


The Effects of Ethanol and Acetic acid on Behaviour of Extranidal Workers of the Narrow-Headed Ant Formica exsecta (Hymenoptera, Formicidae) during a Field Experiment
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. The Role of Animal Models in the Research on Human Mental Health and Mental Disorders
1.2. Insect Models Used in the Research on Drug and Ethanol Addiction
1.3. Methods of Administration of Ethanol to Insects
1.4. Interrelationships between the Consumption of/Exposure to Ethanol and Behaviour, Cognitive Proceses and Depression-like States Encountered in Insects
1.5. Preference for Ethanol as an Evolutionary Adaptation
1.6. Ants as Subjects in the Research Carried out in the Field of Behavioural Neuroscience and Neuropsychopharmacology
1.7. Field Versus Laboratory Studies of the Effects of Ethanol on Insect Behaviour and the Main Aims of the Present Research
1.8. Ant Species
2. Materials and Methods
2.1. Field Site
2.2. Experimental Compounds and Their Concentrations
2.3. Tests
2.4. Behavioural Categories
- (1)
- Ignoring the experimental object by an ant passing close to it;
- (2)
- Stopping close to the experimental object accompanied by movements of the antennae;
- (3)
- Withdrawal from the experimental object immediately after stopping close to it;
- (4)
- Bypassing the experimental object (giving it a wide berth, but within a distance no longer than 2–3 ant body lengths);
- (5)
- Jumping away from the experimental object after an antennal contact with it;
- (6)
- Antennal contact/contacts with the experimental object;
- (7)
- Nibbling of the experimental object;
- (8)
- Nibbling of the experimental object accompanied by gaster flexing, a form of threatening behaviour;
