
Current Status of Trypanosoma grosi and Babesia microti in Small Mammals in the Republic of Korea

Division of Vectors and Parasitic Diseases, Korea Disease Control and Prevention Agency, 187 Osongsaenmyeong 2-ro, Osong-eup, Heungdeok-gu, Cheongju 28159, Republic of Korea
*
Author to whom correspondence should be addressed.
Animals 2024, 14(7), 989; https://doi.org/10.3390/ani14070989
Submission received: 4 February 2024
/
Revised: 13 March 2024
/
Accepted: 21 March 2024
/
Published: 22 March 2024
(This article belongs to the Special Issue Parasites and Parasitic Diseases in Small Animals)
Abstract
:Simple Summary
Parasitic protozoa are a common cause of vector-borne disease outbreaks and have been identified in different groups of wild animal species. Small mammals, such as rodents and shrews, play an important role in the transmission and maintenance of parasites. To understand the risk of parasitic protozoa, it is essential to have a good understanding of the status of transmitting vectors. This study aimed to investigate the diversity and current status of parasitic protozoa, including Trypanosoma, Babesia, and Theileria, from small mammals in the Republic of Korea. We found that the prevalence of Trypanosoma grosi was 23.9% (79/331) and Babesia microti was 10% (33/331), while Theileria was not detected in small mammals. These results can be used to raise awareness of parasite infection in the Republic of Korea.
Abstract
Small mammals, such as rodents and shrews, are natural reservoir hosts of zoonotic diseases, including parasitic protozoa. To assess the risk of rodent-borne parasitic protozoa in the Republic of Korea (ROK), this study investigated the status of parasitic protozoa, namely Trypanosoma, Babesia, and Theileria, in small mammals. In total, 331 blood samples from small mammals were analyzed for parasites using PCR and sequenced. Samples were positive for Trypanosoma grosi (23.9%; n = 79) and Babesia microti (10%; n = 33) but not Theileria. Small mammals from Seogwipo-si showed the highest infection rate of T. grosi (48.4%), while the highest B. microti infection rate was observed in those from Gangneung-si (25.6%). Sequence data revealed T. grosi to be of the AKHA strain. Phylogenetic analysis of B. microti revealed the US and Kobe genotypes. B. microti US-type–infected small mammals were detected throughout the country, but the Kobe type was only detected in Seogwipo-si. To our knowledge, this is the first nationwide survey that confirmed T. grosi and B. microti infections at the species level in small mammals in the ROK and identified the Kobe type of B. microti. These results provide valuable information for further molecular epidemiological studies on these parasites.
1. Introduction
Small mammals, such as rodents and shrews, are well-known hosts and reservoirs of zoonotic diseases that pose a crucial threat to human health. Over 2000 species of rodents are distributed worldwide and live closely with humans [1,2]. Of these, approximately 200 species are reservoirs of more than 60 zoonotic diseases caused by viruses, bacteria, and parasites [3,4]. Yersinia pestis, Salmonella, and Hantavirus are among the most important pathogens affecting public health [5,6].
Zoonotic parasites are a common cause of vector-borne disease outbreaks and have been identified in different groups of wild animal species. Rodents play an important role in the transmission and maintenance of zoonotic parasites [6,7]. Most parasites in the blood and tissues of rodents can be transmitted to humans through contaminated food, water, rodent urine and feces, or via ectoparasites [8]. Therefore, it is necessary to investigate zoonotic parasite infections in small mammals to reduce exposure risk and predict future trends in pathogen prevalence and distribution according to seasonal and environmental changes.
Several zoonotic parasitic protozoa, such as Trypanosoma and Babesia, which cause trypanosomiasis and babesiosis, respectively, have gained importance as infectious agents. Trypanosoma is a parasitic hemoflagellate protozoan belonging to the Trypanosomatidae family that can infect animals and humans [9,10]. Certain species of this parasite may be transmitted by blood-feeding arthropods such as Triatoma sp. (kissing bug) and Glossina sp. (tsetse fly). Trypanosoma cause serious diseases in humans, such as Chagas disease (also known as American trypanosomiasis), caused by Trypanosoma cruzi, and African trypanosomiasis, caused by the Trypanosoma brucei complex [11].
Piroplasms are parasitic protozoa belonging to the genera Babesia and Theileria, which are the causative agents of babesiosis and theileriosis, respectively. Both intraerythrocytic protozoan parasites are transmitted by ticks [12,13] and are recognized for their important economic effects on the livestock industry and human health. Zoonotic babesiosis, mainly caused by Babesia microti, has recently posed a serious public health risk worldwide, in contrast to theileriosis, which has no zoonotic potential [14,15]. Approximately 100 Babesia species are known to infect animals and humans. The common clinical symptoms of babesiosis include fever, chills, fatigue, and headache [16,17]. The incidence of babesiosis has increased between 2011 and 2019 in the United States [18]. Cases of Babesia infections have been reported in various countries worldwide, including China, Germany, Canada, Australia, Japan, and the Republic of Korea (ROK) [19,20].
Therefore, given the importance of these three parasitic protozoa within the livestock industry and/or Public Health and as they can also be found in wild small mammals as reservoir hosts, we used molecular methods to investigate the diversity and current status of parasitic protozoa, including Trypanosoma, Babesia, and Theileria, in small mammals in the ROK.
2. Materials and Methods
2.1. Ethical Approval
All animal experiments were performed according to the guidelines for ethical conduct in the care and use of animals and approved by the Institutional Animal Care and Use Committee of the Korea Disease Control and Prevention Agency (approval number: KDCA-102-22). Permission to capture small mammals was obtained from each site in accordance with the Wildlife Protection and Management Act of the ROK.
