
Role of Platinum Nanozymes in the Oxidative Stress Response of Salmonella Typhimurium
 , , ,
, , ,  , and
, and 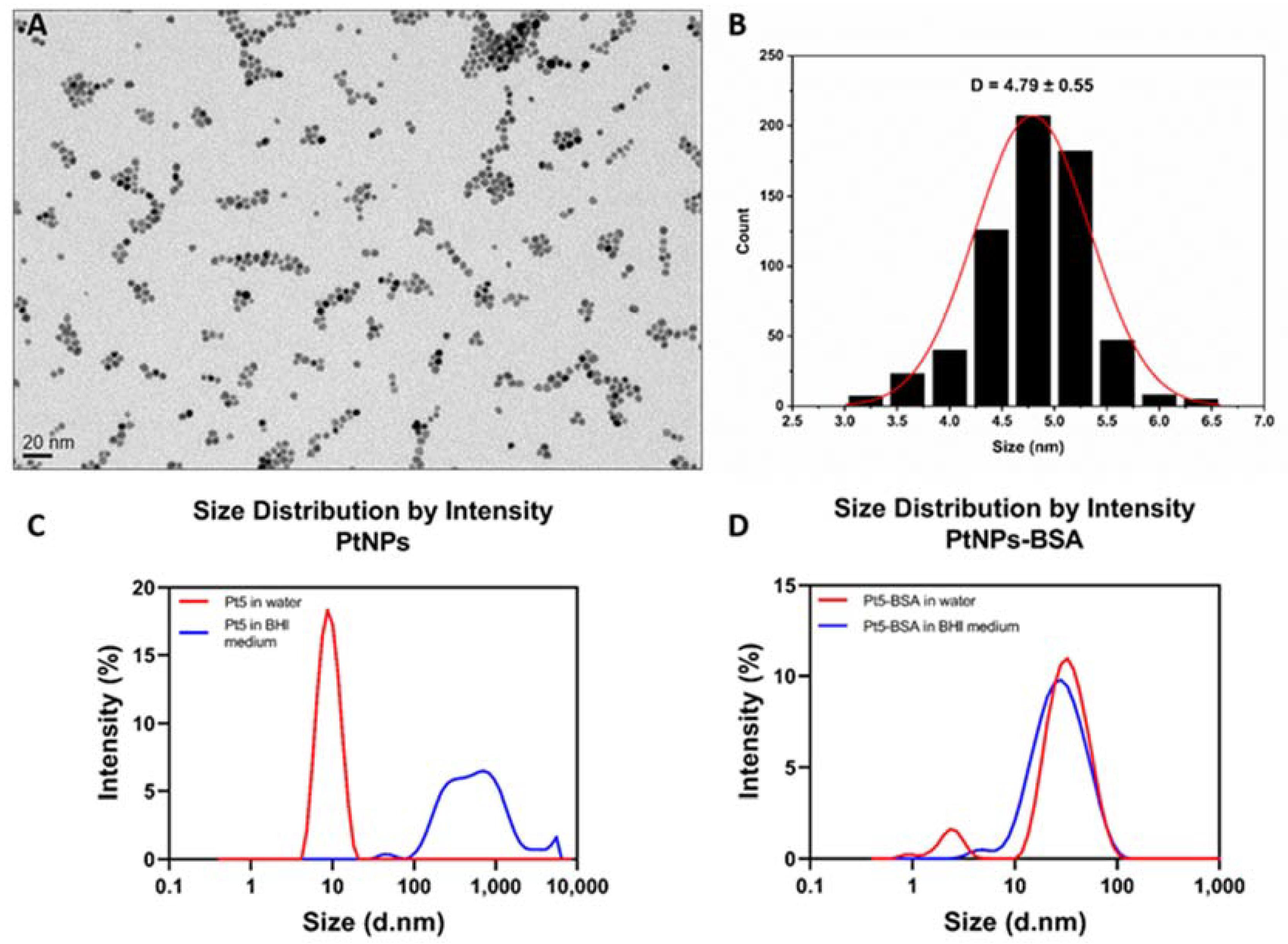{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Platinum Nanoparticle Synthesis, Functionalization and Characterization
2.3. Biocidal Effect of PtNPs
2.4. Hydrogen Peroxide Scavenging and Sensitivity
2.5. Untargeted Metabolomics by UHPLC-HRMS
2.6. Malondialdehyde TBARS Assay
2.7. Statistical Analysis
3. Results and Discussion
3.1. Synthesis and Characterization of Platinum Nanoparticles
3.2. Effect of PtNPs on Salmonella Typhimurium Growth
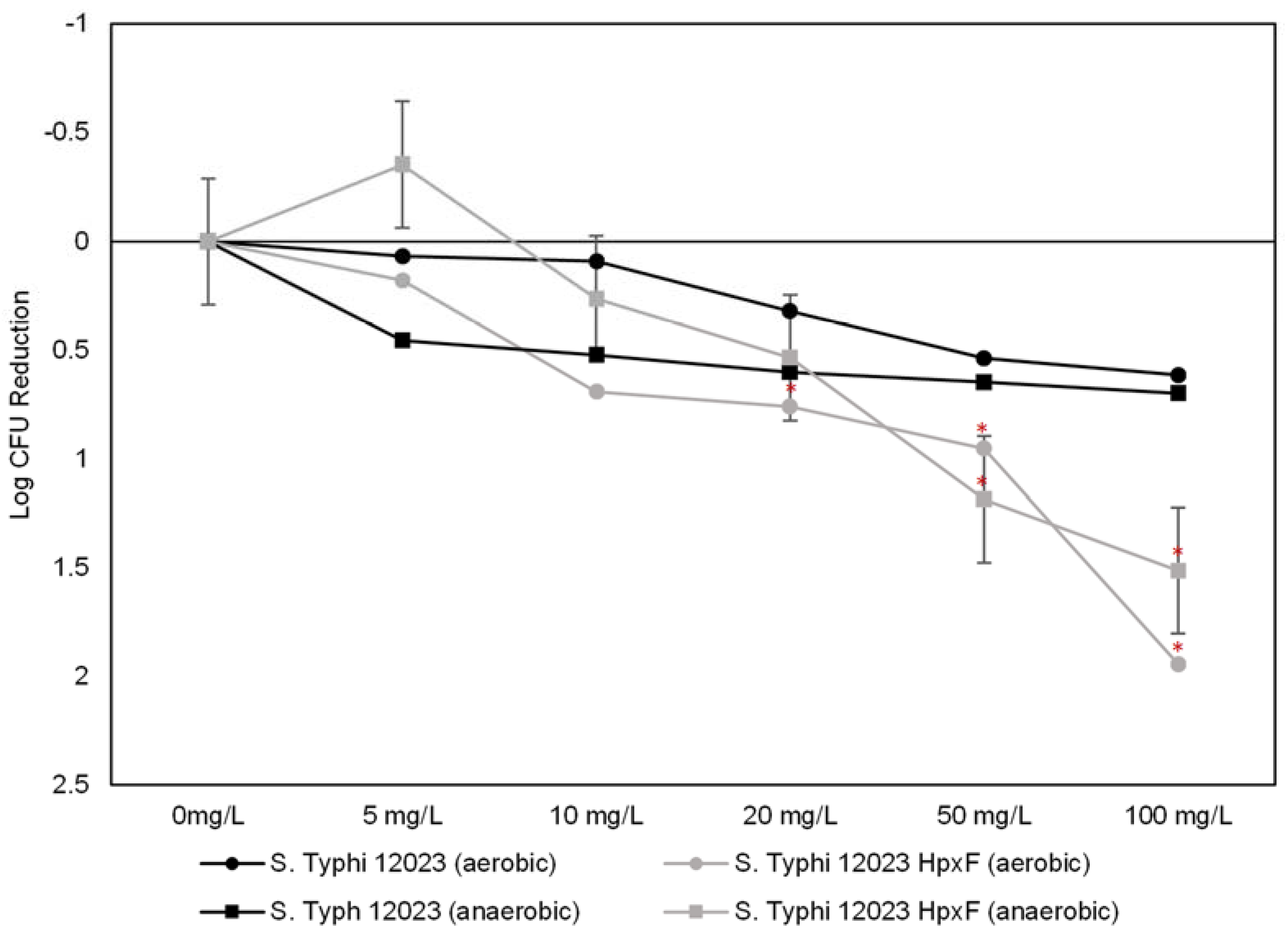
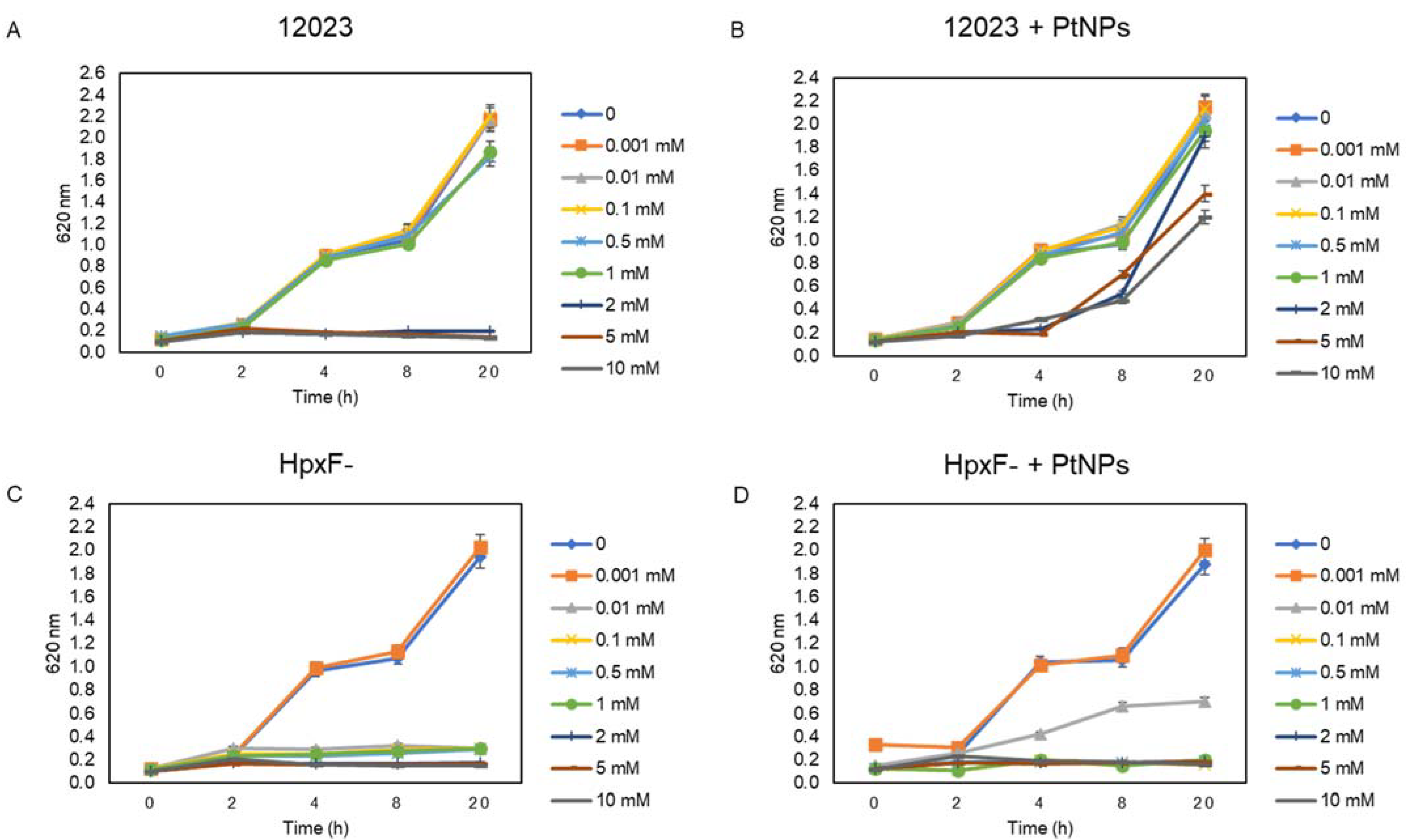
3.3. The Combined Effect of PtNPs and H2O2 on Salmonella Typhimurium
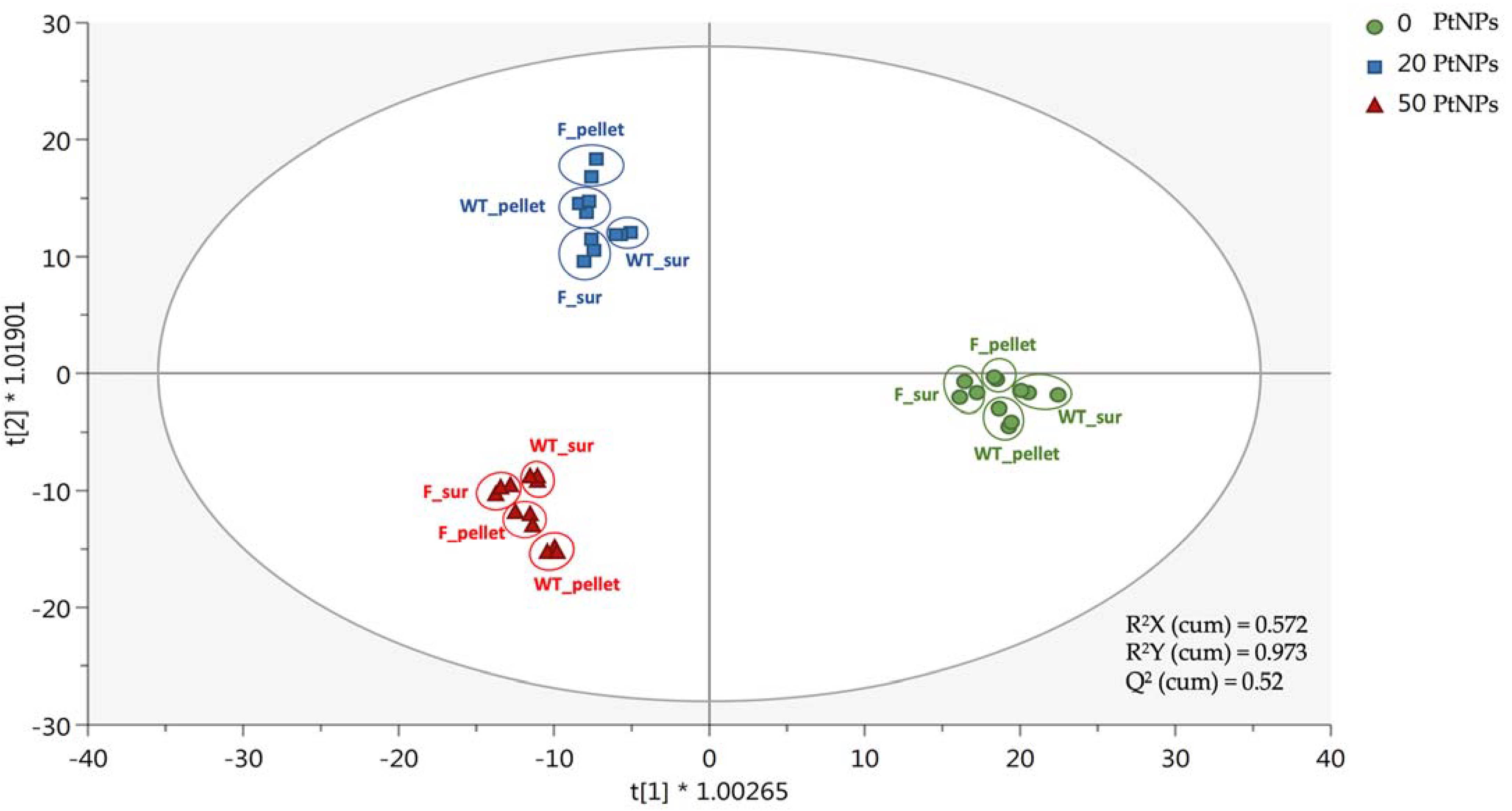
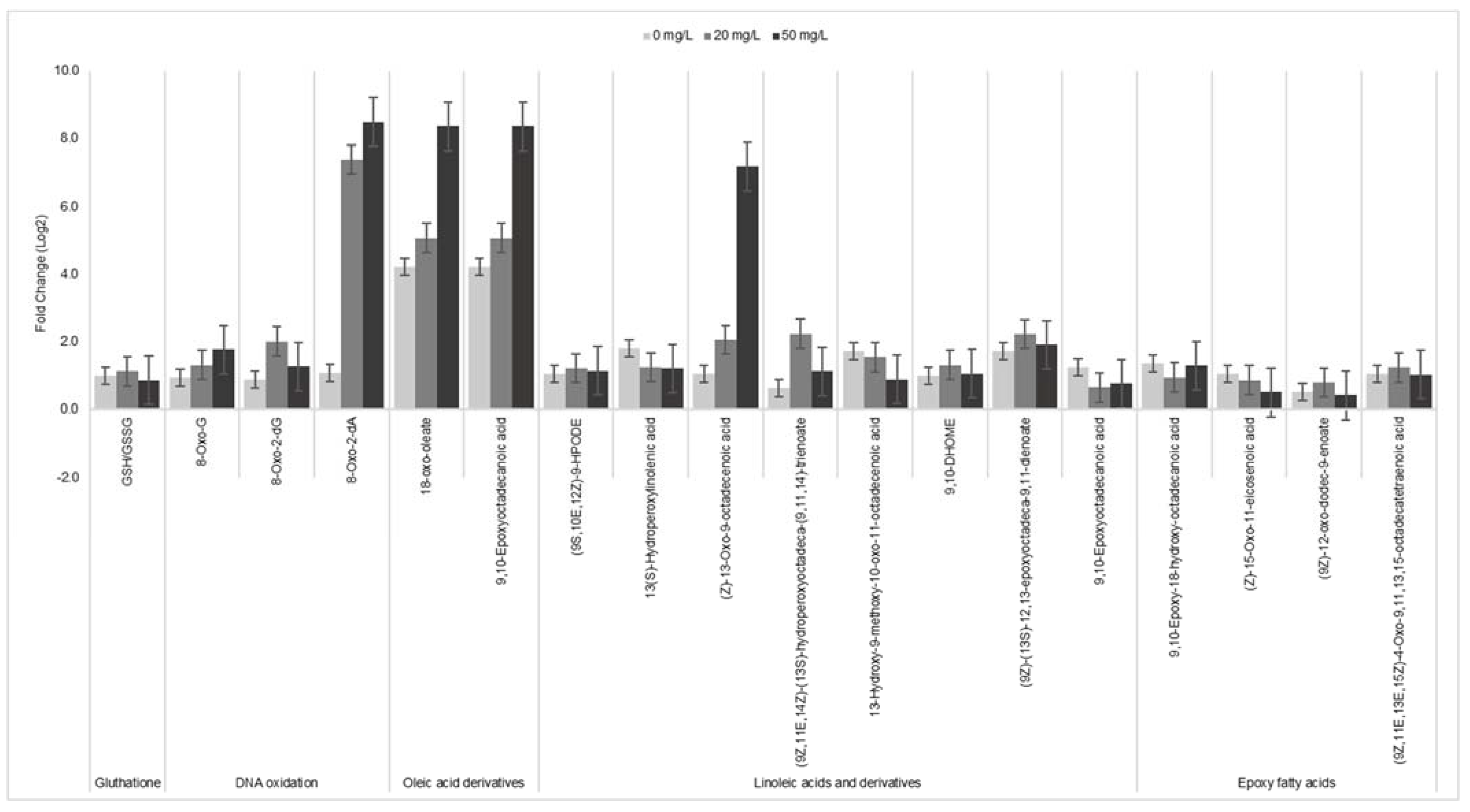
3.4. PtNPs-Induced ROS, Affect Membrane Lipids and Oxidize DNA
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ozdal, M.; Gurkok, S. Recent advances in nanoparticles as antibacterial agent. ADMET DMPK 2022, 10, 115. [Google Scholar] [CrossRef]
- Manke, A.; Wang, L.; Rojanasakul, Y. Mechanisms of nanoparticle-induced oxidative stress and toxicity. Biomed Res. Int. 2013, 2013, 942916. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-López, E.; Gomes, D.; Esteruelas, G.; Bonilla, L.; Lopez-Machado, A.L.; Galindo, R.; Cano, A.; Espina, M.; Ettcheto, M.; Camins, A.; et al. Metal-Based Nanoparticles as Antimicrobial Agents: An Overview. Nanomaterials 2020, 10, 292. [Google Scholar] [CrossRef] [Green Version]
- Beyth, N.; Houri-Haddad, Y.; Domb, A.; Khan, W.; Hazan, R. Alternative antimicrobial approach: Nano-antimicrobial materials. Evid. Based. Complement. Alternat. Med. 2015, 2015, 246012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, P.; Huang, X.; Ye, N.; Lu, Q.; Zhang, G.; Peng, S.; Wang, H.; Liu, Y. Cytotoxicity of Metal-Based Nanoparticles: From Mechanisms and Methods of Evaluation to Pathological Manifestations. Adv. Sci. 2022, 9, 2106049. [Google Scholar] [CrossRef] [PubMed]
- Dror-Ehre, A.; Mamane, H.; Belenkova, T.; Markovich, G.; Adin, A. Silver nanoparticle-E. coli colloidal interaction in water and effect on E. coli survival. J. Colloid Interface Sci. 2009, 339, 521–526. [Google Scholar] [CrossRef]