- (9)
- Nibbling at the substrate close to the experimental object;
- (10)
- Self-grooming of the antennae close to the experimental object;
- (11)
- Self-grooming of the gaster close to the experimental object;
- (12)
- Charge at a nestmate observed close to the experimental object;
- (13)
- Open-mandible threat directed at a nestmate close to the experimental object;
- (14)
- Dragging of a nestmate close to the experimental object;
- (15)
- Antennal contact/contacts with a nestmate close to the experimental object;
- (16)
- Allogrooming of a nestmate close to the experimental object.
2.5. Statistical Analysis of the Data
3. Results
3.1. General Remarks
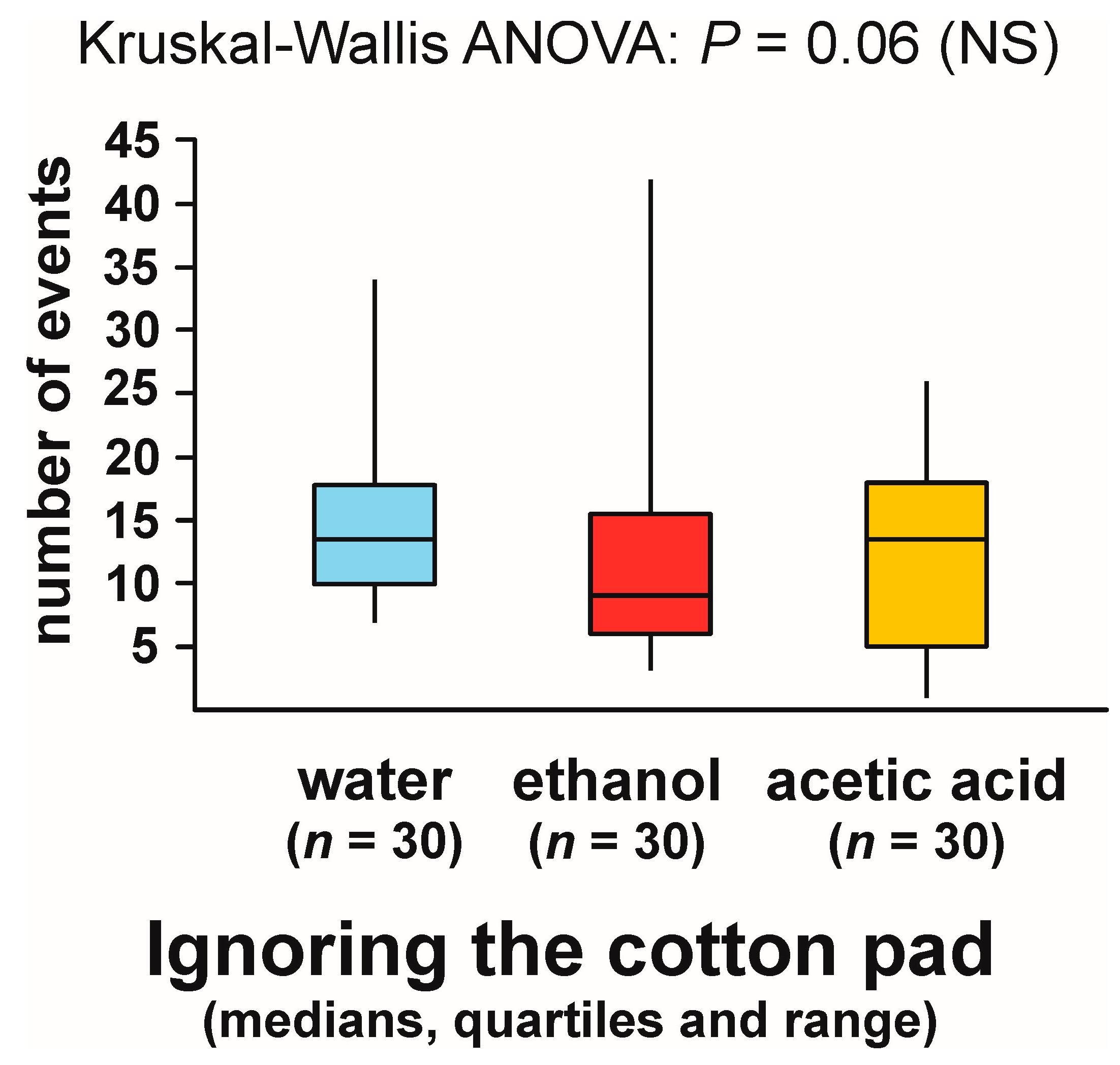
3.2. Ignoring the Experimental Object
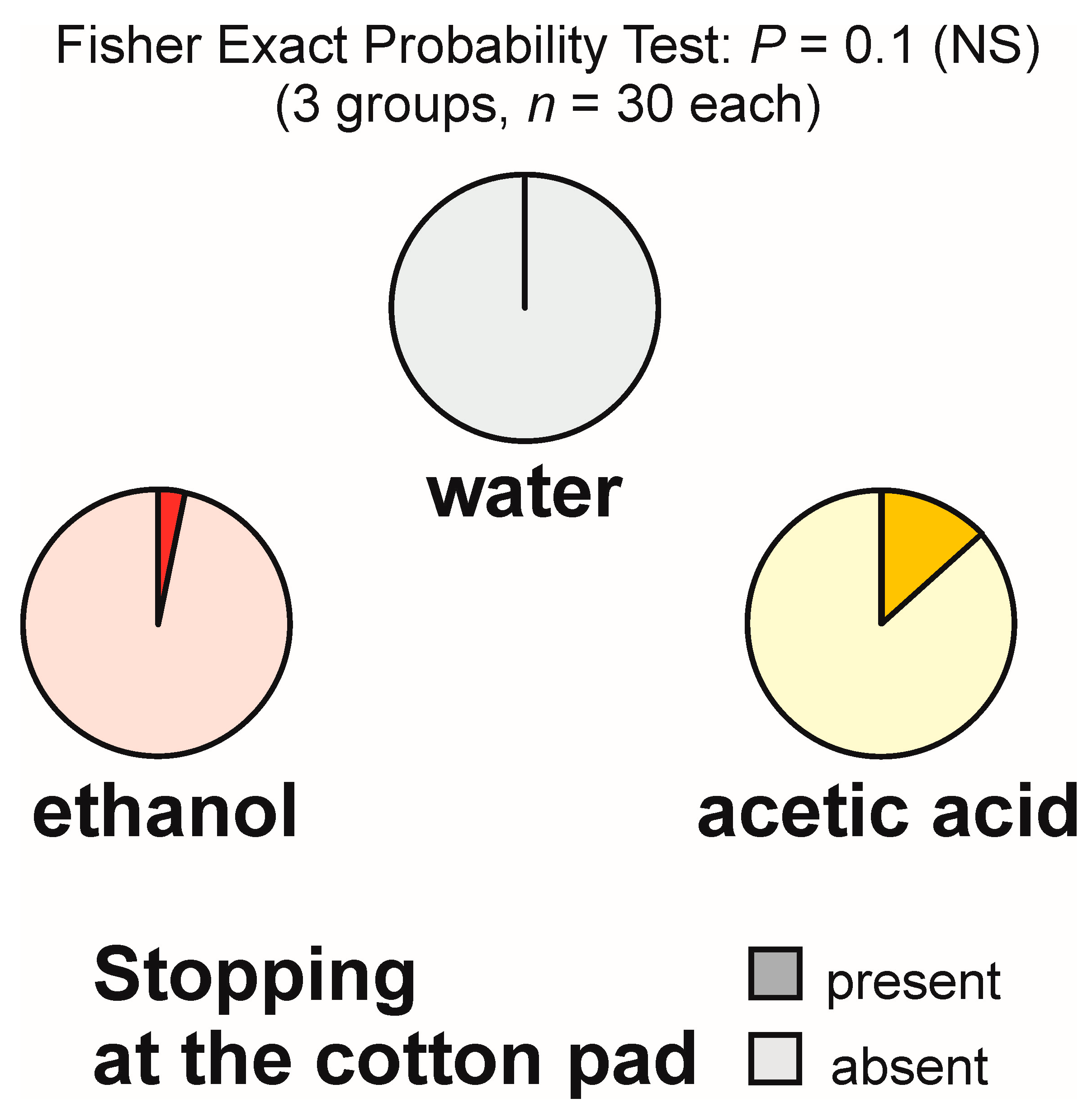