2.2. Small Mammal Sampling
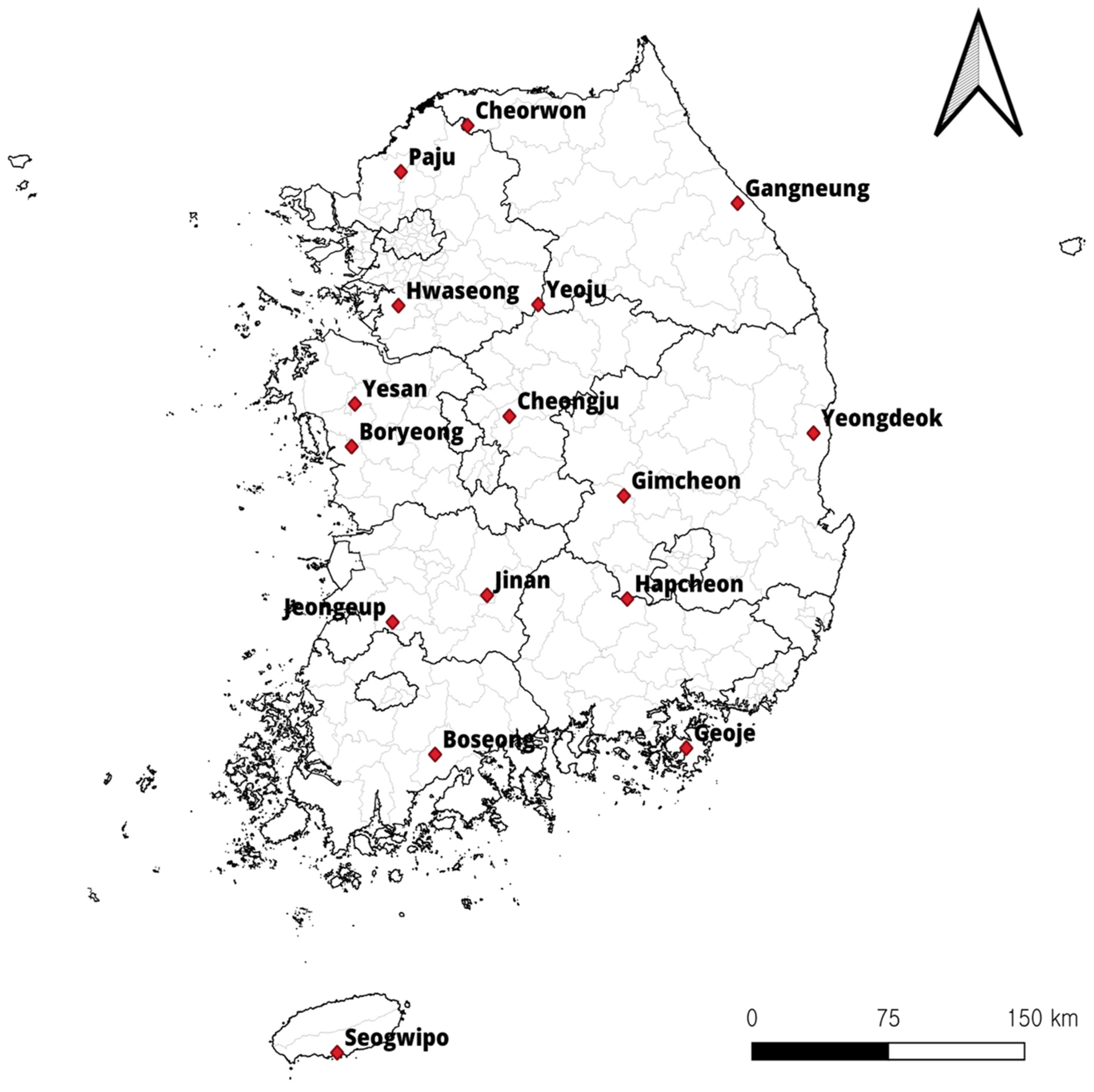
This study was conducted at 16 sites in the ROK in 2021 (Figure 1). Trapping was performed in the spring (March and April) and autumn (October and November). Small mammals were captured using Sherman folding live traps (BioQuip Products, Rancho Dominguez, CA, USA) baited with pieces of cheese crackers set in various habitats, including rice paddy fields, dry paddy fields, waterways, meadows, and reservoirs. Live small mammals were transferred to the laboratory and euthanized with carbon dioxide. After the identification of small mammals [2], blood was drawn from the heart using syringes, preserved in EDTA vacutainer tubes (BD, Franklin Lakes, NJ, USA), and stored at −20 °C until further use.
2.3. Molecular Detection of Parasites in Small Mammals
Genomic DNA was extracted from 331 small mammal blood samples (100 μL each) using the QIAamp DNA Blood Mini Kit, following the manufacturer’s protocol (Qiagen, Hilden, Germany). To detect parasite infection, we used the extracted DNA and amplified the ITS1 gene of Trypanosoma spp. using TRYP1R (5′-GGAAGCCAAGTCATCCATCG-3′) and TRYP1S (5′-CGTCCCTGCCATTTGTACACAC-′3) primer sets targeting for ~623 bp fragments [21]. PCR was performed with 20 μL using an AccuPower PCR PreMix (Bioneer, Daejeon, Republic of Korea). Each PCR mixture was composed of 1 μL of each oligonucleotide primer (10 pmol/μL), 3 μL of genomic DNA as template, and 15 μL of distilled water. The conditions were initial denaturation at 95 °C for 5 min, followed by 35 cycles of 30 s at 95 °C, 1 min at 55 °C, and 1 min at 72 °C, with a final extension at 72 °C for 10 min in a C1000TM Thermal Cycler (Bio-Rad, Hercules, CA, USA). Thus, we amplified the 18S rRNA gene of Babesia and Theileria to obtain a 561 bp fragment using the commercial AccuPower Babesia & Theileria PCR Kit (Bioneer, Daejeon, Republic of Korea) according to the manufacturer’s instructions. Briefly, the lyophilized premix and primer comprised 3 μL genomic DNA as template, and 17 μL distilled water. The PCR reaction mixture was subjected to thermal cycling at 95 °C for 5 min, followed by 40 cycles of 20 s at 95 °C, and 50 s at 59 °C, with a final extension at 72 °C for 5 min. The genomic DNA of B. microti and T. cruzi, obtained from American Type Culture Collection (ATCC, Manassas, VA, USA), and Theileria, obtained from Bioneer (Daejeon, Republic of Korea), were used as positive controls. A suitable positive control and negative control were included in each amplification reaction. Amplification products were analyzed using a QIAxcel capillary electrophoresis system (Qiagen). The DNA extraction, PCR amplification, and automated electrophoresis were performed in separate rooms to prevent contamination.
2.4. Sequencing and Phylogenetic Analysis
Positive PCR products were purified using QIAquick PCR purification kits (Qiagen) and sequenced using Sanger sequencing and an ABI PRISM 3730xl analyzer (Life Technologies, Carlsbad, CA, USA). The nucleotide sequences were compared with reference sequences obtained from GenBank using nucleotide BLAST (National Center for Biotechnology Information, NCBI). All sequences used in phylogenetic analyses were downloaded from GenBank, available through the NCBI. A phylogenetic tree was constructed using the neighbor-joining method and p-distance model in MEGA X (Pennsylvania State University, State College, PA, USA). The sequences obtained in this study were submitted to GenBank (accession numbers: OP804252, OP804253, OP804254, OP297200, and OP297201).
2.5. Statistical Analysis
Statistical analyses were performed using IBM SPSS Statistics software version 26 (IBM, Armonk, NY, USA). Pearson’s chi-square test or Fisher’s exact test was used to examine the association between parasitic infections in small mammals and related factors, including small mammal species, seasonal habitat type, and trapping site. p < 0.05 was considered significant.
3. Results
3.1. Prevalence of Parasitic Protozoa in Small Mammals in the ROK
Over the two trapping seasons in 2021, 331 small mammals belonging to eight genera and ten species were collected from 16 sites in the ROK. The results showed that 14 sites harbored parasitic protozoa, T. grosi or B. microti, infecting small mammals, with the exceptions being Boryeong-si and Cheongju-si (Table 1); Theileria was not detected in any of the samples. The small mammals collected from Seogwipo-si had the highest T. grosi infection rate at 48.4% (15/31). The results also showed that 10 sites harbored B. microti, with the highest B. microti infection rate being 32.6% (14/43) in Gangneung-si. The infection rates in Seogwipo-si and Gangneung-si were significantly higher than those in other sites (p < 0.05). Also, our results showed that one genotype of B. microti was confirmed in each site. Among positive samples of B. microti, the US type was found to be the dominant genotype in the ROK; only the B. microti Kobe type was detected in Seogwipo-si, which is the first confirmation from small mammals in the ROK (Table 1).
