
Melanoma Cellular Signaling Transduction Pathways Targeted by Polyphenols Action Mechanisms

 ,
,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
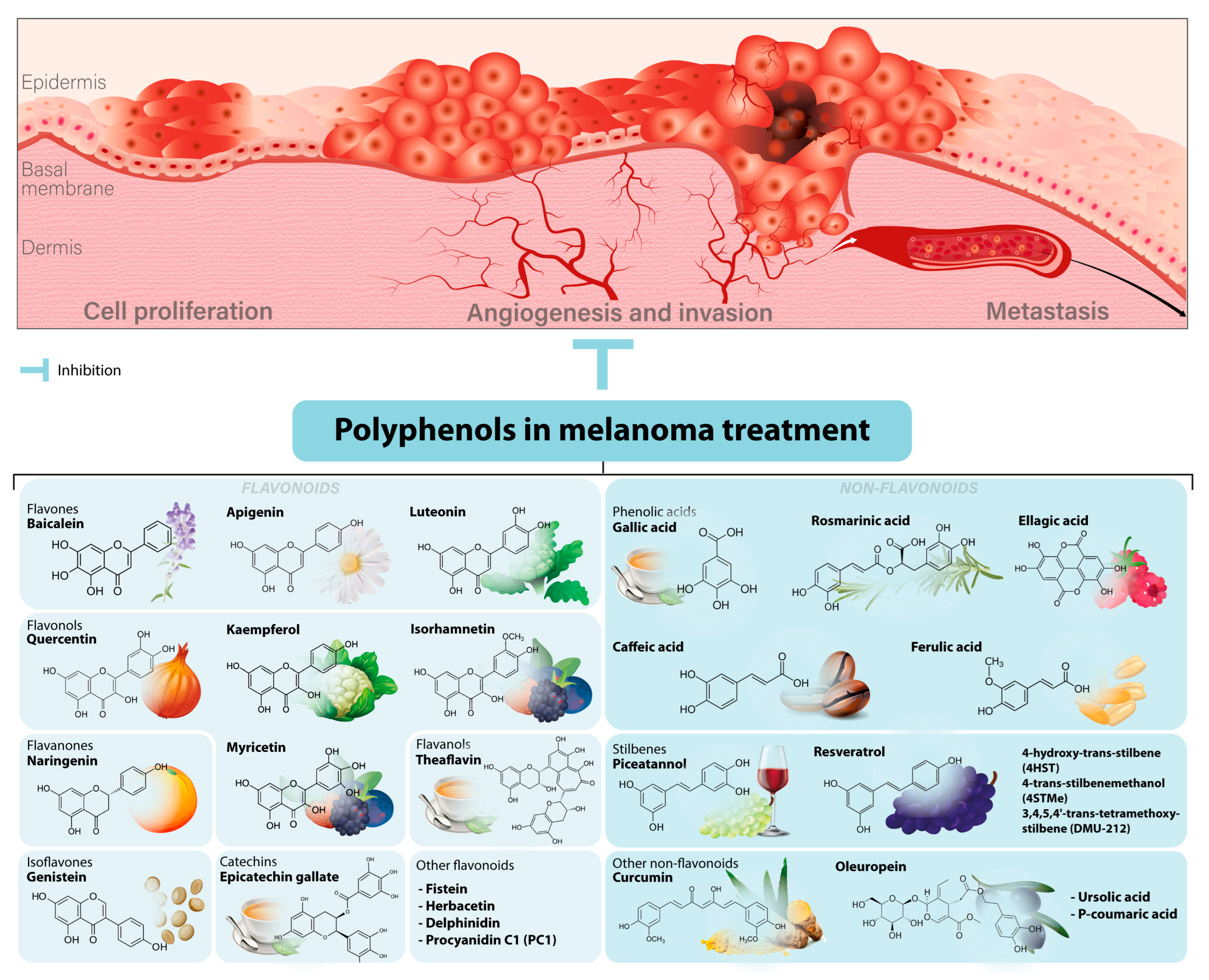
2. Effects of Polyphenols on Melanoma Initiation
2.1. Inhibition of Cell Growth
2.1.1. Cell Cycle Arrest
2.1.2. Antiproliferative Effect of MAPK Pathway Regulation
2.2. Apoptosis
2.2.1. p53-Dependent Apoptosis
2.2.2. p53-Independent Apoptosis
2.2.3. Oxidative Stress in Apoptotic Pathway Regulation
2.3. Autophagy
Regulation of AKT/PKB Pathway
3. Effects of Polyphenols on Tumor Progression in Melanoma
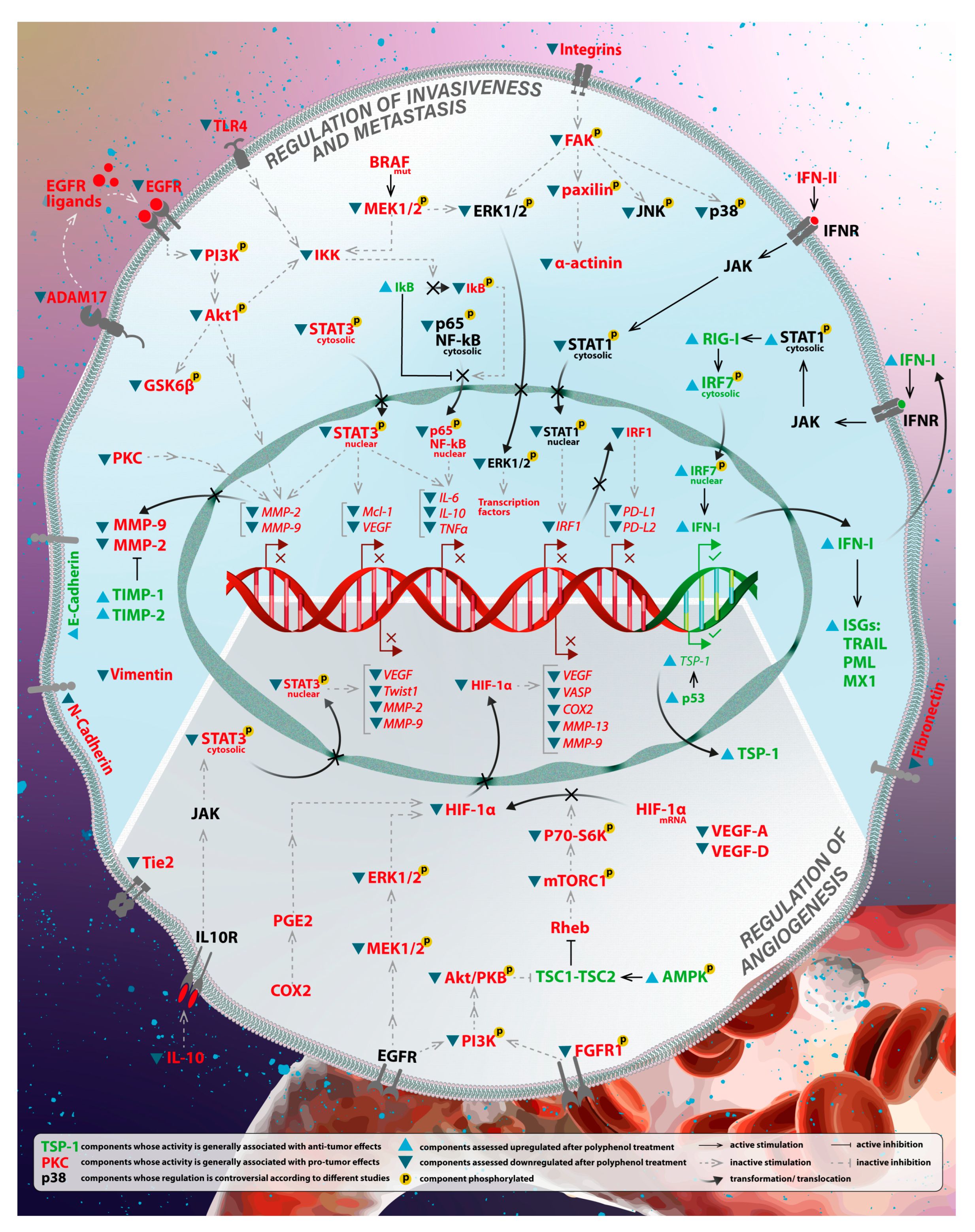
3.1. Regulation of Invasiveness and Metastasis
3.1.1. Modulation of the Immune System
3.1.2. Downregulation of Matrix Metalloproteinases
3.2. Regulation of Angiogenesis
4. The Relationship of Polyphenols and miRNAs in Melanoma
5. Clinical Implications of Polyphenols and Combinatorial Therapy Approach
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 4EBP1 | Eukaryotic translation initiation factor 4E binding protein 1 |
| ADAM17 | ADAM metallopeptidase domain 17 |
| AKT (PKB) | Protein kinase B |
| AMPK | AMP-activated protein kinase |
| ANG1/2 | Angiopoietin 1/2 |
| ANT-1 | Adenine nucleotide translocator-1 |
| ATM | Ataxia-telangiectasia mutated kinase |
| BAX | BCL-2-associated X |
| BCL-2 | B-cell lymphoma 2 apoptosis regulator |
| BCL2A1 | BCL-2-related protein A1 |
| bFGF | Basic fibroblast growth factor |
| BID | BH3 interacting domain death agonist |
| BIRC5 (survivin) | Baculoviral IAP repeat containing 5 |
| BRAF | BRAF proto-oncogene |
| CASP | Caspase |
| CD147 (BSG) | Basigin |
| CDK | Cyclin-dependent kinase |
| CDKN1A (p21) | Cyclin-dependent kinase inhibitor 1A |
| CDKN1B (p27) | Cyclin-dependent kinase inhibitor 1B |
| CHK1 (CHEK1) | Checkpoint kinase 1 |
| CHK2 (CHEK2) | Checkpoint kinase 2 |
| CHOP | C/EBP homologous protein |
| c-KIT (KIT) | Tyrosine-protein kinase KIT |
| COX-2 | Cyclooxygenase 2 |
| CREB | cAMP-response element binding protein |
| CSC | Cancer stem cells |
| CyPD | Cyclophilin-D |
| DDR | DNA damage response |
| DISC | Death-inducing signaling complex |
| DVL-2 | Disheveled-2 |
| ECM | Extracellular matrix |
| EF24 | Diphenyl difluoroketone |
| EGCG | Epigallocatechin-3-gallate |
| EGFR | Epidermal growth factor receptor |
| eIF2α | Eukaryotic initiation factor 2 alpha |
| EMT | Epithelial–mesenchymal transition |
| ER | Endoplasmic reticulum |
| ERK | Extracellular signal-regulated kinase |
| EZH2 | Enhancer of zeste 2 polycomb repressive complex 2 subunit |
| FAK | Focal adhesion kinase |
| Fas | Fas cell surface death receptor |
| FasL | Fas ligand |
| FGFR1 | Fibroblast growth factor receptor |
| GSK3β | Glycogen synthase kinase 3 beta |
| GTPs | Green tea polyphenols |
| HDAC | Histone deacetylase |
| HIF-1α | Hypoxia-inducible factor 1 alpha |
| HMGA2 | High-mobility group AT-hook 2 |
| IFN | Interferon |
| IGF-1 | Insulin-like growth factor 1 |
| IHC | Immunohistochemistry |
| IkB | Inhibitor of nuclear factor kappa B |
| IL | Interleukin |
| IRF7 | Interferon regulatory factor 7 |
| JAK1 | Janus kinase 1 |
| JNK | JUN N-terminal kinase |
| KRAS | Kirsten rat sarcoma virus protein |
| LC3 (MAP1LC3A) | Microtubule-associated protein 1 light chain 3 alpha |
| MAPK | Mitogen-activated protein kinase |
| MCL-1 | Myeloid cell leukemia 1 apoptosis regulator |
| MEK (MAP2K) | Mitogen-activated protein kinase kinase |
| miRNA (miR) | microRNA |
| MITF | Microphthalmia-associated transcription factor |
| MMP | Matrix metalloproteinases |
| mPTP | Mitochondrial permeability transition pore |
| mTORC | Mechanistic target of rapamycin kinase |
| ncRNA | Non-coding RNA |
| NF-κb | Nuclear factor kappa B |
| NQO2 | Quinone reductase 2 |
| NRF2 | Nuclear factor erythroid 2-related factor 2 |
| P70S6K (S6K) | Ribosomal protein S6 kinase |
| PARP | Poly(ADP-ribose) polymerase 1 |
| PCNA | Proliferating cell nuclear antigen |
| PDCD4 | Programmed cell death 4 |
| PDL-1 | Programmed death ligand 1 |
| PDL-2 | Programmed death ligand 2 |
| PEBP | Polyphenol-enriched blueberry preparation |
| PFKFB4 | Phosphofructo-2-kinase/fructose-2,6-bisphosphatases 4 |
| PI3K | Phosphoinositide 3-kinase |
| PIK3R3 | Phosphoinositide-3-kinase regulatory subunit 3 |
| PKC | Protein kinase C |
| PKM2 | Pyruvate kinase M2 |
| pRB | Protein retinoblastoma |
| PTEN | Phosphatase and tensin homolog |
| RIG-I | Retinoic acid-inducible gene I protein |
| ROS | Reactive oxygen species |
| SAPK | Stress-activated protein kinase |
| siRNA | Small interfering RNA |
| SOX-10 | SRY-Box transcription factor 10 |
| STAT1 | Signal transducer and activator of transcription 1 |
| STAT3 | Signal transducer and activator of transcription 3 |
| TFG | TRK-fused gene protein |
| TGF | Transforming growth factor |
| TIMP | Tissue inhibitors of metalloproteinases |
| TLR4 | Toll-like receptor 4 |
| TME | Tumor microenvironment |
| TNF-α | Tumor necrosis factor alpha |
| TRADD | Tumor necrosis factor receptor-1-associated protein |
| TSP-1 | Thrombospondin-1 |
| TWIST1 | Twist family BHLH transcription factor 1 |
| Tyr | Tyrosinase |
| ULK1 | Unc-51-like autophagy activating kinase 1 |
| UTR | Untranslated region |
| UV-B | Ultraviolet B light |
| VASP | Vasodilator-stimulated phosphoprotein |
| VEGF | Vascular endothelial growth factor |
| Wnt | Wingless-Int |
| WT | Wild type |
| TKO | TSP1 knockout |
| XIAP | X-linked inhibitor of apoptosis protein |
| ZEB1 | Zinc finger E-box binding homeobox 1 |
References
- Kontogianni, G.; Piroti, G.; Maglogiannis, I.; Chatziioannou, A.; Papadodima, O. Dissecting the Mutational Landscape of Cutaneous Melanoma: An Omic Analysis Based on Patients from Greece. Cancers 2018, 10, 96. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Rabbie, R.; Ferguson, P.; Molina-Aguilar, C.; Adams, D.J.; Robles-Espinoza, C.D. Melanoma Subtypes: Genomic Profiles, Prognostic Molecular Markers and Therapeutic Possibilities. J. Pathol. 2019, 247, 539–551. [Google Scholar] [CrossRef] [PubMed]
- Dzwierzynski, W.W. Managing Malignant Melanoma. Plast. Reconstr. Surg. 2013, 132, 446e–460e. [Google Scholar] [CrossRef]
- Mishra, H.; Mishra, P.K.; Ekielski, A.; Jaggi, M.; Iqbal, Z.; Talegaonkar, S. Melanoma Treatment: From Conventional to Nanotechnology. J. Cancer Res. Clin. Oncol. 2018, 144, 2283–2302. [Google Scholar] [CrossRef]
- Gamba, C.S.; Clarke, C.A.; Keegan, T.H.M.; Tao, L.; Swetter, S.M. Melanoma Survival Disadvantage in Young, Non-Hispanic White Males Compared with Females. JAMA Dermatol. 2013, 149, 912–920. [Google Scholar] [CrossRef]
