Hydrogen Sulfide Attenuates Aortic Remodeling in Aortic Dissection Associating with Moderated Inflammation and Oxidative Stress through a NO-Dependent Pathway

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Development of Aortic Dissection Model in Mice
2.2. External Aortic Diameter Measurement
2.3. Histological and TUNEL Analysis
2.4. Immunohistochemistry
2.5. Cell Culture and Treatments
2.6. Measurement of SOD Activity, Malondialdehyde Level, and NO Level
2.7. Quantitative Reverse Transcription-Polymerase Chain Reaction
2.8. Western Blotting
2.9. Detection of Reactive Oxygen Species
2.10. Cell Proliferation Assay
2.11. Statistical Analysis
3. Results
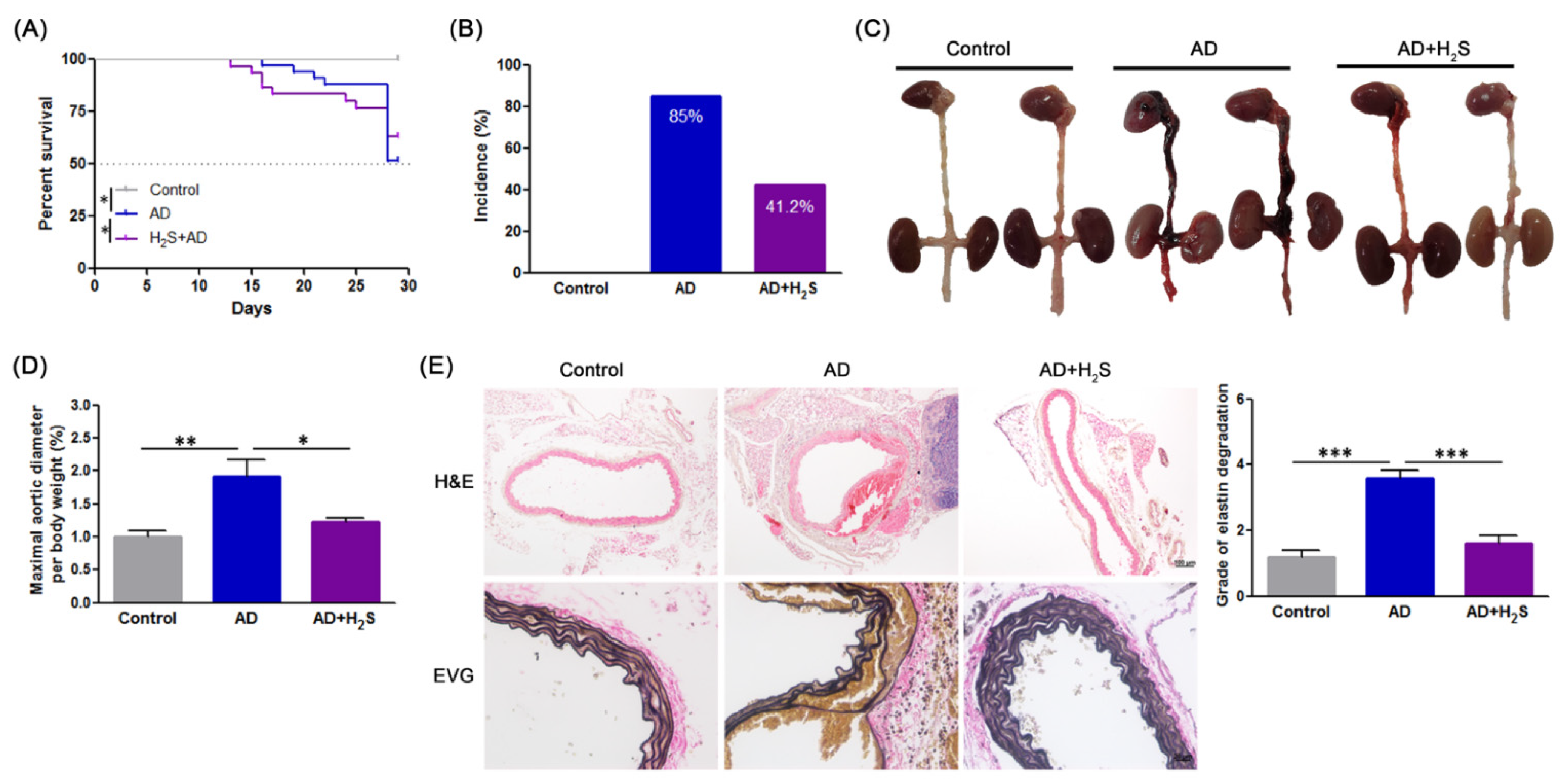
3.1. H2S Suppresses the Development of AD in Animal Models
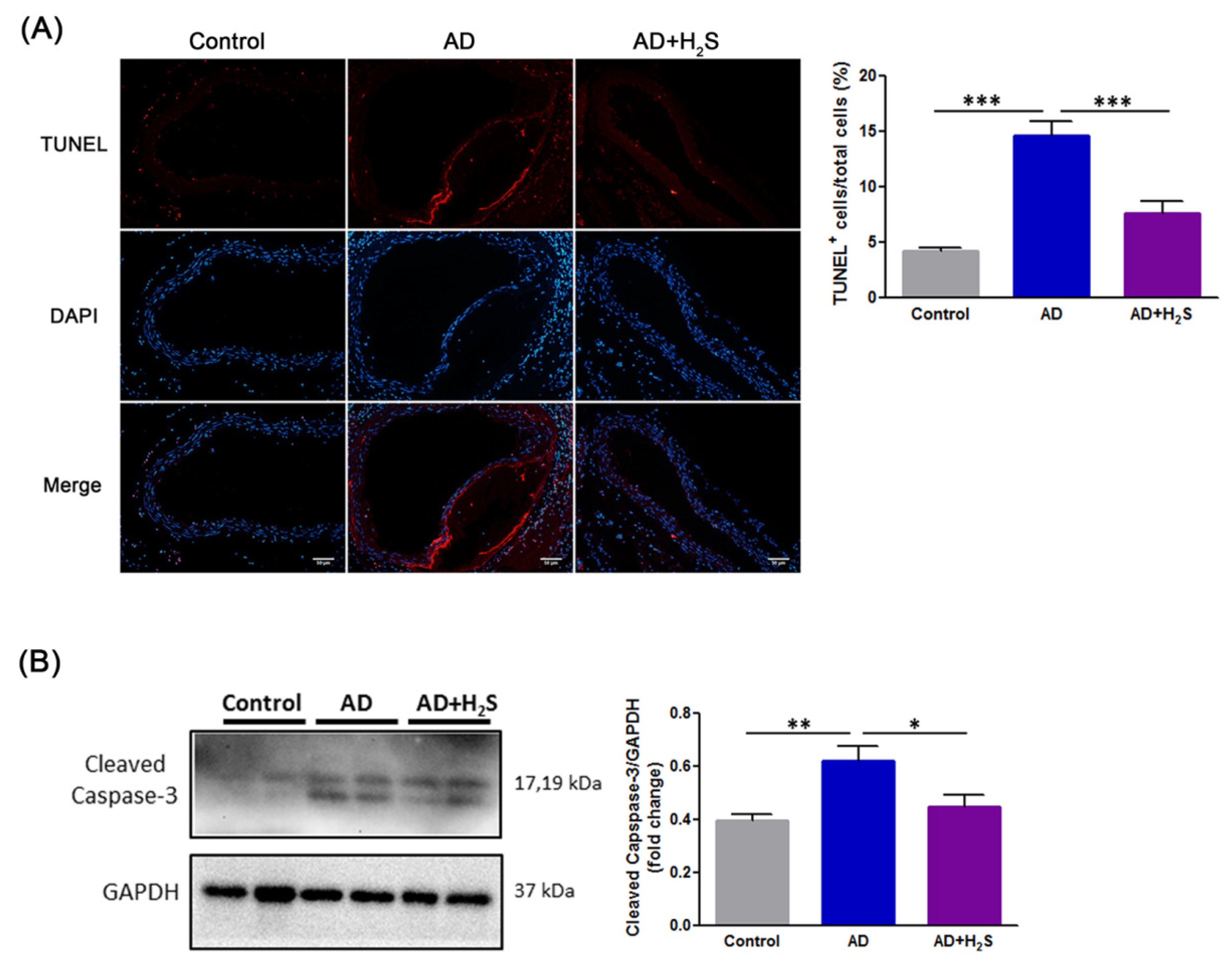
3.2. H2S Mitigates Cell Death of Aortic Dissection
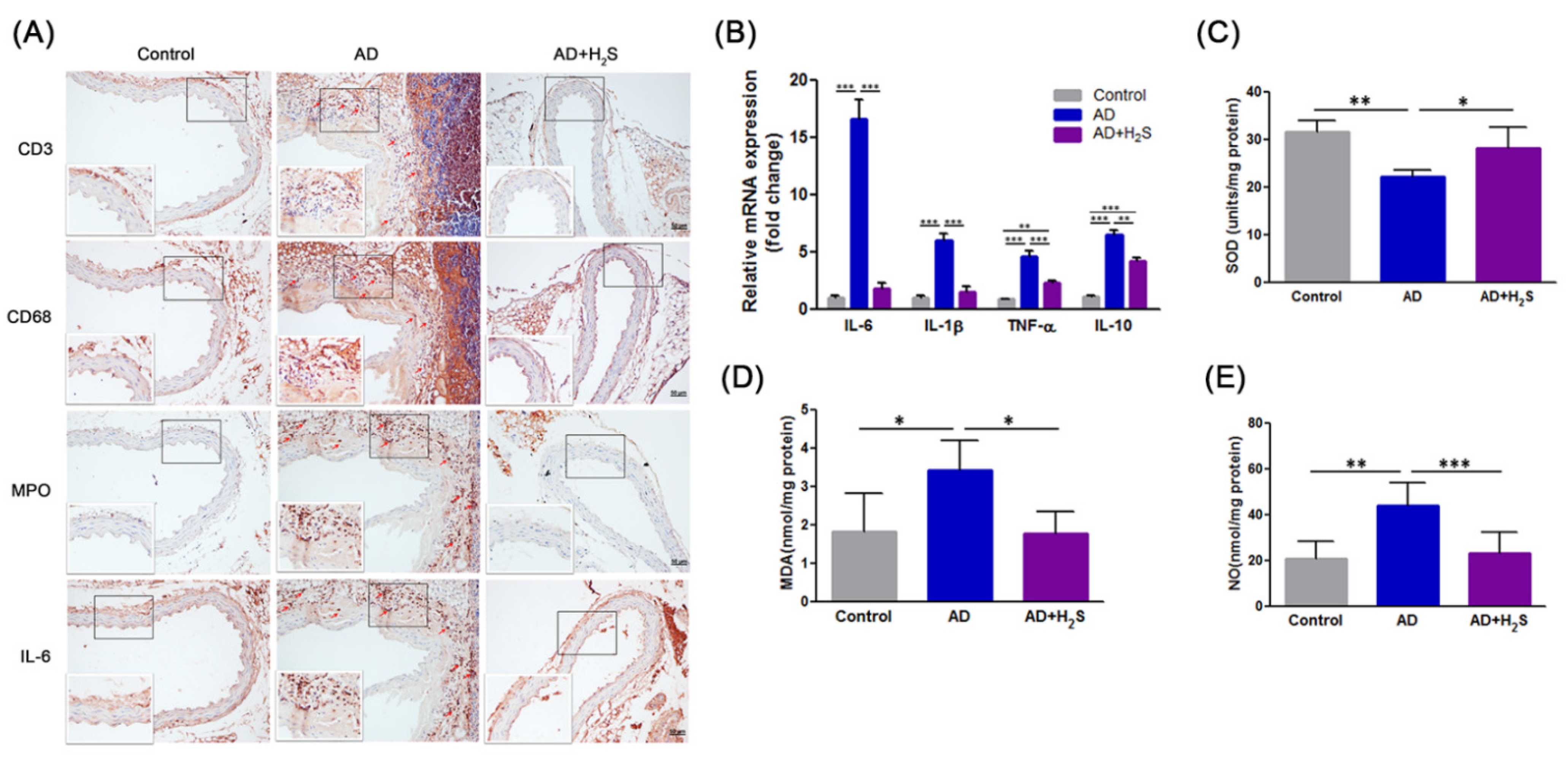
3.3. H2S Inhibits the Accumulation of Inflammatory Cells and Cytokine Profile Expression and Oxidative Stress in AD Mice Aorta
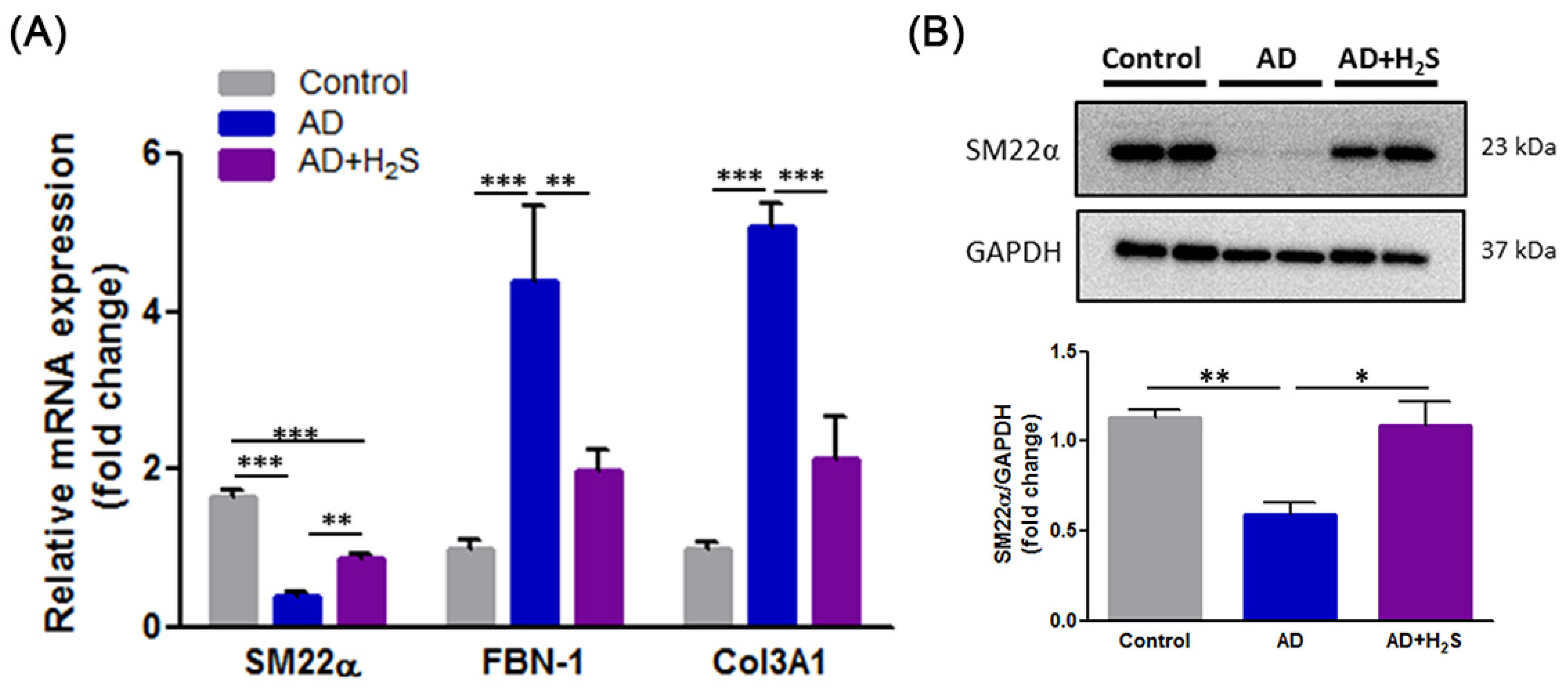
3.4. H2S Reverses the Expression of Synthetic Phenotypic Markers in AD Aorta
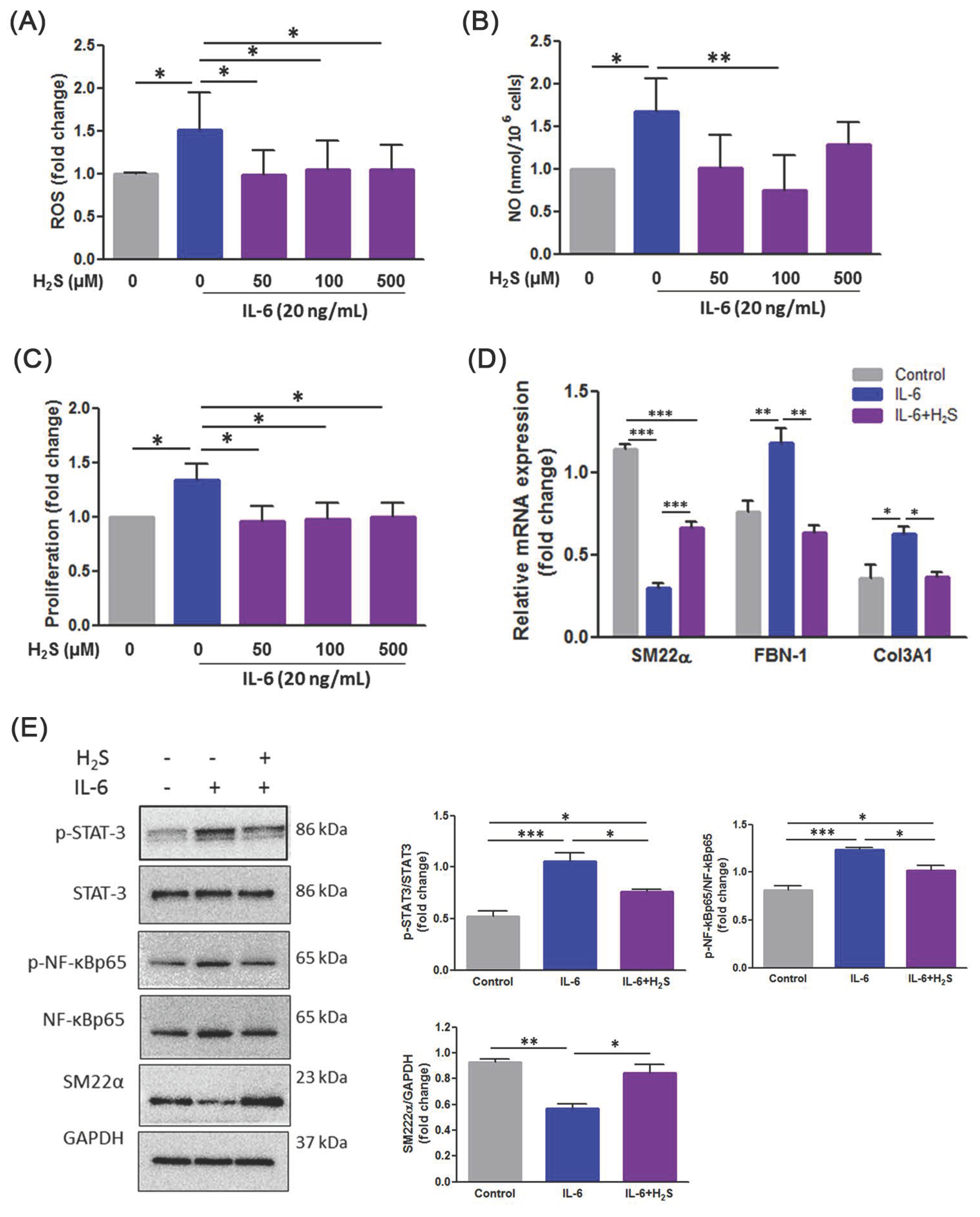
3.5. H2S Ameliorates Oxidative Stress and Phenotypic Switch in IL-6-Indcued VSMCs
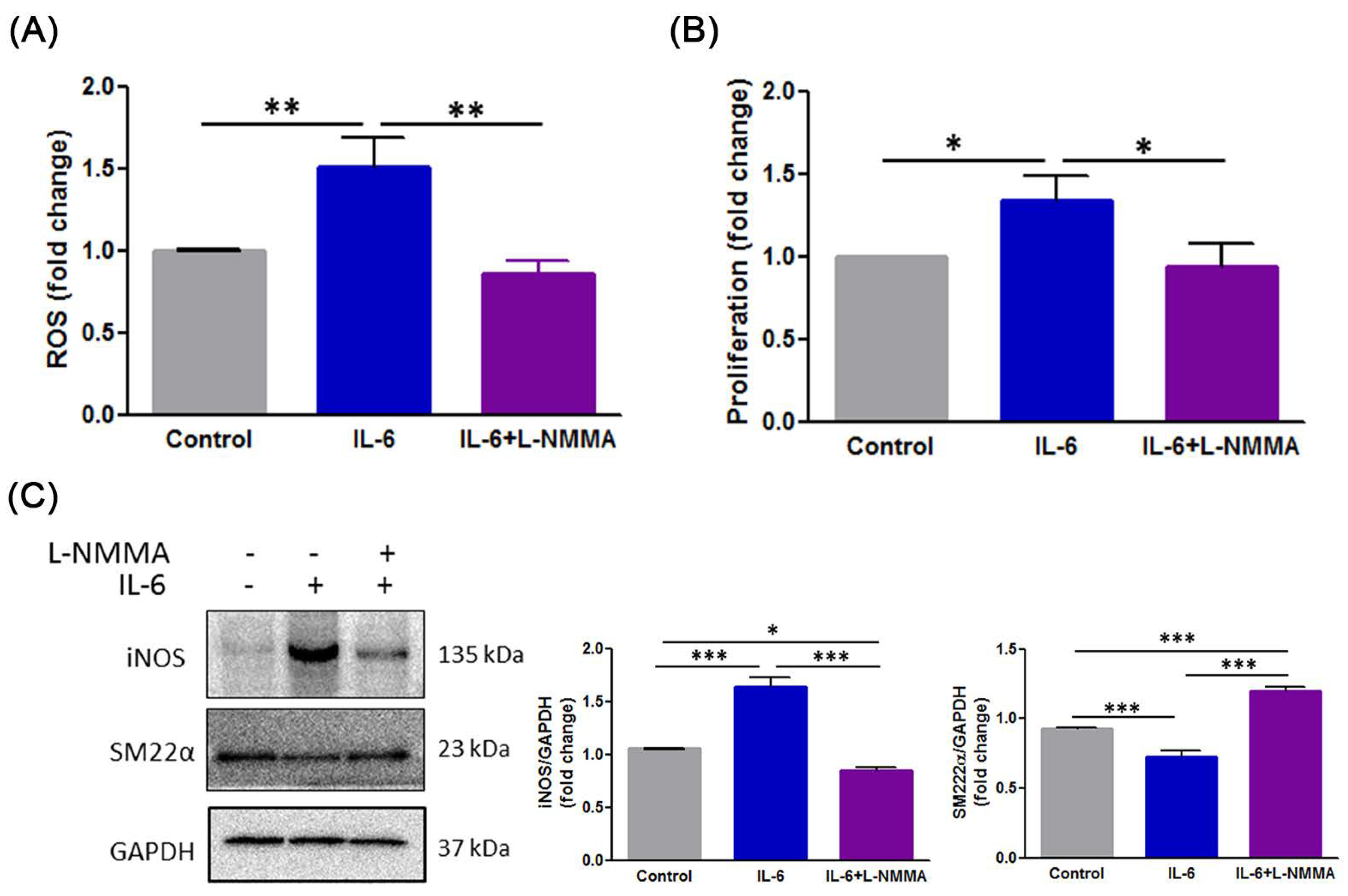
3.6. H2S Modulates IL-6-Induced Phenotype Switch of VSMCs through the ROS-NO-iNOS Pathway
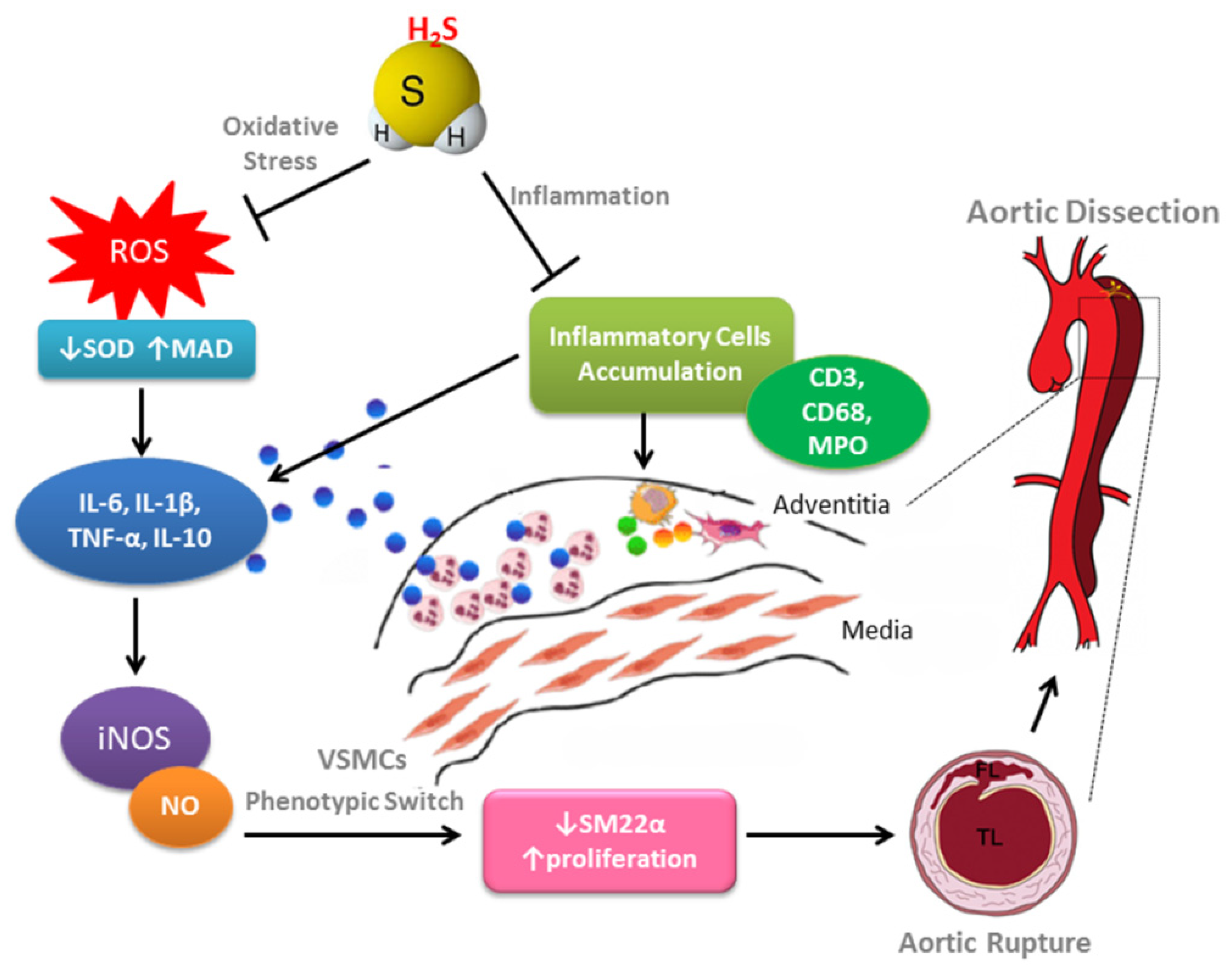