- El Badawy, A.M.; Silva, R.G.; Morris, B.; Scheckel, K.G.; Suidan, M.T.; Tolaymat, T.M. Surface charge-dependent toxicity of silver nanoparticles. Environ. Sci. Technol. 2011, 45, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Dallas, P.; Sharma, V.K.; Zboril, R. Silver polymeric nanocomposites as advanced antimicrobial agents: Classification, synthetic paths, applications, and perspectives. Adv. Colloid Interface Sci. 2011, 166, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Navarro, E.; Piccapietra, F.; Wagner, B.; Marconi, F.; Kaegi, R.; Odzak, N.; Sigg, L.; Behra, R. Toxicity of silver nanoparticles to Chlamydomonas reinhardtii. Environ. Sci. Technol. 2008, 42, 8959–8964. [Google Scholar] [CrossRef]
- Buckley, J.J.; Lee, A.F.; Olivi, L.; Wilson, K. Hydroxyapatite supported antibacterial Ag3PO4 nanoparticles. J. Mater. Chem. 2010, 20, 8056–8063. [Google Scholar] [CrossRef] [Green Version]
- Klasen, H.J. A historical review of the use of silver in the treatment of burns. II. Renewed interest for silver. Burns 2000, 26, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.; Goshisht, M.K. Recent Advances and Mechanistic Insights into Antibacterial Activity, Antibiofilm Activity, and Cytotoxicity of Silver Nanoparticles. ACS Appl. Bio Mater. 2022, 5, 1391–1463. [Google Scholar] [CrossRef]
- Rizzello, L.; Pompa, P.P. Nanosilver-based antibacterial drugs and devices: Mechanisms, methodological drawbacks, and guidelines. Chem. Soc. Rev. 2014, 43, 1501. [Google Scholar] [CrossRef] [PubMed]
- Kumawat, M.; Madhyastha, H.; Singh, M.; Revaprasadu, N.; Srinivas, S.P.; Kumar, H.; Id, D. Double functionalized haemocompatible silver nanoparticles control cell inflammatory homeostasis. PloS ONE 2022, 17, e0276296. [Google Scholar] [CrossRef]
- Estevez, M.B.; Casaux, M.L.; Fraga, M.; Faccio, R.; Alborés, S. Biogenic Silver Nanoparticles as a Strategy in the Fight Against Multi-Resistant Salmonella enterica Isolated From Dairy Calves. Front. Bioeng. Biotechnol. 2021, 9, 314. [Google Scholar] [CrossRef]
- Seong, M.; Lee, D.G. Silver Nanoparticles Against Salmonella enterica Serotype Typhimurium: Role of Inner Membrane Dysfunction. Curr. Microbiol. 2017, 74, 661–670. [Google Scholar] [CrossRef]
