
Tryptophan Metabolism in Bipolar Disorder in a Longitudinal Setting

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants and Procedure
2.2. Laboratory Methods
2.3. Materials
2.4. Statistical Analyses
3. Results
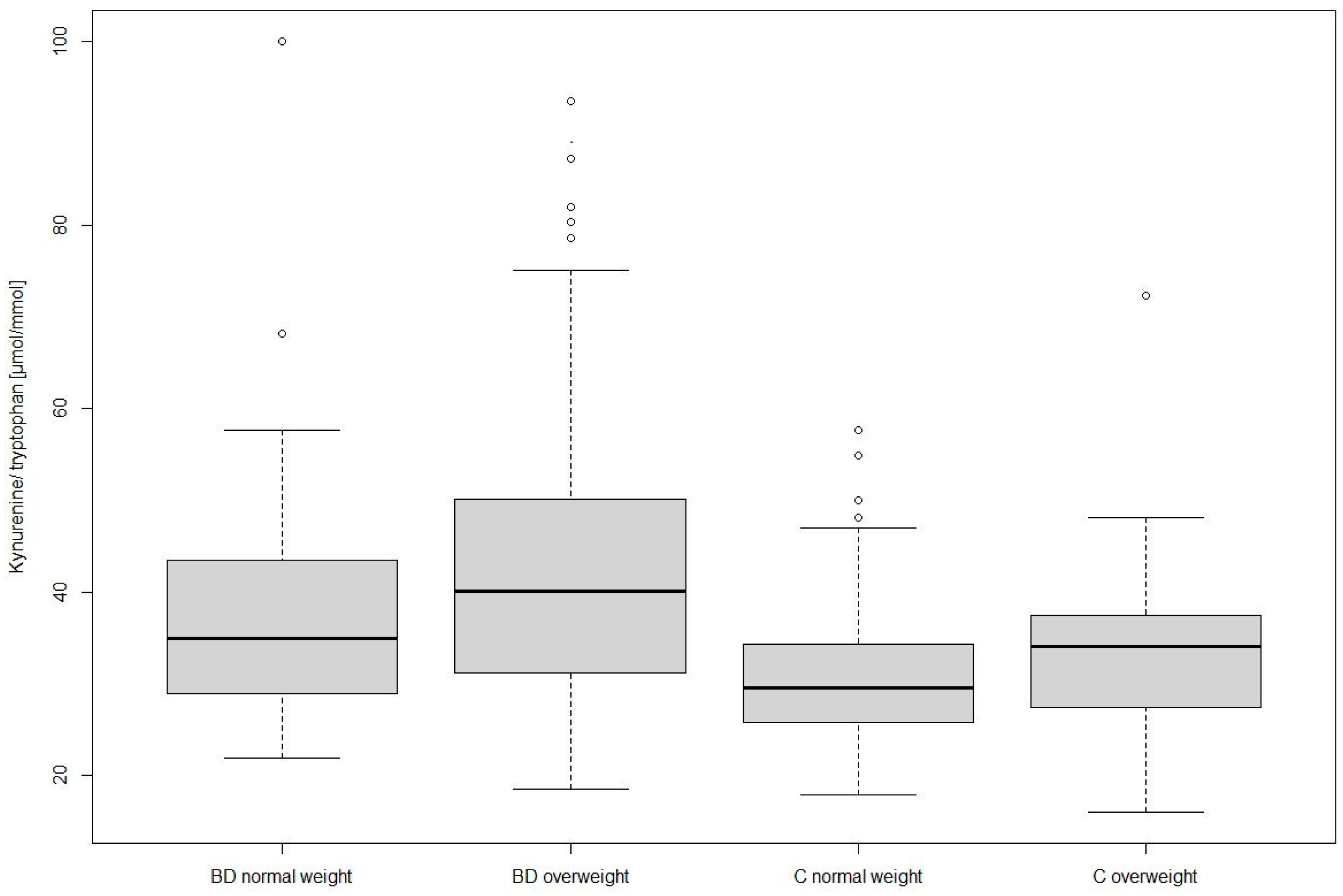
3.1. Differences between Groups and Weight Classes
3.2. Longitudinal Setting
4. Discussion
Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brietzke, E.; Mansur, R.B.; Soczynska, J.; Powell, A.M.; McIntyre, R.S. A theoretical framework informing research about the role of stress in the pathophysiology of bipolar disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 39, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.; Maes, M. Bipolar disorder: Role of immune-inflammatory cytokines, oxidative and nitrosative stress and tryptophan catabolites. Curr. Psychiatry Rep. 2015, 17, 8. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.M.; Su, T.P.; Li, C.T.; Tsai, S.J.; Chen, M.H.; Tu, P.C.; Chiou, W.F. Comparison of pro-inflammatory cytokines among patients with bipolar disorder and unipolar depression and normal controls. Bipolar. Disord. 2015, 17, 269–277. [Google Scholar] [CrossRef]
- Maes, M.; Meltzer, H.Y.; Bosmans, E.; Bergmans, R.; Vandoolaeghe, E.; Ranjan, R.; Desnyder, R. Increased plasma concentrations of interleukin-6, soluble interleukin-6, soluble interleukin-2 and transferrin receptor in major depression. J. Affect. Disord. 1995, 34, 301–309. [Google Scholar] [CrossRef]
- Munkholm, K.; Braüner, J.V.; Kessing, L.V.; Vinberg, M. Cytokines in bipolar disorder vs. healthy control subjects: A systematic review and meta-analysis. J. Psychiatr. Res. 2013, 47, 1119–1133. [Google Scholar] [CrossRef]
- Brown, N.C.; Andreazza, A.C.; Young, L.T. An updated meta-analysis of oxidative stress markers in bipolar disorder. Psychiatry Res. 2014, 218, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Gostner, J.M.; Becker, K.; Fuchs, D.; Sucher, R. Redox regulation of the immune response. Redox Rep. 2013, 18, 88–94. [Google Scholar] [CrossRef]
- Zhao, Z.; Okusaga, O.O.; Quevedo, J.; Soares, J.C.; Teixeira, A.L. The potential association between obesity and bipolar disorder: A meta-analysis. J. Affect. Disord. 2016, 202, 120–123. [Google Scholar] [CrossRef]
- Kucyi, A.; Alsuwaidan, M.T.; Liauw, S.S.; Mcintyre, R.S. Aerobic physical exercise as a possible treatment for neurocognitive dysfunction in bipolar disorder. Postgrad. Med. 2010, 122, 107–116. [Google Scholar] [CrossRef]
- Lackner, N.; Bengesser, S.A.; Birner, A.; Painold, A.; Fellendorf, F.T.; Platzer, M.; Reininghaus, B.; Weiss, E.M.; Mangge, H.; McIntyre, R.S.; et al. Abdominal obesity is associated with impaired cognitive function in euthymic bipolar individuals. World J. Biol. Psychiatry 2016, 17, 535–546. [Google Scholar] [CrossRef]