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erbel, R.; Aboyans, V.; Boileau, C.; Bossone, E.; Bartolomeo, R.D.; Eggebrecht, H.; Evangelista, A.; Falk, V.; Frank, H.; Gaemperli, O.; et al. 2014 ESC Guidelines on the diagnosis and treatment of aortic diseases: Document covering acute and chronic aortic diseases of the thoracic and abdominal aorta of the adult. The Task Force for the Diagnosis and Treatment of Aortic Diseases of the European Society of Cardiology (ESC). Eur. Heart J. 2014, 35, 2873–2926. [Google Scholar]
- Chen, L.W.; Wu, X.J.; Lu, L.; Zhang, G.C.; Yang, G.F.; Yang, Z.W.; Dong, Y.; Cao, H.; Chen, Q. Total arch repair for acute type A aortic dissection with 2 modified techniques: Open single-branched stent graft placement and reinforcement of the dissected arch vessel stump with stent graft. Circulation 2011, 123, 2536–2541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiellaro-Rafferty, K.; Weiss, D.; Joseph, G.; Wan, W.; Gleason, R.L.; Taylor, W.R. Catalase overexpression in aortic smooth muscle prevents pathological mechanical changes underlying abdominal aortic aneurysm formation. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H355–H362. [Google Scholar] [CrossRef] [Green Version]
- del Porto, F.; Proietta, M.; Tritapepe, L.; Miraldi, F.; Koverech, A.; Cardelli, P.; Tabacco, F.; de Santis, V.; Vecchione, A.; Mitterhofer, A.P.; et al. Inflammation and immune response in acute aortic dissection. Ann. Med. 2010, 42, 622–629. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Guo, D.C.; Estrera, A.L.; Safi, H.J.; Huynh, T.T.; Yin, Z.; Cao, S.N.; Lin, J.; Kurian, T.; Buja, L.M.; et al. Characterization of the inflammatory and apoptotic cells in the aortas of patients with ascending thoracic aortic aneurysms and dissections. J. Thorac. Cardiovasc. Surg. 2006, 131, 671–678. