
Effects of Isochlorogenic Acid on Ewes Rumen Fermentation, Microbial Diversity and Ewes Immunity of Different Physiological Stages

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Material
2.2. Experimental Design
2.3. Sample Collection and Processing
2.4. Rumen Fermentation Index Determination
2.5. Analysis of Rumen Microbial Diversity
2.6. Blood Immune Index Determination
2.7. Statistical Analysis
3. Results
3.1. Effects of ICGA on Ewes Rumen Fermentation at Different Physiological Stages
3.2. Effects of ICGA on Ewes Rumen Microbial Diversity at Different Physiological Stages
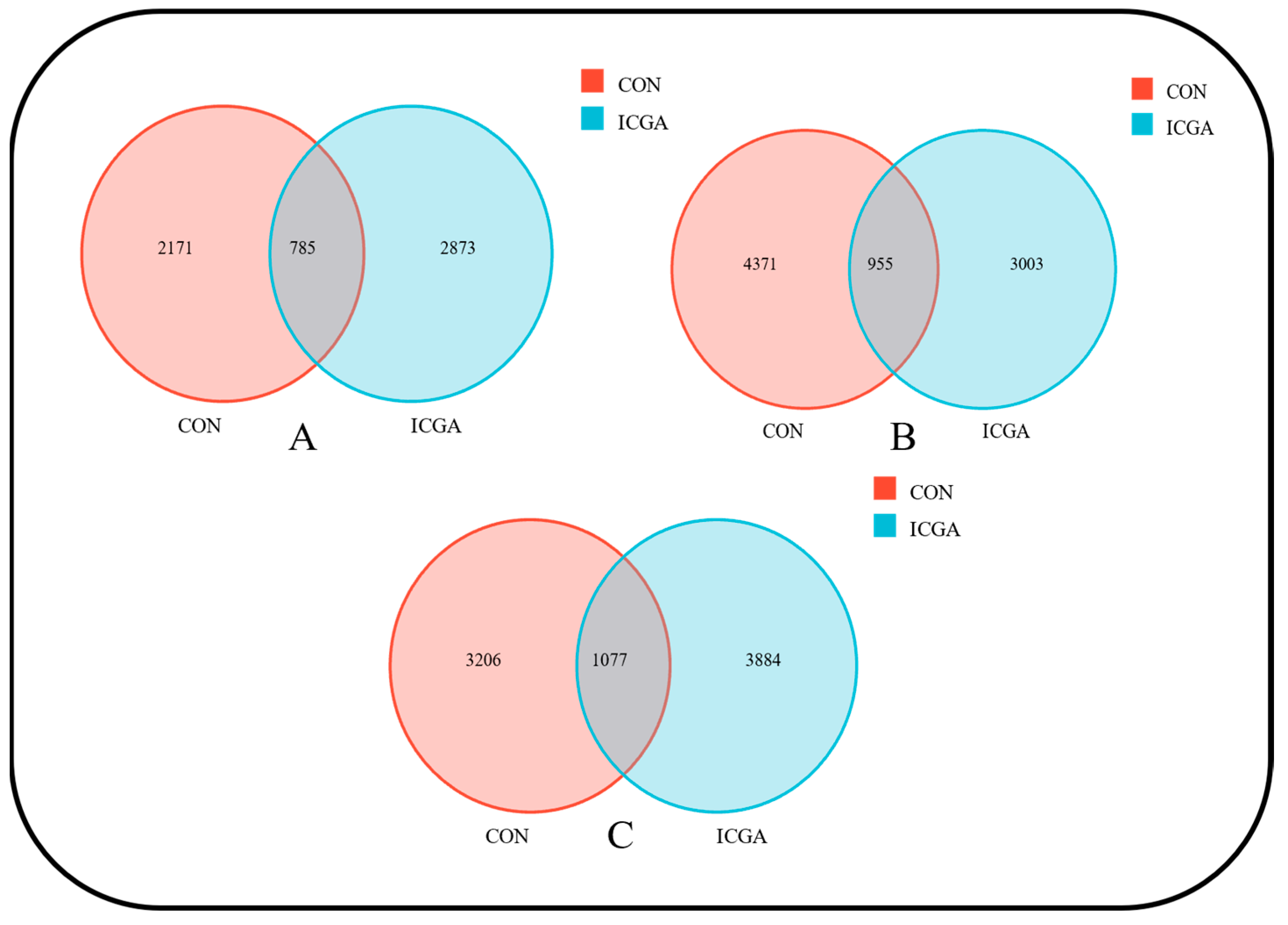
3.2.1. Effect on the Number of Operational Taxonomic Units (OUT)
3.2.2. Effect on Dilution Curve
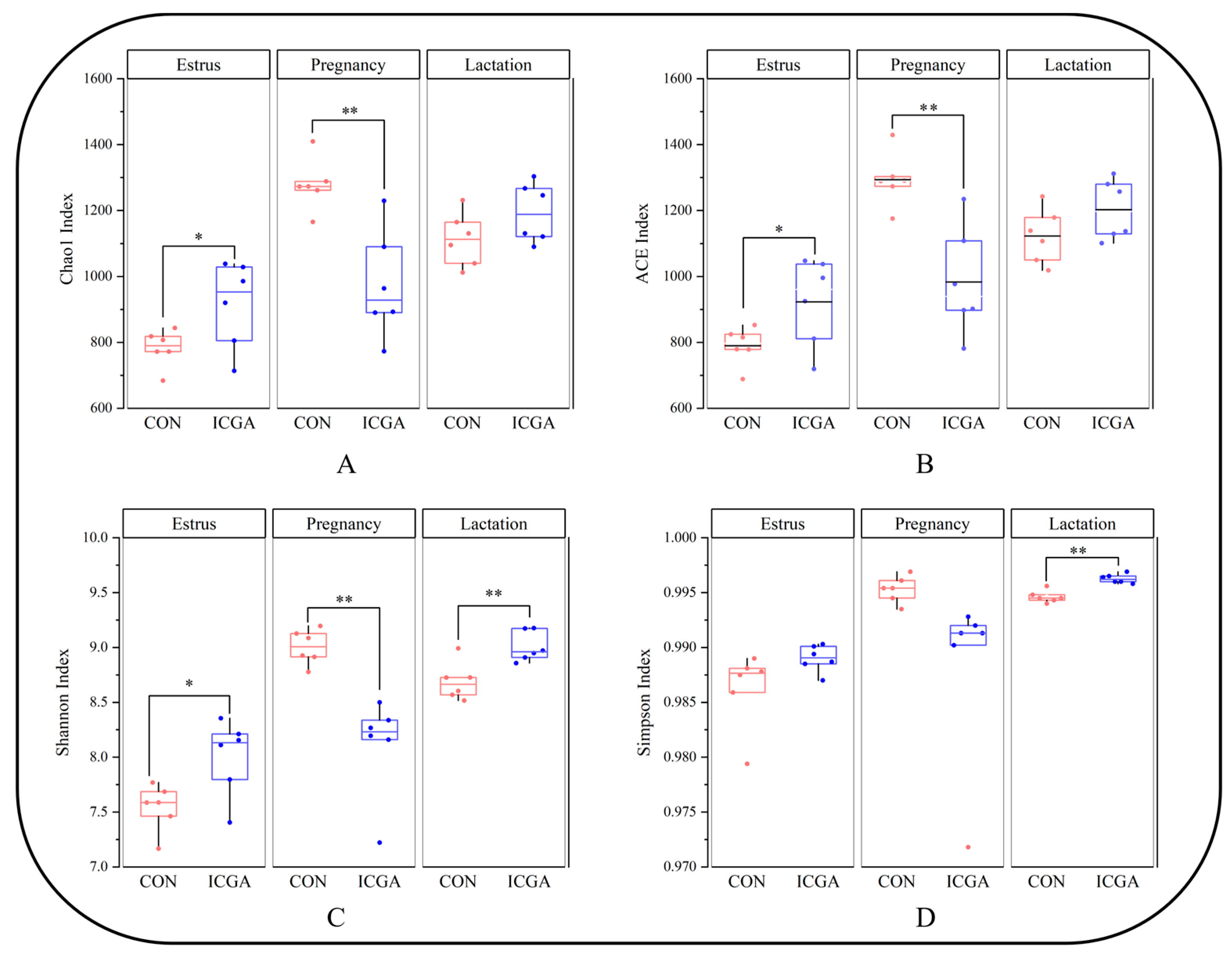
3.2.3. Effect on Alpha Diversity
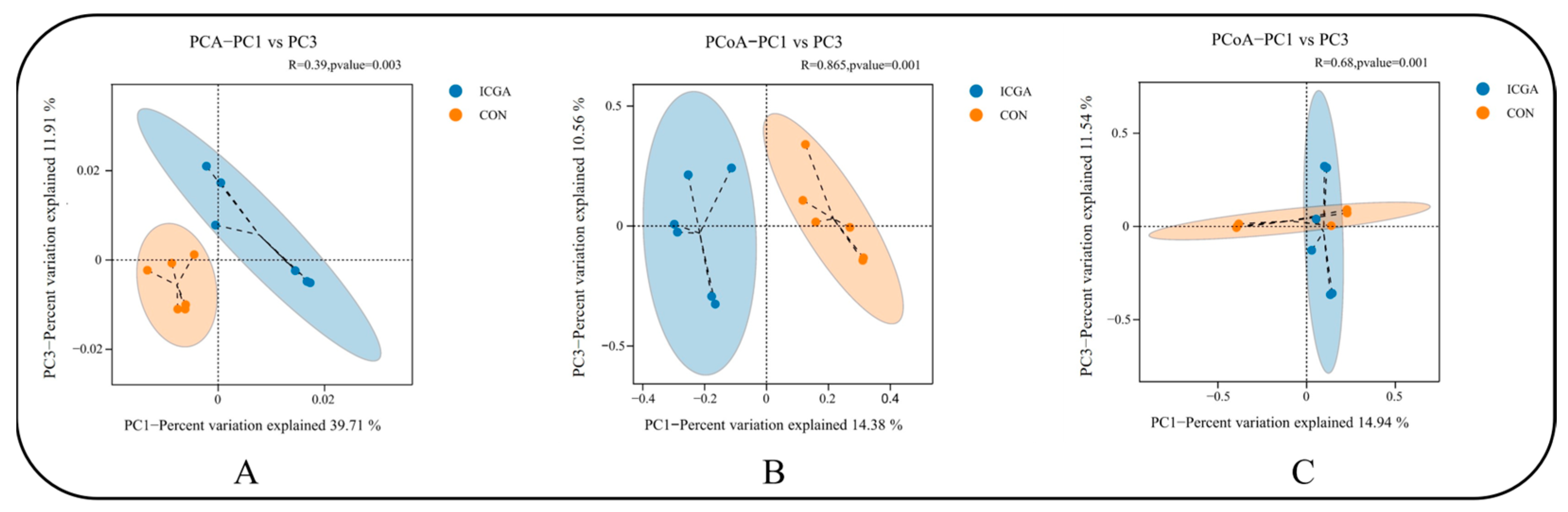
3.2.4. Effect on Beta Diversity
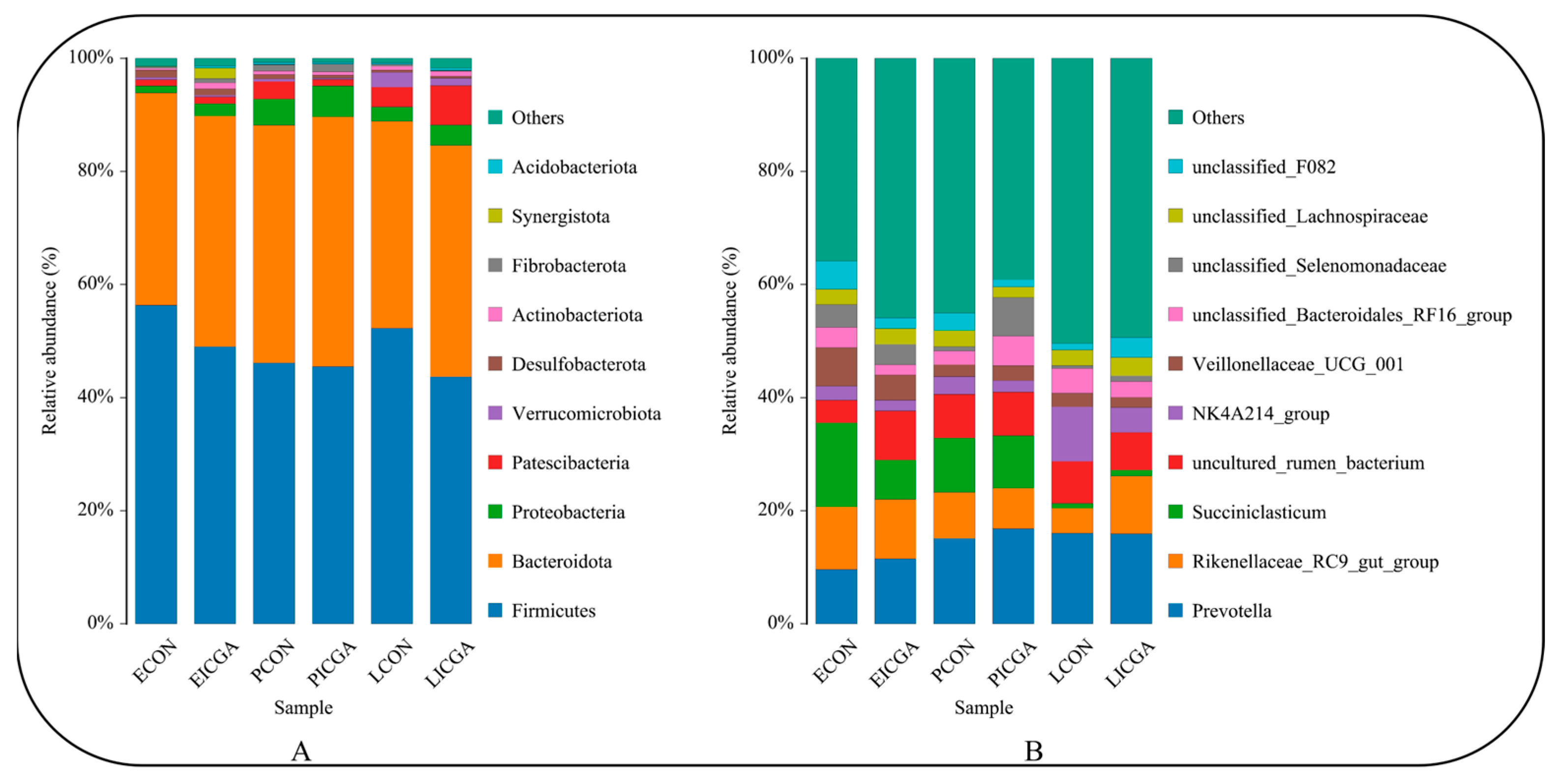
3.2.5. Effect on Bacterial Composition and Structure at the Phylum Level
3.2.6. Effect on Bacterial Composition and Structure at the Genus Level
3.2.7. Effect on LEfSe Difference
3.3. Effects of ICGA on Ewes Blood Immune Index at Different Physiological Stages
4. Discussion
4.1. Effects of ICGA on Ewes Rumen Fermentation at Different Physiological Stages
4.2. Effects of ICGA on Ewes Rumen Microbial Diversity at Different Physiological Stages
4.3. Effects of ICGA on Ewes Blood Immune Index at Different Physiological Stages
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Y.C.; Zhao, Q.R.; Zhai, T.C.; Si, W. Structural transition of protein intake in urban China: Stage characteristics and driving forces. Agribusiness 2023, 39, 1559–1577. [Google Scholar] [CrossRef]
- Ma, Z.F. Research on the Construction and Application of Dairy Big Data Platform from the Perspective of Grassland Agriculture; Lanzhou University: Lanzhou, China, 2020. [Google Scholar]
- Li, X.L.; Yuan, Q.H.; Wan, L.Q.; He, F. Perspectives on livestock production systems in China. Rangel. J. 2008, 30, 211–220. [Google Scholar] [CrossRef]
- Khanal, P.; Nielsen, M.O. Impacts of prenatal nutrition on animal production and performance: A focus on growth and metabolic and endocrine function in sheep. J. Anim. Sci. Biotechnol. 2017, 8, 75. [Google Scholar] [CrossRef]
- Zhang, S.Z.; Zhang, S.B.; Han, J.Y.; Liu, H.B.; Chen, Q.L.; Yang, J.F.; Shi, Z.H.; Wang, Y.Z.; Li, X.L.; Song, L.G. Effects of compound Chinese herbal medicine additive on reproductive performance, some blood biochemical indexes and reproductive hormones of Hu sheep ewes. Heilongjiang J. Anim. Sci. Vet. Med. Sci. 2020, 16, 118–120. [Google Scholar]