3.2. Prevalence and Phylogenetic Analysis of T. grosi and B. microti in Small Mammals
Three species, Apodemus agrarius (striped field mouse), Crocidura sp., and Apodemus peninsulae (Korean field mouse), were confirmed to be infected with parasitic protozoa. A. agrarius was highly dominant, accounting for 87.6% (290/331) of trapped small mammals, and co-infected with T. grosi and B. microti in six individuals (Table 2).
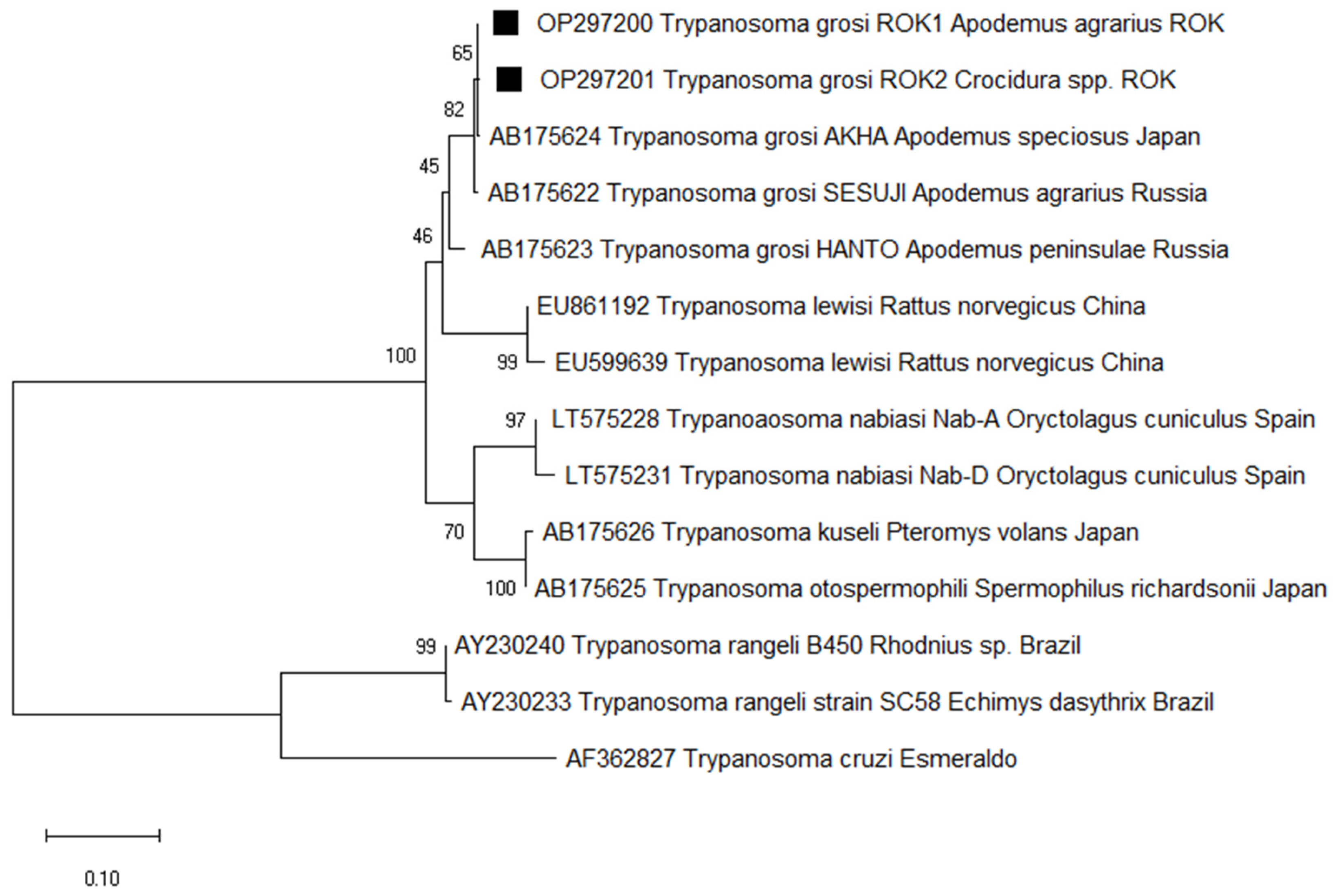
To detect Trypanosoma in small mammals, we target the ITS1 gene of the protozoa. Of the 331 samples, 79 (23.9%) were positive and sequencing revealed 98.8% identity with T. grosi (AB175624), identified in two species of small mammals: A. agrarius (26.5%) and Crocidura sp. (8.7%). Of the 79 sequences identified as T. grosi, two representative sequences were selected without duplicates of sequences and host source. Therefore, the two T. grosi ITS1 sequences were deposited in GenBank under the accession numbers OP297200 and OP297201. Phylogenetic analysis revealed that OP297200 (from A. agrarius) and OP297201 (from Crocidura sp.) are closely related with the previously reported T. grosi AKHA strain sequence (AB175624), which was isolated from A. speciosus in Japan (Figure 2).