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Shen, Y.H.; Russell, L.; Coselli, J.S.; LeMaire, S.A. Molecular mechanisms of thoracic aortic dissection. J. Surg. Res. 2013, 184, 907–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhang, J.; Fu, W.; Guo, D.; Jiang, J.; Wang, Y. Association of smooth muscle cell phenotypes with extracellular matrix disorders in thoracic aortic dissection. J. Vasc. Surg. 2012, 56, 1698–1709.e1. [Google Scholar] [CrossRef] [Green Version]
- Clement, M.; Chappell, J.; Raffort, J.; Lareyre, F.; Vandestienne, M.; Taylor, A.L.; Finigan, A.; Harrison, J.; Bennett, M.R.; Bruneval, P.; et al. Vascular Smooth Muscle Cell Plasticity and Autophagy in Dissecting Aortic Aneurysms. Arter. Thromb. Vasc. Biol. 2019, 39, 1149–1159. [Google Scholar] [CrossRef]
- Szabo, C. Gasotransmitters in cancer: From pathophysiology to experimental therapy. Nat. Rev. Drug. Discov. 2016, 15, 185–203. [Google Scholar] [CrossRef] [Green Version]
- Kamoun, P. Endogenous production of hydrogen sulfide in mammals. Amino Acids 2004, 26, 243–254. [Google Scholar] [CrossRef]
- Li, L.; Hsu, A.; Moore, P.K. Actions and interactions of nitric oxide, carbon monoxide and hydrogen sulphide in the cardiovascular system and in inflammation—A tale of three gases! Pharmacol. Ther. 2009, 123, 386–400. [Google Scholar] [CrossRef] [PubMed]
- Shimada, S.; Fukai, M.; Wakayama, K.; Ishikawa, T.; Kobayashi, N.; Kimura, T.; Yamashita, K.; Kamiyama, T.; Shimamura, T.; Taketomi, A.; et al. Hydrogen sulfide augments survival signals in warm ischemia and reperfusion of the mouse liver. Surg. Today 2015, 45, 892–903. [Google Scholar] [CrossRef]
- Kasinath, B.S. Hydrogen sulfide to the rescue in obstructive kidney injury. Kidney Int. 2014, 85, 1255–1258. [Google Scholar] [CrossRef] [Green Version]