- Hossain, M.F.; Islam, M.T.; Islam, M.A.; Akhtar, S. Cultivation and uses of stevia (Stevia rebaudiana Bertoni): A review. Afr. J. Food Agric. Nutr. Dev. 2017, 17, 12745–12757. [Google Scholar] [CrossRef]
- Angelini, L.G.; Martini, A.; Passera, B.; Tavarini, S. Cultivation of Stevia rebaudiana Bertoni and associated challenges. In Sweeteners; Springer International Publishing AG: Cham, Switzerland, 2018; pp. 35–85. [Google Scholar]
- Chen, F.; Du, E.C.; Fan, Q.W.; Wei, J.T. The application and research progress of chlorogenic acids in livestock and poultry production. Hubei Agric. Sci. 2020, 59, 10–13+16. [Google Scholar]
- Xi, L.S.; Mu, T.H.; Sun, H.N. Research progress of chlorogenic acids at home and abroad. Acta Agric. Nucl. Sin. 2014, 28, 292–301. [Google Scholar]
- Chen, J.L.; Chen, D.W.; Yu, B.; Luo, Y.H.; Zheng, P.; Mao, X.B.; Yu, J.; Luo, J.Q.; Huang, Z.Q.; Yan, H.; et al. Chlorogenic acid attenuates oxidative stress-Induced intestinal mucosa disruption in weaned pigs. Front. Vet. Sci. 2022, 9, 806253. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.L.; Liu, Y.; Hu, J.J.; Gao, Y.Y.; Ma, Y.N.; Wen, D.L. Chlorogenic acid-induced gut microbiota improves metabolic endotoxemia. Front. Endocrinol. 2021, 12, 762691. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q. Effects of Chlorogenic Acid on Growth Performance, Gastrointestinal Tissue Morphology and Microflora of Hu Sheep. Master’s Thesis, Henan Agricultural University, Zhengzhou, China, 2022. [Google Scholar]
- Liu, Y.; Zhang, Y.; Bai, D.; Li, Y.Q.; He, X.L.; Ito, K.; Liu, K.X.; Tan, H.Q.; Zhen, B.K.; Ma, Y.B. Dietary supplementation with chlorogenic acid enhances antioxidant capacity, which promotes growth, jejunum barrier function, and cecum microbiota in broilers under high stocking density stress. Animals 2023, 13, 303. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Zhen, J.D.; Liu, W. Research progress on biological activity and extraction process of isochlorogenic acid. Feed Ind. 2021, 42, 54–59. [Google Scholar]
- Li, Y.C.; Niu, Z.P.; Xu, M.L.; Tian, H.; Gao, W. Comparative analysis of chlorogenic acids in stevia and other plants. China Food Addit. 2021, 32, 1–6. [Google Scholar]
- Li, S.Y.; Jiao, T.; Qi, S.; Zhang, X.; Wang, H.N.; Zhao, S.G. In vitro study of the effects of isochlorogenic acid supplementation on nutrient digestion and rumen fermentation in sheep diets. J. Zhejiang Agric. 2024, accepted. [Google Scholar]