- Bochis, O.V.; Irimie, A.; Pichler, M.; Berindan-Neagoe, I. The Role of Skp2 and Its Substrate CDKN1B (P27) in Colorectal Cancer. J. Gastrointestin. Liver Dis. 2015, 24, 225–234. [Google Scholar] [CrossRef]
- Braicu, C.; Pileczki, V.; Irimie, A.; Berindan-Neagoe, I. P53siRNA Therapy Reduces Cell Proliferation, Migration and Induces Apoptosis in Triple Negative Breast Cancer Cells. Mol. Cell. Biochem. 2013, 381, 61–68. [Google Scholar] [CrossRef]
- Grozav, A.; Balacescu, O.; Balacescu, L.; Cheminel, T.; Berindan-Neagoe, I.; Therrien, B. Synthesis, Anticancer Activity, and Genome Profiling of Thiazolo Arene Ruthenium Complexes. J. Med. Chem. 2015, 58, 8475–8490. [Google Scholar] [CrossRef]
- Chiorean, R.; Braicu, C.; Berindan-Neagoe, I. Another Review on Triple Negative Breast Cancer. Are We on the Right Way towards the Exit from the Labyrinth? Breast 2013, 22, 1026–1033. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, H.G. Anti-Cancer Drug Discovery and Development: Bcl-2 Family Small Molecule Inhibitors. Commun. Integr. Biol. 2012, 5, 557. [Google Scholar] [CrossRef] [PubMed]
- Kozovska, Z.; Gabrisova, V.; Kucerova, L. Malignant Melanoma: Diagnosis, Treatment and Cancer Stem Cells. Neoplasma 2016, 63, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Chanda, M.; Cohen, M.S. Advances in the Discovery and Development of Melanoma Drug Therapies. Expert Opin. Drug Discov. 2021, 16, 1319–1347. [Google Scholar] [CrossRef]
- Bentebibel, S.E.; Diab, A. Cytokines in the Treatment of Melanoma. Curr. Oncol. Rep. 2021, 23, 83. [Google Scholar] [CrossRef] [PubMed]
- Carlino, M.S.; Larkin, J.; Long, G.V. Immune Checkpoint Inhibitors in Melanoma. Lancet 2021, 398, 1002–1014. [Google Scholar] [CrossRef]
- Mirzavi, F.; Barati, M.; Soleimani, A.; Vakili-Ghartavol, R.; Jaafari, M.R.; Soukhtanloo, M. A Review on Liposome-Based Therapeutic Approaches against Malignant Melanoma. Int. J. Pharm. 2021, 599, 120413. [Google Scholar] [CrossRef]
- Cassano, R.; Cuconato, M.; Calviello, G.; Serini, S.; Trombino, S. Recent Advances in Nanotechnology for the Treatment of Melanoma. Molecules 2021, 26, 785. [Google Scholar] [CrossRef]
- Cheng, Y.C.; Chang, Y.A.; Chen, Y.J.; Sung, H.M.; Bogeski, I.; Su, H.L.; Hsu, Y.L.; Wang, H.M.D. The Roles of Extracellular Vesicles in Malignant Melanoma. Cells 2021, 10, 2740. [Google Scholar] [CrossRef]
- Trager, M.H.; Geskin, L.J.; Saenger, Y.M. Oncolytic Viruses for the Treatment of Metastatic Melanoma. Curr. Treat. Options Oncol. 2020, 21, 26. [Google Scholar] [CrossRef]
- Demirsoy, S.; Martin, S.; Maes, H.; Agostinis, P. Adapt, Recycle, and Move on: Proteostasis and Trafficking Mechanisms in Melanoma. Front. Oncol. 2016, 6, 240. [Google Scholar] [CrossRef] [Green Version]
- Garbe, C.; Peris, K.; Hauschild, A.; Saiag, P.; Middleton, M.; Bastholt, L.; Grob, J.J.; Malvehy, J.; Newton-Bishop, J.; Stratigos, A.J.; et al. Diagnosis and Treatment of Melanoma. European Consensus-Based Interdisciplinary Guideline—Update 2016. Eur. J. Cancer 2016, 63, 201–217. [Google Scholar] [CrossRef]
- Bommareddy, P.K.; Silk, A.W.; Kaufman, H.L. Intratumoral Approaches for the Treatment of Melanoma. Cancer J. 2017, 23, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Zanoaga, O.; Jurj, A.; Raduly, L.; Cojocneanu-Petric, R.; Fuentes-Mattei, E.; Wu, O.; Braicu, C.; Gherman, C.D.; Berindan-Neagoe, I. Implications of Dietary ω-3 and ω-6 Polyunsaturated Fatty Acids in Breast Cancer. Exp. Ther. Med. 2018, 15, 1167. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, Y.-F. Natural Compounds as Anticancer Agents: Experimental Evidence. World J. Exp. Med. 2012, 2, 45. [Google Scholar] [CrossRef]
- Khan, H.; Nabavi, S.M.; Sureda, A.; Mehterov, N.; Gulei, D.; Berindan-Neagoe, I.; Taniguchi, H.; Atanasov, A.G. Therapeutic Potential of Songorine, a Diterpenoid Alkaloid of the Genus Aconitum. Eur. J. Med. Chem. 2018, 153, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [PubMed]
- Chinembiri, T.N.; Du Plessis, L.H.; Gerber, M.; Hamman, J.H.; Du Plessis, J. Review of Natural Compounds for Potential Skin Cancer Treatment. Molecules 2014, 19, 11679–11721. [Google Scholar] [CrossRef]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural Polyphenols: Chemical Classification, Definition of Classes, Subcategories, and Structures. J. AOAC Int. 2019, 102, 1397–1400. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhou, J. The Application of Polyphenols in Food Preservation. Adv. Food Nutr. Res. 2021, 98, 35–99. [Google Scholar]
- Almeida, T.C.; da Silva, G.N.; de Souza, D.V.; de Moraes Malinverni, A.C.; Aguiar, O.; Estadella, D.; Ribeiro, D.A. Resveratrol Effects in Oral Cancer Cells: A Comprehensive Review. Med. Oncol. 2021, 38, 97. [Google Scholar] [CrossRef]
- Behroozaghdam, M.; Dehghani, M.; Zabolian, A.; Kamali, D.; Javanshir, S.; Hasani Sadi, F.; Hashemi, M.; Tabari, T.; Rashidi, M.; Mirzaei, S.; et al. Resveratrol in Breast Cancer Treatment: From Cellular Effects to Molecular Mechanisms of Action. Cell. Mol. Life Sci. 2022, 79, 539. [Google Scholar] [CrossRef]
- Vernousfaderani, E.K.; Akhtari, N.; Rezaei, S.; Rezaee, Y.; Shiranirad, S.; Mashhadi, M.; Hashemi, A.; Khankandi, H.P.; Behzad, S. Resveratrol and Colorectal Cancer: A Molecular Approach to Clinical Researches. Curr. Top. Med. Chem. 2021, 21, 2634–2646. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Zhou, M.; Huang, D.; Wasan, H.S.; Zhang, K.; Sun, L.; Huang, H.; Ma, S.; Shen, M.; Ruan, S. Resveratrol Inhibits the Invasion and Metastasis of Colon Cancer through Reversal of Epithelial- Mesenchymal Transition via the AKT/GSK-3β/Snail Signaling Pathway. Mol. Med. Rep. 2019, 20, 2783–2795. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, L.; Zhu, F.; Han, X.; Sun, L.; Chen, K. The Cytotoxicity Effect of Resveratrol: Cell Cycle Arrest and Induced Apoptosis of Breast Cancer 4T1 Cells. Toxins 2019, 11, 731. [Google Scholar] [CrossRef]
- Gong, C.; Xia, H. Resveratrol Suppresses Melanoma Growth by Promoting Autophagy through Inhibiting the PI3K/AKT/MTOR Signaling Pathway. Exp. Ther. Med. 2020, 19, 1878–1886. [Google Scholar] [CrossRef] [PubMed]
- Silk, N.; Reich, J.; Sinha, R.; Chawla, S.; Geary, K.; Zhang, D. The Effects of Resveratrol on Prostate Cancer through Targeting the Tumor Microenvironment. J. Xenobiotics 2021, 11, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Vladu, A.F.; Ficai, D.; Ene, A.G.; Ficai, A. Combination Therapy Using Polyphenols: An Efficient Way to Improve Antitumoral Activity and Reduce Resistance. Int. J. Mol. Sci. 2022, 23, 10244. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (Poly)Phenolics in Human Health: Structures, Bioavailability, and Evidence of Protective Effects against Chronic Diseases. Antioxid. Redox Signal 2013, 18, 1818–1892. [Google Scholar] [CrossRef]