- McIntyre, R.S.; Danilewitz, M.; Liauw, S.S.; Kemp, D.E.; Nguyen, H.T.; Kahn, L.S.; Kucyi, A.; Soczynska, J.K.; Woldeyohannes, H.O.; Lachowski, A.; et al. Bipolar disorder and metabolic syndrome: An international perspective. J. Affect. Disord. 2010, 126, 366–387. [Google Scholar] [CrossRef]
- Geddes, J.R.; Miklowitz, D.J. Treatment of bipolar disorder. Lancet 2013, 381, 1672–1682. [Google Scholar] [CrossRef]
- Swartz, H.A.; Fagiolini, A. Cardiovascular disease and bipolar disorder: Risk and clinical implications. J. Clin. Psychiatry 2012, 73, 1563–1565. [Google Scholar] [CrossRef]
- Leboyer, M.; Soreca, I.; Scott, J.; Frye, M.; Henry, C.; Tamouza, R.; Kupfer, D.J. Can bipolar disorder be viewed as a multi-system inflammatory disease? J. Affect. Disord. 2012, 141, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widner, B.; Laich, A.; Sperner-Unterweger, B.; Ledochowski, M.; Fuchs, D. Neopterin production, tryptophan degradation, and mental depression—What is the link? Brain Behav. Immun. 2002, 16, 590–595. [Google Scholar] [CrossRef]
- Dai, X.; Zhu, B.T. Indoleamine 2,3-dioxygenase tissue distribution and cellular localization in mice: Implications for its biological functions. Histochem. Cytochem. 2010, 58, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badawy, A.A. Tryptophan availability for kynurenine pathway metabolism across the life span: Control mechanisms and focus on aging, exercise, diet and nutritional supplements. Neuropharmacology 2017, 112, 248–263. [Google Scholar] [CrossRef] [PubMed]
- Musso, T.; Gusella, G.L.; Brooks, A.; Longo, D.L.; Varesio, L. Interleukin-4 inhibits indoleamine 2,3-dioxygenase expression in human monocytes. Blood 1994, 83, 1408–1411. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.W.; Feng, G. Relationship between interferon-γ, indoleamine 2,3-dioxygenase, and tryptophan catabolism. FASEB J. 1991, 5, 2516–2522. [Google Scholar] [CrossRef]
- Fuchs, D.; Möller, A.A.; Reibnegger, G.; Stöckle, E.; Werner, E.R.; Wachter, H. Decreased serum tryptophan in patients with HIV-1 infection correlates with increased serum neopterin and with neurologic/psychiatric symptoms. J. Acquir. Immune Defic. Syndr. 1990, 3, 873–876. [Google Scholar]
- Myint, A.M. Kynurenines: From the perspective of major psychiatric disorders. FEBS J. 2012, 279, 1375–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michels, N.; Clarke, G.; Olavarria-Ramirez, L.; Gómez-Martínez, S.; Díaz, L.E.; Marcos, A.; Widhalm, K.; Carvalho, L.A. Psychosocial stress and inflammation driving tryptophan breakdown in children and adolescents: A cross-sectional analysis of two cohorts. Psychoneuroendocrinology 2018, 94, 104–111. [Google Scholar] [CrossRef] [PubMed]
- O’Farrell, K.; Harkin, A. Stress-related regulation of the kynurenine pathway: Relevance to neuropsychiatric and degenerative disorders. Neuropharmacology 2017, 112 Pt B, 307–323. [Google Scholar] [CrossRef] [Green Version]