- Panáček, A.; Kvítek, L.; Smékalová, M.; Večeřová, R.; Kolář, M.; Röderová, M.; Dyčka, F.; Šebela, M.; Prucek, R.; Tomanec, O.; et al. Bacterial resistance to silver nanoparticles and how to overcome it. Nat. Nanotechnol. 2018, 13, 65–71. [Google Scholar] [CrossRef]
- Wu, J.; Hou, S.; Ren, D.; Mather, P.T. Antimicrobial properties of nanostructured hydrogel webs containing silver. Biomacromolecules 2009, 10, 2686–2693. [Google Scholar] [CrossRef]
- Holt, K.B.; Bard, A.J. Interaction of silver(I) ions with the respiratory chain of Escherichia coli: An electrochemical and scanning electrochemical microscopy study of the antimicrobial mechanism of micromolar Ag+. Biochemistry 2005, 44, 13214–13223. [Google Scholar] [CrossRef] [PubMed]
- Penders, J.; Stolzoff, M.; Hickey, D.J.; Andersson, M.; Webster, T.J. Shape-dependent antibacterial effects of non- cytotoxic gold nanoparticles. Int. J. Nanomed. 2017, 12, 2457–2468. [Google Scholar] [CrossRef] [Green Version]
- Chlumsky, O.; Purkrtova, S.; Michova, H.; Sykorova, H.; Slepicka, P.; Fajstavr, D.; Ulbrich, P.; Viktorova, J.; Demnerova, K. Antimicrobial properties of palladium and platinum nanoparticles: A new tool for combating food-borne pathogens. Int. J. Mol. Sci. 2021, 22, 7892. [Google Scholar] [CrossRef] [PubMed]
- Gunes, S.; He, Z.; van Acken, D.; Malone, R.; Cullen, P.J.; Curtin, J.F. Platinum nanoparticles inhibit intracellular ROS generation and protect against cold atmospheric plasma-induced cytotoxicity. Nanomed. Nanotechnol. Biol. Med. 2021, 36, 102436. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Cao, Z.; Liu, X.; Cui, Z.; Li, Z.; Liang, Y.; Zhu, S.; Wu, S. Noble metal-based nanomaterials as antibacterial agents. J. Alloys Compd. 2022, 904, 164091. [Google Scholar] [CrossRef]
- Tahir, K.; Nazir, S.; Ahmad, A.; Li, B.; Khan, A.U.; Khan, Z.U.H.; Khan, F.U.; Khan, Q.U.; Khan, A.; Rahman, A.U. Facile and green synthesis of phytochemicals capped platinum nanoparticles and in vitro their superior antibacterial activity. J. Photochem. Photobiol. B Biol. 2017, 166, 246–251. [Google Scholar] [CrossRef]