- Tokuda, K.; Kida, K.; Marutani, E.; Crimi, E.; Bougaki, M.; Khatri, A.; Kimura, H.; Ichinose, F. Inhaled hydrogen sulfide prevents endotoxin-induced systemic inflammation and improves survival by altering sulfide metabolism in mice. Antioxid. Redox Signal. 2012, 17, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Yang, Z.; Zhang, M.; Dong, Q.; Wang, X.; Lan, A.; Zeng, F.; Chen, P.; Wang, C.; Feng, J. Hydrogen sulfide protects against chemical hypoxia-induced cytotoxicity and inflammation in HaCaT cells through inhibition of ROS/NF-kappaB/COX-2 pathway. PLoS ONE 2011, 6, e21971. [Google Scholar]
- Du, J.; Huang, Y.; Yan, H.; Zhang, Q.; Zhao, M.; Zhu, M.; Liu, J.; Chen, S.X.; Bu, D.; Tang, C.; et al. Hydrogen sulfide suppresses oxidized low-density lipoprotein (ox-LDL)-stimulated monocyte chemoattractant protein 1 generation from macrophages via the nuclear factor kappaB (NF-kappaB) pathway. J. Biol. Chem. 2014, 289, 9741–9753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Dong, J.H.; Teng, X.; Jin, S.; Xue, H.M.; Liu, S.Y.; Guo, Q.; Shen, W.; Ni, X.C.; Wu, Y.M. Hydrogen sulfide improves endothelial dysfunction in hypertension by activating peroxisome proliferator-activated receptor delta/endothelial nitric oxide synthase signaling. J. Hypertens. 2018, 36, 651–665. [Google Scholar] [CrossRef]
- Elrod, J.W.; Calvert, J.W.; Morrison, J.; Doeller, J.E.; Kraus, D.W.; Tao, L.; Jiao, X.; Scalia, R.; Kiss, L.; Szabo, C.; et al. Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial function. Proc. Natl. Acad. Sci. USA 2007, 104, 15560–15565. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhao, X.; Jin, H.; Wei, H.; Li, W.; Bu, D.; Tang, X.; Ren, Y.; Tang, C.; Du, J. Role of hydrogen sulfide in the development of atherosclerotic lesions in apolipoprotein E knockout mice. Arter. Thromb. Vasc. Biol. 2009, 29, 173–179. [Google Scholar] [CrossRef]