- Myint, A.M.; Kim, Y.K.; Verkerk, R.; Park, S.H.; Scharpé, S.; Steinbusch, H.W.; Leonard, B.E. Tryptophan breakdown pathway in bipolar mania. J. Affect. Disord. 2007, 102, 65–72. [Google Scholar] [CrossRef]
- Brandacher, G.; Hoeller, E.; Fuchs, D.; Weiss, H.G. Chronic immune activation underlies morbid obesity: Is IDO a key player? Curr. Drug Metab. 2007, 8, 289–295. [Google Scholar] [CrossRef] [PubMed]
- De Bie, J.; Guest, J.; Guillemin, G.J.; Grant, R. Central kynurenine pathway shift with age in women. J. Neurochem. 2016, 136, 995–1003. [Google Scholar] [CrossRef] [Green Version]
- Reininghaus, E.Z.; Dalkner, N.; Riedrich, K.; Fuchs, D.; Gostner, J.M.; Reininghaus, B. Sex Specific Changes in Tryptophan Breakdown over a 6 Week Treatment Period. Front. Psychiatry 2019, 10, 74. [Google Scholar] [CrossRef]
- Miura, H.; Ando, Y.; Noda, Y.; Isobe, K.; Ozaki, N. Long-lasting effects of inescapable-predator stress on brain tryptophan metabolism and the behavior of juvenile mice. Stress 2011, 14, 262–272. [Google Scholar] [CrossRef]
- Widner, B.; Leblhuber, F.; Walli, J.; Tilz, G.P.; Demel, U.; Fuchs, D. Tryptophan degradation and immune activation in Alzheimer’s disease. J. Neural Transm. 2000, 107, 343–353. [Google Scholar] [CrossRef]
- Maddison, D.C.; Giorgini, F. The kynurenine pathway and neurodegenerative disease. Semin. Cell Dev. Biol. 2015, 40, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Bartoli, F.; Misiak, B.; Callovini, T.; Cavaleri, D.; Cioni, R.M.; Crocamo, C.; Savitz, J.B.; Carrà, G. The kynurenine pathway in bipolar disorder: A meta-analysis on the peripheral blood levels of tryptophan and related metabolites. Mol. Psychiatry 2020, 26, 3419–3429. [Google Scholar] [CrossRef]
- Marx, W.; McGuinness, A.J.; Rocks, T.; Ruusunen, A.; Cleminson, J.; Walker, A.J.; Gomes-da-Costa, S.; Lane, M.; Sanches, M.; Diaz, A.P.; et al. The kynurenine pathway in major depressive disorder, bipolar disorder, and schizophrenia: A meta-analysis of 101 studies. Mol. Psychiatry 2020, 26, 4158–4178. [Google Scholar] [CrossRef] [PubMed]
- Trepci, A.; Sellgren, C.M.; Pålsson, E.; Brundin, L.; Khanlarkhani, N.; Schwieler, L.; Landén, M.; Erhardt, S. Central levels of tryptophan metabolites in subjects with bipolar disorder. Eur. Neuropsychopharmacol. 2021, 43, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Sellgren, C.M.; Gracias, J.; Jungholm, O.; Perlis, R.H.; Engberg, G.; Schwieler, L.; Landen, M.; Erhardt, S. Peripheral and central levels of kynurenic acid in bipolar disorder subjects and healthy controls. Transl. Psychiatry 2019, 9, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, C.; Batchelor, J.R.; Fuchs, D.; Hausen, A.; Lang, A.; Niederwieser, D.; Reibnegger, G.; Swetly, P.; Troppmair, J.; Wachter, H. Immune response-associated production of neopterin. Release from macrophages primarily under control of interferon-gamma. J. Exp. Med. 1984, 160, 310–316. [Google Scholar] [CrossRef] [Green Version]