- Gao, F.; Shao, T.; Yu, Y.; Xiong, Y.; Yang, L. Surface-bound reactive oxygen species generating nanozymes for selective antibacterial action. Nat. Commun. 2021, 12, 745. [Google Scholar] [CrossRef] [PubMed]
- Pedone, D.; Moglianetti, M.; De Luca, E.; Bardi, G.; Pompa, P.P. Platinum nanoparticles in nanobiomedicine. Chem. Soc. Rev. 2017, 46, 4951. [Google Scholar] [CrossRef] [PubMed]
- Fang, G.; Li, W.; Shen, X.; Perez-Aguilar, J.M.; Chong, Y.; Gao, X.; Chai, Z.; Chen, C.; Ge, C.; Zhou, R. Differential Pd-nanocrystal facets demonstrate distinct antibacterial activity against Gram-positive and Gram-negative bacteria. Nat. Commun. 2018, 9, 129. [Google Scholar] [CrossRef] [Green Version]
- Rana, K.; Nayak, S.R.; Bihary, A.; Sahoo, A.K.; Mohanty, K.C.; Palo, S.K.; Sahoo, D.; Pati, S.; Dash, P. Association of quorum sensing and biofilm formation with Salmonella virulence: Story beyond gathering and cross-talk. Arch. Microbiol. 2021, 203, 5887–5897. [Google Scholar] [CrossRef]
- Fu, J.; Qi, L.; Hu, M.; Liu, Y.; Yu, K.; Liu, Q.; Liu, X. Salmonella proteomics under oxidative stress reveals coordinated regulation of antioxidant defense with iron metabolism and bacterial virulence. J. Proteomics 2017, 157, 52–58. [Google Scholar] [CrossRef]
- Rhen, M. Salmonella and Reactive Oxygen Species: A Love-Hate Relationship. J. Innate Immun. 2019, 11, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Hébrard, M.; Viala, J.P.M.; Méresse, S.; Barras, F.; Aussel, L. Redundant hydrogen peroxide scavengers contribute to Salmonella virulence and oxidative stress resistance. J. Bacteriol. 2009, 191, 4605–4614. [Google Scholar] [CrossRef] [Green Version]
- Moglianetti, M.; De Luca, E.; Pedone, D.; Marotta, R.; Catelani, T.; Sartori, B.; Amenitsch, H.; Retta, S.F.; Pompa, P.P. Platinum nanozymes recover cellular ROS homeostasis in an oxidative stress-mediated disease model. Nanoscale 2016, 8, 3739–3752. [Google Scholar] [CrossRef] [PubMed]
- De Luca, E.; Pedone, D.; Moglianetti, M.; Pulcini, D.; Perrelli, A.; Retta, S.F.; Pompa, P.P. Multifunctional Platinum@BSA-Rapamycin Nanocarriers for the Combinatorial Therapy of Cerebral Cavernous Malformation. ACS Omega 2018, 3, 15389–15398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Cui, Y.; Guo, X.; Chen, S.; Sun, H.; Wang, L.; Wang, J.; Zhao, Y.; Liu, Z. Biocompatible bovine serum albumin stabilized platinum nanoparticles for the oxidation of morin. New J. Chem. 2019, 43, 8774–8780. [Google Scholar] [CrossRef]