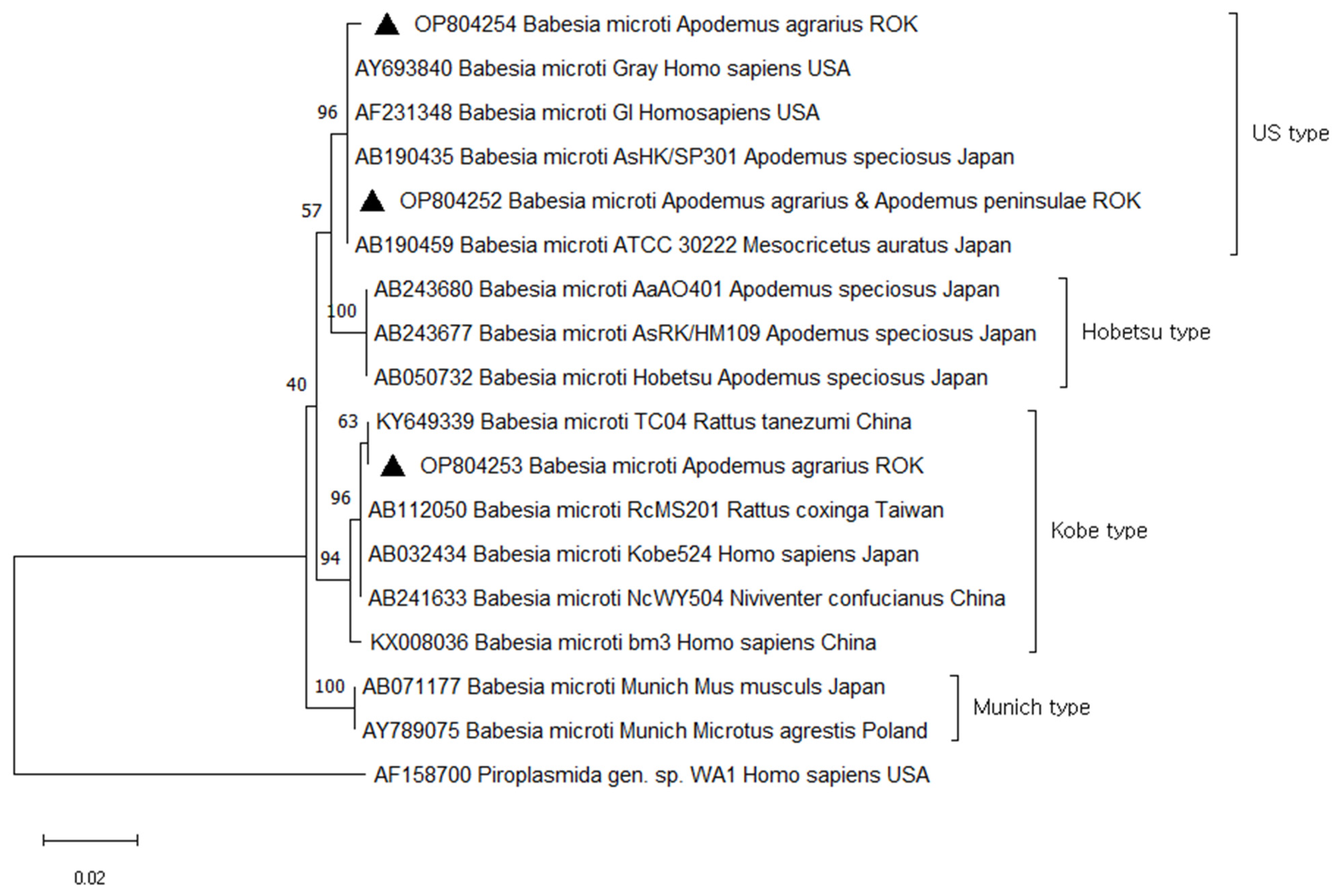
A total of 33 (10.0%) samples were positive and sequencing revealed 100.0% similarity to B. microti, found in two species of small mammals: A. agrarius (11%) and A. peninsulae (50%). Of the 33 sequences identified as B. microti, three representative sequences were selected without duplicates of sequences and host source. The three B. microti 18S rRNA sequences were deposited in GenBank under accession numbers OP804252, OP804253, and OP804254. Phylogenetic analysis revealed that OP804252 (from A. agrarius and A. peninsulae) OP804254 (from A. agrarius) were closely related to B. microti US type (AY693840, AF231348, AB190435, and AB190459). These clusters contained B. microti US type that have been isolated from small mammals or humans in different regions, including USA (AY693840 and AF231348 from human) and Japan (AB190435 from A. speciosus and AB190459 from M. auratus). OP804253 (from A. agrarius) belonged to the B. microti Kobe type (KY649339 from R. tanezumi, AB112050 from R. coxinga, AB032434 from Human, AB241633 from N. confucianus, and KX008036 from human) (Figure 3).
3.3. Relationships between Season, Ecological Habitat, and T. grosi and B. microti Infections
In ecological habitats, the prevalence of T. grosi in small mammals from reservoirs was significantly lower than that in small mammals from other habitats (rice paddy fields, dry paddy fields, watery ways, and meadows; p = 0.006). The seasonal T. grosi positivity rate was higher in spring (26.4%; 46/174) than in autumn (21.0%; 33/157); however, the difference was not significant. The infection rate of T. grosi in A. agrarius was significantly higher than that in other species (p = 0.0014). The infection rates of B. microti per ecological habitat, season, and small mammal species are shown in Table 3. The seasonal B. microti positivity rate was 9.8% (17/174) for small mammals in spring and 10.2% (16/157) in autumn. There were no significant differences between seasons, small mammal species, and ecological habitats with respect to B. microti infection. However, B. microti DNA from small mammals was consistently detected throughout the study period in all habitats (Table 3).
4. Discussion
This study was a nationwide investigation of parasitic protozoa, including Trypanosoma, Babesia, and Theileria, in small mammals from 16 sites across the ROK and demonstrates a wide prevalence of parasitic protozoa. Over two trapping seasons in 2021, 331 small mammals belonging to eight genera and ten species were collected from 16 sites. Blood samples from small mammals were analyzed for parasites using PCR and positive samples were sequenced. Samples were positive for Trypanosoma grosi (23.9%) and Babesia microti (10.0%) but not Theileria. These results suggest that the diversity of parasitic protozoa in the ROK is high and even new species can be involved in the infections.
Trypanosoma is a flagellate blood parasite found in every vertebrate class [22,23]. Over 500 species of Trypanosoma have been recorded worldwide [24,25,26]. Trypanosomes can be categorized into two groups based on their transmission route from the vector to the host [27]. The first group, known as salivarian trypanosomes, includes T. brucei, the causative agent of human African trypanosomiasis; this species develops in the midgut of the vector, migrates to the salivary glands or proboscis, and is transmitted to the host through the vector’s saliva during the biting process. The second group, stercorarian trypanosomes, includes T. cruzi, the causative agent of Chagas disease. Stercorarian trypanosomes develop in the hindgut of the vector and are transmitted to the host either through the ingestion of vectors, such as fleas, or the contamination of bite wounds with vector feces [26,27]. A total of 44 Trypanosoma species have been identified in 144 rodent species, the majority of which belong to the stercorarian group [11,27]. Most of stercorarian trypanosomes are considered nonpathogenic, except for T. cruzi. To date, there has been no investigation of wild small mammals infected with Trypanosoma in the ROK, resulting in a critical knowledge gap.
Trypanosoma grosi belongs to the nonpathogenic stercorarian group [23,26] and has been detected in species of Apodemus from several countries, including Russia (A. sylvaticus), France (A. sylvaticus), Japan (A. speciosus), and China (A. agrarius) [26,28,29,30]. To the best of our knowledge, T. grosi has not yet been reported in small mammals in the ROK. Our results revealed that there were two species of small mammals (A. agrarius and Crocidura sp.) infected with T. grosi in the ROK, with a high infection rate of 23.9% compared to other protozoa such as Babesia and Theileria. This is also the first confirmed report of co-infection of B. microti and T. grosi in small mammals at the molecular level. To date, there has been no evidence of human infection with T. grosi, however, several cases have been reported of humans infected with T. lewisi or T. lewisi-like (T. grosi-involved group) [31,32,33]. Although T. grosi is harmless in humans, further investigation to maximize the understanding of these diseases should be considered. This study may be helpful in future molecular epidemiological studies of T. grosi in the ROK.
