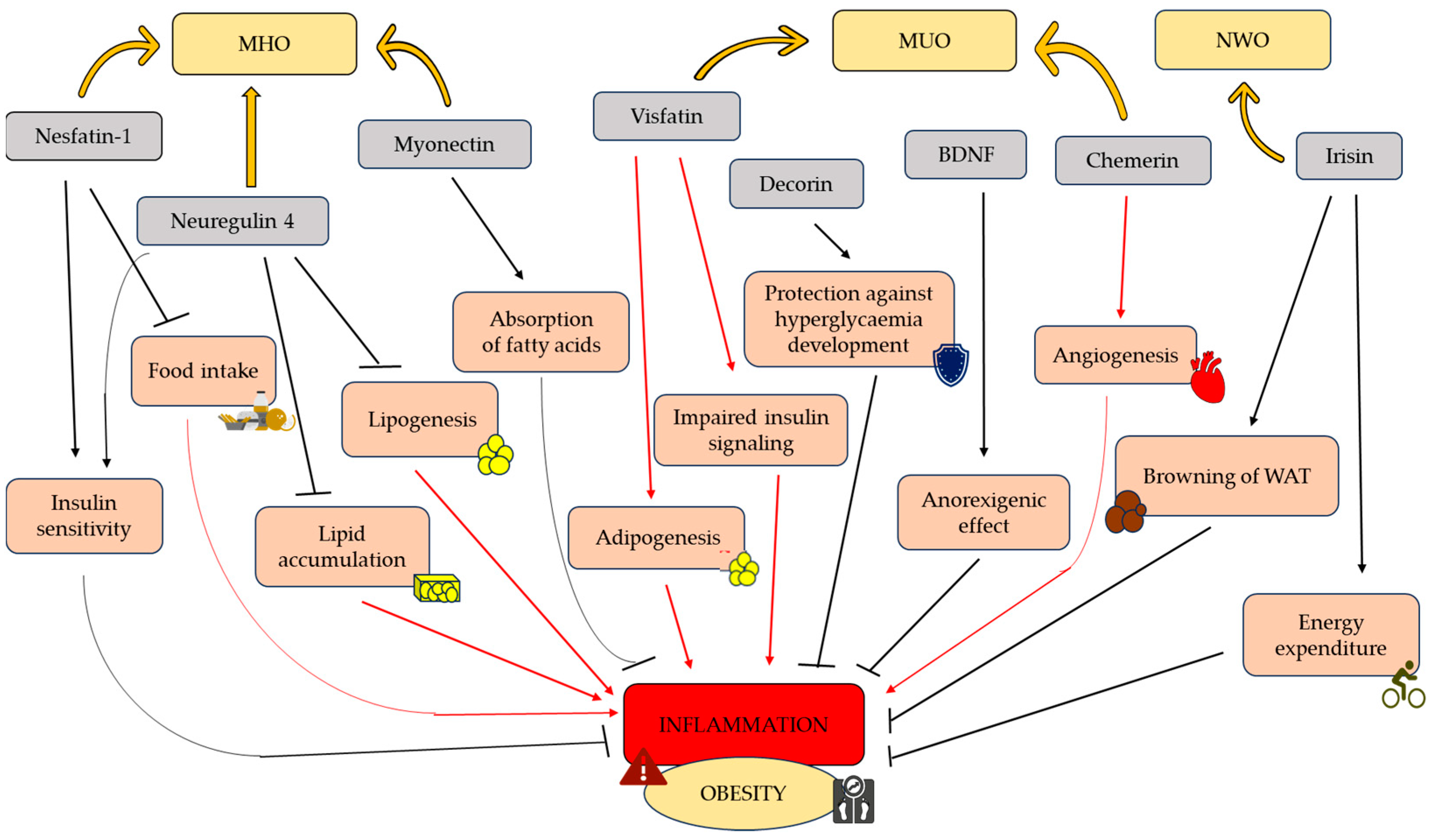
3. Adipokines and Myokines Protecting from Obesity
3.1. Nestatin-1

{kind=link}
| Adipokines/Myokines | The Potential Role in Obesity and Its Phenotypes | References |
|---|---|---|
| Nesfatin-1 | Regulator of energy homeostasis, food intake, and glucose metabolism; potential anorexigenic factor; | [38] |
| Obese people (especially MUO-like phenotype) demonstrate lower nesfatin-1 concentrations than non-obese individuals; | [51,59] | |
| Nesfatin-1 levels negatively correlate with BMI, body AT percentage, body AT weight, and blood glucose levels. | [51] | |
| Neuregulin 4 | Inhibits lipogenesis and lipid accumulation, reduces chronic inflammation; | [61] |
| Neuregulin 4 is decreased in obese individuals with metabolic syndrome (MUO-like phenotype) and negatively correlated with waist circumference, body AT percentage, BMI, LDL cholesterol, and fasting glucose concentration. | [62] | |
| Myonectin | Influences lipid homeostasis in the liver and AT, regulates energy metabolism; | [63] |
| Myonectin can be correlated to MHO-like phenotype; exercises increase levels of myonectin in obese subjects and decrease IR. | [64] | |
| Irisin | Supports WAT browning, increases energy expenditure, regulates glucose metabolic homeostasis; | [65] |
| Reduces inflammatory processes; | [66,67] | |
| Conflicting results occur in irisin concentration in obesity phenotypes. The decrease levels of irisine in MUO obese has been showed. | [68,69] | |
| While, another study indicated its increase concentration. | [70] | |
| Decorin | Is involved in inflammatory processes and maintaining glucose tolerance; | [71] |
| Decorin concentration is increased in obesity. | [72] | |
| BDNF | Possess anorexigenic effects; regulates energy homeostasis; | [73,74] |
| Conflicting results occurs BDNF concentration in obesity. Data indicated its decrease concentration. | [75] | |
| On the other hand, another study indicated its increase levels. | [76] | |
| Visfatin | Enhances adipogenesis, promotes pro-inflammatory processes and IR, contribute to cholesterol accumulation; | [77,78] |
| Obese individuals (MUO-like phenotype) present higher visfatin levels; | [79] | |
| Visfatin levels positively correlate with IR indicators such as glucose and insulin concentration and HOMA-IR index value. | [80] | |
| Chemerin | Promotes AT growth by inducing angiogenesis and increasing its vascularization, increases inflammation in AT; | [81] |
| Chemerin levels are lower in MHO compared to MUO; | [82] | |
| In patients with obesity, chemerin levels positively correlates with obesity markers (HOMA-IR, BMI, AT percentage, waist circumference, WHR, triglycerides, total cholesterol). | [83] |
3.2. Neuregulin 4
3.3. Myonectin
3.4. Irisin
3.5. Decorin
3.6. BDNF
4. Adipokines and Myokines Predisposing to Obesity
4.1. Visfatin
4.2. Chemerin
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- 2017 Global Nutrition Report. Available online: https://globalnutritionreport.org/reports/2017-global-nutrition-report/ (accessed on 20 April 2023).