- Rocchetti, G.; Michelini, S.; Pizzamiglio, V.; Masoero, F.; Lucini, L. A combined metabolomics and peptidomics approach to discriminate anomalous rind inclusion levels in Parmigiano Reggiano PDO grated hard cheese from different ripening stages. Food Res. Int. 2021, 149, 110654. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; Vandergheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]
- Tsugawa, H.; Kind, T.; Nakabayashi, R.; Yukihira, D.; Tanaka, W.; Cajka, T.; Saito, K.; Fiehn, O.; Arita, M. Hydrogen Rearrangement Rules: Computational MS/MS Fragmentation and Structure Elucidation Using MS-FINDER Software. Anal. Chem. 2016, 88, 7946–7958. [Google Scholar] [CrossRef]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The role of reporting standards for metabolite annotation and identification in metabolomic studies. Gigascience 2013, 2, 2047-217X-2-13. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2018, 47, 687–692. [Google Scholar] [CrossRef]
- Zhang, Z.; Huang, R. Analysis of Malondialdehyde, Chlorophyll Proline, Soluble Sugar, and Glutathione Content in Arabidopsis seedling. Bio-Protocol 2013, 3, e817. [Google Scholar] [CrossRef]
- Jin, P.; Niu, X.; Gao, Z.; Xue, X.; Zhang, F.; Cheng, W.; Ren, C.; Du, H.; Manyande, A.; Chen, H. Ultrafine Platinum Nanoparticles Supported on Covalent Organic Frameworks As Stable and Reusable Oxidase-Like Catalysts for Cellular Glutathione Detection. ACS Appl. Nano Mater. 2021, 4, 5834–5841. [Google Scholar] [CrossRef]
- Deng, H.; He, S.; Lin, X.; Yang, L.; Lin, Z.; Chen, R.; Peng, H.; Chen, W. Target-triggered inhibiting oxidase-mimicking activity of platinum nanoparticles for ultrasensitive colorimetric detection of silver ion. Chinese Chem. Lett. 2019, 30, 1659–1662. [Google Scholar] [CrossRef]
- Song, H.; Ma, C.; Wang, L.; Zhu, Z. Platinum Nanoparticles-Deposited Multiwalled Carbon Nanotubes as a NADH Nanoscale characterization and applications. Nanoscale 2021, 12, 19284–19292. [Google Scholar] [CrossRef] [PubMed]