- Cháirez-Ramírez, M.H.; de la Cruz-López, K.G.; García-Carrancá, A. Polyphenols as Antitumor Agents Targeting Key Players in Cancer-Driving Signaling Pathways. Front. Pharmacol. 2021, 12, 710304. [Google Scholar] [CrossRef]
- Liu, Z.; Hu, M. Natural Polyphenol Disposition via Coupled Metabolic Pathways. Expert Opin. Drug Metab. Toxicol. 2007, 3, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P.E. Polyphenols and Human Health: Prevention of Disease and Mechanisms of Action. Nutrients 2010, 2, 1106–1131. [Google Scholar] [CrossRef] [PubMed]
- Durazzo, A.; Lucarini, M.; Camilli, E.; Marconi, S.; Gabrielli, P.; Lisciani, S.; Gambelli, L.; Aguzzi, A.; Novellino, E.; Santini, A.; et al. Dietary Lignans: Definition, Description and Research Trends in Databases Development. Molecules 2018, 23, 2351. [Google Scholar] [CrossRef] [PubMed]
- Lecour, S.; Lamont, K.T. Natural Polyphenols and Cardioprotection. Mini Rev. Med. Chem. 2011, 11, 1191–1199. [Google Scholar]
- Serino, A.; Salazar, G. Protective Role of Polyphenols against Vascular Inflammation, Aging and Cardiovascular Disease. Nutrients 2018, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kaur, M.; Katnoria, J.K.; Nagpal, A.K. Polyphenols in Food: Cancer Prevention and Apoptosis Induction. Curr. Med. Chem. 2018, 25, 4740–4757. [Google Scholar] [CrossRef]
- Kang, S.; Wang, Z.; Li, B.; Gao, X.; He, W.; Cao, S.; Cai, Y.; Chen, H. Anti-Tumor Effects of Resveratrol on Malignant Melanoma Is Associated with Promoter Demethylation of RUNX3 Gene. Pharmazie 2019, 74, 163–167. [Google Scholar]
- Tabrez, S.; Jabir, N.R.; Adhami, V.M.; Khan, M.I.; Moulay, M.; Kamal, M.A.; Mukhtar, H. Nanoencapsulated Dietary Polyphenols for Cancer Prevention and Treatment: Successes and Challenges. Nanomedicine 2020, 15, 1147–1162. [Google Scholar] [CrossRef]
- Morris, G.; Gamage, E.; Travica, N.; Berk, M.; Jacka, F.N.; O’Neil, A.; Puri, B.K.; Carvalho, A.F.; Bortolasci, C.C.; Walder, K.; et al. Polyphenols as Adjunctive Treatments in Psychiatric and Neurodegenerative Disorders: Efficacy, Mechanisms of Action, and Factors Influencing Inter-Individual Response. Free Radic. Biol. Med. 2021, 172, 101–122. [Google Scholar]
- Stamatakos, M.; Palla, V.; Karaiskos, I.; Xiromeritis, K.; Alexiou, I.; Pateras, I.; Kontzoglou, K. Cell Cyclins: Triggering Elements of Cancer or Not? World, J. Surg. Oncol. 2010, 8, 111. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, B.; Cailing, E.; Liu, J.; Zhang, Q.; Liu, J.; Chen, N.; Chen, R.; Zhu, R. Resveratrol Inhibits the Proliferation of Human Melanoma Cells by Inducing G1/S Cell Cycle Arrest and Apoptosis. Mol. Med. Rep. 2015, 11, 400–404. [Google Scholar] [CrossRef]
- Gatouillat, G.; Balasse, E.; Joseph-Pietras, D.; Morjani, H.; Madoulet, C. Resveratrol Induces Cell-Cycle Disruption and Apoptosis in Chemoresistant B16 Melanoma. J. Cell. Biochem. 2010, 110, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Xiang, W.; Wang, F.F.; Wang, R.; Ding, Y. Curcumin Inhibited Growth of Human Melanoma A375 Cells via Inciting Oxidative Stress. Biomed. Pharmacother. 2017, 95, 1177–1186. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Han, X.; Zheng, S.; Li, Z.; Sha, Y.; Ni, J.; Sun, Z.; Qiao, S.; Song, Z. Curcumin Induces Autophagy, Inhibits Proliferation and Invasion by Downregulating AKT/MTOR Signaling Pathway in Human Melanoma Cells. Oncol. Rep. 2016, 35, 1065–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Androutsopoulos, V.P.; Fragiadaki, I.; Tosca, A. Activation of ERK1/2 Is Required for the Antimitotic Activity of the Resveratrol Analogue 3,4,5,4’-Tetramethoxystilbene (DMU-212) in Human Melanoma Cells. Exp. Dermatol. 2015, 24, 632–634. [Google Scholar] [CrossRef]
- Heo, J.R.; Kim, S.M.; Hwang, K.A.; Kang, J.H.; Choi, K.C. Resveratrol Induced Reactive Oxygen Species and Endoplasmic Reticulum Stress-mediated Apoptosis, and Cell Cycle Arrest in the A375SM Malignant Melanoma Cell Line. Int. J. Mol. Med. 2018, 42, 1427–1435. [Google Scholar] [CrossRef]
- Alaswad, H.A.; Mahbub, A.A.; Le Maitre, C.L.; Jordan-mahy, N. Molecular Action of Polyphenols in Leukaemia and Their Therapeutic Potential. Int. J. Mol. Sci. 2021, 22, 3085. [Google Scholar] [CrossRef]
- Patra, S.; Pradhan, B.; Nayak, R.; Behera, C.; Das, S.; Patra, S.K.; Efferth, T.; Jena, M.; Bhutia, S.K. Dietary Polyphenols in Chemoprevention and Synergistic Effect in Cancer: Clinical Evidences and Molecular Mechanisms of Action. Phytomedicine 2021, 90, 153554. [Google Scholar] [CrossRef]
- Burotto, M.; Chiou, V.L.; Lee, J.M.; Kohn, E.C. The MAPK Pathway across Different Malignancies: A New Perspective. Cancer 2014, 120, 3446–3456. [Google Scholar] [CrossRef]
- Kim, S.H.; Yoo, E.S.; Woo, J.S.; Han, S.H.; Lee, J.H.; Jung, S.H.; Kim, H.J.; Jung, J.Y. Antitumor and Apoptotic Effects of Quercetin on Human Melanoma Cells Involving JNK/P38 MAPK Signaling Activation. Eur. J. Pharmacol. 2019, 860, 172568. [Google Scholar] [CrossRef]
- Androutsopoulos, V.P.; Fragiadaki, I.; Spandidos, D.A.; Tosca, A. The Resveratrol Analogue, 3,4,5,4’-trans-Tetramethoxystilbene, Inhibits the Growth of A375 Melanoma Cells through Multiple Anticancer Modes of Action. Int. J. Oncol. 2016, 49, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yan, B.; Meng, S.; Zhou, L.; Xu, Y.; Du, W.; Shan, L. Theaflavin Induces Apoptosis of A375 Human Melanoma Cells and Inhibits Tumor Growth in Xenograft Zebrafishes through P53- and JNK-Related Mechanism. Front. Pharmacol. 2020, 11, 1317. [Google Scholar] [CrossRef] [PubMed]
- Jiang, A.J.; Jiang, G.; Li, L.T.; Zheng, J.N. Curcumin Induces Apoptosis through Mitochondrial Pathway and Caspases Activation in Human Melanoma Cells. Mol. Biol. Rep. 2015, 42, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Ekmekcioglu, S.; Walch, E.T.; Tang, C.H.; Grimm, E.A. Inhibition of Nuclear Factor-KappaB and Nitric Oxide by Curcumin Induces G2/M Cell Cycle Arrest and Apoptosis in Human Melanoma Cells. Melanoma Res. 2004, 14, 165–171. [Google Scholar] [CrossRef]
- Marín, Y.E.; Wall, B.A.; Wang, S.; Namkoong, J.; Martino, J.J.; Suh, J.; Lee, H.J.; Rabson, A.B.; Yang, C.S.; Chen, S.; et al. Curcumin Downregulates the Constitutive Activity of NF-KappaB and Induces Apoptosis in Novel Mouse Melanoma Cells. Melanoma Res. 2007, 17, 274–283. [Google Scholar] [CrossRef]