- World Health Organization. Obesity: Preventing and Managing the Global Epidemic: Report of a WHO Consultation; World Health Organization: Geneva, Switzerland, 2000. [Google Scholar]
- Agha, M.; Agha, R. The Rising Prevalence of Obesity: Part A: Impact on Public Health. Int. J. Surg. Oncol. 2017, 2, e17. [Google Scholar] [CrossRef]
- Zorena, K.; Jachimowicz-Duda, O.; Ślęzak, D.; Robakowska, M.; Mrugacz, M. Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications. Int. J. Mol. Sci. 2020, 21, E3570. [Google Scholar] [CrossRef]
- Febbraio, M.A.; Pedersen, B.K. Contraction-Induced Myokine Production and Release: Is Skeletal Muscle an Endocrine Organ? Exerc. Sport. Sci. Rev. 2005, 33, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Itoh, N. FGF21 as a Hepatokine, Adipokine, and Myokine in Metabolism and Diseases. Front. Endocrinol. 2014, 5, 107. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.-J.; Lee, D.S.; Kim, W.K.; Han, B.S.; Lee, S.C.; Bae, K.-H. Metabolic Adaptation in Obesity and Type II Diabetes: Myokines, Adipokines and Hepatokines. Int. J. Mol. Sci. 2016, 18, 8. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Zhao, H.; Yin, C.; Lan, X.; Wu, L.; Du, X.; Griffiths, H.R.; Gao, D. Adipokines, Hepatokines and Myokines: Focus on Their Role and Molecular Mechanisms in Adipose Tissue Inflammation. Front. Endocrinol. 2022, 13, 873699. [Google Scholar] [CrossRef]
- Cinti, S. Adipose Organ Development and Remodeling. Compr. Physiol. 2018, 8, 1357–1431. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Dos Santos, A.R.; de Oliveira Zanuso, B.; Miola, V.F.B.; Barbalho, S.M.; Santos Bueno, P.C.; Flato, U.A.P.; Detregiachi, C.R.P.; Buchaim, D.V.; Buchaim, R.L.; Tofano, R.J.; et al. Adipokines, Myokines, and Hepatokines: Crosstalk and Metabolic Repercussions. Int. J. Mol. Sci. 2021, 22, 2639. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gil, A.M.; Elizondo-Montemayor, L. The Role of Exercise in the Interplay between Myokines, Hepatokines, Osteokines, Adipokines, and Modulation of Inflammation for Energy Substrate Redistribution and Fat Mass Loss: A Review. Nutrients 2020, 12, 1899. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in Inflammation and Metabolic Disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef]
- Ruderman, N.; Chisholm, D.; Pi-Sunyer, X.; Schneider, S. The Metabolically Obese, Normal-Weight Individual Revisited. Diabetes 1998, 47, 699–713. [Google Scholar] [CrossRef]
- Sims, E.A. Are There Persons Who Are Obese, but Metabolically Healthy? Metabolism 2001, 50, 1499–1504. [Google Scholar] [CrossRef] [PubMed]
- Graf, C.; Ferrari, N. Metabolic Health—The Role of Adipo-Myokines. Int. J. Mol. Sci. 2019, 20, 6159. [Google Scholar] [CrossRef] [PubMed]
- Lejawa, M.; Osadnik, K.; Osadnik, T.; Pawlas, N. Association of Metabolically Healthy and Unhealthy Obesity Phenotypes with Oxidative Stress Parameters and Telomere Length in Healthy Young Adult Men. Analysis of the MAGNETIC Study. Antioxidants 2021, 10, 93. [Google Scholar] [CrossRef]
- Severinsen, M.C.K.; Pedersen, B.K. Muscle–Organ Crosstalk: The Emerging Roles of Myokines. Endocr. Rev. 2020, 41, 594–609. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Febbraio, M.A. Muscles, Exercise and Obesity: Skeletal Muscle as a Secretory Organ. Nat. Rev. Endocrinol. 2012, 8, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Khansari, N.; Shakiba, Y.; Mahmoudi, M. Chronic Inflammation and Oxidative Stress as a Major Cause of Age- Related Diseases and Cancer. Recent Pat. Inflamm. Allergy Drug Discov. 2009, 3, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Barbalho, S.M.; Prado Neto, E.V.; De Alvares Goulart, R.; Bechara, M.D.; Baisi Chagas, E.F.; Audi, M.; Guissoni Campos, L.M.; Landgraf Guiger, E.; Buchaim, R.L.; Buchaim, D.V.; et al. Myokines: A Descriptive Review. J. Sports Med. Phys. Fitness 2020, 60, 1583–1590. [Google Scholar] [CrossRef]
- Chung, H.S.; Choi, K.M. Adipokines and Myokines: A Pivotal Role in Metabolic and Cardiovascular Disorders. Curr. Med. Chem. 2018, 25, 2401–2415. [Google Scholar] [CrossRef]
- Landecho, M.F.; Tuero, C.; Valentí, V.; Bilbao, I.; de la Higuera, M.; Frühbeck, G. Relevance of Leptin and Other Adipokines in Obesity-Associated Cardiovascular Risk. Nutrients 2019, 11, 2664. [Google Scholar] [CrossRef]
- Farhadipour, M.; Depoortere, I. The Function of Gastrointestinal Hormones in Obesity-Implications for the Regulation of Energy Intake. Nutrients 2021, 13, 1839. [Google Scholar] [CrossRef]
- Pluta, W.; Dudzińska, W.; Lubkowska, A. Metabolic Obesity in People with Normal Body Weight (MONW)-Review of Diagnostic Criteria. Int. J. Environ. Res. Public. Health 2022, 19, 624. [Google Scholar] [CrossRef]
- Goossens, G.H. The Metabolic Phenotype in Obesity: Fat Mass, Body Fat Distribution, and Adipose Tissue Function. Obes. Facts 2017, 10, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Choi, M.-S. Obesity and Its Metabolic Complications: The Role of Adipokines and the Relationship between Obesity, Inflammation, Insulin Resistance, Dyslipidemia and Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.-L.; Ren, J.-X.; Zhang, P.; Jin, H.; Qu, Y.; Yu, Y.; Guo, Z.-N.; Yang, Y. Strong Association of Waist Circumference (WC), Body Mass Index (BMI), Waist-to-Height Ratio (WHtR), and Waist-to-Hip Ratio (WHR) with Diabetes: A Population-Based Cross-Sectional Study in Jilin Province, China. J. Diabetes Res. 2021, 2021, 8812431. [Google Scholar] [CrossRef] [PubMed]
- Mayoral, L.P.-C.; Andrade, G.M.; Mayoral, E.P.-C.; Huerta, T.H.; Canseco, S.P.; Rodal Canales, F.J.; Cabrera-Fuentes, H.A.; Cruz, M.M.; Pérez Santiago, A.D.; Alpuche, J.J.; et al. Obesity Subtypes, Related Biomarkers & Heterogeneity. Indian. J. Med. Res. 2020, 151, 11–21. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzo, A.; Soldati, L.; Sarlo, F.; Calvani, M.; Di Lorenzo, N.; Di Renzo, L. New Obesity Classification Criteria as a Tool for Bariatric Surgery Indication. World J. Gastroenterol. 2016, 22, 681–703. [Google Scholar] [CrossRef]
- Oliveros, E.; Somers, V.K.; Sochor, O.; Goel, K.; Lopez-Jimenez, F. The Concept of Normal Weight Obesity. Prog. Cardiovasc. Dis. 2014, 56, 426–433. [Google Scholar] [CrossRef]
- Blüher, M. Metabolically Healthy Obesity. Endocr. Rev. 2020, 41, bnaa004. [Google Scholar] [CrossRef]
- Després, J.-P. Body Fat Distribution and Risk of Cardiovascular Disease: An Update. Circulation 2012, 126, 1301–1313. [Google Scholar] [CrossRef] [PubMed]
- Di Daniele, N.; Petramala, L.; Di Renzo, L.; Sarlo, F.; Della Rocca, D.G.; Rizzo, M.; Fondacaro, V.; Iacopino, L.; Pepine, C.J.; De Lorenzo, A. Body Composition Changes and Cardiometabolic Benefits of a Balanced Italian Mediterranean Diet in Obese Patients with Metabolic Syndrome. Acta Diabetol. 2013, 50, 409–416. [Google Scholar] [CrossRef]
- The Metabolic Syndrome—A New Worldwide Definition—Abstract—Europe PMC. Available online: http://europepmc.org/article/MED/16182882 (accessed on 20 April 2023).