Babesia and Theileria are parasitic protozoa, mainly transmitted by ticks, that can infect a variety of domestic and wild animals. The prevalence of Babesia in small mammals was as high as 20.8% (32/154) in the ROK in 2001 [34]. However, there was a low infection rate of 0.52% (3/578) in 2008, reported for two districts (Gangwon-do and Gyeonggi-do) [35]. Both studies only detected the B. microti US type in small mammals. Compared to previous studies, our study showed that the infection rate of Babesia in small mammals in the ROK decreased from 20.8% in 2001 to 10.0% in 2021. Additionally, previous studies only confirmed the B. microti US type, but we used molecular methods to target Babesia 18S rRNA from small mammals in the ROK to identify two genetic groups of B. microti: the US and Kobe types. B. microti US-type–infected small mammals were distributed across various sites of the ROK, whereas B. microti Kobe-type–infected small mammals were detected only in Seogwipo-si. However, molecular evidence of Babesia genera in ticks has already been reported in the ROK, such as B. duncani, B. venatorum, B. divergens, and B. microti US type [36,37,38,39]. According to a recent report, the US type of B. microti was detected in ticks from small mammals in the ROK in 2017, while the Kobe type was only detected in ticks from the southern region of the ROK (Goheung-gun and Jeju island) [40]. This result is similar to that of our study, which showed that the Kobe type of B. microti was only detected in small mammals collected from Seogwipo-si, Jeju island. In the ROK, the first report of human babesiosis was reported in 1988. Around 16 cases of human babesiosis have been recorded since, most of which were imported from other countries [41,42]. Two cases were known to be locally infected with Babesia sp. KO-1 and B. motasi-like [37,43]. To date, these studies have revealed that there have been no cases of human infection by vectors transmitting B. microti in the ROK. Therefore, this study reinforces the importance of further studies addressing changes in Babesia microti genotypes in the study area.
Some species of Theileria such as T. annulata and T. parva are pathogenic to livestock and cause high rates of mortality in sheep and cattle [44]. Other Theileria spp. are thought to be less virulent, probably because of evolutionary interactions between Theileria and the host [45]. Recently, several studies have investigated the presence of Theileria spp. in various wild animals, including rodents. For example, Theileria peramelis was identified in Rattus (black rat) and Perameles nasuta (long-nosed bandicoots) in Australia [46]. Theileria sp. were successfully amplified from the rodents Le. Striatus and Praomys sp. in Gabon [47]. Those findings have not been confirmed for the zoonotic potential of Theileria. Therefore, further research is needed to investigate the potential consequences of infection in wildlife, especially the effect of pathogens spreading to native wildlife.
Our study has a few limitations: (1) we did not double-check positivity of T. grosi and/or B. microti in small mammals through microscopic examination. The difficulty with identifying species-level parasitic protozoa by microscopy has been described in several studies. Therefore, we used molecular methods to investigate the diversity and current status of parasitic protozoa at the species level in the small mammals. (2) only blood samples from small mammals were used to investigate the prevalence of parasitic protozoa. Further studies including the diversity and rate of parasitic infection from ectoparasites (such as fleas and ticks) in small mammals should be conducted to better understand these parasitic protozoa circulating in small mammals.
In this study, 33 of 331 samples (10.0%) were positive for B. microti in two small mammal species, A. agrarius and A. peninsula. These two small mammal species were confirmed to play an important role as reservoirs of B. microti, especially because A. agrarius is abundant and widely distributed across the country [48]. Although no statistical differences were observed between B. microti infection and season or habitat, B. microti DNA in small mammals was consistently detected over the study period and sites. Therefore, our findings suggest that small mammals infected with T. grosi and B. microti are widely and non-seasonally distributed throughout the ROK; however, there is no detection of Theileria. Additionally, this is the first study to detect B. microti Kobe type in small mammals in the ROK. These results provide useful information for further molecular epidemiological studies on parasites, especially those on T. grosi and B. microti transmission to humans or animals via vectors.
5. Conclusions
This study aimed to investigate the diversity and status of parasitic protozoa, including Trypanosoma grosi, Babesia microti, and Theileria, in small mammals in the ROK. Here, we present the rate of parasitic protozoa infection in small mammals in the study area. T. grosi was detected in small mammals for the first time. Furthermore, B. microti US-type–infected small mammals were detected throughout the country, but the Kobe type was only detected in Seogwipo-si. These results provide valuable information for further molecular epidemiological studies on these parasites.
Author Contributions
Conceptualization, H.-I.S.; methodology, H.J.K. and B.H.; writing original draft preparation, H.J.K.; writing review and editing, J.-W.J., H.-I.L. and H.-I.S. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by the Korea Disease Control and Prevention Agency (grant number: 6331-311).
Institutional Review Board Statement
All animal experiments were performed according to the guidelines for ethical conduct in the care and use of animals and were approved by the Institutional Animal Care and Use Committee of the Korea Disease Control and Prevention Agency (approval number: KDCA-102-22).
Informed Consent Statement
Not applicable.
Data Availability Statement
The sequence data generated in the current study are available in the GenBank repository under the accession numbers OP804252, OP804253, OP804254, OP297200, and OP297201. The datasets used and/or analyzed during the present study are available upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Capizzi, D.; Bertolino, S.; Mortelliti, A. Rating the rat: Global patterns and research priorities in impacts and management of rodent pests. Mamm. Rev. 2014, 44, 148–162. [Google Scholar] [CrossRef]