- Murr, C.; Widner, B.; Wirleitner, B.; Fuchs, D. Neopterin as a marker for immune system activation. Curr. Drug Metab. 2002, 3, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Schröcksnadel, K.; Wirleitner, B.; Winkler, C.; Fuchs, D. Monitoring tryptophan metabolism in chronic immune activation. Clin. Chim. Acta 2006, 364, 82–90. [Google Scholar] [CrossRef]
- Capuron, L.; Schroecksnadel, S.; Féart, C.; Aubert, A.; Higueret, D.; Barberger-Gateau, P.; Layé, S.; Fuchs, D. Chronic low-grade inflammation in elderly persons is associated with altered tryptophan and tyrosine metabolism: Role in neuropsychiatric symptoms. Biol. Psychiatry 2011, 70, 175–182. [Google Scholar] [CrossRef]
- Hoekstra, R.; Fekkes, D.; Pepplinkhuizen, L.; Loonen, A.J.; Tuinier, S.; Verhoeven, W.M. Nitric oxide and neopterin in bipolar affective disorder. Neuropsychobiology 2006, 54, 75–81. [Google Scholar] [CrossRef]
- Van den Ameele, S.; Fuchs, D.; Coppens, V.; de Boer, P.; Timmers, M.; Sabbe, B.; Morrens, M. Markers of Inflammation and Monoamine Metabolism Indicate Accelerated Aging in Bipolar Disorder. Front. Psychiatry 2018, 9, 250. [Google Scholar] [CrossRef] [Green Version]
- Reininghaus, E.Z.; McIntyre, R.S.; Reininghaus, B.; Geisler, S.; Bengesser, S.A.; Lackner, N.; Hecht, K.; Birner, A.; Kattnig, F.; Unterweger, R.; et al. Tryptophan breakdown is increased in euthymic overweight individuals with bipolar disorder: A preliminary report. Bipolar. Disord. 2014, 16, 432–440. [Google Scholar] [CrossRef]
- Wittchen, H.U.; Wunderlich, U.; Gruschwitz, S.; Zaudig, M. SKID-I: Strukturiertes Klinisches Interview für DSM-IV, Achse I: Psychische Störungen; Hogrefe: Göttingen, Germany, 1997. [Google Scholar]
- Geisler, S.; Mayersbach, P.; Becker, K.; Schennach, H.; Fuchs, D.; Gostner, J.M. Serum tryptophan, kynurenine, phenylalanine, tyrosine and neopterin concentrations in 100 healthy blood donors. Pteridines 2015, 26, 31–36. [Google Scholar] [CrossRef]
- Widner, B.; Werner, E.R.; Schennach, H.; Wachter, H.; Fuchs, D. Simultaneous measurement of serum tryptophan and kynurenine by HPLC. Clin. Chem. 1997, 43, 2424–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laich, A.; Neurauter, G.; Widner, B.; Fuchs, D. More rapid method for simultaneous measurement of tryptophan and kynurenine by HPLC. Clin. Chem. 2002, 48, 579–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widner, B.; Werner, E.R.; Schennach, H.; Fuchs, D. An HPLC method to determine tryptophan and kynurenine in serum simultaneously. Adv. Exp. Med. Biol. 1999, 467, 827–832. [Google Scholar] [CrossRef]