- Yang, H.Z.; Zhang, J.; Zeng, J.; Liu, S.; Zhou, F.; Zhang, F.; Giampieri, F.; Cianciosi, D.; Forbes-Hernandez, T.Y.; Ansary, J.; et al. Resveratrol Inhibits the Proliferation of Melanoma Cells by Modulating Cell Cycle. Int. J. Food Sci. Nutr. 2020, 71, 84–93. [Google Scholar] [CrossRef]
- Fulda, S.; Debatin, K.M. Extrinsic versus Intrinsic Apoptosis Pathways in Anticancer Chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar] [CrossRef]
- Chiu, C.T.; Hsuan, S.W.; Lin, H.H.; Hsu, C.C.; Chou, F.P.; Chen, J.H. Hibiscus Sabdariffa Leaf Polyphenolic Extract Induces Human Melanoma Cell Death, Apoptosis, and Autophagy. J. Food Sci. 2015, 80, H649–H658. [Google Scholar] [CrossRef]
- Pourhanifeh, M.H.; Abbaszadeh-Goudarzi, K.; Goodarzi, M.; Piccirillo, S.G.M.; Shafiee, A.; Hajighadimi, S.; Moradizarmehri, S.; Asemi, Z.; Mirzaei, H. Resveratrol: A New Potential Therapeutic Agent for Melanoma? Curr. Med. Chem. 2021, 28, 687–711. [Google Scholar] [CrossRef]
- Zhao, H.; Han, L.; Jian, Y.; Ma, Y.; Yan, W.; Chen, X.; Xu, H.; Li, L. Resveratrol Induces Apoptosis in Human Melanoma Cell through Negatively Regulating Erk/PKM2/Bcl-2 Axis. Onco. Targets. Ther. 2018, 11, 8995–9006. [Google Scholar] [CrossRef]
- Bian, Y.; Wei, J.; Zhao, C.; Li, G. Natural Polyphenols Targeting Senescence: A Novel Prevention and Therapy Strategy for Cancer. Int. J. Mol. Sci. 2020, 21, 684. [Google Scholar] [CrossRef] [PubMed]
- Pandima Devi, K.; Rajavel, T.; Daglia, M.; Nabavi, S.F.; Bishayee, A.; Nabavi, S.M. Targeting MiRNAs by Polyphenols: Novel Therapeutic Strategy for Cancer. Semin. Cancer Biol. 2017, 46, 146–157. [Google Scholar] [CrossRef]
- Heenatigala Palliyage, G.; Singh, S.; Ashby, C.R.; Tiwari, A.K.; Chauhan, H. Pharmaceutical Topical Delivery of Poorly Soluble Polyphenols: Potential Role in Prevention and Treatment of Melanoma. AAPS PharmSciTech 2019, 20, 250. [Google Scholar] [CrossRef] [PubMed]
- Habibie; Yokoyama, S.; Abdelhamed, S.; Awale, S.; Sakurai, H.; Hayakawa, Y.; Saiki, I. Survivin Suppression through STAT3/β-Catenin Is Essential for Resveratrol-Induced Melanoma Apoptosis. Int. J. Oncol. 2014, 45, 895–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free Radicals, Metals and Antioxidants in Oxidative Stress-Induced Cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Oxidative Stress and Cancer: Have We Moved Forward? Biochem. J. 2007, 401, 1–11. [Google Scholar] [CrossRef]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of Oxidative Stress as an Anticancer Strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar] [CrossRef]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in Cancer Therapy: The Bright Side of the Moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef]
- Glasauer, A.; Chandel, N.S. Targeting Antioxidants for Cancer Therapy. Biochem. Pharmacol. 2014, 92, 90–101. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef]
- Russo, G.L.; Tedesco, I.; Spagnuolo, C.; Russo, M. Antioxidant Polyphenols in Cancer Treatment: Friend, Foe or Foil? Semin. Cancer Biol. 2017, 46, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, Y.; Ohishi, T.; Nakamura, Y.; Fukutomi, R.; Miyoshi, N. Anti-Cancer Effects of Dietary Polyphenols via ROS-Mediated Pathway with Their Modulation of MicroRNAs. Molecules 2022, 27, 3816. [Google Scholar] [CrossRef]
- Ouyang, J.; Zhu, K.; Liu, Z.; Huang, J. Prooxidant Effects of Epigallocatechin-3-Gallate in Health Benefits and Potential Adverse Effect. Oxid. Med. Cell. Longev. 2020, 2020, 9723686. [Google Scholar] [CrossRef]
- Nakamae, I.; Morimoto, T.; Shima, H.; Shionyu, M.; Fujiki, H.; Yoneda-Kato, N.; Yokoyama, T.; Kanaya, S.; Kakiuchi, K.; Shirai, T.; et al. Curcumin Derivatives Verify the Essentiality of ROS Upregulation in Tumor Suppression. Molecules 2019, 24, 4067. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.T.; Lee, S.H.; Kim, J., II; Kim, Y.M. Quercetin Regulates the Sestrin 2-AMPK-P38 MAPK Signaling Pathway and Induces Apoptosis by Increasing the Generation of Intracellular ROS in a P53-Independent Manner. Int. J. Mol. Med. 2014, 33, 863–869. [Google Scholar] [CrossRef]
- Kocyigit, A.; Guler, E.M. Curcumin Induce DNA Damage and Apoptosis through Generation of Reactive Oxygen Species and Reducing Mitochondrial Membrane Potential in Melanoma Cancer Cells. Cell. Mol. Biol. 2017, 63, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Yu, T.; Wang, W.; Pan, K.; Shi, D.; Sun, H. Curcumin-Induced Melanoma Cell Death Is Associated with Mitochondrial Permeability Transition Pore (MPTP) Opening. Biochem. Biophys. Res. Commun. 2014, 448, 15–21. [Google Scholar] [CrossRef]
- Parzych, K.R.; Klionsky, D.J. An Overview of Autophagy: Morphology, Mechanism, and Regulation. Antioxid. Redox Signal 2014, 20, 460–473. [Google Scholar] [CrossRef]
- Xu, Z.; Han, X.; Ou, D.; Liu, T.; Li, Z.; Jiang, G.; Liu, J.; Zhang, J. Targeting PI3K/AKT/MTOR-Mediated Autophagy for Tumor Therapy. Appl. Microbiol. Biotechnol. 2020, 104, 575–587. [Google Scholar] [CrossRef]
- Wang, M.; Yu, T.; Zhu, C.; Sun, H.; Qiu, Y.; Zhu, X.; Li, J. Resveratrol Triggers Protective Autophagy through the Ceramide/Akt/MTOR Pathway in Melanoma B16 Cells. Nutr. Cancer 2014, 66, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Guenther, G.G.; Peralta, E.R.; Rosales, K.R.; Wong, S.Y.; Siskind, L.J.; Edinger, A.L. Ceramide Starves Cells to Death by Downregulating Nutrient Transporter Proteins. Proc. Natl. Acad. Sci. USA 2008, 105, 17402–17407. [Google Scholar] [CrossRef]
- Levy, J.M.M.; Thorburn, A. Targeting Autophagy during Cancer Therapy to Improve Clinical Outcomes. Pharmacol. Ther. 2011, 131, 130–141. [Google Scholar] [CrossRef]
- Junco, J.J.; Mancha-Ramirez, A.; Malik, G.; Wei, S.J.; Kim, D.J.; Liang, H.; Slaga, T.J. Ursolic Acid and Resveratrol Synergize with Chloroquine to Reduce Melanoma Cell Viability. Melanoma Res. 2015, 25, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.H.; Shudo, T.; Yoshida, T.; Sugiyama, Y.; Si, J.Y.; Tsukano, C.; Takemoto, Y.; Kakizuka, A. Ellagic Acid, Extracted from Sanguisorba Officinalis, Induces G1 Arrest by Modulating PTEN Activity in B16F10 Melanoma Cells. Genes Cells 2019, 24, 688–704. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.; Kumazoe, M.; Murata, K.; Fujimura, Y.; Tachibana, H. Procyanidin C1 Inhibits Melanoma Cell Growth by Activating 67-kDa Laminin Receptor Signaling. Mol. Nutr. Food Res. 2020, 64, 1900986. [Google Scholar] [CrossRef]
- Chen, Y.; Li, H.; Zhang, G.; Wu, Y.; Xiao, J.; Liu, J.; Qiu, P.; Liu, X.; Sun, L.; Du, B.; et al. Synergistic Inhibitory Effect of Resveratrol and TK/GCV Therapy on Melanoma Cells. J. Cancer Res. Clin. Oncol. 2020, 146, 1489–1499. [Google Scholar] [CrossRef]