- Kondo, K.; Bhushan, S.; King, A.L.; Prabhu, S.D.; Hamid, T.; Koenig, S.; Murohara, T.; Predmore, B.L.; Gojon, G., Sr.; Gojon, G., Jr.; et al. H(2)S protects against pressure overload-induced heart failure via upregulation of endothelial nitric oxide synthase. Circulation 2013, 127, 1116–1127. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhou, J.; Jing, Z.; Xiao, Y.; Sun, Y.; Wu, Y.; Sun, H. Glucocorticoids Regulate the Vascular Remodeling of Aortic Dissection Via the p38 MAPK-HSP27 Pathway Mediated by Soluble TNF-RII. EBioMedicine 2018, 27, 247–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middleton, R.K.; Lloyd, G.M.; Bown, M.J.; Cooper, N.J.; London, N.J.; Sayers, R.D. The pro-inflammatory and chemotactic cytokine microenvironment of the abdominal aortic aneurysm wall: A protein array study. J. Vasc. Surg. 2007, 45, 574–580. [Google Scholar] [CrossRef] [Green Version]
- An, Z.; Qiao, F.; Lu, Q.; Ma, Y.; Liu, Y.; Lu, F.; Xu, Z. Interleukin-6 downregulated vascular smooth muscle cell contractile proteins via ATG4B-mediated autophagy in thoracic aortic dissection. Heart Vessel. 2017, 32, 1523–1535. [Google Scholar] [CrossRef] [PubMed]
- Son, B.K.; Sawaki, D.; Tomida, S.; Fujita, D.; Aizawa, K.; Aoki, H.; Akishita, M.; Manabe, I.; Komuro, I.; Friedman, S.L.; et al. Granulocyte macrophage colony-stimulating factor is required for aortic dissection/intramural haematoma. Nat. Commun. 2015, 6, 6994. [Google Scholar] [CrossRef] [Green Version]