- Hamilton, M. A rating scale for depression. J. Neurol. Neurosurg. Psychiatry 1960, 23, 56–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, R.C.; Biggs, J.T.; Ziegler, V.E.; Meyer, D.A. A rating scale for mania: Reliability, validity and sensitivity. Br. J. Psychiatry 1978, 133, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, T.; Bauer, I.E.; Meyer, T.D.; Kapczinski, F.; Soares, J.C. Neuroprogression and cognitive functioning in bipolar disorder: A systematic review. Curr. Psychiatry Rep. 2015, 17, 75. [Google Scholar] [CrossRef] [PubMed]
- Badawy, A.A. Kynurenine pathway and human systems. Exp. Gerontol. 2020, 129, 110770. [Google Scholar] [CrossRef]
- Gostner, J.M.; Geisler, S.; Stonig, M.; Mair, L.; Sperner-Unterweger, B.; Fuchs, D. Tryptophan Metabolism and Related Pathways in Psychoneuroimmunology: The Impact of Nutrition and Lifestyle. Neuropsychobiology 2020, 79, 89–99. [Google Scholar] [CrossRef]
- Schwarcz, R.; Stone, T.W. The kynurenine pathway and the brain: Challenges, controversies and promises. Neuropharmacology. 2017, 112 Pt B, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Cox, A.J.; West, N.P.; Cripps, A.W. Obesity, inflammation, and the gut microbiota. Lancet Diabetes Endocrinol. 2015, 3, 207–215. [Google Scholar] [CrossRef]
- Reininghaus, B.; Riedrich, K.; Dalkner, N.; Bengesser, S.A.; Birner, A.; Platzer, M.; Hamm, C.; Gostner, J.M.; Fuchs, D.; Reininghaus, E.Z. Changes in the tryptophan-kynurenine axis in association to therapeutic response in clinically depressed patients undergoing psychiatric rehabilitation. Psychoneuroendocrinology 2018, 94, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Dalkner, N.; Platzer, M.; Bengesser, S.A.; Birner, A.; Fellendorf, F.T.; Queissner, R.; Painold, A.; Mangge, H.; Fuchs, D.; Reininghaus, B.; et al. The role of tryptophan metabolism and food craving in the relationship between obesity and bipolar disorder. Clin. Nutr. 2018, 37, 1744–1751. [Google Scholar] [CrossRef]
- Raheja, U.K.; Fuchs, D.; Giegling, I.; Brenner, L.A.; Rovner, S.F.; Mohyuddin, I.; Weghuber, D.; Mangge, H.; Rujescu, D.; Postolache, T.T. In psychiatrically healthy individuals, overweight women but not men have lower tryptophan levels. Pteridines 2015, 26, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangge, H.; Summers, K.L.; Meinitzer, A.; Zelzer, S.; Almer, G.; Prassl, R.; Schnedl, W.J.; Reininghaus, E.; Paulmichl, K.; Weghuber, D.; et al. Obesity-related dysregulation of the tryptophan-kynurenine metabolism: Role of age and parameters of the metabolic syndrome. Obesity 2014, 22, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Kubera, M.; Obuchowiczwa, E.; Goehler, L.; Brzeszcz, J. Depression’s multiple comorbidities explained by (neuro)inflammatory and oxidative & nitrosative stress pathways. Neuro Endocrinol. Lett. 2011, 32, 7–24. [Google Scholar] [PubMed]
- Ghosh, M.; Wang, H.D.; McNeill, J.R. Role of oxidative stress and nitric oxide in regulation of spontaneous tone in aorta of DOCA-salt hypertensive rats. Br. J. Pharmacol. 2004, 141, 562–573. [Google Scholar] [CrossRef]