- Wang, B.; Zhang, M.; Wang, S.; Wang, C.; Wang, J.; Li, L.; Zhang, L.; Ren, Y.; Han, C.; Zhao, Y.; et al. Dynamic Status of Metabolically Healthy Overweight/Obesity and Metabolically Unhealthy and Normal Weight and the Risk of Type 2 Diabetes Mellitus: A Cohort Study of a Rural Adult Chinese Population. Obes. Res. Clin. Pract. 2018, 12, 61–71. [Google Scholar] [CrossRef]
- Schröder, H.; Ramos, R.; Baena-Díez, J.M.; Mendez, M.A.; Canal, D.J.; Fíto, M.; Sala, J.; Elosua, R. Determinants of the Transition from a Cardiometabolic Normal to Abnormal Overweight/Obese Phenotype in a Spanish Population. Eur. J. Nutr. 2014, 53, 1345–1353. [Google Scholar] [CrossRef]
- Kramer, C.K.; Zinman, B.; Retnakaran, R. Are Metabolically Healthy Overweight and Obesity Benign Conditions?: A Systematic Review and Meta-Analysis. Ann. Intern. Med. 2013, 159, 758. [Google Scholar] [CrossRef] [PubMed]
- Dore, R.; Levata, L.; Lehnert, H.; Schulz, C. Nesfatin-1: Functions and Physiology of a Novel Regulatory Peptide. J. Endocrinol. 2017, 232, R45–R65. [Google Scholar] [CrossRef]
- Prinz, P.; Goebel-Stengel, M.; Teuffel, P.; Rose, M.; Klapp, B.F.; Stengel, A. Peripheral and Central Localization of the Nesfatin-1 Receptor Using Autoradiography in Rats. Biochem. Biophys. Res. Commun. 2016, 470, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Dong, J.; Jiang, Z.-Y. Nesfatin-1 Influences the Excitability of Glucosensing Neurons in the Hypothalamic Nuclei and Inhibits the Food Intake. Regul. Pept. 2012, 177, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Oh-I, S.; Shimizu, H.; Satoh, T.; Okada, S.; Adachi, S.; Inoue, K.; Eguchi, H.; Yamamoto, M.; Imaki, T.; Hashimoto, K.; et al. Identification of Nesfatin-1 as a Satiety Molecule in the Hypothalamus. Nature 2006, 443, 709–712. [Google Scholar] [CrossRef]
- Foo, K.S.; Brismar, H.; Broberger, C. Distribution and Neuropeptide Coexistence of Nucleobindin-2 mRNA/Nesfatin-like Immunoreactivity in the Rat CNS. Neuroscience 2008, 156, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Pałasz, A.; Janas-Kozik, M.; Borrow, A.; Arias-Carrión, O.; Worthington, J.J. The Potential Role of the Novel Hypothalamic Neuropeptides Nesfatin-1, Phoenixin, Spexin and Kisspeptin in the Pathogenesis of Anxiety and Anorexia Nervosa. Neurochem. Int. 2018, 113, 120–136. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, L.; Yang, M.; Liu, H.; Boden, G.; Yang, G. Increased Plasma Levels of Nesfatin-1 in Patients with Newly Diagnosed Type 2 Diabetes Mellitus. Exp. Clin. Endocrinol. Diabetes 2012, 120, 91–95. [Google Scholar] [CrossRef]
- Shimizu, H.; Mori, M. The Brain-Adipose Axis: A Review of Involvement of Molecules. Nutr. Neurosci. 2005, 8, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Stengel, A.; Goebel, M.; Wang, L.; Rivier, J.; Kobelt, P.; Mönnikes, H.; Lambrecht, N.W.G.; Taché, Y. Central Nesfatin-1 Reduces Dark-Phase Food Intake and Gastric Emptying in Rats: Differential Role of Corticotropin-Releasing Factor2 Receptor. Endocrinology 2009, 150, 4911–4919. [Google Scholar] [CrossRef] [PubMed]
- Maejima, Y.; Sedbazar, U.; Suyama, S.; Kohno, D.; Onaka, T.; Takano, E.; Yoshida, N.; Koike, M.; Uchiyama, Y.; Fujiwara, K.; et al. Nesfatin-1-Regulated Oxytocinergic Signaling in the Paraventricular Nucleus Causes Anorexia through a Leptin-Independent Melanocortin Pathway. Cell Metab. 2009, 10, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.K.; Hallschmid, M.; Kern, W.; Lehnert, H.; Randeva, H.S. Decreased Cerebrospinal Fluid/Plasma Ratio of the Novel Satiety Molecule, Nesfatin-1/NUCB-2, in Obese Humans: Evidence of Nesfatin-1/NUCB-2 Resistance and Implications for Obesity Treatment. J. Clin. Endocrinol. Metab. 2011, 96, E669–E673. [Google Scholar] [CrossRef] [PubMed]
- Saldanha, J.F.; Carrero, J.J.; Lobo, J.C.; Stockler-Pinto, M.B.; Leal, V.O.; Calixto, A.; Geloneze, B.; Mafra, D. The Newly Identified Anorexigenic Adipokine Nesfatin-1 in Hemodialysis Patients: Are There Associations with Food Intake, Body Composition and Inflammation? Regul. Pept. 2012, 173, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Anwar, G.M.; Yamamah, G.; Ibrahim, A.; El-Lebedy, D.; Farid, T.M.; Mahmoud, R. Nesfatin-1 in Childhood and Adolescent Obesity and Its Association with Food Intake, Body Composition and Insulin Resistance. Regul. Pept. 2014, 188, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, T.; Shimizu, H.; Yamada, M.; Osaki, A.; Oh-I, S.; Ariyama, Y.; Takahashi, H.; Okada, S.; Hashimoto, K.; Satoh, T.; et al. Fasting Concentrations of Nesfatin-1 Are Negatively Correlated with Body Mass Index in Non-Obese Males. Clin. Endocrinol. 2010, 73, 484–490. [Google Scholar] [CrossRef]
- Abaci, A.; Catli, G.; Anik, A.; Kume, T.; Bober, E. The Relation of Serum Nesfatin-1 Level with Metabolic and Clinical Parameters in Obese and Healthy Children. Pediatr. Diabetes 2013, 14, 189–195. [Google Scholar] [CrossRef]
- Hofmann, T.; Stengel, A.; Ahnis, A.; Buße, P.; Elbelt, U.; Klapp, B.F. NUCB2/Nesfatin-1 Is Associated with Elevated Scores of Anxiety in Female Obese Patients. Psychoneuroendocrinology 2013, 38, 2502–2510. [Google Scholar] [CrossRef]
- Gajewska, J.; Ambroszkiewicz, J.; Szamotulska, K.; Rowicka, G.; Strucińska, M.; Klemarczyk, W.; Chełchowska, M. Associations between Oxidant/Antioxidant Status and Circulating Adipokines in Non-Obese Children with Prader–Willi Syndrome. Antioxidants 2023, 12, 927. [Google Scholar] [CrossRef] [PubMed]
- Dokumacioglu, E.; Iskender, H.; Sahin, A.; Erturk, E.Y.; Kaynar, O. Serum Levels of Nesfatin-1 and Irisin in Obese Children. Eur. Cytokine Netw. 2020, 31, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Does NUCB2/Nesfatin-1 Influence Eating Behaviors in Obese Patients with Binge Eating Disorder? Toward a Neurobiological Pathway—PMC. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9864089/#B19-nutrients-15-00348 (accessed on 18 April 2023).