- Yoo, T.K.; Kim, J.S.; Hyun, T.K. Polyphenolic Composition and Anti-Melanoma Activity of White Forsythia (Abeliophyllum Distichum Nakai) Organ Extracts. Plants 2020, 9, 757. [Google Scholar] [CrossRef]
- Zhang, L.; Meng, S.; Yan, B.; Chen, J.; Zhou, L.; Shan, L.; Wang, Y. Anti-Proliferative, Pro-Apoptotic, Anti-Migrative and Tumor-Inhibitory Effects and Pleiotropic Mechanism of Theaflavin on B16F10 Melanoma Cells. Onco. Targets. Ther. 2021, 14, 1291–1304. [Google Scholar] [CrossRef]
- Martino, R.; Barreiro Arcos, M.L.; Peralta, I.; Marrassini, C.; Saint Martin, E.M.; Cogoi, L.; Cremaschi, G.; Alonso, M.R.; Anesini, C. Antiproliferative Activity of Aqueous and Polyphenol-Rich Extracts of Larrea Divaricata Cav. on a Melanoma Cell Line. Nat. Prod. Res. 2022, 36, 4431–4434. [Google Scholar] [CrossRef]
- Jensen, J.D.; Dunn, J.H.; Luo, Y.; Liu, W.; Fujita, M.; Dellavalle, R.P. Ellagic Acid Inhibits Melanoma Growth in Vitro. Dermatol. Rep. 2011, 3, e36. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.J.; Dong, Y.; Sun, C.X.; Zhang, X.H. Resveratrol Inhibits Proliferation, Promotes Differentiation and Melanogenesis in HT-144 Melanoma Cells through Inhibition of MEK/ERK Kinase Pathway. Microb. Pathog. 2017, 111, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, N.S.; Srivastava, R.A.K. Curcumin and Quercetin Synergistically Inhibit Cancer Cell Proliferation in Multiple Cancer Cells and Modulate Wnt/β-Catenin Signaling and Apoptotic Pathways in A375 Cells. Phytomedicine 2019, 52, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Duan, R.; Liang, X.; Chai, B.; Zhou, Y.; Du, H.; Suo, Y.; Chen, Z.; Li, Q.; Huang, X. Isorhamnetin Induces Melanoma Cell Apoptosis via the PI3K/Akt and NF- κ B Pathways. Biomed. Res. Int. 2020, 2020, 1057943. [Google Scholar] [CrossRef]
- Katerinaki, E.; Evans, G.S.; Lorigan, P.C.; MacNeil, S. TNF-Alpha Increases Human Melanoma Cell Invasion and Migration in Vitro: The Role of Proteolytic Enzymes. Br. J. Cancer 2003, 89, 1123–1129. [Google Scholar] [CrossRef] [PubMed]
- Sajadimajd, S.; Bahramsoltani, R.; Iranpanah, A.; Kumar Patra, J.; Das, G.; Gouda, S.; Rahimi, R.; Rezaeiamiri, E.; Cao, H.; Giampieri, F.; et al. Advances on Natural Polyphenols as Anticancer Agents for Skin Cancer. Pharmacol. Res. 2020, 151, 104584. [Google Scholar] [CrossRef]
- Tyciakova, S.; Valova, V.; Svitkova, B.; Matuskova, M. Overexpression of TNFα Induces Senescence, Autophagy and Mitochondrial Dysfunctions in Melanoma Cells. BMC Cancer 2021, 21, 507. [Google Scholar] [CrossRef]
- Afaq, F.; Malik, A.; Syed, D.; Maes, D.; Matsui, M.S.; Mukhtar, H. Pomegranate Fruit Extract Modulates UV-B-Mediated Phosphorylation of Mitogen-Activated Protein Kinases and Activation of Nuclear Factor Kappa B in Normal Human Epidermal Keratinocytes Paragraph Sign. Photochem. Photobiol. 2005, 81, 38–45. [Google Scholar] [CrossRef]
- Pal, H.C.; Sharma, S.; Strickland, L.R.; Katiyar, S.K.; Ballestas, M.E.; Athar, M.; Elmets, C.A.; Afaq, F. Fisetin Inhibits Human Melanoma Cell Invasion through Promotion of Mesenchymal to Epithelial Transition and by Targeting MAPK and NFκB Signaling Pathways. PLoS ONE 2014, 9, e86338. [Google Scholar] [CrossRef]
- Xu, A.; Lee, J.; Zhao, Y.; Wang, Y.; Li, X.; Xu, P. Potential Effect of EGCG on the Anti-Tumor Efficacy of Metformin in Melanoma Cells. J. Zhejiang Univ. Sci. B 2021, 22, 548–562. [Google Scholar] [CrossRef]
- Xiao, P.; Yang, J.; Sun, J.; Guo, L. Anticancer Effects of Kaempferol in A375 Human Malignant Melanoma Cells Are Mediated via Induction of Apoptosis, Cell Cycle Arrest, Inhibition of Cell Migration and Downregula-Tion of m-TOR/PI3K/AKT Pathway. J. Balk. Union Oncol. 2018, 23, 218–223. [Google Scholar]
- Kim, H.; Park, J.; Tak, K.H.; Bu, S.Y.; Kim, E. Chemopreventive Effects of Curcumin on Chemically Induced Mouse Skin Carcinogenesis in BK5.Insulin-like Growth Factor-1 Transgenic Mice. In Vitro Cell. Dev. Biol. Anim. 2014, 50, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chen, J.; Quan, J.; Xiang, D. Rosmarinic Acid Inhibits Proliferation and Migration, Promotes Apoptosis and Enhances Cisplatin Sensitivity of Melanoma Cells through Inhibiting ADAM17/EGFR/AKT/GSK3β Axis. Bioengineered 2021, 12, 3065–3076. [Google Scholar] [CrossRef]
- Hasnat, M.A.; Pervin, M.; Lim, J.H.; Lim, B.O. Apigenin Attenuates Melanoma Cell Migration by Inducing Anoikis through Integrin and Focal Adhesion Kinase Inhibition. Molecules 2015, 20, 21157–21166. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Wang, J.; Wu, Q.; Qian, J.; Yang, C.; Bo, P. Genistein Inhibits the Growth and Regulates the Migration and Invasion Abilities of Melanoma Cells via the FAK/Paxillin and MAPK Pathways. Oncotarget 2017, 8, 21674–21691. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Chang, L.; Qu, Y.; Liang, J.; Jin, W.; Xia, X. Tea Polyphenols Inhibit the Proliferation, Migration, and Invasion of Melanoma Cells through the down-Regulation of TLR4. Int. J. Immunopathol. Pharmacol. 2018, 32, 0394632017739531. [Google Scholar] [CrossRef]
- Borden, E.C. Interferons α and β in Cancer: Therapeutic Opportunities from New Insights. Nat. Rev. Drug Discov. 2019, 18, 219–234. [Google Scholar] [CrossRef]
- Peng, D.; Chen, L.; Sun, Y.; Sun, L.; Yin, Q.; Deng, S.; Niu, L.; Lou, F.; Wang, Z.; Xu, Z.; et al. Melanoma Suppression by Quercein Is Correlated with RIG-I and Type I Interferon Signaling. Biomed. Pharmacother. 2020, 125, 109984. [Google Scholar] [CrossRef]
- Menon, D.R.; Li, Y.; Yamauchi, T.; Osborne, D.G.; Vaddi, P.K.; Wempe, M.F.; Zhai, Z.; Fujita, M. EGCG Inhibits Tumor Growth in Melanoma by Targeting JAK-STAT Signaling and Its Downstream PD-L1/PD-L2-PD1 Axis in Tumors and Enhancing Cytotoxic T-Cell Responses. Pharmaceuticals 2021, 14, 1081. [Google Scholar] [CrossRef]
- Mondal, S.; Adhikari, N.; Banerjee, S.; Amin, S.A.; Jha, T. Matrix Metalloproteinase-9 (MMP-9) and Its Inhibitors in Cancer: A Minireview. Eur. J. Med. Chem. 2020, 194, 112260. [Google Scholar] [CrossRef]
- Zhang, X.M.; Huang, S.P.; Xu, Q. Quercetin Inhibits the Invasion of Murine Melanoma B16-BL6 Cells by Decreasing pro-MMP-9 via the PKC Pathway. Cancer Chemother. Pharmacol. 2004, 53, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.H.; Tse, A.K.W.; Kwan, H.Y.; Yu, H.; Cheng, C.Y.; Su, T.; Fong, W.F.; Yu, Z.L. Quercetin Exerts Anti-Melanoma Activities and Inhibits STAT3 Signaling. Biochem. Pharmacol. 2014, 87, 424–434. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Jiang, W.; Yu, D.; Yan, Z. Luteolin Inhibits Proliferation and Induces Apoptosis of Human Melanoma Cells In Vivo and In Vitro by Suppressing MMP-2 and MMP-9 through the PI3K/AKT Pathway. Food Funct. 2019, 10, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.O.; Cho, E.J.; Jeong, J.W.; Park, C.; Hong, S.H.; Hwang, H.J.; Moon, S.K.; Son, C.G.; Kim, W.J.; Choi, Y.H. Baicalein Inhibits the Migration and Invasion of B16F10 Mouse Melanoma Cells through Inactivation of the PI3K/Akt Signaling Pathway. Biomol. Ther. 2017, 25, 213–221. [Google Scholar] [CrossRef]