- Ren, Q.M. Study on the Structure and Function of Microbial Community in Different Niches of Yak Rumen. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2022. [Google Scholar]
- Wang, J.; Zhang, L.Q.; Wang, W.L.; Zhao, Z.W.; Ma, Q.; Wen, X.F. Effects of caragana korshinskii fermented feed on growth performance and rumen microflora of Ningxia Tan sheep. J. Anim. Nutr. 2023, 35, 1035–1045. [Google Scholar]
- Guo, H.; Yang, H.C.; Gao, Q.X.; Xin, G.X. Rumen microbiota and its regulation. Pratacultural Sci. 2022, 39, 371–380. [Google Scholar]
- Shen, Y.Y.; Tong, J.J.; Xiong, B.H.; Jiang, L.S. Research progress of multi-omics technology in the interaction mechanism between rumen microorganisms and host in dairy cows. Chin. Milk Ind. 2021, 8, 68–75. [Google Scholar]
- Zhao, C.X.; Liu, G.W.; Li, X.B.; Guan, Y.; Wang, Y.Z.; Yuan, X.; Sun, G.Q.; Wang, Z.; Li, X.W. Inflammatory mechanism of rumenitis in dairy cows with subacute ruminal acidosis. BMC Vet. Res. 2018, 14, 135. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.X.; Bian, Q.; Zhang, S.P.; Wang, F.; Liu, H.L.; Zhang, J.B. The relationship between rumen health, body health and reproductive performance in dairy cows. Hunan Feed 2021, 3, 31–34. [Google Scholar]
- Rey, J.; Otálora DD, X.; Atxaerandio, R.; Mandaluniz, N.; García-Rodríguez, A.; González-Recio, O.; Lopez-García, A.; Ruiz, R.; Goiri, A. Effect of chitosan on ruminal fermentation and microbial communities, methane emissions, and productive performance of dairy cattle. Animals 2023, 13, 2861. [Google Scholar] [CrossRef]
- Feng, Z.C.; Gao, M. Improvement of colorimetric method for determination of ammonia nitrogen in rumen fluid. Anim. Husb. Feed Sci. 2010, 31, 37. [Google Scholar]
- Luo, T.; Li, Y.T.; Zhang, W.Y.; Liu, J.X.; Shi, H.B. Rumen and fecal microbiota profiles associated with immunity of young and adult goats. Front. Immunol. 2022, 13, 978402. [Google Scholar] [CrossRef]
- Li, H.T.; Yang, Z.B.; Yang, W.R.; Jiang, S.Z.; Zhang, G.G.; Zhang, Q.; Zhang, C.Y. Effects of different dietary types on rumen pH, ammonia nitrogen and volatile fatty acid concentration in Dorper sheep. Chin. J. Anim. Husb. 2017, 53, 67–72. [Google Scholar]
- Zhang, H.T. Effects of Different Nutritional Levels on Reproductive Performance (Estrus Time) of Ultra-Early-Weaned Small-Tailed Han Ewes. Master’s Thesis, Gansu Agricultural University, Lanzhou, China, 2004. [Google Scholar]
- Weston, R.H. The effect of pregnancy and early lactation on the digestion of a medium-quality roughage. Aust. J. Agric. Res. 1988, 39, 659–669. [Google Scholar] [CrossRef]