- Anzai, A.; Shimoda, M.; Endo, J.; Kohno, T.; Katsumata, Y.; Matsuhashi, T.; Yamamoto, T.; Ito, K.; Yan, X.; Shirakawa, K.; et al. Adventitial CXCL1/G-CSF expression in response to acute aortic dissection triggers local neutrophil recruitment and activation leading to aortic rupture. Circ. Res. 2015, 116, 612–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Ventura, J.L.; Madrigal-Matute, J.; Martinez-Pinna, R.; Ramos-Mozo, P.; Blanco-Colio, L.M.; Moreno, J.A.; Tarin, C.; Burillo, E.; Fernandez-Garcia, C.E.; Egido, J.; et al. Erythrocytes, leukocytes and platelets as a source of oxidative stress in chronic vascular diseases: Detoxifying mechanisms and potential therapeutic options. Thromb. Haemost. 2012, 108, 435–442. [Google Scholar]
- Liao, M.; Liu, Z.; Bao, J.; Zhao, Z.; Hu, J.; Feng, X.; Feng, R.; Lu, Q.; Mei, Z.; Liu, Y.; et al. A proteomic study of the aortic media in human thoracic aortic dissection: Implication for oxidative stress. J. Thorac. Cardiovasc. Surg. 2008, 136, 65–72, 72.e1–e3. [Google Scholar] [CrossRef] [Green Version]
- Angouras, D.; Sokolis, D.P.; Dosios, T.; Kostomitsopoulos, N.; Boudoulas, H.; Skalkeas, G.; Karayannacos, P.E. Effect of impaired vasa vasorum flow on the structure and mechanics of the thoracic aorta: Implications for the pathogenesis of aortic dissection. Eur. J. Cardiothorac Surg. 2000, 17, 468–473. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Sun, Y.; Wu, Y.; Zhu, J.; Gao, B.; Yan, H.; Zhao, Z.; Zhou, J.; Jing, Z. Downregulation of Talin-1 expression associates with increased proliferation and migration of vascular smooth muscle cells in aortic dissection. BMC Cardiovasc. Disord. 2017, 17, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.L.; Wang, Z.W. The expression and significance of IL-6, IFN-gamma, SM22alpha, and MMP-2 in rat model of aortic dissection. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 560–568. [Google Scholar]