- Wang, Y.; Li, Z.; Zhang, X.; Xiang, X.; Li, Y.; Mulholland, M.W.; Zhang, W. Nesfatin-1 Promotes Brown Adipocyte Phenotype. Sci. Rep. 2016, 6, 34747. [Google Scholar] [CrossRef]
- Tekin, T.; Cicek, B.; Konyaligil, N. Regulatory Peptide Nesfatin-1 and Its Relationship with Metabolic Syndrome. Eurasian J. Med. 2019, 51, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Başar, O.; Akbal, E.; Köklü, S.; Koçak, E.; Tuna, Y.; Ekiz, F.; Gültuna, S.; Yιlmaz, F.M.; Aydoğan, T. A Novel Appetite Peptide, Nesfatin-1 in Patients with Non-Alcoholic Fatty Liver Disease. Scand. J. Clin. Lab. Investig. 2012, 72, 479–483. [Google Scholar] [CrossRef]
- Alotibi, M.N.; Alnoury, A.M.; Alhozali, A.M. Serum nesfatin-1 and galanin concentrations in the adult with metabolic syndrome. Relationships to insulin resistance and obesity. Saudi Med. J. 2019, 40, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Gao, M.; Liu, D. Preventing High Fat Diet-Induced Obesity and Improving Insulin Sensitivity through Neuregulin 4 Gene Transfer. Sci. Rep. 2016, 6, 26242. [Google Scholar] [CrossRef]
- Cai, C.; Lin, M.; Xu, Y.; Li, X.; Yang, S.; Zhang, H. Association of Circulating Neuregulin 4 with Metabolic Syndrome in Obese Adults: A Cross-Sectional Study. BMC Med. 2016, 14, 165. [Google Scholar] [CrossRef]
- Seldin, M.M.; Peterson, J.M.; Byerly, M.S.; Wei, Z.; Wong, G.W. Myonectin (CTRP15), a Novel Myokine That Links Skeletal Muscle to Systemic Lipid Homeostasis. J. Biol. Chem. 2012, 287, 11968–11980. [Google Scholar] [CrossRef]
- Pourranjbar, M.; Arabnejad, N.; Naderipour, K.; Rafie, F. Effects of Aerobic Exercises on Serum Levels of Myonectin and Insulin Resistance in Obese and Overweight Women. J. Med. Life 2018, 11, 381–386. [Google Scholar] [CrossRef]
- Perakakis, N.; Triantafyllou, G.A.; Fernández-Real, J.M.; Huh, J.Y.; Park, K.H.; Seufert, J.; Mantzoros, C.S. Physiology and Role of Irisin in Glucose Homeostasis. Nat. Rev. Endocrinol. 2017, 13, 324–337. [Google Scholar] [CrossRef]
- Mazur-Bialy, A.I.; Bilski, J.; Pochec, E.; Brzozowski, T. New Insight into the Direct Anti-Inflammatory Activity of a Myokine Irisin against Proinflammatory Activation of Adipocytes. Implication for Exercise in Obesity. J. Physiol. Pharmacol. 2017, 68, 243–251. [Google Scholar] [PubMed]
- Mazur-Bialy, A.I. Irisin Acts as a Regulator of Macrophages Host Defense. Life Sci. 2017, 176, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Yosaee, S.; Basirat, R.; Hamidi, A.; Esteghamati, A.; Khodadost, M.; Shidfar, F.; Bitarafan, V.; Djafarian, K. Serum Irisin Levels in Metabolically Healthy versus Metabolically Unhealthy Obesity: A Case-Control Study. Med. J. Islam. Repub. Iran. 2020, 34, 46. [Google Scholar] [CrossRef] [PubMed]
- Castillo, E.C.; Elizondo-Montemayor, L.; Hernández-Brenes, C.; Rodríguez-Sánchez, D.G.; Silva-Platas, C.; Marín-Obispo, L.M.; Rodríguez-Gutierrez, N.A.; Treviño, V.; García-Rivas, G. Integrative Analysis of Lipid Profiles in Plasma Allows Cardiometabolic Risk Factor Clustering in Children with Metabolically Unhealthy Obesity. Oxid. Med. Cell Longev. 2020, 2020, 2935278. [Google Scholar] [CrossRef] [PubMed]
- Abulmeaty, M.M.A.; Aldisi, D.; Aljuraiban, G.S.; Almajwal, A.; El Shorbagy, E.; Almuhtadi, Y.; Albaran, B.; Aldossari, Z.; Alsager, T.; Razak, S.; et al. Association of Gastric Myoelectrical Activity with Ghrelin, Gastrin, and Irisin in Adults with Metabolically Healthy and Unhealthy Obesity. Front. Physiol. 2022, 13, 815026. [Google Scholar] [CrossRef]
- Frey, H.; Schroeder, N.; Manon-Jensen, T.; Iozzo, R.V.; Schaefer, L. Biological Interplay between Proteoglycans and Their Innate Immune Receptors in Inflammation. FEBS J. 2013, 280, 2165–2179. [Google Scholar] [CrossRef]
- Burmitskaya, Y.V.; Vasyukova, O.V.; Okorokov, P.L.; Zuraeva, Z.T.; Bezlepkina, O.B. Adipomyokines in children with obesity. Probl. Endokrinol. 2023, 69, 87–95. [Google Scholar] [CrossRef]
- Nakazato, M.; Hashimoto, K.; Shimizu, E.; Niitsu, T.; Iyo, M. Possible Involvement of Brain-Derived Neurotrophic Factor in Eating Disorders. IUBMB Life 2012, 64, 355–361. [Google Scholar] [CrossRef]
- Nicholson, J.R.; Peter, J.-C.; Lecourt, A.-C.; Barde, Y.-A.; Hofbauer, K.G. Melanocortin-4 Receptor Activation Stimulates Hypothalamic Brain-Derived Neurotrophic Factor Release to Regulate Food Intake, Body Temperature and Cardiovascular Function. J. Neuroendocrinol. 2007, 19, 974–982. [Google Scholar] [CrossRef]
- Corripio, R.; Gónzalez-Clemente, J.-M.; Jacobo, P.-S.; Silvia, N.; Lluis, G.; Joan, V.; Assumpta, C. Plasma Brain-Derived Neurotrophic Factor in Prepubertal Obese Children: Results from a 2-Year Lifestyle Intervention Programme. Clin. Endocrinol. 2012, 77, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, P.; Tortorella, A.; Martiadis, V.; Serritella, C.; Fuschino, A.; Maj, M. Opposite Changes in the Serum Brain-Derived Neurotrophic Factor in Anorexia Nervosa and Obesity. Psychosom. Med. 2004, 66, 744. [Google Scholar] [CrossRef] [PubMed]
- Sethi, J.K.; Vidal-Puig, A. Visfatin: The Missing Link between Intra-Abdominal Obesity and Diabetes? Trends Mol. Med. 2005, 11, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Dahl, T.B.; Yndestad, A.; Skjelland, M.; Øie, E.; Dahl, A.; Michelsen, A.; Damås, J.K.; Tunheim, S.H.; Ueland, T.; Smith, C.; et al. Increased Expression of Visfatin in Macrophages of Human Unstable Carotid and Coronary Atherosclerosis: Possible Role in Inflammation and Plaque Destabilization. Circulation 2007, 115, 972–980. [Google Scholar] [CrossRef] [PubMed]
- Lejawa, M.; Osadnik, K.; Czuba, Z.; Osadnik, T.; Pawlas, N. Association of Metabolically Healthy and Unhealthy Obesity Phenotype with Markers Related to Obesity, Diabetes among Young, Healthy Adult Men. Analysis of MAGNETIC Study. Life 2021, 11, 1350. [Google Scholar] [CrossRef]
- Nourbakhsh, M.; Nourbakhsh, M.; Gholinejad, Z.; Razzaghy-Azar, M. Visfatin in Obese Children and Adolescents and Its Association with Insulin Resistance and Metabolic Syndrome. Scand. J. Clin. Lab. Investig. 2015, 75, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Ernst, M.C.; Sinal, C.J. Chemerin: At the Crossroads of Inflammation and Obesity. Trends Endocrinol. Metab. 2010, 21, 660–667. [Google Scholar] [CrossRef]