- Youn, J.Y.; Siu, K.L.; Lob, H.E.; Itani, H.; Harrison, D.G.; Cai, H. Role of vascular oxidative stress in obesity and metabolic syndrome. Diabetes 2014, 63, 2344–2355. [Google Scholar] [CrossRef] [Green Version]
- Vadodaria, K.C.; Stern, S.; Marchetto, M.C.; Gage, F.H. Serotonin in psychiatry: In vitro disease modeling using patient-derived neurons. Cell Tissue Res. 2018, 371, 161–170. [Google Scholar] [CrossRef]
- Anderson, G.; Jacob, A.; Bellivier, F.; Geoffroy, P.A. Bipolar Disorder: The Role of the Kynurenine and Melatonergic Pathways. Curr. Pharm. Des. 2016, 22, 987–1012. [Google Scholar] [CrossRef] [PubMed]
- Christen, S.; Peterhans, E.; Stocker, R. Antioxidant activities of some tryptophan metabolites: Possible implication for inflammatory diseases. Proc. Natl. Acad. Sci. USA 1990, 87, 2506–2510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuda, S.; Nishiyama, N.; Saito, H.; Katsuki, H. Hydrogen peroxide-mediated neuronal cell death induced by an endogenous neurotoxin, 3-hydroxykynurenine. Proc. Natl. Acad. Sci. USA 1996, 93, 12553–12558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birner, A.; Platzer, M.; Bengesser, S.A.; Dalkner, N.; Fellendorf, F.T.; Queissner, R.; Pilz, R.; Rauch, P.; Maget, A.; Hamm, C.; et al. Increased breakdown of kynurenine towards its neurotoxic branch in bipolar disorder. PLoS ONE 2017, 12, e0172699. [Google Scholar] [CrossRef] [PubMed]
- Platzer, M.; Dalkner, N.; Fellendorf, F.T.; Birner, A.; Bengesser, S.A.; Queissner, R.; Kainzbauer, N.; Pilz, R.; Herzog-Eberhard, S.; Hamm, C.; et al. Tryptophan breakdown and cognition in bipolar disorder. Psychoneuroendocrinology 2017, 81, 144–150. [Google Scholar] [CrossRef]
- Opitz, C.A.; Litzenburger, U.M.; Sahm, F.; Ott, M.; Tritschler, I.; Trump, S.; Schumacher, T.; Jestaedt, L.; Schrenk, D.; Weller, M.; et al. An endogenous tumour-promoting ligand of the human aryl hydrocarbon receptor. Nature 2011, 478, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Bock, K.W. Aryl hydrocarbon receptor (AHR) functions in NAD+ metabolism, myelopoiesis and obesity. Biochem. Pharmacol. 2019, 163, 128–132. [Google Scholar] [CrossRef]
- Thomas, S.R.; Terentis, A.C.; Cai, H.; Takikawa, O.; Levina, A.; Lay, P.A.; Freewan, M.; Stocker, R. Post-translational regulation of human indoleamine 2,3-dioxygenase activity by nitric oxide. J. Biol. Chem. 2007, 282, 23778–23787. [Google Scholar] [CrossRef] [Green Version]
- Sathyasaikumar, K.V.; Tararina, M.; Wu, H.Q.; Neale, S.A.; Weisz, F.; Salt, T.E.; Schwarcz, R. Xanthurenic Acid Formation from 3-Hydroxykynurenine in the Mammalian Brain: Neurochemical Characterization and Physiological Effects. Neuroscience 2017, 367, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Frick, B.; Schroecksnadel, K.; Neurauter, G.; Leblhuber, F.; Fuchs, D. Increasing production of homocysteine and neopterin and degradation of tryptophan with older age. Clin. Biochem. 2004, 37, 684–687. [Google Scholar] [CrossRef]