- Zhang, F.; Diao, Q.Y. Effects of energy on the health of ewes and their lambs during late pregnancy. Chin. Anim. Vet. 2017, 44, 1369–1374. [Google Scholar]
- Du, H.D.; Na, R.H. Research progress on physiological metabolism and microbial changes of ruminants during gestation and lactation and their effects on offspring development. J. Anim. Husb. Vet. Med. 2023, 54, 4458–4467. [Google Scholar]
- Kaske, M.; Groth, A. Changes in factors affecting the rate of digesta passage during pregnancy and lactation in sheep fed on hay. Reprod. Nutr. Dev. 1997, 37, 573–588. [Google Scholar] [CrossRef]
- Liu, Y.J. Study on the Differences in Physiological Metabolism and Rumen Internal Environment of Dairy Cows with Different Milk Yields. Master’s Thesis, Jilin Agricultural University, Changchun, China, 2021. [Google Scholar]
- Cui, C.Y.; Li, W.J.; Zhang, F.; Wang, Y.L.; Wu, Q.C.; Wang, W.K.; Jiang, Y.W.; Chen, H.W.; Yang, H.J. Effects of Bacillus subtilis on rumen microbial fermentation and degradation characteristics of peanut vine, oat and alfalfa hay. Chin. J. Anim. Husb. 2023, 59, 266–271. [Google Scholar]
- Luo, Z.J.; Ma, L.; Pu, D.P.; Qin, X. Meta-analysis of the effects of rumen fermentation product acetic acid and propionic acid ratio on production performance and milk composition of dairy cows. Chin. Anim. Husb. Vet. Med. 2021, 48, 1613–1624. [Google Scholar]
- Xiang, K.H.; Hu, X.Y.; Li, S.; Wang, Y.; Zhang, Y.X.; Zhang, N.S. Research progress on the changes and influencing factors of rumen flora in dairy cows during perinatal period. Vet. Med. Prog. 2022, 43, 93–97. [Google Scholar]
- Guo, Y.J. Comparison of Rumen Outflow Rate and Microflora of Ewes during Non-Pregnancy, Pregnancy and Lactation; Shanxi Agricultural University: Jinzhong, China, 2021. [Google Scholar]
- Yang, D.; Yin, F.Y.; He, R.H.; Tian, R.G.; Wu, J.H. Research progress on rumen and intestinal microorganisms in ruminants. Anim. Husb. Feed Sci. 2021, 42, 28–38. [Google Scholar]
- Chaput, N.; Lepage, P.; Coutzac, C.; Soularue, E.; Roux, K.L.; Monot, C.; Boselli, L.; Routier, E.; Cassard, L.; Collins, M.; et al. Baseline gut microbiota predicts clinical response and colitis in metastatic melanoma patients treated with ipilimumab. Ann. Oncol. 2017, 28, 1368–1379. [Google Scholar] [CrossRef]