- Spooner, R.; Yilmaz, Ö. The Role of Reactive-Oxygen-Species in Microbial Persistence and Inflammation. Int. J. Mol. Sci. 2011, 12, 334. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hu, C.; Shao, L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017, 12, 1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horst, S.A.; Jaeger, T.; Denkel, L.A.; Rouf, S.F.; Rhen, M.; Bange, F.C. Thiol Peroxidase Protects Salmonella enterica from Hydrogen Peroxide Stress In Vitro and Facilitates Intracellular Growth. J. Bacteriol. 2010, 192, 2929. [Google Scholar] [CrossRef] [Green Version]
- Chlumsky, O.; Purkrtova, S.; Michova, H.; Svarcova, V.; Slepicka, P.; Fajstavr, D.; Ulbrich, P.; Demnerova, K. The effect of gold and silver nanoparticles, chitosan and their combinations on bacterial biofilms of food-borne pathogens. Biofouling 2020, 36, 222–233. [Google Scholar] [CrossRef]
- Hashimoto, M.; Yanagiuchi, H.; Kitagawa, H.; Honda, Y. Inhibitory effect of platinum nanoparticles on biofilm formation of oral bacteria. Nano Biomed. 2018, 9, 77–82. [Google Scholar] [CrossRef]
- Ramkumar, V.S.; Pugazhendhi, A.; Prakash, S.; Ahila, N.K.; Vinoj, G.; Selvam, S.; Kumar, G.; Kannapiran, E.; Rajendran, R.B. ScienceDirect Synthesis of platinum nanoparticles using seaweed Padina gymnospora and their catalytic activity as PVP / PtNPs nanocomposite towards biological applications. Biomed. Pharmacother. 2017, 92, 479–490. [Google Scholar] [CrossRef]
- Ayaz Ahmed, K.B.; Raman, T.; Anbazhagan, V. Platinum nanoparticles inhibit bacteria proliferation and rescue zebrafish from bacterial infection. RSC Adv. 2016, 6, 44415–44424. [Google Scholar] [CrossRef]
- Pawcenis, D.; Twardowska, E.; Leśniak, M.; Jędrzejczyk, R.J.; Sitarz, M.; Profic-Paczkowska, J. TEMPO-oxidized cellulose for in situ synthesis of Pt nanoparticles. Study of catalytic and antimicrobial properties. Int. J. Biol. Macromol. 2022, 213, 738–750. [Google Scholar] [CrossRef]
- Ismail, N.A.S.; Lee, J.X.; Yusof, F. Platinum Nanoparticles: The Potential Antioxidant in the Human Lung Cancer Cells. Antioxidants 2022, 11, 986. [Google Scholar] [CrossRef]
- Nakanishi, H.; Hamasaki, T.; Kinjo, T.; Yan, H.; Nakamichi, N.; Kabayama, S.; Teruya, K.; Shirahata, S. Low concentration platinum nanoparticles effectively scavenge reactive oxygen species in rat skeletal L6 cells. Nano Biomed. Eng. 2013, 5, 76–85. [Google Scholar] [CrossRef] [Green Version]