- Sanidas, E.; Velliou, M.; Papadopoulos, D.; Anastasiou, T.; Michalaki, E.; Golemati, S.; Barbetseas, J. METABOLICALLY HEALTHY OBESITY AND RISK FACTORS. THE ROLE OF ADIPOSE TISSUE. J. Hypertens. 2022, 40 (Suppl. 1), e268. [Google Scholar] [CrossRef]
- Li, Y.; Shi, B.; Li, S. Association between Serum Chemerin Concentrations and Clinical Indices in Obesity or Metabolic Syndrome: A Meta-Analysis. PLoS ONE 2014, 9, e113915. [Google Scholar] [CrossRef]
- Harari, D.; Tzahar, E.; Romano, J.; Shelly, M.; Pierce, J.H.; Andrews, G.C.; Yarden, Y. Neuregulin-4: A Novel Growth Factor That Acts through the ErbB-4 Receptor Tyrosine Kinase. Oncogene 1999, 18, 2681–2689. [Google Scholar] [CrossRef]
- Hayes, N.V.L.; Blackburn, E.; Boyle, M.M.; Russell, G.A.; Frost, T.M.; Morgan, B.J.T.; Gullick, W.J. Expression of Neuregulin 4 Splice Variants in Normal Human Tissues and Prostate Cancer and Their Effects on Cell Motility. Endocr. Relat. Cancer 2011, 18, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Dunn, M.; Sinha, P.; Campbell, R.; Blackburn, E.; Levinson, N.; Rampaul, R.; Bates, T.; Humphreys, S.; Gullick, W.J. Co-Expression of Neuregulins 1, 2, 3 and 4 in Human Breast Cancer. J. Pathol. 2004, 203, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.O.; Friis-Hansen, L.; Poulsen, S.S.; Federspiel, B.; Sorensen, B.S. Expression of the EGF Family in Gastric Cancer: Downregulation of HER4 and Its Activating Ligand NRG4. PLoS ONE 2014, 9, e94606. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-X.; Zhao, X.-Y.; Meng, Z.-X.; Kern, M.; Dietrich, A.; Chen, Z.; Cozacov, Z.; Zhou, D.; Okunade, A.L.; Su, X.; et al. The Brown Fat-Enriched Secreted Factor Nrg4 Preserves Metabolic Homeostasis through Attenuating Hepatic Lipogenesis. Nat. Med. 2014, 20, 1436–1443. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Xu, Y.; Zhang, Z.; Gao, C.; Zhu, J.; Li, H.; Wan, Q. Decreased Plasma Neuregulin 4 Levels Are Associated with Peripheral Neuropathy in Chinese Patients with Newly Diagnosed Type 2 Diabetes: A Cross-Sectional Study. Cytokine 2019, 113, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Xu, Y.; Wan, Q.; Feng, J.; Li, H.; Yang, J.; Zhong, H.; Zhang, Z. Plasma Neuregulin 4 Levels Are Associated with Metabolic Syndrome in Patients Newly Diagnosed with Type 2 Diabetes Mellitus. Dis. Markers 2018, 2018, 6974191. [Google Scholar] [CrossRef]
- Park, T.-J.; Park, A.; Kim, J.; Kim, J.-Y.; Han, B.S.; Oh, K.-J.; Lee, E.W.; Lee, S.C.; Bae, K.-H.; Kim, W.K. Myonectin Inhibits Adipogenesis in 3T3-L1 Preadipocytes by Regulating P38 MAPK Pathway. BMB Rep. 2021, 54, 124–129. [Google Scholar] [CrossRef]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-Dependent Myokine That Drives Brown-Fat-like Development of White Fat and Thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Arias-Loste, M.T.; Ranchal, I.; Romero-Gómez, M.; Crespo, J. Irisin, a Link among Fatty Liver Disease, Physical Inactivity and Insulin Resistance. Int. J. Mol. Sci. 2014, 15, 23163–23178. [Google Scholar] [CrossRef]
- Yong Qiao, X.; Nie, Y.; Xian Ma, Y.; Chen, Y.; Cheng, R.; Yao Yinrg, W.; Hu, Y.; Ming Xu, W.; Zhi Xu, L. Irisin Promotes Osteoblast Proliferation and Differentiation via Activating the MAP Kinase Signaling Pathways. Sci. Rep. 2016, 6, 18732. [Google Scholar] [CrossRef]
- Maak, S.; Norheim, F.; Drevon, C.A.; Erickson, H.P. Progress and Challenges in the Biology of FNDC5 and Irisin. Endocr. Rev. 2021, 42, 436–456. [Google Scholar] [CrossRef]
- Jedrychowski, M.P.; Wrann, C.D.; Paulo, J.A.; Gerber, K.K.; Szpyt, J.; Robinson, M.M.; Nair, K.S.; Gygi, S.P.; Spiegelman, B.M. Detection and Quantitation of Circulating Human Irisin by Tandem Mass Spectrometry. Cell Metab. 2015, 22, 734–740. [Google Scholar] [CrossRef]
- Yu, Q.; Kou, W.; Xu, X.; Zhou, S.; Luan, P.; Xu, X.; Li, H.; Zhuang, J.; Wang, J.; Zhao, Y.; et al. FNDC5/Irisin Inhibits Pathological Cardiac Hypertrophy. Clin. Sci. 2019, 133, 611–627. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Zhang, J.; Ren, Y.; Du, Z.; Li, T.; Wang, T.; Zhang, L.; Wang, M.; Wu, Z.; Lv, Y.; et al. Irisin Reverses Intestinal Epithelial Barrier Dysfunction during Intestinal Injury via Binding to the Integrin αVβ5 Receptor. J. Cell. Mol. Med. 2020, 24, 996–1009. [Google Scholar] [CrossRef]
- Kim, H.; Wrann, C.D.; Jedrychowski, M.; Vidoni, S.; Kitase, Y.; Nagano, K.; Zhou, C.; Chou, J.; Parkman, V.-J.A.; Novick, S.J.; et al. Irisin Mediates Effects on Bone and Fat via αV Integrin Receptors. Cell 2018, 175, 1756–1768.e17. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, N.; Walden, T.B.; Shabalina, I.G.; Timmons, J.A.; Cannon, B.; Nedergaard, J. Chronic Peroxisome Proliferator-Activated Receptor γ (PPARγ) Activation of Epididymally Derived White Adipocyte Cultures Reveals a Population of Thermogenically Competent, UCP1-Containing Adipocytes Molecularly Distinct from Classic Brown Adipocytes. J. Biol. Chem. 2010, 285, 7153–7164. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Cohen, P.; Spiegelman, B.M. Adaptive Thermogenesis in Adipocytes: Is Beige the New Brown? Genes. Dev. 2013, 27, 234–250. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.-H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige Adipocytes Are a Distinct Type of Thermogenic Fat Cell in Mouse and Human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, F.W. Browning and Thermogenic Programing of Adipose Tissue. Best. Pract. Res. Clin. Endocrinol. Metab. 2016, 30, 479–485. [Google Scholar] [CrossRef]
- Tang, H.; Yu, R.; Liu, S.; Huwatibieke, B.; Li, Z.; Zhang, W. Irisin Inhibits Hepatic Cholesterol Synthesis via AMPK-SREBP2 Signaling. eBioMedicine 2016, 6, 139–148. [Google Scholar] [CrossRef]
- Xin, C.; Liu, J.; Zhang, J.; Zhu, D.; Wang, H.; Xiong, L.; Lee, Y.; Ye, J.; Lian, K.; Xu, C.; et al. Irisin Improves Fatty Acid Oxidation and Glucose Utilization in Type 2 Diabetes by Regulating the AMPK Signaling Pathway. Int. J. Obes. 2016, 40, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.-Y.; Shi, C.-X.; Gao, R.; Sun, H.-J.; Xiong, X.-Q.; Ding, L.; Chen, Q.; Li, Y.-H.; Wang, J.-J.; Kang, Y.-M.; et al. Irisin Inhibits Hepatic Gluconeogenesis and Increases Glycogen Synthesis via the PI3K/Akt Pathway in Type 2 Diabetic Mice and Hepatocytes. Clin. Sci. 2015, 129, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Sahar, N.E.; Javaid, H.M.A.; Pak, E.S.; Liang, G.; Wang, Y.; Ha, H.; Huh, J.Y. Exercise-Induced Irisin Decreases Inflammation and Improves NAFLD by Competitive Binding with MD2. Cells 2021, 10, 3306. [Google Scholar] [CrossRef]