- Shi, A.M.; Li, T.; Zheng, Y.; Song, Y.H.; Wang, H.T.; Wang, N.; Dong, L.; Shi, H.T. Chlorogenic acid improves NAFLD by regulating gut microbiota and GLP-1. Front. Pharmacol. 2021, 12, 693048. [Google Scholar] [CrossRef]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the bovine rumen bacterial community from birth to adulthood. Isme J. 2013, 7, 1069–1079. [Google Scholar] [CrossRef]
- Dodd, D.; Moon, Y.H.; Swaminathan, K.; Mackie, R.; Cann, I. Transcriptomic analyses of xylan degradation by prevotella bryantii and insights into energy acquisition by xylanolytic Bacteroidetes. Biol. Chem. 2010, 285, 30261–30273. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.N.; Zhang, Y.M.; Ao, C.J.; Zhang, T.L.; Zhang, X.F.; Bai, C.; Zhao, Y.B.; Qi, J.W. Effects of γ-polyglutamic acid on rumen fermentation and microflora structure of Small Tail Han sheep. Chin. J. Anim. Husb. 2021, 57, 136–142. [Google Scholar]
- Long, T.H.; Ou Yang, K.H.; Fang, Y.T.; Chen, X.F.; Wang, C.B.; Li, Y.J.; Ou Yang, K.H.; Qiu, Q.H. Effects of dietary lysine supplementation on rumen fermentation characteristics and microbial community structure. Feed Ind. 2023, 44, 1–9. [Google Scholar]
- Zhang, K. Study on the Vertical Effect of Maternal Microorganisms on Gastrointestinal Flora and Intestinal Function of Offspring during Pregnancy-Lactation Period; Northwest University of Agriculture and Forestry: Xianyang, China, 2022. [Google Scholar]
- Stevenson, D.M.; Weimer, P.J. Dominance of prevotella and low abundance of classical ruminal bacterial species in the bovine rumen revealed by relative quantification real-time PCR. Appl. Microbiol. Biotechnol. 2007, 75, 165–174. [Google Scholar] [CrossRef]
- Scheifinger, C.C.; Linehan, B.; Wolin, M.J. H2 production by selenomonas ruminantium in the absence and presence of methanogenic bacteria. Appl. Microbiol. 1975, 29, 480–483. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.W.; Chi, Z.; Deng, M.; Tian, H.C.; Zhong, C.M.; Sun, B.L.; Guo, Y.Q. Effects of dietary concentrate and chromium propionate levels on growth performance, fecal fermentation parameters and microflora of Leizhou goats. Ann. Anim. Nutr. 2021, 33, 5781–5793. [Google Scholar]
- Zhao, G.L.; Niu, Y.; Zhang, R.Q.; Dai, B.; Yang, C.M. Effects of yeast culture on growth performance, immune function and caecal microflora of yellow-feathered broilers. Ann. Anim. Nutr. 2022, 34, 7701–7710. [Google Scholar]
- Van, G.N.O. Succiniclasticum ruminis gen. nov., sp. nov., a ruminal bacterium converting succinate to propionate as the sole energy-yielding mechanism. Int. J. Syst. Bacteriol. 1995, 45, 297–300. [Google Scholar]