- Felcht, M.; Thomas, M. Angiogenesis in Malignant Melanoma. J. Dtsch. Dermatol. Ges. 2015, 13, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Kang, G.; Wang, T.; Huang, H. Tumor Angiogenesis and Anti-Angiogenic Gene Therapy for Cancer. Oncol. Lett. 2018, 16, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Braicu, C.; Chiorean, R.; Irimie, A.; Chira, S.; Tomuleasa, C.; Neagoe, E.; Paradiso, A.; Achimas-Cadariu, P.; Lazar, V.; Berindan-Neagoe, I. Novel Insight into Triple-Negative Breast Cancers, the Emerging Role of Angiogenesis, and Antiangiogenic Therapy. Expert Rev. Mol. Med. 2016, 18, e18. [Google Scholar] [CrossRef]
- Trapp, V.; Parmakhtiar, B.; Papazian, V.; Willmott, L.; Fruehauf, J.P. Anti-Angiogenic Effects of Resveratrol Mediated by Decreased VEGF and Increased TSP1 Expression in Melanoma-Endothelial Cell Co-Culture. Angiogenesis 2010, 13, 305–315. [Google Scholar] [CrossRef]
- Song, H.; Lim, D.Y.; Jung, J.I.; Cho, H.J.; Park, S.Y.; Kwon, G.T.; Kang, Y.H.; Lee, K.W.; Choi, M.S.; Park, J.H.Y. Dietary Oleuropein Inhibits Tumor Angiogenesis and Lymphangiogenesis in the B16F10 Melanoma Allograft Model: A Mechanism for the Suppression of High-Fat Diet-Induced Solid Tumor Growth and Lymph Node Metastasis. Oncotarget 2017, 8, 32027–32042. [Google Scholar] [CrossRef]
- Yang, G.W.; Jiang, J.S.; Lu, W.Q. Ferulic Acid Exerts Anti-Angiogenic and Anti-Tumor Activity by Targeting Fibroblast Growth Factor Receptor 1-Mediated Angiogenesis. Int. J. Mol. Sci. 2015, 16, 24011–24031. [Google Scholar] [CrossRef]
- Lee, S.H.W.; Koo, B.S.E.; Park, S.Y.I.; Kim, Y.M.I. Anti-Angiogenic Effects of Resveratrol in Combination with 5-Fluorouracil on B16 Murine Melanoma Cells. Mol. Med. Rep. 2015, 12, 2777–2783. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.K.; Lee, K.W.; Byun, S.; Lee, E.J.; Kim, J.E.; Bode, A.M.; Dong, Z.; Lee, H.J. Myricetin Inhibits UVB-Induced Angiogenesis by Regulating PI-3 Kinase In Vivo. Carcinogenesis 2010, 31, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Peng, B.; Nayak, Y.; Wang, C.; Si, F.; Liu, X.; Dou, J.; Xu, H.; Peng, G. Baicalein and Baicalin Promote Melanoma Apoptosis and Senescence via Metabolic Inhibition. Front. Cell Dev. Biol. 2020, 8, 836. [Google Scholar] [CrossRef]
- Mirzoeva, S.; Tong, X.; Bridgeman, B.B.; Plebanek, M.P.; Volpert, O.V. Apigenin Inhibits UVB-Induced Skin Carcinogenesis: The Role of Thrombospondin-1 as an Anti-Inflammatory Factor. Neoplasia 2018, 20, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Lee, D.H.; Jang, H.; Park, S.Y.; Seol, J.W. Naringenin Exerts Anticancer Effects by Inducing Tumor Cell Death and Inhibiting Angiogenesis in Malignant Melanoma. Int. J. Med. Sci. 2020, 17, 3049–3057. [Google Scholar] [CrossRef]
- Agilan, B.; Rajendra Prasad, N.; Kanimozhi, G.; Karthikeyan, R.; Ganesan, M.; Mohana, S.; Velmurugan, D.; Ananthakrishnan, D. Caffeic Acid Inhibits Chronic UVB-Induced Cellular Proliferation Through JAK-STAT3 Signaling in Mouse Skin. Photochem. Photobiol. 2016, 92, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Keravis, T.; Favot, L.; Abusnina, A.A.; Anton, A.; Justiniano, H.; Soleti, R.; Alibrahim, E.A.; Simard, G.; Andriantsitohaina, R.; Lugnier, C. Delphinidin Inhibits Tumor Growth by Acting on VEGF Signalling in Endothelial Cells. PLoS ONE 2015, 10, e0145291. [Google Scholar] [CrossRef]
- Cao, H.H.; Chu, J.H.; Kwan, H.Y.; Su, T.; Yu, H.; Cheng, C.Y.; Fu, X.Q.; Guo, H.; Li, T.; Tse, A.K.W.; et al. Inhibition of the STAT3 Signaling Pathway Contributes to Apigenin-Mediated Anti-Metastatic Effect in Melanoma. Sci. Rep. 2016, 6, 21731. [Google Scholar] [CrossRef]
- Li, L.; Fan, P.; Chou, H.; Li, J.; Wang, K.; Li, H. Herbacetin Suppressed MMP9 Mediated Angiogenesis of Malignant Melanoma through Blocking EGFR-ERK/AKT Signaling Pathway. Biochimie 2019, 162, 198–207. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- González-Gómez, P.; Sánchez, P.; Mira, H. MicroRNAs as Regulators of Neural Stem Cell-Related Pathways in Glioblastoma Multiforme. Mol. Neurobiol. 2011, 44, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Zhang, D.; Wan, X.; Bai, Y.; Yuan, C.; Wang, T.; Yuan, D.; Zhang, C.; Liu, C. Chlorogenic Acid Suppresses MiR-155 and Ameliorates Ulcerative Colitis through the NF-ΚB/NLRP3 Inflammasome Pathway. Mol. Nutr. Food Res. 2020, 64, 2000452. [Google Scholar] [CrossRef] [PubMed]
- Farshbaf-Khalili, A.; Farajnia, S.; Pourzeinali, S.; Shakouri, S.K.; Salehi-Pourmehr, H. The Effect of Nanomicelle Curcumin Supplementation and Nigella Sativa Oil on the Expression Level of MiRNA-21, MiRNA-422a, and MiRNA-503 Gene in Postmenopausal Women with Low Bone Mass Density: A Randomized, Triple-Blind, Placebo-Controlled Clinical Trial with Factorial Design. Phytother. Res. 2021, 35, 6216–6227. [Google Scholar] [PubMed]
- Biersack, B. Current State of Phenolic and Terpenoidal Dietary Factors and Natural Products as Non-Coding RNA/MicroRNA Modulators for Improved Cancer Therapy and Prevention. Non-Coding RNA Res. 2016, 1, 12–34. [Google Scholar] [CrossRef]
- Ohishi, T.; Hayakawa, S.; Miyoshi, N. Involvement of MicroRNA Modifications in Anticancer Effects of Major Polyphenols from Green Tea, Coffee, Wine, and Curry. Crit. Rev. Food Sci. Nutr. 2022, 54, 2000452. [Google Scholar] [CrossRef]
- Carpi, S.; Polini, B.; Manera, C.; Digiacomo, M.; Salsano, J.E.; Macchia, M.; Scoditti, E.; Nieri, P. MiRNA Modulation and Antitumor Activity by the Extra-Virgin Olive Oil Polyphenol Oleacein in Human Melanoma Cells. Front. Pharmacol. 2020, 11, 574317. [Google Scholar] [CrossRef]
- Du, M.; Zhang, Z.; Gao, T. Piceatannol Induced Apoptosis through Up-Regulation of MicroRNA-181a in Melanoma Cells. Biol. Res. 2017, 50, 36. [Google Scholar] [CrossRef]
- Shehzad, A.; Lee, J.; Lee, Y.S. Curcumin in Various Cancers. Biofactors 2013, 39, 56–68. [Google Scholar] [CrossRef]
- Lelli, D.; Pedone, C.; Sahebkar, A. Curcumin and Treatment of Melanoma: The Potential Role of MicroRNAs. Biomed. Pharmacother. 2017, 88, 832–834. [Google Scholar] [CrossRef]