- Liu, S.; Cui, F.; Ning, K.; Wang, Z.; Fu, P.; Wang, D.; Xu, H. Role of Irisin in Physiology and Pathology. Front. Endocrinol. 2022, 13, 962968. [Google Scholar] [CrossRef]
- Parr, E.B.; Camera, D.M.; Burke, L.M.; Phillips, S.M.; Coffey, V.G.; Hawley, J.A. Circulating MicroRNA Responses between “High” and “Low” Responders to a 16-Wk Diet and Exercise Weight Loss Intervention. PLoS ONE 2016, 11, e0152545. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Wang, X.; Wu, K.; Liu, K.; Wang, S.; Chen, X. Irisin Attenuates H2O2-Induced Apoptosis in Cardiomyocytes via microRNA-19b/AKT/mTOR Signaling Pathway. Int. J. Clin. Exp. Pathol. 2017, 10, 7707–7717. [Google Scholar] [PubMed]
- LeBleu, V.S.; O’Connell, J.T.; Gonzalez Herrera, K.N.; Wikman, H.; Pantel, K.; Haigis, M.C.; de Carvalho, F.M.; Damascena, A.; Domingos Chinen, L.T.; Rocha, R.M.; et al. PGC-1α Mediates Mitochondrial Biogenesis and Oxidative Phosphorylation in Cancer Cells to Promote Metastasis. Nat. Cell Biol. 2014, 16, 992–1003. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Ando, D.; Takamatsu, K.; Goto, K. Resistance Exercise Induces a Greater Irisin Response than Endurance Exercise. Metab. Clin. Exp. 2015, 64, 1042–1050. [Google Scholar] [CrossRef]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic Inflammation in Fat Plays a Crucial Role in the Development of Obesity-Related Insulin Resistance. J. Clin. Investig. 2003, 112, 1821–1830. [Google Scholar] [CrossRef]
- Smitka, K.; Marešová, D. Adipose Tissue as an Endocrine Organ: An Update on Pro-Inflammatory and Anti-Inflammatory Microenvironment. Prague Med. Rep. 2015, 116, 87–111. [Google Scholar] [CrossRef]
- Kern, P.A.; Ranganathan, S.; Li, C.; Wood, L.; Ranganathan, G. Adipose Tissue Tumor Necrosis Factor and Interleukin-6 Expression in Human Obesity and Insulin Resistance. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E745–E751. [Google Scholar] [CrossRef]
- Belviranli, M.; Okudan, N.; Çelik, F. Association of Circulating Irisin with Insulin Resistance and Oxidative Stress in Obese Women. Horm. Metab. Res. 2016, 48, 653–657. [Google Scholar] [CrossRef]
- Khajebishak, Y.; Faghfouri, A.H.; Soleimani, A.; Ilaei, S.; Peyrovi, S.; Madani, S.; Payahoo, L. The Potential Relationship between Serum Irisin Concentration with Inflammatory Cytokines, Oxidative Stress Biomarkers, Glycemic Indices and Lipid Profiles in Obese Patients with Type 2 Diabetes Mellitus: A Pilot Study. J. ASEAN Fed. Endocr. Soc. 2023, 38, 45–51. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Y.; Wang, F.; Donelan, W.; Zona, M.C.; Li, S.; Reeves, W.; Ding, Y.; Tang, D.; Yang, L. Effects of Irisin on the Differentiation and Browning of Human Visceral White Adipocytes. Am. J. Transl. Res. 2019, 11, 7410–7421. [Google Scholar] [PubMed]
- Zhang, H.-J.; Zhang, X.-F.; Ma, Z.-M.; Pan, L.-L.; Chen, Z.; Han, H.-W.; Han, C.-K.; Zhuang, X.-J.; Lu, Y.; Li, X.-J.; et al. Irisin Is Inversely Associated with Intrahepatic Triglyceride Contents in Obese Adults. J. Hepatol. 2013, 59, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Shanaki, M.; Moradi, N.; Emamgholipour, S.; Fadaei, R.; Poustchi, H. Lower Circulating Irisin Is Associated with Nonalcoholic Fatty Liver Disease and Type 2 Diabetes. Diabetes Metab. Syndr. Clin. Res. Rev. 2017, 11, S467–S472. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, S.A.; Kountouras, J.; Anastasilakis, A.D.; Geladari, E.V.; Mantzoros, C.S. Irisin in Patients with Nonalcoholic Fatty Liver Disease. Metab. Clin. Exp. 2014, 63, 207–217. [Google Scholar] [CrossRef]
- Jia, J.; Yu, F.; Wei, W.-P.; Yang, P.; Zhang, R.; Sheng, Y.; Shi, Y.-Q. Relationship between Circulating Irisin Levels and Overweight/Obesity: A Meta-Analysis. World J. Clin. Cases 2019, 7, 1444–1455. [Google Scholar] [CrossRef] [PubMed]
- Stengel, A.; Hofmann, T.; Goebel-Stengel, M.; Elbelt, U.; Kobelt, P.; Klapp, B.F. Circulating Levels of Irisin in Patients with Anorexia Nervosa and Different Stages of Obesity—Correlation with Body Mass Index. Peptides 2013, 39, 125–130. [Google Scholar] [CrossRef]
- Crujeiras, A.B.; Pardo, M.; Casanueva, F.F. Irisin: “fat” or Artefact. Clin. Endocrinol. 2015, 82, 467–474. [Google Scholar] [CrossRef]
- Montes-Nieto, R.; Martínez-García, M.Á.; Luque-Ramírez, M.; Escobar-Morreale, H.F. Differences in Analytical and Biological Results between Older and Newer Lots of a Widely Used Irisin Immunoassay Question the Validity of Previous Studies. Clin. Chem. Lab. Med. (CCLM) 2016, 54, e199–e201. [Google Scholar] [CrossRef] [PubMed]
- Mehrabian, S.; Taheri, E.; Karkhaneh, M.; Qorbani, M.; Hosseini, S. Association of Circulating Irisin Levels with Normal Weight Obesity, Glycemic and Lipid Profile. J. Diabetes Metab. Disord. 2016, 15, 17. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Zhong, J.; Dong, L. The Role of Decorin in Autoimmune and Inflammatory Diseases. J. Immunol. Res. 2022, 2022, 1283383. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ge, Y.; Cheng, Q.; Zhang, Q.; Fang, L.; Zheng, J. Decorin Is a Pivotal Effector in the Extracellular Matrix and Tumour Microenvironment. Oncotarget 2018, 9, 5480–5491. [Google Scholar] [CrossRef] [PubMed]
- Bolton, K.; Segal, D.; McMillan, J.; Jowett, J.; Heilbronn, L.; Abberton, K.; Zimmet, P.; Chisholm, D.; Collier, G.; Walder, K. Decorin Is a Secreted Protein Associated with Obesity and Type 2 Diabetes. Int. J. Obes. 2008, 32, 1113–1121. [Google Scholar] [CrossRef]
- Svärd, J.; Røst, T.H.; Sommervoll, C.E.N.; Haugen, C.; Gudbrandsen, O.A.; Mellgren, A.E.; Rødahl, E.; Fernø, J.; Dankel, S.N.; Sagen, J.V.; et al. Absence of the Proteoglycan Decorin Reduces Glucose Tolerance in Overfed Male Mice. Sci. Rep. 2019, 9, 4614. [Google Scholar] [CrossRef]