- Jung, I.L.; Kim, I.G. Transcription of ahpC, katG, and katE genes in Escherichia coli is regulated by polyamines: Polyamine-deficient mutant sensitive to H2O2-induced oxidative damage. Biochem. Biophys. Res. Commun. 2003, 301, 915–922. [Google Scholar] [CrossRef]
- Liu, X.; Omar, M.; Abrahante, J.E.; Nagaraja, K.V.; Vidovic, S. Insights into the oxidative stress response of salmonella enterica serovar enteritidis revealed by the next generation sequencing approach. Antioxidants 2020, 9, 849. [Google Scholar] [CrossRef] [PubMed]
- Piao, M.J.; Kang, K.A.; Lee, I.K.; Kim, H.S.; Kim, S.; Choi, J.Y.; Choi, J.; Hyun, J.W. Silver nanoparticles induce oxidative cell damage in human liver cells through inhibition of reduced glutathione and induction of mitochondria-involved apoptosis. Toxicol. Lett. 2011, 201, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Pujalté, I.; Passagne, I.; Brouillaud, B.; Tréguer, M.; Durand, E.; Ohayon-Courtès, C.; L’Azou, B. Cytotoxicity and oxidative stress induced by different metallic nanoparticles on human kidney cells. Part. Fibre Toxicol. 2011, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Huang, X.; Sun, W.; Too, H.Z.; Laserna, A.K.C.; Li, S.F.Y. A global metabolomic insight into the oxidative stress and membrane damage of copper oxide nanoparticles and microparticles on microalga Chlorella vulgaris. Environ. Pollut. 2020, 258, 113647. [Google Scholar] [CrossRef]
- Fanoro, O.T.; Parani, S.; Maluleke, R.; Lebepe, T.C.; Varghese, R.J.; Mgedle, N.; Mavumengwana, V.; Oluwafemi, O.S. Biosynthesis of smaller-sized platinum nanoparticles using the leaf extract of combretum erythrophyllum and its antibacterial activities. Antibiotics 2021, 10, 1275. [Google Scholar] [CrossRef]
- Seixas, A.F.; Quendera, A.P.; Sousa, J.P.; Silva, A.F.Q.; Arraiano, C.M.; Andrade, J.M. Bacterial Response to Oxidative Stress and RNA Oxidation. Front. Genet. 2022, 12, 2789. [Google Scholar] [CrossRef]