- Zhang, P.; Bai, H.; Liu, G.; Wang, H.; Chen, F.; Zhang, B.; Zeng, P.; Wu, C.; Peng, C.; Huang, C.; et al. MicroRNA-33b, Upregulated by EF24, a Curcumin Analog, Suppresses the Epithelial-to-Mesenchymal Transition (EMT) and Migratory Potential of Melanoma Cells by Targeting HMGA2. Toxicol. Lett. 2015, 234, 151–161. [Google Scholar] [CrossRef]
- Yang, C.H.; Yue, J.; Sims, M.; Pfeffer, L.M. The Curcumin Analog EF24 Targets NF-ΚB and MiRNA-21, and Has Potent Anticancer Activity In Vitro and In Vivo. PLoS ONE 2013, 8, e71130. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Cao, Y. Curcumin Inhibits the Growth and Metastasis of Melanoma via MiR-222-3p/SOX10/Notch Axis. Dis. Markers 2022, 2022, 3129781. [Google Scholar] [CrossRef]
- Alsadi, N.; Mallet, J.-F.; Matar, C. MiRNA-200b Signature in the Prevention of Skin Cancer Stem Cells by Polyphenol-Enriched Blueberry Preparation. J. Cancer Prev. 2021, 26, 162. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Tsukamoto, S.; Huang, Y.; Makio, A.; Kumazoe, M.; Yamashita, S.; Tachibana, H. Epigallocatechin-3-O-Gallate up-Regulates MicroRNA-Let-7b Expression by Activating 67-KDa Laminin Receptor Signaling in Melanoma Cells. Sci. Rep. 2016, 6, 19225. [Google Scholar] [CrossRef]
- Wu, K.; Wei, Y.; Yu, Y.; Shan, M.; Tang, Y.; Sun, Y. Green Tea Polyphenols Inhibit Malignant Melanoma Progression via Regulating Circ_MITF/MiR-30e-3p/HDAC2 Axis. Biotechnol. Appl. Biochem. 2022, 69, 808–821. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Cui, L. Resveratrol Suppresses Melanoma by Inhibiting NF-ΚB/MiR-221 and Inducing TFG Expression. Arch. Dermatol. Res. 2017, 309, 823–831. [Google Scholar] [CrossRef]
- Zhao, S.; Tang, L.; Chen, W.; Su, J.; Li, F.; Chen, X.; Wu, L. Resveratrol-Induced Apoptosis Is Associated with Regulating the MiR-492/CD147 Pathway in Malignant Melanoma Cells. Naunyn. Schmiedebergs. Arch. Pharmacol. 2021, 394, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.X.; Young, L.C. Nutrition: The Future of Melanoma Prevention? J. Am. Acad. Dermatol. 2014, 71, 151–160. [Google Scholar] [CrossRef]
- Harris, Z.; Donovan, M.G.; Branco, G.M.; Limesand, K.H.; Burd, R. Quercetin as an Emerging Anti-Melanoma Agent: A Four-Focus Area Therapeutic Development Strategy. Front. Nutr. 2016, 3, 48. [Google Scholar] [CrossRef]
- Niedzwiecki, A.; Roomi, M.W.; Kalinovsky, T.; Rath, M. Anticancer Efficacy of Polyphenols and Their Combinations. Nutrients 2016, 8, 552. [Google Scholar] [CrossRef]
- Mirzaei, H.; Naseri, G.; Rezaee, R.; Mohammadi, M.; Banikazemi, Z.; Mirzaei, H.R.; Salehi, H.; Peyvandi, M.; Pawelek, J.M.; Sahebkar, A. Curcumin: A New Candidate for Melanoma Therapy? Int. J. Cancer 2016, 139, 1683–1695. [Google Scholar] [CrossRef] [PubMed]
- Witika, B.A.; Makoni, P.A.; Matafwali, S.K.; Mweetwa, L.L.; Shandele, G.C.; Walker, R.B. Enhancement of Biological and Pharmacological Properties of an Encapsulated Polyphenol: Curcumin. Molecules 2021, 26, 4244. [Google Scholar] [CrossRef] [PubMed]
- Faião-Flores, F.; Suarez, J.A.Q.; Fruet, A.C.; Maria-Engler, S.S.; Pardi, P.C.; Maria, D.A. Curcumin Analog DM-1 in Monotherapy or Combinatory Treatment with Dacarbazine as a Strategy to Inhibit In Vivo Melanoma Progression. PLoS ONE 2015, 10, e0118702. [Google Scholar] [CrossRef] [PubMed]
- Nihal, M.; Roelke, C.T.; Wood, G.S. Anti-Melanoma Effects of Vorinostat in Combination with Polyphenolic Antioxidant (-)-Epigallocatechin-3-Gallate (EGCG). Pharm. Res. 2010, 27, 1103–1114. [Google Scholar] [CrossRef]
- Pal, H.C.; Diamond, A.C.; Strickland, L.R.; Kappes, J.C.; Katiyar, S.K.; Elmets, C.A.; Athar, M.; Afaq, F. Fisetin, a Dietary Flavonoid, Augments the Anti-Invasive and Anti-Metastatic Potential of Sorafenib in Melanoma. Oncotarget 2016, 7, 1227–1241. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nr. | Compound | Cell Lines | Upregulated Targets | Downregulated Targets | Effects | Ref. |
|---|---|---|---|---|---|---|
| 1. | Ellagic acid | 1205Lu M852c A375 | - | IL-1β IL-8 NF-κb | Inhibition of proliferation by induction of cell cycle arrest at G1-phase; Induction of apoptosis; | [101] |
| 2. | Resveratrol | SK-MEL-2 SK-MEL-2 UACC257 B16-BL6 +In vivo | - | β-catenin STAT3 Survivin | Induction of apoptosis; | [74] |
| 3. | Resveratrol | B16 | Ceramides LC3-II | AKT mTORC1 4E-BP1 S6K | Induction of mitochondria-dependent apoptosis; Induction of autophagy and halt of protein translation; | [91] |
| 4. | Curcumin | WM-115 | Cytochrome C (cytosolic) | - | Decrease in mitochondrial membrane potential; Opening of mitochondrial permeability transition pore via cyclophilin-D and adenine nucleotide translocator-1 (ANT-1) association; | [88] |
| 5. | Resveratrol | A375 SK-MEL-31 | p53 BAX Caspase-9 Caspase-3 | BCL-2 | Reduction of cells in S phase; Inhibition of proliferation by induction of G1/S cell cycle arrest; Induction of mitochondria-dependent apoptosis; | [51] |
| 6. | Curcumin | A375 MV3 M14 MRC-5 | γ-H2AX t-BAX Cleaved caspase-8 (procaspase 8) Cleaved caspase-3 (procaspase 3) p53 (lower doses of curcumin) | MCL-1 BCL-2 BAX (full-length) NF-κb-p65 p38 p53 (higher doses of curcumin) | Inhibition of proliferation; Induction of intrinsic and extrinsic apoptotic pathways; Induction of DNA fragmentation, DNA double-strand breaks; | [63] |
| 7. | Hibiscus leaf polyphenolic extract, rich in epicatechin gallate | WS1 A375 B16F10 | Active-caspase-3 Active-caspase-8 Active-caspase-9 t-BID BAX Cytochrome c (cytosolic) Fas (membrane) FasL PI3K class III Beclin1 LC3-II p62 ATG5/12 conjugate ATG 16 ATG9 | BCL-2 p-AKT p-mTOR | Inhibition of cell growth; Induction of intrinsic and extrinsic apoptotic pathways, also supported by apoptotic bodies formation; Induction of autophagy, also supported by the formation of acidic autophagolysosomal vacuoles as well; | [68] |
| 8. | Resveratrol Ursolic acid Resveratrol + chloroquine Ursolic acid + chloroquine | B16F10 A375 | LC3-II | Beclin-1 p62 | Reduction of cell growth; Induction of apoptosis and decrease in autophagosomes levels; | [94] |
| 9. | DMU-212 (3,4,5,4′-tetramethoxystilbene, a resveratrol analog) | A375 MeWo M5 Bro | p21 P53 Cyclin B1 BAX Caspase 3 Caspase 9 p-ERK1/2 | Cyclin A2 p-CHK 2 BCL-2 | Inhibition of proliferation by induction of cell cycle arrest at prometaphase G2/M; Induction of apoptosis; | [55] |
| 10. | Curcumin | A375 C8161 +In vivo | LC3-II | p-AKT p-mTOR p-P70S6K | Inhibition of proliferation through cell cycle arrest in the G2/M phase; Inhibition of invasiveness; Induction of autophagy; | [54] |
| 11. | 3,4,5,4′trans-tetramethoxystilbene (resveratrol analog) | A375 | JNK p-p38 p38 Aurora A | - | Inhibition of proliferation through cell cycle arrest in G2/M phase and prometaphase stage; Inhibition of cell migration and attachment to the collagen-coated surface; | [61] |