- Iozzo, R.V.; Schaefer, L. Proteoglycans in Health and Disease: Novel Regulatory Signaling Mechanisms Evoked by the Small Leucine-Rich Proteoglycans. FEBS J. 2010, 277, 3864–3875. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, S.; Zeng, L.; Li, J.; Klionsky, D.J.; Kroemer, G.; Jiang, J.; Tang, D.; Kang, R. DCN Released from Ferroptotic Cells Ignites AGER-Dependent Immune Responses. Autophagy 2022, 18, 2036–2049. [Google Scholar] [CrossRef]
- Hirata, B.K.S.; Pedroso, A.P.; Machado, M.M.F.; Neto, N.I.P.; Perestrelo, B.O.; de Sá, R.D.C.C.; Alonso-Vale, M.I.C.; Nogueira, F.N.; Oyama, L.M.; Ribeiro, E.B.; et al. Ginkgo Biloba Extract Modulates the Retroperitoneal Fat Depot Proteome and Reduces Oxidative Stress in Diet-Induced Obese Rats. Front. Pharmacol. 2019, 10, 686. [Google Scholar] [CrossRef]
- Zhang, J.; Wright, W.; Bernlohr, D.A.; Cushman, S.W.; Chen, X. Alterations of the Classic Pathway of Complement in Adipose Tissue of Obesity and Insulin Resistance. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E1433–E1440. [Google Scholar] [CrossRef]
- Groeneveld, T.W.L.; Oroszlán, M.; Owens, R.T.; Faber-Krol, M.C.; Bakker, A.C.; Arlaud, G.J.; McQuillan, D.J.; Kishore, U.; Daha, M.R.; Roos, A. Interactions of the Extracellular Matrix Proteoglycans Decorin and Biglycan with C1q and Collectins. J. Immunol. 2005, 175, 4715–4723. [Google Scholar] [CrossRef]
- Ward, M.; Ajuwon, K.M. Regulation of Pre-adipocyte Proliferation and Apoptosis by the Small Leucine-rich Proteoglycans, Biglycan and Decorin. Cell Prolif. 2011, 44, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Leibrock, J.; Lottspeich, F.; Hohn, A.; Hofer, M.; Hengerer, B.; Masiakowski, P.; Thoenen, H.; Barde, Y.A. Molecular Cloning and Expression of Brain-Derived Neurotrophic Factor. Nature 1989, 341, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, P.; Ng, C.F.; Pang, B.P.S.; Chan, W.S.; Tse, M.C.L.; Bi, X.; Kwan, H.-L.R.; Brobst, D.; Herlea-Pana, O.; Yang, X.; et al. Muscle-Generated BDNF (Brain Derived Neurotrophic Factor) Maintains Mitochondrial Quality Control in Female Mice. Autophagy 2021, 18, 1367–1384. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Bomberg, E.; Billington, C.; Levine, A.; Kotz, C.M. Brain-Derived Neurotrophic Factor in the Hypothalamic Paraventricular Nucleus Increases Energy Expenditure by Elevating Metabolic Rate. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R992–R1002. [Google Scholar] [CrossRef]
- Xu, B.; Xie, X. Neurotrophic Factor Control of Satiety and Body Weight. Nat. Rev. Neurosci. 2016, 17, 282–292. [Google Scholar] [CrossRef]
- Yiannakas, A.; Rosenblum, K. The Insula and Taste Learning. Front. Mol. Neurosci. 2017, 10, 335. [Google Scholar] [CrossRef] [PubMed]
- Bumb, J.M.; Bach, P.; Grosshans, M.; Wagner, X.; Koopmann, A.; Vollstädt-Klein, S.; Schuster, R.; Wiedemann, K.; Kiefer, F. BDNF Influences Neural Cue-Reactivity to Food Stimuli and Food Craving in Obesity. Eur. Arch. Psychiatry Clin. Neurosci. 2021, 271, 963–974. [Google Scholar] [CrossRef]
- Thorleifsson, G.; Walters, G.B.; Gudbjartsson, D.F.; Steinthorsdottir, V.; Sulem, P.; Helgadottir, A.; Styrkarsdottir, U.; Gretarsdottir, S.; Thorlacius, S.; Jonsdottir, I.; et al. Genome-Wide Association Yields New Sequence Variants at Seven Loci That Associate with Measures of Obesity. Nat. Genet. 2009, 41, 18–24. [Google Scholar] [CrossRef]
- Speliotes, E.K.; Willer, C.J.; Berndt, S.I.; Monda, K.L.; Thorleifsson, G.; Jackson, A.U.; Allen, H.L.; Lindgren, C.M.; Luan, J.; Mägi, R.; et al. Association Analyses of 249,796 Individuals Reveal Eighteen New Loci Associated with Body Mass Index. Nat. Genet. 2010, 42, 937–948. [Google Scholar] [CrossRef]
- Yeo, G.S.H.; Connie Hung, C.-C.; Rochford, J.; Keogh, J.; Gray, J.; Sivaramakrishnan, S.; O’Rahilly, S.; Farooqi, I.S. A de Novo Mutation Affecting Human TrkB Associated with Severe Obesity and Developmental Delay. Nat. Neurosci. 2004, 7, 1187–1189. [Google Scholar] [CrossRef] [PubMed]
- Sandrini, L.; Di Minno, A.; Amadio, P.; Ieraci, A.; Tremoli, E.; Barbieri, S.S. Association between Obesity and Circulating Brain-Derived Neurotrophic Factor (BDNF) Levels: Systematic Review of Literature and Meta-Analysis. Int. J. Mol. Sci. 2018, 19, 2281. [Google Scholar] [CrossRef] [PubMed]
- Elsner, V.R.; Dorneles, G.P.; Santos, M.A.; da Silva, I.M.; Romão, P.; Peres, A. Exercise-Induced BDNF Production by Mononuclear Cells of Sedentary and Physically Active Obese Men. J. Sports Med. Phys. Fitness 2020, 60, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Ognjanovic, S.; Bao, S.; Yamamoto, S.Y.; Garibay-Tupas, J.; Samal, B.; Bryant-Greenwood, G.D. Genomic Organization of the Gene Coding for Human Pre-B-Cell Colony Enhancing Factor and Expression in Human Fetal Membranes. J. Mol. Endocrinol. 2001, 26, 107–117. [Google Scholar] [CrossRef]
- Kralisch, S.; Klein, J.; Lossner, U.; Bluher, M.; Paschke, R.; Stumvoll, M.; Fasshauer, M. Interleukin-6 Is a Negative Regulator of Visfatin Gene Expression in 3T3-L1 Adipocytes. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E586–E590. [Google Scholar] [CrossRef]
- Abdalla, M.M.I. Role of Visfatin in Obesity-Induced Insulin Resistance. World J. Clin. Cases 2022, 10, 10840–10851. [Google Scholar] [CrossRef]
- Zhou, F.; Pan, Y.; Huang, Z.; Jia, Y.; Zhao, X.; Chen, Y.; Diao, J.; Wan, Q.; Cui, X. Visfatin Induces Cholesterol Accumulation in Macrophages through Up-Regulation of Scavenger Receptor-A and CD36. Cell Stress. Chaperones 2013, 18, 643–652. [Google Scholar] [CrossRef]
- Romacho, T.; Sánchez-Ferrer, C.F.; Peiró, C. Visfatin/Nampt: An Adipokine with Cardiovascular Impact. Mediat. Inflamm. 2013, 2013, 946427. [Google Scholar] [CrossRef]
- Chen, S.; Sun, L.; Gao, H.; Ren, L.; Liu, N.; Song, G. Visfatin and Oxidative Stress Influence Endothelial Progenitor Cells in Obese Populations. Endocr. Res. 2015, 40, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Heo, Y.J.; Choi, S.-E.; Jeon, J.Y.; Han, S.J.; Kim, D.J.; Kang, Y.; Lee, K.W.; Kim, H.J. Visfatin Induces Inflammation and Insulin Resistance via the NF-κB and STAT3 Signaling Pathways in Hepatocytes. J. Diabetes Res. 2019, 2019, 4021623. [Google Scholar] [CrossRef]
- Chang, Y.-C.; Chang, T.-J.; Lee, W.-J.; Chuang, L.-M. The Relationship of Visfatin/Pre-B-Cell Colony-Enhancing Factor/Nicotinamide Phosphoribosyltransferase in Adipose Tissue with Inflammation, Insulin Resistance, and Plasma Lipids. Metabolism 2010, 59, 93–99. [Google Scholar] [CrossRef] [PubMed]