- Wuriliga Aladar Cui, S.; Xiao, M.; Gong, J. Effects of selenium on rumen fermentation and rumen microflora of grazing sheep under heat stress. Chin. Feed 2022, 3, 8–16. [Google Scholar]
- Yang, L. Effects of Leonurine on Immune and Antioxidation in Broilers and the Related Mechanisms. Ph.D. Thesis, Shihezi University, Shihezi, China, 2019. [Google Scholar]
- Gao, Y. Effects of Feeding Acidified Milk on Growth Performance, Blood Immunologic Indicator and Fecal Microorganism Diversity in Holstein Calves; Heilongjiang Bayi Agricultural University: Daqing, China, 2018. [Google Scholar]
- Zhang, Y.Q.; Liu, Y.Z.; Yang, Y.F.; Li, J.G. Effects of feeding ginseng powder to sows during the perinatal period on their reproductive performance and blood immunoglobulins. Pig Breed. 2019, 2, 41–44. [Google Scholar]
- Zhang, L.; Bai, H.; Wang, Z.T. Changes of hCG and IL-1β, IL-6, TNF-α serum level in different phase of pregnancy. J. Armed Police Med. Coll. 2008, 10, 864–868. [Google Scholar]
- Feng, D.L.; Yin, C.X.; You, H. A study on the correlation between placental IL-6 mRNA and amniotic fluid IL-6 level with preeclampsia. Chin. J. Matern. Child Health 2001, 16, 765–767. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological Period | Estrus (7.5 Months Old) | Pregnancy (7.5–12.5 Months Old) | Lactation (12.5–14.5 Months Old) | |
|---|---|---|---|---|
| Formula composition/% | Whole plant silage corn | 20 | 10 | 22 |
| Wheat straw | 25 | 25 | 10 | |
| Alfalfa | 25 | 30 | 33 | |
| Corn grain | 13 | 15 | 15 | |
| Bran | 2 | 5 | 5 | |
| Sesame cake | 8 | 7 | 7 | |
| Cottonseed meal | 3 | 4 | 4 | |
| Baking soda | 0.5 | 0.5 | 0.5 | |
| Calcium hydrogen phosphate | 1 | 1 | 1 | |
| Salt | 0.5 | 0.5 | 0.5 | |
| Premix 1 | 2 | 2 | 2 | |
| Total | 100 | 100 | 100 | |
| Nutritional level | Dry matter, DM (%) | 80.19 | 85.91 | 82.85 |
| Digestible energy, DE/(MJ kg −1) | 11.01 | 11.53 | 13.03 | |
| Metabolizable energy, ME/(MJ kg −1) | 9.07 | 8.85 | 9.12 | |
| Calcium, Ca (%) | 1.94 | 1.07 | 1.08 | |
| Phosphorus, P (%) | 0.71 | 0.54 | 0.58 | |
| Crude protein, CP (%) | 12.89 | 13.76 | 13.07 | |
| Index | Estrus | Pregnancy | Lactation | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CON | ICGA | p Value | CON | ICGA | p Value | CON | ICGA | p Value | |
| pH | 6.55 ± 0.05 b | 6.91 ± 0.02 a | 0.002 | 6.73 ± 0.13 a | 6.28 ± 0.02 b | 0.025 | 6.52 ± 0.15 a | 6.93 ± 0.04 a | 0.053 |
| Ammonia nitrogen (mg dL−1) | 8.62 ± 0.19 b | 10.50 ± 0.60 a | 0.040 | 4.79 ± 0.78 a | 4.16 ± 0.20 a | 0.481 | 8.10 ± 0.94 a | 4.73 ± 0.25 a | 0.061 |