- Sawosz, E.; Chwalibog, A.; Szeliga, J.; Sawosz, F.; Grodzik, M.; Rupiewicz, M.; Niemiec, T.; Kacprzyk, K. Visualization of gold and platinum nanoparticles interacting with Salmonella Enteritidis and Listeria monocytogenes. Int. J. Nanomed. 2010, 5, 631. [Google Scholar] [CrossRef] [Green Version]
- Bogomolnaya, L.M.; Andrews, K.D.; Talamantes, M.; Maple, A.; Ragoza, Y.; Vazquez-Torres, A.; Andrews-Polymenis, H. The ABC-Type Efflux Pump MacAB Protects Salmonella enterica serovar Typhimurium from Oxidative Stress. MBio 2013, 4, e00630-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redfern, B.H. The Incorporation of Lipids into the Cellular Membrane of Salmonella. Bachelor’s Thesis, University of Southern Mississippi, Hattiesburg, MS, USA, May 2019. [Google Scholar]
- Wang, R.Y.; Ou, Y.; Zeng, X.A.; Guo, C.J. Membrane fatty acids composition and fluidity modification in Salmonella Typhimurium by culture temperature and resistance under pulsed electric fields. Int. J. Food Sci. Technol. 2019, 54, 2236–2245. [Google Scholar] [CrossRef]
- Chanana, V.; Majumdar, S.; Rishi, P. Involvement of caspase-3, lipid peroxidation and TNF-α in causing apoptosis of macrophages by coordinately expressed Salmonella phenotype under stress conditions. Mol. Immunol. 2007, 44, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Planchon, M.; Léger, T.; Spalla, O.; Huber, G.; Ferrari, R. Metabolomic and proteomic investigations of impacts of titanium dioxide nanoparticles on Escherichia coli. PLoS ONE 2017, 12, e0178437. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belloso Daza, M.V.; Scarsi, A.; Gatto, F.; Rocchetti, G.; Pompa, P.P.; Cocconcelli, P.S. Role of Platinum Nanozymes in the Oxidative Stress Response of Salmonella Typhimurium. Antioxidants 2023, 12, 1029. https://doi.org/10.3390/antiox12051029
Belloso Daza MV, Scarsi A, Gatto F, Rocchetti G, Pompa PP, Cocconcelli PS. Role of Platinum Nanozymes in the Oxidative Stress Response of Salmonella Typhimurium. Antioxidants. 2023; 12(5):1029. https://doi.org/10.3390/antiox12051029
Chicago/Turabian StyleBelloso Daza, Mireya Viviana, Anna Scarsi, Francesca Gatto, Gabriele Rocchetti, Pier Paolo Pompa, and Pier Sandro Cocconcelli. 2023. "Role of Platinum Nanozymes in the Oxidative Stress Response of Salmonella Typhimurium" Antioxidants 12, no. 5: 1029. https://doi.org/10.3390/antiox12051029