- Dinerman, J.L.; Lowenstein, C.J.; Snyder, S.H. Molecular mechanisms of nitric oxide regulation. Potential relevance to cardiovascular disease. Circ. Res. 1993, 73, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Schackelford, R.E.; Misra, U.K.; Florine-Casteel, K.; Thai, S.F.; Pizzo, S.V.; Adams, D.O. Oxidized low density lipoprotein suppresses activation of NF kappa B in macrophages via a pertussis toxin-sensitive signaling mechanism. J. Biol. Chem. 1995, 270, 3475–3478. [Google Scholar] [CrossRef] [Green Version]
- Kibbe, M.; Billiar, T.; Tzeng, E. Inducible nitric oxide synthase and vascular injury. Cardiovasc. Res. 1999, 43, 650–657. [Google Scholar] [CrossRef]
- Sha, Y.; Marshall, H.E. S-nitrosylation in the regulation of gene transcription. Biochim. Biophys. Acta 2012, 1820, 701–711. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.G.; Fujii, J.; Asahi, M.; Okado, A.; Fujiwara, N.; Taniguchi, N. Roles of purine nucleotides and adenosine in enhancing NOS II gene expression in interleukin-1 beta-stimulated rat vascular smooth muscle cells. Free Radic Res. 1997, 26, 409–418. [Google Scholar] [CrossRef]
- Ikeda, U.; Kurosaki, K.; Ohya, K.; Shimada, K. Adenosine stimulates nitric oxide synthesis in vascular smooth muscle cells. Cardiovasc. Res. 1997, 35, 168–174. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Brahmbhatt, S.; Gupta, A.; Sharma, A.C. Duration of streptozotocin-induced diabetes differentially affects p38-mitogen-activated protein kinase (MAPK) phosphorylation in renal and vascular dysfunction. Cardiovasc. Diabetol. 