- Han, B.A.; Schmidt, J.P.; Bowden, S.E.; Drake, J.M. Rodent reservoirs of future zoonotic diseases. Proc. Natl. Acad. Sci. USA 2015, 112, 7039–7044. [Google Scholar] [CrossRef] [PubMed]
- Morand, S.; Jittapalapong, S.; Kosoy, M. Rodents as hosts of infectious diseases: Biological and ecological characteristics. Vector Borne Zoonotic Dis. 2015, 15, 1. [Google Scholar] [CrossRef]
- Jahan, N.A.; Lindsey, L.L.; Larsen, P.A. The role of peridomestic rodents as reservoirs for zoonotic foodborne pathogens. Vector-Borne Zoonotic Dis. 2021, 21, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Shimi, A.; Keyhani, M.; Hedayati, K. Studies on salmonellosis in the house mouse, Mus musculus. Lab. Anim. 1979, 13, 33–34. [Google Scholar] [CrossRef] [PubMed]
- Meerburg, B.G. Rodents are a risk factor for the spreading of pathogens on farms. Vet. Microbiol. 2010, 142, 464–465. [Google Scholar] [CrossRef]
- Dahmana, H.; Granjon, L.; Diagne, C.; Davoust, B.; Fenollar, F.; Mediannikov, O. Rodents as hosts of pathogens and related zoonotic disease risk. Pathogens 2020, 9, 202. [Google Scholar] [CrossRef] [PubMed]
- Rabiee, M.H.; Mahmoudi, A.; Siahsarvie, R.; Kryštufek, B.; Mostafavi, E. Rodent-borne diseases and their public health importance in Iran. PLoS Negl. Trop. Dis. 2018, 12, e0006256. [Google Scholar] [CrossRef] [PubMed]
- Seifollahi, Z.; Sarkari, B.; Motazedian, M.H.; Asgari, Q.; Ranjbar, M.J.; Abdolahi Khabisi, S. Protozoan parasites of rodents and their zoonotic significance in Boyer-Ahmad District, Southwestern Iran. Vet. Med. Int. 2016, 2016, 3263868. [Google Scholar] [CrossRef]
- Tijjani, M.; Majid, R.A.; Abdullahi, S.A.; Unyah, N.Z. Detection of rodent-borne parasitic pathogens of wild rats in Serdang, Selangor, Malaysia: A potential threat to human health. Int. J. Parasitol. Parasit. Wildl. 2020, 11, 174–182. [Google Scholar] [CrossRef]
- Pumhom, P.; Pognon, D.; Yangtara, S.; Thaprathorn, N.; Milocco, C.; Douangboupha, B.; Herder, S.; Chaval, Y.; Morand, S.; Jittapalapong, S.; et al. Molecular prevalence of Trypanosoma spp. in wild rodents of Southeast Asia: Influence of human settlement habitat. Epidemiol. Infect. 2014, 142, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Bishop, R.; Musoke, A.; Morzaria, S.; Gardner, M.; Nene, V. Theileria: Intracellular protozoan parasites of wild and domestic ruminants transmitted by ixodid ticks. Parasitology 2004, 129, S271–S283. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J. Human babesiosis. Int. J. Parasitol. 2019, 49, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Uilenberg, G. International collaborative research: Significance of tick-borne hemoparasitic diseases to world animal health. Vet. Parasitol. 1995, 57, 19–41. [Google Scholar] [CrossRef]
- Duh, D.; Punda-Polić, V.; Trilar, T.; Avsic-Zupanc, T. Molecular detection of Theileria sp. in ticks and naturally infected sheep. Vet. Parasitol. 2008, 151, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Homer, M.J.; Aguilar-Delfin, I.; Telford, S.R., 3rd; Krause, P.J.; Persing, D.H. Babesiosis. Clin. Microbiol. Rev. 2000, 13, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J. Babesiosis diagnosis and treatment. Vector Borne Zoonotic Dis. 2003, 3, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Swanson, M.; Pickrel, A.; Williamson, J.; Montgomery, S. Trends in Reported Babesiosis Cases—United States, 2011–2019. MMWR Morb. Mortal Wkly. Rep. 2023, 72, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Young, K.M.; Corrin, T.; Wilhelm, B.; Uhland, C.; Greig, J.; Mascarenhas, M.; Waddell, L.A. Zoonotic Babesia: A scoping review of the global evidence. PLoS ONE 2019, 14, e0226781. [Google Scholar] [CrossRef]
- Hussain, S.; Hussain, A.; Aziz, M.U.; Song, B.; Zeb, J.; George, D.; Li, J.; Sparagano, O. A review of zoonotic babesiosis as an emerging public health threat in Asia. Pathogens 2022, 11, 23. [Google Scholar] [CrossRef]
- Desquesnes, M.; Ravel, S.; Cuny, G. PCR identification of Trypanosoma lewisi, a common parasite of laboratory rats. Kinetoplastid Biol. Dis. 2002, 1, 2. [Google Scholar] [CrossRef]
- Goodrich, I.; McKee, C.; Kosoy, M. Trypanosoma (Herpetosoma) diversity in rodents and lagomorphs of New Mexico with a focus on epizootological aspects of infection in Southern Plains woodrats (Neotoma micropus). PLoS ONE 2020, 15, e0244803. [Google Scholar] [CrossRef]
- Desquesnes, M.; Gonzatti, M.; Sazmand, A.; Thévenon, S.; Bossard, G.; Boulangé, A.; Gimonneau, G.; Truc, P.; Herder, S.; Ravel, S.; et al. A review on the diagnosis of animal trypanosomoses. Parasit. Vector. 2022, 15, 64. [Google Scholar] [CrossRef]
- Nussbaum, K.; Honek, J.; Cadmus, C.M.; Efferth, T. Trypanosomatid parasites causing neglected diseases. Curr. Med. Chem. 2010, 17, 1594–1617. [Google Scholar] [CrossRef]
- Votýpka, J.; d’Avila-Levy, C.M.; Grellier, P.; Maslov, D.A.; Lukeš, J.; Yurchenko, V. New approaches to systematics of Trypanosomatidae: Criteria for taxonomic (re)description. Trends Parasitol. 2015, 31, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Magri, A.; Galuppi, R.; Fioravanti, M. Autochthonous Trypanosoma spp. in European mammals: A brief journey amongst the neglected trypanosomes. Pathogens 2021, 10, 334. [Google Scholar] [CrossRef]
- Hoare, C.A. The trypanosomes of mammals. A zoological monograph. Med. J. Aust. 1972, 1, 140. [Google Scholar] [CrossRef]
- Cox, F.E.G. Protozoan parasites of British small rodents. Mamm. Rev. 1987, 17, 59–66. [Google Scholar] [CrossRef]
- Sato, H.; Osanai, A.; Kamiya, H.; Obara, Y.; Jiang, W.; Zhen, Q.; Chai, J.; Une, Y.; Ito, M. Characterization of SSU and LSU rRNA genes of three Trypanosoma (Herpetosoma) grosi isolates maintained in Mongolian jirds. Parasitology 2005, 130, 157–167. [Google Scholar] [CrossRef]
- Guan, G.; Niu, Q.; Yang, J.; Li, Y.; Gao, J.; Luo, J.; Yin, H. Trypanosoma (Herpetosoma) grosi: First isolation from Chinese striped field mouse (Apodemus agrarius). Parasitol. Int. 2011, 60, 101–104. [Google Scholar] [CrossRef]
- Howie, S.; Guy, M.; Fleming, L.; Bailey, W.; Noyes, H.; Faye, J.A.; Pepin, J.; Greenwood, B.; Whittle, H.; Molyneux, D.; et al. A Gambian infant with fever and an unexpected blood film. PLoS Med. 2006, 3, 1508–1512. [Google Scholar] [CrossRef] [PubMed]