- El-Mesallamy, H.O.; Kassem, D.H.; El-Demerdash, E.; Amin, A.I. Vaspin and Visfatin/Nampt Are Interesting Interrelated Adipokines Playing a Role in the Pathogenesis of Type 2 Diabetes Mellitus. Metabolism 2011, 60, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Haider, D.G.; Schindler, K.; Schaller, G.; Prager, G.; Wolzt, M.; Ludvik, B. Increased Plasma Visfatin Concentrations in Morbidly Obese Subjects Are Reduced after Gastric Banding. J. Clin. Endocrinol. Metab. 2006, 91, 1578–1581. [Google Scholar] [CrossRef] [PubMed]
- Salama, H.M.; Galal, A.; Motawie, A.A.; Kamel, A.F.; Ibrahim, D.M.; Aly, A.A.; Hassan, E.A. Adipokines Vaspin and Visfatin in Obese Children. Open Access Maced. J. Med. Sci. 2015, 3, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Hu, W.; Wang, M.; Xiao, Y. The Role of the Adipocytokines Vaspin and Visfatin in Vascular Endothelial Function and Insulin Resistance in Obese Children. BMC Endocr. Disord. 2019, 19, 127. [Google Scholar] [CrossRef]
- Alnowihi, S.M.; Al Doghaither, H.A.; Osman, N.N. Serum Visfatin Concentration and Its Relationship with Sex Hormones in Obese Saudi Women. Int. J. Health Sci. 2020, 14, 9–13. [Google Scholar]
- Kim, J.-E.; Kim, J.-S.; Jo, M.-J.; Cho, E.; Ahn, S.-Y.; Kwon, Y.-J.; Ko, G.-J. The Roles and Associated Mechanisms of Adipokines in Development of Metabolic Syndrome. Molecules 2022, 27, 334. [Google Scholar] [CrossRef] [PubMed]
- Zabel, B.A.; Kwitniewski, M.; Banas, M.; Zabieglo, K.; Murzyn, K.; Cichy, J. Chemerin Regulation and Role in Host Defense. Am. J. Clin. Exp. Immunol. 2014, 3, 1–19. [Google Scholar]
- Ernst, M.C.; Issa, M.; Goralski, K.B.; Sinal, C.J. Chemerin Exacerbates Glucose Intolerance in Mouse Models of Obesity and Diabetes. Endocrinology 2010, 151, 1998–2007. [Google Scholar] [CrossRef]
- Bondue, B.; Wittamer, V.; Parmentier, M. Chemerin and Its Receptors in Leukocyte Trafficking, Inflammation and Metabolism. Cytokine Growth Factor. Rev. 2011, 22, 331–338. [Google Scholar] [CrossRef]
- Mariani, F.; Roncucci, L. Chemerin/chemR23 Axis in Inflammation Onset and Resolution. Inflamm. Res. 2015, 64, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Adya, R.; Tan, B.K.; Chen, J.; Randeva, H.S. Identification of Chemerin Receptor (ChemR23) in Human Endothelial Cells: Chemerin-Induced Endothelial Angiogenesis. Biochem. Biophys. Res. Commun. 2010, 391, 1762–1768. [Google Scholar] [CrossRef]
- Weigert, J.; Neumeier, M.; Wanninger, J.; Filarsky, M.; Bauer, S.; Wiest, R.; Farkas, S.; Scherer, M.N.; Schäffler, A.; Aslanidis, C.; et al. Systemic Chemerin Is Related to Inflammation Rather than Obesity in Type 2 Diabetes. Clin. Endocrinol. 2010, 72, 342–348. [Google Scholar] [CrossRef]
- Bozaoglu, K.; Curran, J.E.; Stocker, C.J.; Zaibi, M.S.; Segal, D.; Konstantopoulos, N.; Morrison, S.; Carless, M.; Dyer, T.D.; Cole, S.A.; et al. Chemerin, a Novel Adipokine in the Regulation of Angiogenesis. J. Clin. Endocrinol. Metab. 2010, 95, 2476–2485. [Google Scholar] [CrossRef] [PubMed]
- Fatima, S.S.; Bozaoglu, K.; Rehman, R.; Alam, F.; Memon, A.S. Elevated Chemerin Levels in Pakistani Men: An Interrelation with Metabolic Syndrome Phenotypes. PLoS ONE 2013, 8, e57113. [Google Scholar] [CrossRef]
- Tönjes, A.; Scholz, M.; Breitfeld, J.; Marzi, C.; Grallert, H.; Gross, A.; Ladenvall, C.; Schleinitz, D.; Krause, K.; Kirsten, H.; et al. Genome Wide Meta-Analysis Highlights the Role of Genetic Variation in RARRES2 in the Regulation of Circulating Serum Chemerin. PLoS Genet. 2014, 10, e1004854. [Google Scholar] [CrossRef] [PubMed]
- Niklowitz, P.; Rothermel, J.; Lass, N.; Barth, A.; Reinehr, T. Link between Chemerin, Central Obesity, and Parameters of the Metabolic Syndrome: Findings from a Longitudinal Study in Obese Children Participating in a Lifestyle Intervention. Int. J. Obes. 2018, 42, 1743–1752. [Google Scholar] [CrossRef]
- Fülöp, P.; Seres, I.; Lőrincz, H.; Harangi, M.; Somodi, S.; Paragh, G. Association of Chemerin with Oxidative Stress, Inflammation and Classical Adipokines in Non-Diabetic Obese Patients. J. Cell Mol. Med. 2014, 18, 1313–1320. [Google Scholar] [CrossRef]
- Corona-Meraz, F.-I.; Navarro-Hernández, R.-E.; Ruíz-Quezada, S.-L.; Madrigal-Ruíz, P.-M.; Castro-Albarrán, J.; Chavarría-Ávila, E.; Guzmán-Ornelas, M.-O.; Gómez-Bañuelos, E.; Petri, M.-H.; Ramírez-Cedano, J.-I.; et al. Inverse Relationship of the CMKLR1 Relative Expression and Chemerin Serum Levels in Obesity with Dysmetabolic Phenotype and Insulin Resistance. Mediators Inflamm. 2016, 2016, 3085390. [Google Scholar] [CrossRef]
- Ernst, M.C.; Haidl, I.D.; Zúñiga, L.A.; Dranse, H.J.; Rourke, J.L.; Zabel, B.A.; Butcher, E.C.; Sinal, C.J. Disruption of the Chemokine-Like Receptor-1 (CMKLR1) Gene Is Associated with Reduced Adiposity and Glucose Intolerance. Endocrinology 2012, 153, 672–682. [Google Scholar] [CrossRef]

| NWO | MONW | MHO | MUO | |
|---|---|---|---|---|
| BMI (kg/m2) | <25 (normal) | <25 (normal) | >30 (elevated) | >30 (elevated) |
| Body AT percentage | ↑ | ↑ | ↑ | ↑ |
| VAT | ↑ | ↑ | Lower compared to MUO | ↑ |
| SAT | ↓ | ↑ | Higher compared to MUO | ↓ |
| Insulin sensitivity | ↓ | ↓ | Favorable | ↓ |
| Inflammatory markers | ↑ | ↑ | Normal | ↑ |
| Blood pressure | ↑ | ↑ | Normal | ↑ |
| MetS | − | + | − | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pelczyńska, M.; Miller-Kasprzak, E.; Piątkowski, M.; Mazurek, R.; Klause, M.; Suchecka, A.; Bucoń, M.; Bogdański, P. The Role of Adipokines and Myokines in the Pathogenesis of Different Obesity Phenotypes—New Perspectives. Antioxidants 2023, 12, 2046. https://doi.org/10.3390/antiox12122046
Pelczyńska M, Miller-Kasprzak E, Piątkowski M, Mazurek R, Klause M, Suchecka A, Bucoń M, Bogdański P. The Role of Adipokines and Myokines in the Pathogenesis of Different Obesity Phenotypes—New Perspectives. Antioxidants. 2023; 12(12):2046. https://doi.org/10.3390/antiox12122046
Chicago/Turabian StylePelczyńska, Marta, Ewa Miller-Kasprzak, Marcin Piątkowski, Roksana Mazurek, Mateusz Klause, Anna Suchecka, Magdalena Bucoń, and Paweł Bogdański. 2023. "The Role of Adipokines and Myokines in the Pathogenesis of Different Obesity Phenotypes—New Perspectives" Antioxidants 12, no. 12: 2046. https://doi.org/10.3390/antiox12122046