{kind=link}
| BD | C | Statistics | |||||
|---|---|---|---|---|---|---|---|
| Normal Weight (n = 80) (M ± SD) | Overweight (n = 146) (M ± SD) | Normal Weight (n = 97) (M ± SD) | Overweight (n = 45) (M ± SD) | Fgroup | Fweightgroup | Fgroup × weight group | |
| Age [years] | 41.33 (13.21) | 45.36 (12.73) | 33.31(14.35) | 41.69 (14.10) | 14.57 ** | 16.38 ** | 2.02 |
| Sex | 40% ♂ 60% ♀ | 58.2% ♂ 41.8% ♀ | 33.0% ♂ 67.0% ♀ | 46.7% ♂ 53.3% ♀ | |||
| BMI [kg/m2] | 22.61 (1.69) | 30.54 (5.12) | 21.68 (1.77) | 29.14 (3.43) | 7.82 ** | 341.63 ** | 0.32 |
| Tryptophan [µmol/L] | 55.69 (10.76) | 57.74 (10.43) | 62.54 (10.55) | 61.40 (7.80) | #18.12 ** | # 0.55 | # 0.79 |
| Kynurenine [µmol/L] | 2.04 (0.62) | 2.43 (0.89) | 1.90 (0.46) | 2.04 (0.51) | # 5.39 | #4.15 * | # 3.17 |
| Kynurenine/tryptophan [µmol/mmol] | 37.22 (11.68) | 42.39 (14.23) | 30.86 (7.95) | 33.73 (9.40) | #19.17 ** | # 2.26 | # 2.26 |
| Neopterin [nmol/L] | a 6.38 (4.59) | b 6.87 (3.17) | c 5.93 (1.71) | d 6.09 (1.67) | 0.10 | 0.11 | 0.19 |
| Mean t1–t3 | t1 | t1–t2 | t2 | t2–t3 | t3 | |
|---|---|---|---|---|---|---|
| Age [years] (M ± SD) | 47.16 (13.29) | |||||
| Sex | 53.3% male 46.7% female | |||||
| BMI [kg/m2] (M ± SD) | 28.63 (5.35) | 28.31 (5.30) | 28.95 (5.50) | 28.90 (5.34) | ||
| Weight group | 25.3% normal 74.7% overweight | |||||
| Time [days] (M ± SD) | 317.51 (176.27) | 384.15 (227.52) | ||||
| Euthymic (HAMD < 14 + YMRS < 9) (YES) | 74.7% | 92.0% | 84.0% | |||
| Illness episode None Mania Depression | 58.7% 12.0% 29.3% | 53.3% 16.0% 29.3% | ||||
| Tryptophan [µmol/L] (M ± SD) | 58.44 (9.46) | 57.89 (9.99) | 59.39 (9.32) | |||
| Kynurenine [µmol/L] (M ± SD) | 2.23 (0.67) | 2.15 (0.65) | 2.21 (0.65) | |||
| Kynurenine/tryptophan [µmol/mmol] (M ± SD) | 38.34 (10.83) | 37.72 (11.90) | 37.73 (12.04) | |||
| Neopterin a [nmol/L] (M ± SD) | 7.09 (3.33) | 7.16 (3.39) | 7.48 (4.31) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fellendorf, F.T.; Gostner, J.M.; Lenger, M.; Platzer, M.; Birner, A.; Maget, A.; Queissner, R.; Tmava-Berisha, A.; Pater, C.A.; Ratzenhofer, M.; et al. Tryptophan Metabolism in Bipolar Disorder in a Longitudinal Setting. Antioxidants 2021, 10, 1795. https://doi.org/10.3390/antiox10111795
Fellendorf FT, Gostner JM, Lenger M, Platzer M, Birner A, Maget A, Queissner R, Tmava-Berisha A, Pater CA, Ratzenhofer M, et al. Tryptophan Metabolism in Bipolar Disorder in a Longitudinal Setting. Antioxidants. 2021; 10(11):1795. https://doi.org/10.3390/antiox10111795
Chicago/Turabian StyleFellendorf, Frederike T., Johanna M. Gostner, Melanie Lenger, Martina Platzer, Armin Birner, Alexander Maget, Robert Queissner, Adelina Tmava-Berisha, Cornelia A. Pater, Michaela Ratzenhofer, and et al. 2021. "Tryptophan Metabolism in Bipolar Disorder in a Longitudinal Setting" Antioxidants 10, no. 11: 1795. https://doi.org/10.3390/antiox10111795