| 12. | Resveratrol | HT-144 | CREB Tyrosinase PBG-D | ERK MEK1/2 | Inhibition of proliferation; Promoting differentiation; | [102] |
| 13. | Curcumin | B16-F10 L-929 | ROS BAX Caspase-3 | BCL-2 | Decrease in cell viability; Induction of apoptosis; Induction of DNA damage; Loss of mitochondrial membrane potential; Induction of oxidative stress; | [87] |
| 14. | Resveratrol | A375SM | p21 p27 ROS p-p38 MAPK p-eIF2α CHOP p53 p-p53 BAX | Cyclin B NRF2 BCL-2 | Inhibition of proliferation by induction of G2/M cell cycle arrest; Induction ROS generation and ER stress; Induction of apoptosis; | [56] |
| 15. | Resveratrol | MV3 A375 | Caspase3 Cleaved PARP1 p53 p53 mRNA p21 p21 mRNA BAX Cytochrome c (cytosolic) | ERK1/2 ERK1/2 mRNA PKM2 PKM2 mRNA BCL-2 | Inhibition of proliferation in p53-independent manner; Induction of mitochondria-dependent apoptosis; | [70] |
| 16. | Curcumin Quercetin | A375 | Caspase 3/7 Cleaved PARP | DVL2 β-catenin Cyclin D1 COX2 AXIN2 BCL-2 | Inhibition of proliferation; Induction of apoptosis; | [103] |
| 17. | Quercetin | A375SM A375P +In vivo | BAX p-JNK p-p38 p-ERK1/2 cleaved PARP | BCL-2 | Decrease in tumor weight (in vivo); Inhibition of proliferation; Induction of apoptosis; | [60] |
| 18. | Resveratrol | B16-F10, A375 | Cleaved caspase-9 Beclin 1 LC3II/LC3I | p62 PI3K p-AKT p-mTOR | Potential induction of apoptosis; Inhibition of migration and invasion and induction of autophagy; | [35] |
| 19. | Isorhamnetin | B16F10 +In vivo | BAX Caspase-3 NF-κb cytosolic | BCL-2 PI3K p-AKT NF-κb nuclear PFKFB4 | Inhibition of proliferation (in vivo and in vitro) and of migration; Induction of intrinsic apoptotic pathway; | [104] |
| 20. | Theaflavin | A375 HFF-1 +In vivo | BAX mRNA BCL-2 mRNA Bim mRNA c-MYC mRNA p21 mRNA p53 mRNA PUMA mRNA ATM, p-ATM CHK1, p-CHK1 p-CHK2 p-P53, c-PARP ASK1, JNK, p-JNK C-JUN, p-C-JUN c-CASP8, c-CASP3 | - | Inhibition of tumor growth (in vivo); Induction of intrinsic apoptosis; | [62] |
| Nr. | Compound | Study Type | Model | Effects | Targets | Ref. |
|---|---|---|---|---|---|---|
| Effects of polyphenols on cell proliferation, migration, and metastasis | ||||||
| 1. | Pomegranate extract | In vitro | Normal human epidermal keratinocytes NHEK | Inhibition of UV-B-mediated phosphorylation of MAPK | ERKl/2, JNK1/2, and p38↓ | [108] |
| 2. | EGCG + metformin | In vitro | B16F10 cells | Inhibition of cell growth and STAT3/NF-κb pathway | STAT3 and NF-κb p65↓ | [110] |
| 3. | EGCG | In vitro | 1205Lu, HS294T, and A375 cells | Inhibition of cell proliferation | STAT1↓ | [119] |
| 4. | EGCG | In vivo | C57BL/6 mice | Inhibition of tumor growth | STAT1↓ | [119] |
| 5. | Curcumin | In vivo | BK5.IGF-1 transgenic (Tg) mice | Inhibition of tumor growth | IGF-1↓ | [112] |
| 6. | Apigenin | In vitro | A2058 and A375 melanoma cells | Suppression of melanoma cells proliferation Induction of anoikis | FAK↓ ERK1/2↓ | [114] |
| 7. | Genistein | In vitro | B16F10 melanoma cells | Inhibition of cell proliferation, migration, and metastasis | p-p38, p-ERK, and p-JNK↓ | [115] |
| 8. | Quercetin | In vitro | B16 and A375 cells | Suppressed proliferation Inhibition of migration and invasion | RIG-I↑ IFN-I↑ STAT1↑ | [118] |
| 9. | Quercetin | In vivo | C57BL/6 J male mice | Inhibits tumor growth | RIG-I↑ IFN-I↑ STAT1↑ | [118] |
| 10. | Quercetin | In vitro | A375 and A2058 cells | Inhibition of proliferation of melanoma cells Suppression of migratory and invasive properties | MCL-1, MMP-2, MMP-9, and VEGF↓ | [122] |
| 11. | Fisetin | In vitro | A375 and Hs294T | Inhibition of melanoma cell invasion | MEK1/2↓ NF-κb↓ | [109] |
| 12. | Rosmarinic acid | In vitro | A375 | Inhibits proliferation and migration | ADAM17/EGFR/AKT/GSK3β↓ | [113] |
| 13. | Luteolin | In vitro | A375 | Inhibited the proliferation, migration, and invasion | MMP-2 and MMP-9↓ TIMP-1 and TIMP-2↑ | [123] |
| 14. | Baicalein | In vitro | B16F10 cells | Inhibits the migration and invasion of cells | MMP-2 and MMP-9↓ | [124] |
| 15. | Kaempferol | In vitro | A375 cell | Inhibition of cell migration | mTOR, phosphorylated (p) mTOR, PI3K, p-PI3K, and AKT↓ | [111] |
| Effects of polyphenols on angiogenesis | ||||||
| 16. | Baicalein and baicalin | In vitro | Mel586 SK-MEL-2 A375 and B16F0 cells | Inhibit melanoma Cell growth and proliferation | mTOR↓ | [133] |
| 17. | Resveratrol | In vitro | Co-culture | Antiangiogenic effects | VEGF↓ TSP-1↑ | [128] |
| 18. | Oleuropein | In vivo | B16F10 melanoma allograft model | Inhibits tumor angiogenesis and lymphangiogenesis | VEGF↓ | [129] |
| 19. | Ferulic acid | In vivo | C57BL/6 mice | Growth-inhibitory activity Inhibition of angiogenesis | FGFR1↓ PI3K↓ AKT↓ | [130] |
| 20. | Resveratrol and 5-FU | In vivo | Balb/c nu/nu mice | Reduced the number of microvascular vessels | AMPK↑ VASP and VEGF↓ | [131] |
| 21. | Myricetin | In vivo | SKH-1 hairless mouse | Inhibits UVB-induced angiogenesis | VEGF↓ | [132] |
| 22. | Apigenin | In vivo | WT mice TKO mice | Inhibition of UVB-induced cutaneous angiogenesis | TSP-1↑ | [134] |
| 23. | Apigenin | In vitro | A375 and G361 cell lines, and murine melanoma B16F10 cells | Inhibition of angiogenesis | VEGF↓ | [138] |
| 24. | Naringenin | In vitro and ex vivo | B16F10 and SK-MEL-28 cells | Inhibition of angiogenesis | Tie2↓ | [135] |
| 25. | Caffeic acid | In vivo | Male Swiss albino mice | Inhibition of angiogenesis and proliferation | TSP-1↑ | [136] |
| 26. | Delphinidin | In vitro | B16-F10 melanoma cell | Antiangiogenic activity | VEGF↓ | [137] |
| 27. | Herbacetin | In vitro | A375 and Hs294T cells | Suppressed angiogenesis | MMP9↓ | [139] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isacescu, E.; Chiroi, P.; Zanoaga, O.; Nutu, A.; Budisan, L.; Pirlog, R.; Atanasov, A.G.; Berindan-Neagoe, I. Melanoma Cellular Signaling Transduction Pathways Targeted by Polyphenols Action Mechanisms. Antioxidants 2023, 12, 407. https://doi.org/10.3390/antiox12020407
Isacescu E, Chiroi P, Zanoaga O, Nutu A, Budisan L, Pirlog R, Atanasov AG, Berindan-Neagoe I. Melanoma Cellular Signaling Transduction Pathways Targeted by Polyphenols Action Mechanisms. Antioxidants. 2023; 12(2):407. https://doi.org/10.3390/antiox12020407
Chicago/Turabian StyleIsacescu, Ecaterina, Paul Chiroi, Oana Zanoaga, Andreea Nutu, Liviuta Budisan, Radu Pirlog, Atanas G. Atanasov, and Ioana Berindan-Neagoe. 2023. "Melanoma Cellular Signaling Transduction Pathways Targeted by Polyphenols Action Mechanisms" Antioxidants 12, no. 2: 407. https://doi.org/10.3390/antiox12020407