| Acetic acid (mmol L−1) | 49.39 ± 3.49 a | 44.18 ± 1.08 a | 0.227 | 26.14 ± 0.46 b | 45.63 ± 2.14 a | 0.009 | 45.30 ± 1.91 a | 38.33 ± 3.72 a | 0.172 |
| Propionic acid (mmol L−1) | 12.91 ± 0.85 a | 9.36 ± 0.46 b | 0.021 | 6.64 ± 0.67 b | 18.99 ± 1.37 a | 0.001 | 15.00 ± 1.15 a | 10.84 ± 1.07 a | 0.057 |
| Isobutyric acid (mmol L−1) | 0.59 ± 0.04 a | 0.66 ± 0.02 a | 0.214 | 0.26 ± 0.02 b | 0.41 ± 0.03 a | 0.011 | 0.55 ± 0.11 a | 0.43 ± 0.01 a | 0.405 |
| Butyric acid (mmol L−1) | 7.99 ± 0.46 a | 4.58 ± 0.20 b | 0.002 | 3.31 ± 0.11 b | 7.26 ± 0.39 a | 0.001 | 6.58 ± 0.79 a | 5.49 ± 0.40 a | 0.284 |
| Isovaleric acid (mmol L−1) | 0.93 ± 0.06 a | 0.95 ± 0.10 a | 0.872 | 0.32 ± 0.00 b | 0.64 ± 0.02 a | 0.003 | 0.88 ± 0.18 a | 0.78 ± 0.04 a | 0.638 |
| Valeric acid (mmol L−1) | 0.52 ± 0.01 a | 0.48 ± 0.03 a | 0.365 | 0.27 ± 0.02 b | 0.57 ± 0.05 a | 0.006 | 0.64 ± 0.06 a | 0.61 ± 0.02 a | 0.715 |
| Acetic acid/Propionic acid | 3.82 ± 0.02 b | 4.74 ± 0.16 a | 0.029 | 4.00 ± 0.34 a | 2.42 ± 0.12 b | 0.012 | 3.04 ± 0.11 a | 3.55 ± 0.21 a | 0.100 |
| Total volatile fatty acids (mmol L−1)) | 71.45 ± 3.69 a | 59.1 ± 2.21 b | 0.045 | 36.88 ± 1.07 b | 78.31 ± 4.78 a | 0.001 | 67.17 ± 3.69 a | 56.39 ± 5.21 a | 0.167 |
| Phylum | Estrus | Pregnancy | Lactation | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CON | ICGA | p Value | CON | ICGA | p Value | CON | ICGA | p Value | |
| Firmicutes | 56.05 ± 0.04 | 48.86 ± 0.02 | 0.99 | 46.42 ± 0.04 | 45.78 ± 0.02 | 1.00 | 51.80 ± 0.03 | 42.78 ± 0.04 | 0.93 |
| Bacteroidota | 37.79 ± 0.04 | 40.78 ± 0.02 | 1.00 | 42.13 ± 0.02 | 42.86 ± 0.02 | 1.00 | 36.92 ± 0.02 | 41.78 ± 0.05 | 0.99 |
| Proteobacteria | 1.26 ± 0.00 | 2.25 ± 0.01 | 1.00 | 4.78 ± 0.01 | 5.68 ± 0.02 | 1.00 | 2.60 ± 0.00 | 3.70 ± 0.00 | 1.00 |
| Patescibacteria | 1.09 ± 0.00 | 1.22 ± 0.00 | 1.00 | 3.11 ± 0.00 | 0.92 ± 0.00 | 0.06 | 3.51 ± 0.00 | 6.92 ± 0.01 | 0.00 |
| Verrucomicrobiota | 0.41 ± 0.00 | 0.28 ± 0.00 | 1.00 | 0.44 ± 0.00 | 0.27 ± 0.00 | 1.00 | 2.63 ± 0.01 | 1.26 ± 0.00 | 0.48 |
| Desulfobacterota | 1.28 ± 0.00 | 1.13 ± 0.00 | 1.00 | 0.80 ± 0.00 | 0.48 ± 0.00 | 0.99 | 0.38 ± 0.00 | 0.33 ± 0.00 | 1.00 |
| Actinobacteriota | 0.27 ± 0.00 | 1.13 ± 0.01 | 0.55 | 0.63 ± 0.00 | 0.66 ± 0.00 | 1.00 | 0.67 ± 0.00 | 0.87 ± 0.00 | 1.00 |
| Fibrobacterota | 0.35 ± 0.00 | 0.72 ± 0.00 | 1.00 | 1.16 ± 0.01 | 1.64 ± 0.01 | 1.00 | 0.38 ± 0.00 | 0.11 ± 0.00 | 1.00 |
| Synergistota | 0.06 ± 0.00 | 1.88 ± 0.01 | 0.00 | 0.04 ± 0.00 | 0.03 ± 0.00 | 1.00 | 0.03 ± 0.00 | 0.05 ± 0.00 | 1.00 |