- Sarataphan, N.; Vongpakorn, M.; Nuansrichay, B.; Autarkool, N.; Keowkarnkah, T.; Rodtian, P.; Stich, R.W.; Jittapalapong, S. Diagnosis of a Trypanosoma lewisi-like (Herpetosoma) infection in a sick infant from Thailand. J. Med. Microbiol. 2007, 56, 1118–1121. [Google Scholar] [CrossRef] [PubMed]
- Truc, P.; Büscher, P.; Cuny, G.; Gonzatti, M.I.; Jannin, J.; Joshi, P.; Juyal, P.; Lun, Z.-R.; Mattioli, R.; Pays, E.; et al. Atypical human infections by animal trypanosomes. PLoS Negl. Trop. Dis. 2013, 7, e2256. [Google Scholar] [CrossRef] [PubMed]
- Zamoto, A.; Tsuji, M.; Wei, Q.; Cho, S.H.; Shin, E.H.; Kim, T.S.; Leonova, G.N.; Hagiwara, K.; Asakawa, M.; Kariwa, H.; et al. Epizootiologic survey for Babesia microti among small wild mammals in northeastern Eurasia and a geographic diversity in the beta-tubulin gene sequences. J. Vet. Med. Sci. 2004, 66, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Lee, S.E.; Jeong, Y.I.; Kim, H.C.; Chong, S.T.; Klein, T.A.; Song, J.W.; Gu, S.H.; Cho, S.H.; Lee, W.J. Prevalence and molecular characterizations of Toxoplasma gondii and Babesia microti from small mammals captured in Gyeonggi and Gangwon Provinces, Republic of Korea. Vet. Parasitol. 2014, 205, 512–517. [Google Scholar] [CrossRef]
- Kang, S.W.; Doan, H.T.; Choe, S.E.; Noh, J.H.; Yoo, M.S.; Reddy, K.E.; Kim, Y.H.; Kweon, C.H.; Jung, S.C.; Chang, K.Y. Molecular investigation of tick-borne pathogens in ticks from grazing cattle in Korea. Parasitol. Int. 2013, 62, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Kim, S.Y.; Song, B.G.; Rho, J.R.; Cho, C.R.; Kim, C.N.; Um, T.H.; Kwak, Y.G.; Cho, S.H.; Lee, S.E. Detection and characterization of an emerging type of Babesia sp. similar to Babesia motasi for the first case of human babesiosis and ticks in Korea. Emerg. Microbes Infect. 2019, 8, 869–878. [Google Scholar] [CrossRef]
- Kim, T.Y.; Kim, S.Y.; Kim, T.K.; Lee, H.I.; Cho, S.H.; Lee, W.G.; Kim, H. Molecular evidence of zoonotic Babesia species, other than B. microti, in ixodid ticks collected from small mammals in the Republic of Korea. Vet. Med. Sci. 2021, 7, 2427–2433. [Google Scholar] [CrossRef] [PubMed]
- Bang, M.S.; Kim, C.M.; Pyun, S.H.; Kim, D.M.; Yun, N.R. Molecular investigation of tick-borne pathogens in ticks removed from tick-bitten humans in the southwestern region of the Republic of Korea. PLoS ONE 2021, 16, e0252992. [Google Scholar] [CrossRef]
- Kim, T.Y.; Kim, S.Y.; Seo, J.; Lee, H.I.; Lee, W.G.; Kim, H. Detection of US and Kobe-type Babesia microti in ticks collected from small mammals of the Republic of Korea. Entomol. Res. 2024, 54, e12708. [Google Scholar] [CrossRef]
- Kwon, H.Y.; Im, J.H.; Park, Y.K.; Durey, A.; Lee, J.S.; Baek, J.H. Two Imported Cases of Babesiosis with Complication or Co-Infection with Lyme Disease in Republic of Korea. Korean J. Parasitol. 2018, 56, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Na, Y.J.; Chai, J.Y.; Jung, B.K.; Lee, H.J.; Song, J.Y.; Je, J.H.; Seo, J.H.; Park, S.H.; Choi, J.S.; Kim, M.J. An Imported Case of Severe falciparum Malaria with Prolonged Hemolytic Anemia Clinically Mimicking a Coinfection with Babesiosis. Korean J. Parasitol. 2014, 52, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Cho, S.H.; Joo, H.N.; Tsuji, M.; Cho, S.R.; Park, I.J.; Chung, G.T.; Ju, J.W.; Cheun, H.I.; Lee, H.W.; et al. First Case of Human Babesiosis in Korea: Detection and Characterization of a Novel Type of Babesia sp. (KO1) Similar to Ovine Babesia. J. Clin. Microbiol. 2007, 45, 2084–2087. [Google Scholar] [CrossRef] [PubMed]
- Okal, M.N.; Odhiambo, B.K.; Otieno, P.; Bargul, J.L.; Masiga, D.; Villinger, J.; Kalayou, S. Anaplasma and Theileria Pathogens in Cattle of Lambwe Valley, Kenya: A Case for Pro-Active Surveillance in the Wildlife–Livestock Interface. Microorganisms 2020, 8, 1830. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.H.; Welburn, S.C.; Woolhouse, M.E. Theileria annulata: Virulence and transmission from single and mixed clone infections in cattle. Exp. Parasitol. 2002, 100, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Egan, S.L.; Taylor, C.L.; Austen, J.M.; Banks, P.B.; Northover, A.S.; Ahlstrom, L.A.; Ryan, U.M.; Irwin, P.J.; Oskam, C.L. Haemoprotozoan surveillance in peri-urban native and introduced wildlife from Australia. Curr. Res. Parasitol. Vector-Borne Dis. 2021, 1, 100052. [Google Scholar] [CrossRef]
- Mangombi, J.B.; N’dilimabaka, N.; Lekana-Douki, J.B.; Banga, O.; Maghendji-Nzondo, S.; Bourgarel, M.; Leroy, E.; Fenollar, F.; Mediannikov, O. First investigation of pathogenic bacteria, protozoa and viruses in rodents and shrews in context of forest-savannah-urban areas interface in the city of Franceville (Gabon). PLoS ONE 2021, 16, e0248244. [Google Scholar] [CrossRef]
- Sakka, H.; Quéré, J.P.; Kartavtseva, I.; Pavlenko, M.; Chelomina, G.; Atopkin, D.; Bogdanov, A.; Michaux, J. Comparative phylogeography of four Apodemus species (Mammalia: Rodentia) in the Asian Far East: Evidence of Quaternary climatic changes in their genetic structure. Biol. J. Linn. Soc. 2010, 100, 797–821. [Google Scholar] [CrossRef]
Figure 1.
Trapping sites (![Animals 14 00989 i001]() ) in the Republic of Korea (ROK). Map was created using the Free and Open Source QGIS (QGIS 3.28.10. Geographic Information System, http://www.qgis.org, accesed on 8 September 2023).
) in the Republic of Korea (ROK). Map was created using the Free and Open Source QGIS (QGIS 3.28.10. Geographic Information System, http://www.qgis.org, accesed on 8 September 2023).
 ) in the Republic of Korea (ROK). Map was created using the Free and Open Source QGIS (QGIS 3.28.10. Geographic Information System, http://www.qgis.org, accesed on 8 September 2023).
) in the Republic of Korea (ROK). Map was created using the Free and Open Source QGIS (QGIS 3.28.10. Geographic Information System, http://www.qgis.org, accesed on 8 September 2023).
Figure 1.
Trapping sites (![Animals 14 00989 i001]() ) in the Republic of Korea (ROK). Map was created using the Free and Open Source QGIS (QGIS 3.28.10. Geographic Information System, http://www.qgis.org, accesed on 8 September 2023).
) in the Republic of Korea (ROK). Map was created using the Free and Open Source QGIS (QGIS 3.28.10. Geographic Information System, http://www.qgis.org, accesed on 8 September 2023).