2005, 4, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagareddy, P.R.; Xia, Z.; McNeill, J.H.; MacLeod, K.M. Increased expression of iNOS is associated with endothelial dysfunction and impaired pressor responsiveness in streptozotocin-induced diabetes. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H2144–H2152. [Google Scholar] [CrossRef] [PubMed]
- Johanning, J.M.; Franklin, D.P.; Han, D.C.; Carey, D.J.; Elmore, J.R. Inhibition of inducible nitric oxide synthase limits nitric oxide production and experimental aneurysm expansion. J. Vasc. Surg. 2001, 33, 579–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, H.-Y.; Hsu, H.-L.; Li, C.-H.; Li, S.-J.; Lin, S.-J.; Shih, C.-M.; Shih, C.-C. Hydrogen Sulfide Attenuates Aortic Remodeling in Aortic Dissection Associating with Moderated Inflammation and Oxidative Stress through a NO-Dependent Pathway. Antioxidants 2021, 10, 682. https://doi.org/10.3390/antiox10050682
Lu H-Y, Hsu H-L, Li C-H, Li S-J, Lin S-J, Shih C-M, Shih C-C. Hydrogen Sulfide Attenuates Aortic Remodeling in Aortic Dissection Associating with Moderated Inflammation and Oxidative Stress through a NO-Dependent Pathway. Antioxidants. 2021; 10(5):682. https://doi.org/10.3390/antiox10050682
Chicago/Turabian StyleLu, Hsin-Ying, Hung-Lung Hsu, Chih-Han Li, Shao-Jung Li, Shing-Jong Lin, Chun-Ming Shih, and Chun-Che Shih. 2021. "Hydrogen Sulfide Attenuates Aortic Remodeling in Aortic Dissection Associating with Moderated Inflammation and Oxidative Stress through a NO-Dependent Pathway" Antioxidants 10, no. 5: 682. https://doi.org/10.3390/antiox10050682