| Acidobacteriota | 0.01 ± 0.00 | 0.35 ± 0.00 | 0.41 | 0.34 ± 0.00 | 0.31 ± 0.00 | 1.00 | 0.26 ± 0.00 | 0.40 ± 0.00 | 1.00 |
| Genus | Estrus | Pregnancy | Lactation | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CON | ICGA | p Value | CON | ICGA | p Value | CON | ICGA | p Value | |
| Prevotella | 9.67 ± 0.01 | 11.49 ± 0.01 | 1.00 | 14.97 ± 0.01 | 16.78 ± 0.01 | 1.00 | 16.09 ± 0.01 | 16.44 ± 0.03 | 1.00 |
| Rikenellaceae_RC9_gu-t_group | 11.16 ± 0.01 | 10.44 ± 0.01 | 1.00 | 8.13 ± 0.01 | 6.89 ± 0.00 | 1.00 | 4.54 ± 0.01 | 10.36 ± 0.01 | 0.00 |
| Succiniclasticum | 14.80 ± 0.01 | 6.98 ± 0.01 | 0.00 | 9.45 ± 0.01 | 9.78 ± 0.02 | 1.00 | 0.88 ± 0.00 | 1.11 ± 0.00 | 1.00 |
| uncultured_rumen_bac-terium | 4.00 ± 0.01 | 8.70 ± 0.02 | 0.23 | 7.90 ± 0.01 | 7.19 ± 0.02 | 1.00 | 7.53 ± 0.01 | 6.76 ± 0.01 | 1.00 |
| NK4A214_group | 2.46 ± 0.00 | 1.89 ± 0.00 | 1.00 | 3.08 ± 0.00 | 2.05 ± 0.00 | 1.00 | 9.54 ± 0.02 | 4.32 ± 0.00 | 0.00 |
| Veillonellaceae_UCG_001 | 6.76 ± 0.01 | 4.45 ± 0.01 | 0.27 | 2.08 ± 0.00 | 2.95 ± 0.01 | 1.00 | 2.4 ± 0.01 | 1.81 ± 0.00 | 1.00 |
| unclassified_Bacteroid-ales_RF16_group | 3.56 ± 0.01 | 1.81 ± 0.01 | 0.91 | 2.40 ± 0.01 | 5.59 ± 0.02 | 0.09 | 4.45 ± 0.01 | 2.79 ± 0.00 | 0.94 |
| unclassified_Selenom-onadaceae | 4.11 ± 0.01 | 3.57 ± 0.01 | 1.00 | 0.72 ± 0.00 | 6.87 ± 0.04 | 0.02 | 0.52 ± 0.00 | 0.94 ± 0.00 | 1.00 |
| unclassified_Lachnospi-raceae | 2.75 ± 0.00 | 2.77 ± 0.01 | 1.00 | 2.87 ± 0.00 | 2.00 ± 0.00 | 1.00 | 2.77 ± 0.01 | 3.2 ± 0.00 | 1.00 |
| unclassified_F082 | 5.09 ± 0.01 | 1.93 ± 0.01 | 0.02 | 3.05 ± 0.00 | 1.14 ± 0.00 | 0.56 | 1.15 ± 0.00 | 3.56 ± 0.00 | 0.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Li, X.; Sha, Y.; Qi, S.; Zhang, X.; Wang, H.; Wang, Z.; Zhao, S.; Jiao, T. Effects of Isochlorogenic Acid on Ewes Rumen Fermentation, Microbial Diversity and Ewes Immunity of Different Physiological Stages. Animals 2024, 14, 715. https://doi.org/10.3390/ani14050715
Li S, Li X, Sha Y, Qi S, Zhang X, Wang H, Wang Z, Zhao S, Jiao T. Effects of Isochlorogenic Acid on Ewes Rumen Fermentation, Microbial Diversity and Ewes Immunity of Different Physiological Stages. Animals. 2024; 14(5):715. https://doi.org/10.3390/ani14050715
Chicago/Turabian StyleLi, Shuyan, Xiongxiong Li, Yuzhu Sha, Shuai Qi, Xia Zhang, Huning Wang, Zhengwen Wang, Shengguo Zhao, and Ting Jiao. 2024. "Effects of Isochlorogenic Acid on Ewes Rumen Fermentation, Microbial Diversity and Ewes Immunity of Different Physiological Stages" Animals 14, no. 5: 715. https://doi.org/10.3390/ani14050715