) in the Republic of Korea (ROK). Map was created using the Free and Open Source QGIS (QGIS 3.28.10. Geographic Information System, http://www.qgis.org, accesed on 8 September 2023).
Figure 2.
Phylogenetic tree based on neighbor-joining analysis of internal transcriptional spacer 1 (ITS1) from GenBank and Trypanosoma grosi-positive small mammal specimens captured in the ROK in 2021. Trypanosoma grosi sequences obtained in this study are denoted by a solid square (■, GenBank accession numbers OP297200 and OP297201). The numbers on the branches indicate bootstrap percentages based on 1000 replication.
Figure 2.
Phylogenetic tree based on neighbor-joining analysis of internal transcriptional spacer 1 (ITS1) from GenBank and Trypanosoma grosi-positive small mammal specimens captured in the ROK in 2021. Trypanosoma grosi sequences obtained in this study are denoted by a solid square (■, GenBank accession numbers OP297200 and OP297201). The numbers on the branches indicate bootstrap percentages based on 1000 replication.

Figure 3.
Phylogenetic tree based on neighbor-joining analysis of Babesia 18S rRNA from GenBank and Babesia microti-positive small mammal specimens captured in the ROK in 2021. Babesia microti sequences obtained in this study are denoted by a solid triangle (▲, GenBank accession numbers OP804252–OP804254). The numbers on the branches indicate bootstrap percentages based on 1000 replication.
Figure 3.
Phylogenetic tree based on neighbor-joining analysis of Babesia 18S rRNA from GenBank and Babesia microti-positive small mammal specimens captured in the ROK in 2021. Babesia microti sequences obtained in this study are denoted by a solid triangle (▲, GenBank accession numbers OP804252–OP804254). The numbers on the branches indicate bootstrap percentages based on 1000 replication.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Molecular prevalence of Trypanosoma grosi and Babesia microti and genetic diversity of B. microti in small mammals collected from different sites of the Republic of Korea (ROK) in 2021.
Table 1.
Molecular prevalence of Trypanosoma grosi and Babesia microti and genetic diversity of B. microti in small mammals collected from different sites of the Republic of Korea (ROK) in 2021.
| Site | No. Tested | No. T. grosi- Positive (%) | No. B. microti- Positive (%) | B. microti Genotype |
|---|---|---|---|---|
| Gangneung-si | 43 | 11 (25.6) | 14 (32.6) §§ | US type |
| Hwaseong-si | 33 | 12 (36.4) | 1 (3.0) | US type |
| Seogwipo-si | 31 | 15 (48.4) § | 2 (6.5) | Kobe type |
| Jinan-gun | 24 | 5 (20.8) | 2 (8.3) | US type |
| Cheorwon-gun | 23 | 6 (64.7) | 1 (4.3) | US type |
| Geoje-si | 23 | 6 (26.1) | 0 (0.0) | |
| Yeoju-si | 22 | 3 (13.6) | 0 (0.0) | |
| Yeongdeok-gun | 20 | 4 (20.0) | 2 (10.0) | US type |
| Paju-si | 19 | 6 (31.6) | 6 (31.6) | US type |
| Jeongeup-si | 18 | 1 (5.6) | 2 (11.1) | US type |
| Gimcheon-si | 18 | 1 (5.6) | 0 (0.0) | |
| Boseong-gun | 17 | 1 (5.9) | 1 (5.9) | US type |
| Hapcheon-gun | 16 | 1 (6.3) | 2 (12.5) | US type |
| Yesan-si | 14 | 2 (14.3) | 0 (0.0) | |
| Boryeong-si | 8 | 0 (0.0) | 0 (0.0) | |
| Cheongju-si | 8 | 0 (0.0) | 0 (0.0) | |
| Total | 331 | 79 (23.9) | 33 (10.0) |
§ Significantly higher T. grosi infection rate in Seogwipo-si vs. other sites; §§ Significantly higher B. microti infection rate in Gangneung-si vs. other sites.
Table 2.
Prevalence of Trypanosoma grosi and Babesia microti in blood samples from small mammals collected from the ROK in 2021.
Table 2.
Prevalence of Trypanosoma grosi and Babesia microti in blood samples from small mammals collected from the ROK in 2021.
| Small Mammal Species | No. Tested | No. of T. grosi- Positive (%) | No. of B. microti- Positive (%) | No. of Coinfection (%) |
|---|---|---|---|---|
| Apodemus agrarius | 290 | 77 (26.5) | 32 (11.0) | 6 (2.0) |
| Crocidura sp. | 23 | 2 (8.7) | 0 | 0 |
| Apodemus peninsulae | 2 | 0 | 1 (50.0) | 0 |
| Craseomys regulus | 4 | 0 | 0 | 0 |
| Craseomys rufocanus | 1 | 0 | 0 | 0 |
| Cricetulus triton | 1 | 0 | 0 | 0 |
| Micromys minutus | 6 | 0 | 0 | 0 |
| Microtus fortis | 2 | 0 | 0 | 0 |
| Myodes regulus | 1 | 0 | 0 | 0 |
| Rattus norvegicus | 1 | 0 | 0 | 0 |
| Total | 331 | 79 (23.9) | 33 (10.0) | 6 (1.8) |
Table 3.
Relationship between season, ecological habitat, and Trypanosoma grosi and Babesia microti infections in small mammals collected from the ROK in 2021.
Table 3.
Relationship between season, ecological habitat, and Trypanosoma grosi and Babesia microti infections in small mammals collected from the ROK in 2021.
| Categories | No. Tested | No. of T. grosi- Positive (%) | p-Value | No. of B. microti- Positive (%) | p-Value | |
|---|---|---|---|---|---|---|
| Season | Spring | 174 | 46 (26.4) | 0.248 | 17 (9.8) | 0.898 |
| Autumn | 157 | 33 (21.0) | 16 (10.2) | |||
| Ecological habitat | Reservoir | 70 | 8 (11.4) | 0.006 | 9 (12.8) | 0.363 |
| Other | 261 | 71(27.2) | 24(9.2) | |||
| Small mammal species | A. agrarius | 290 | 77 (26.5) | 0.0014 | 32 (11.0) | 0.098 |
| Other | 41 | 2 (4.8) | 1 (2.4) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, H.J.; Han, B.; Lee, H.-I.; Ju, J.-W.; Shin, H.-I. Current Status of Trypanosoma grosi and Babesia microti in Small Mammals in the Republic of Korea. Animals 2024, 14, 989. https://doi.org/10.3390/ani14070989
AMA Style
Kim HJ, Han B, Lee H-I, Ju J-W, Shin H-I. Current Status of Trypanosoma grosi and Babesia microti in Small Mammals in the Republic of Korea. Animals. 2024; 14(7):989. https://doi.org/10.3390/ani14070989
Chicago/Turabian StyleKim, Hyun Jung, BoGyeong Han, Hee-Il Lee, Jung-Won Ju, and Hyun-Il Shin. 2024. "Current Status of Trypanosoma grosi and Babesia microti in Small Mammals in the Republic of Korea" Animals 14, no. 7: 989. https://doi.org/10.3390/ani14070989
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.