3.3. Stopping at the Experimental Object
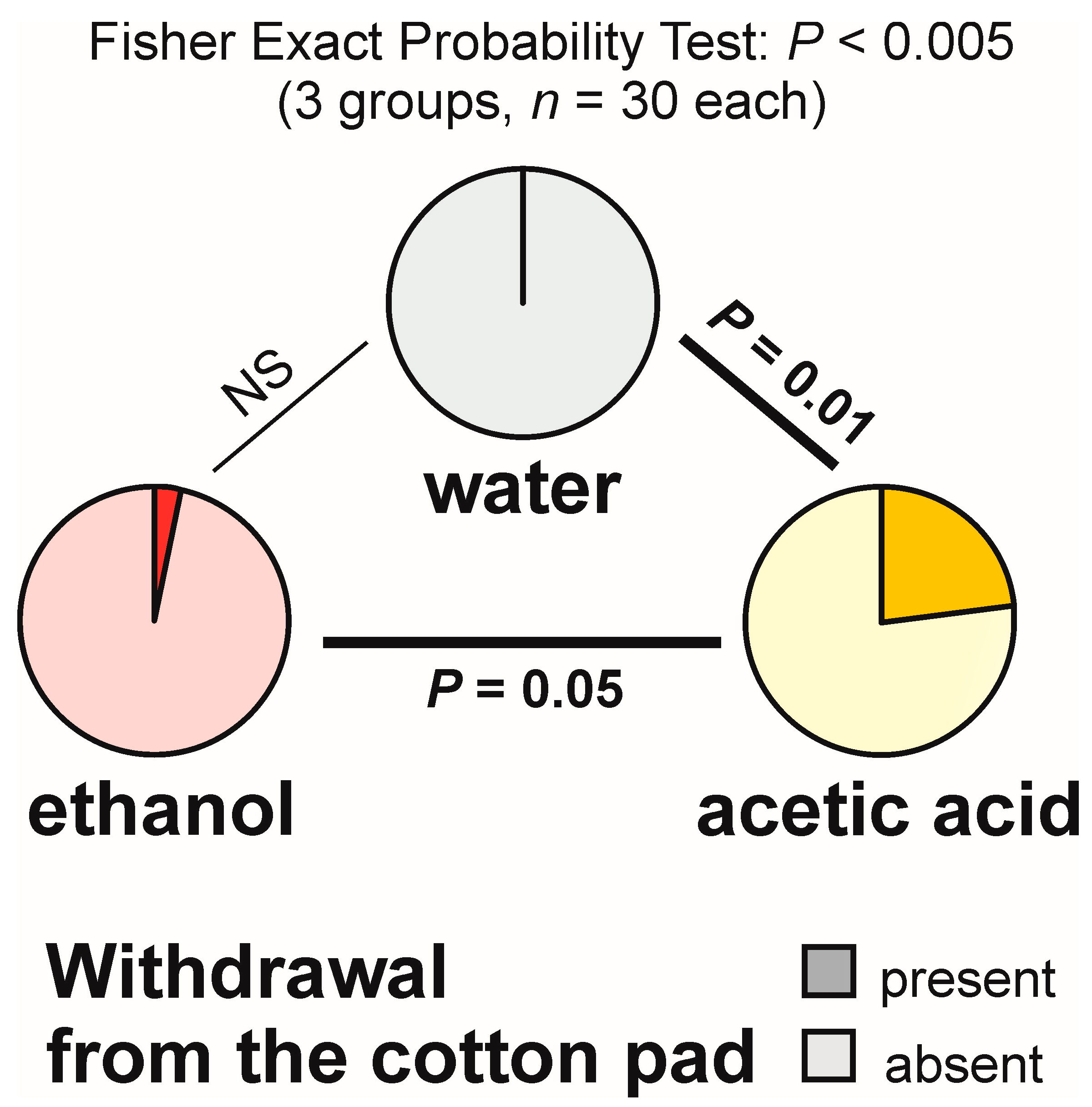
3.4. Withdrawal from the Experimental Object
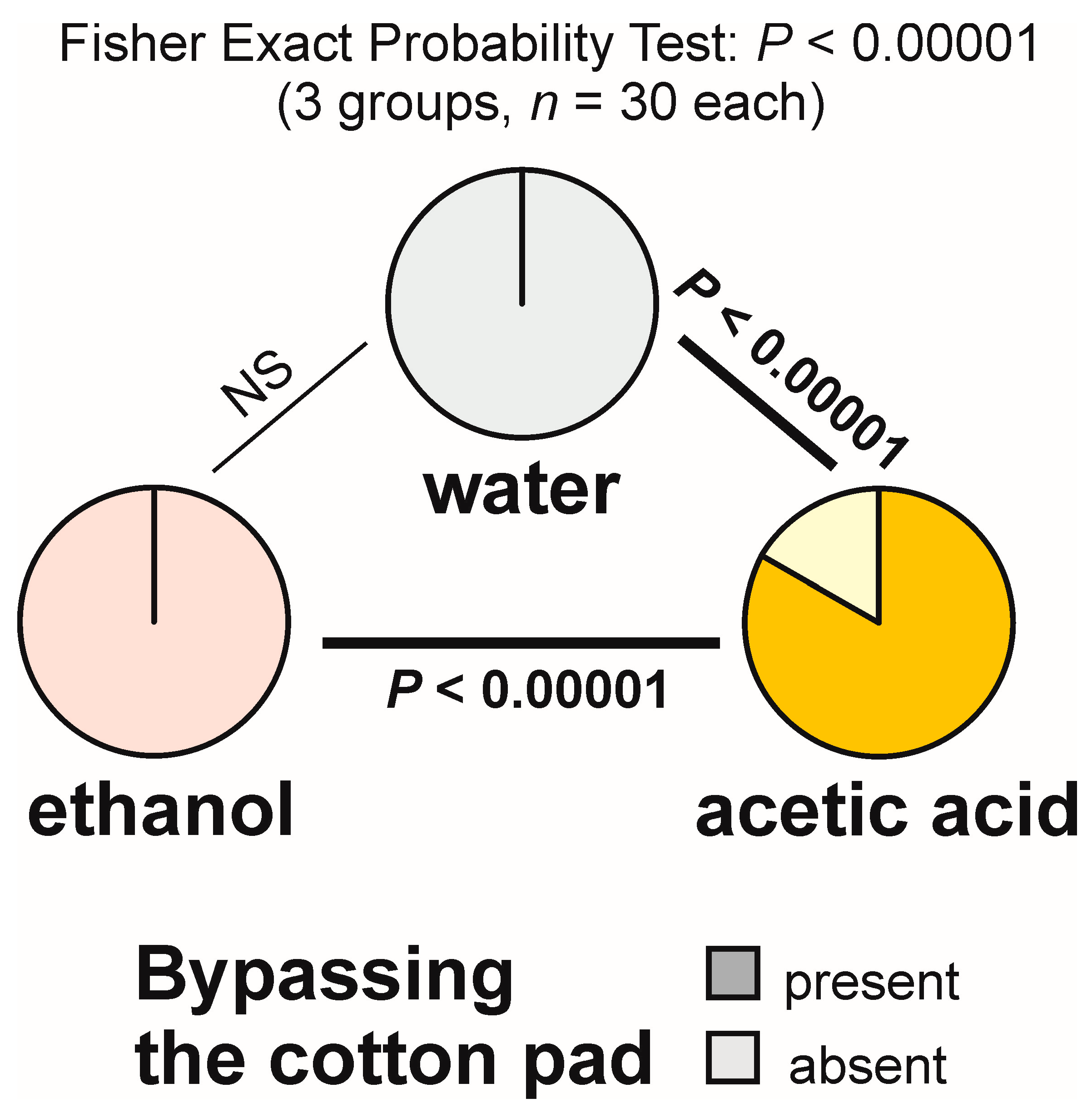
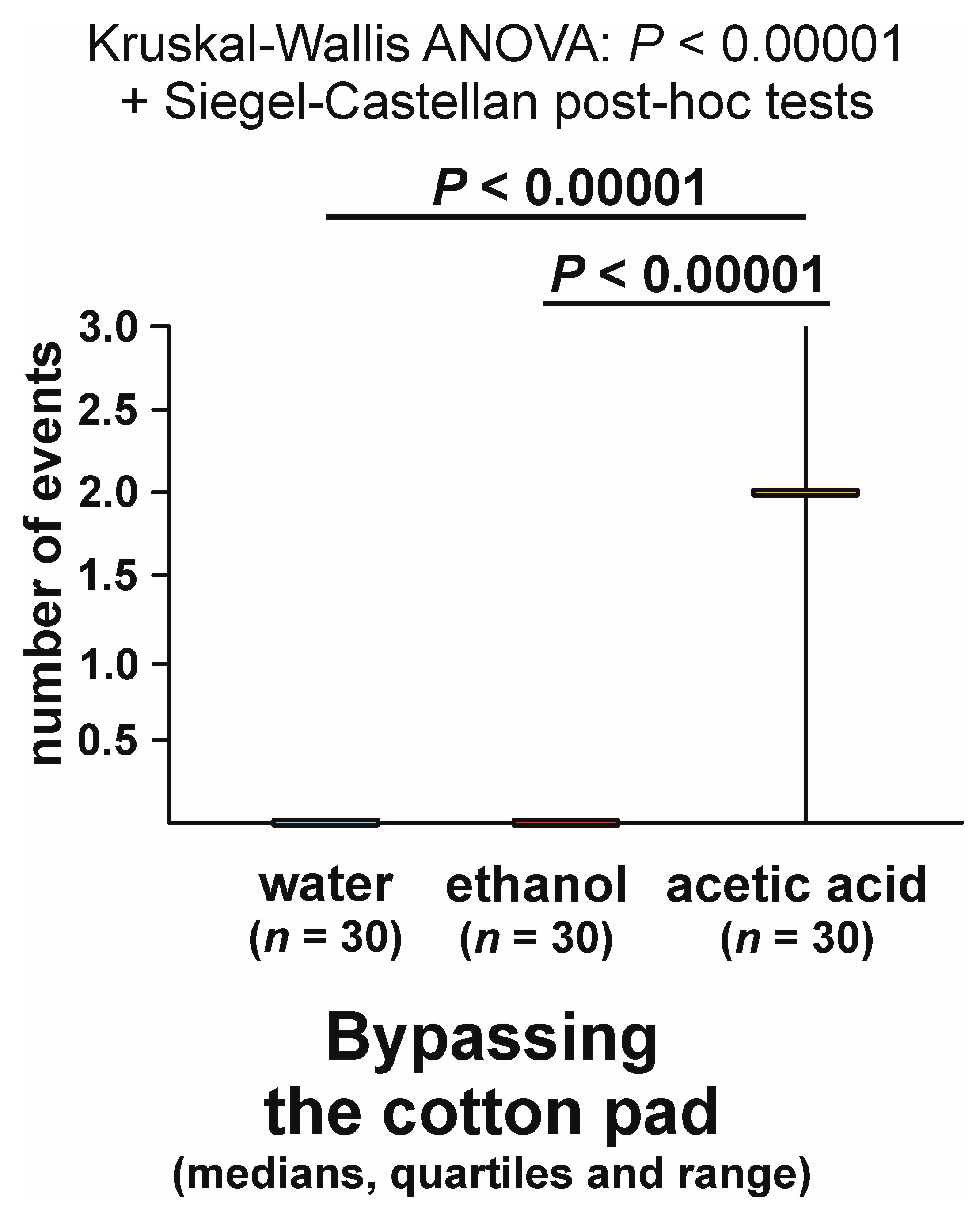
3.5. Bypassing the Experimental Object
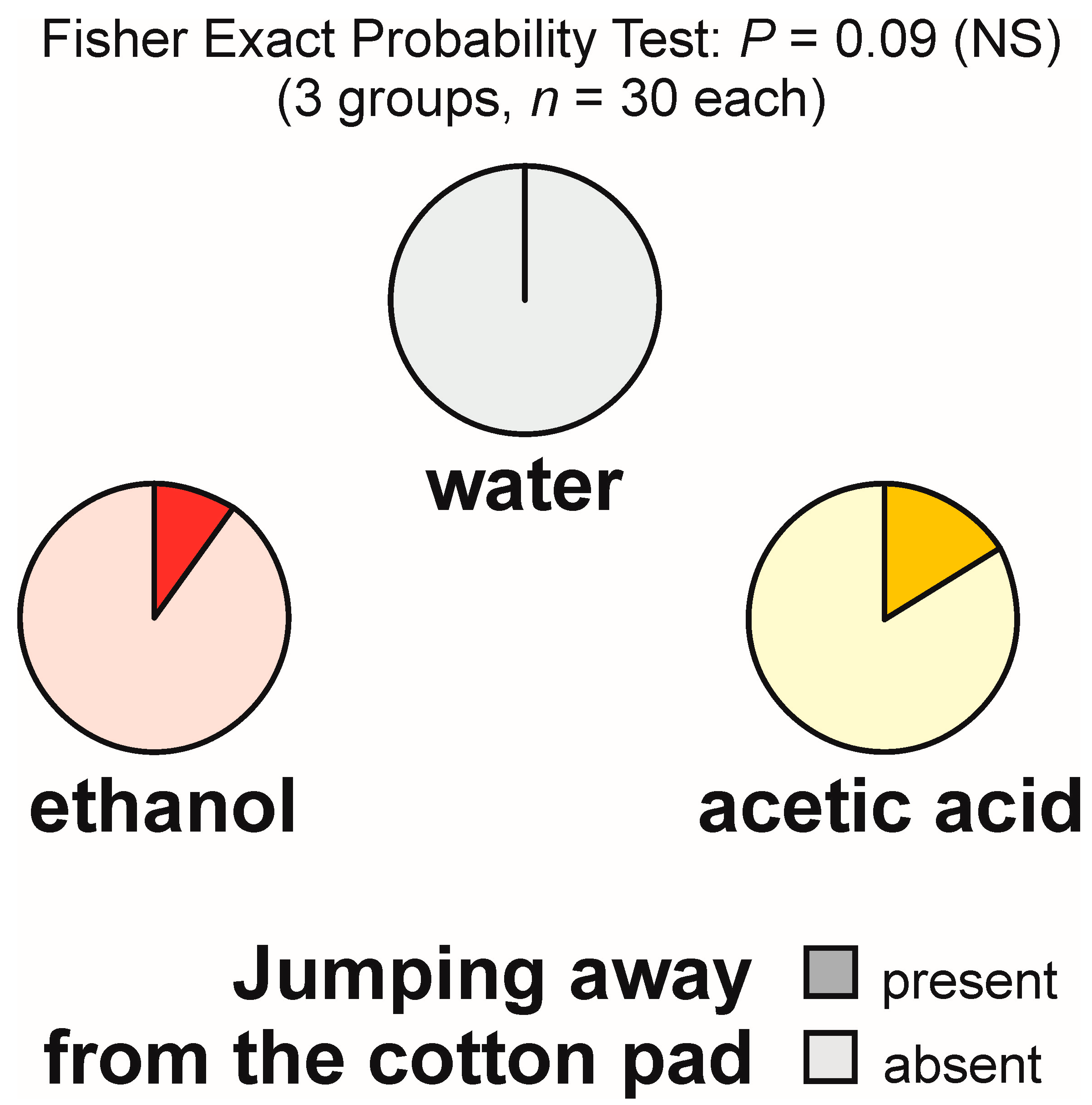
3.6. Jumping Away from the Experimental Object
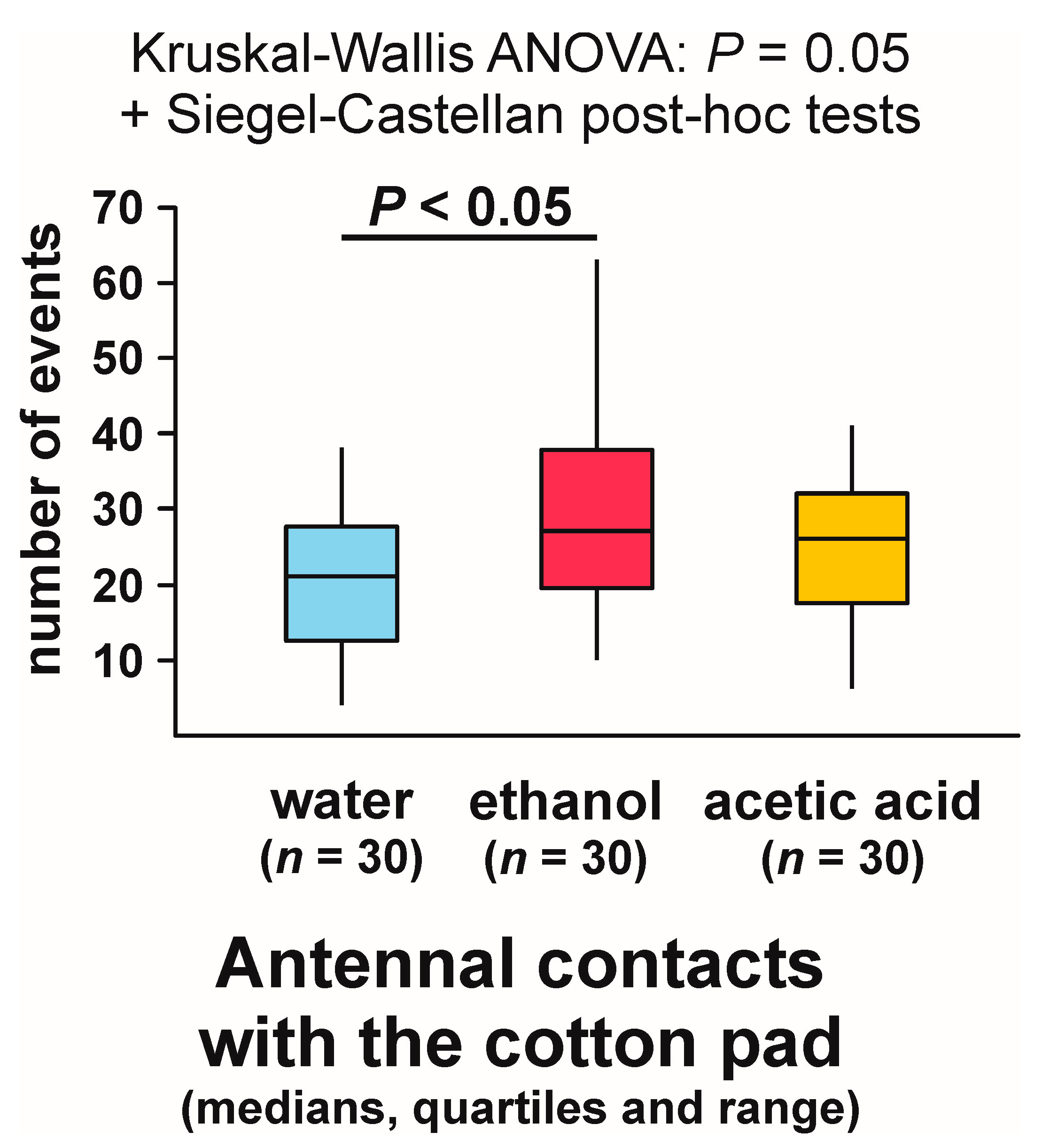
3.7. Antennal Contacts with the Experimental Object
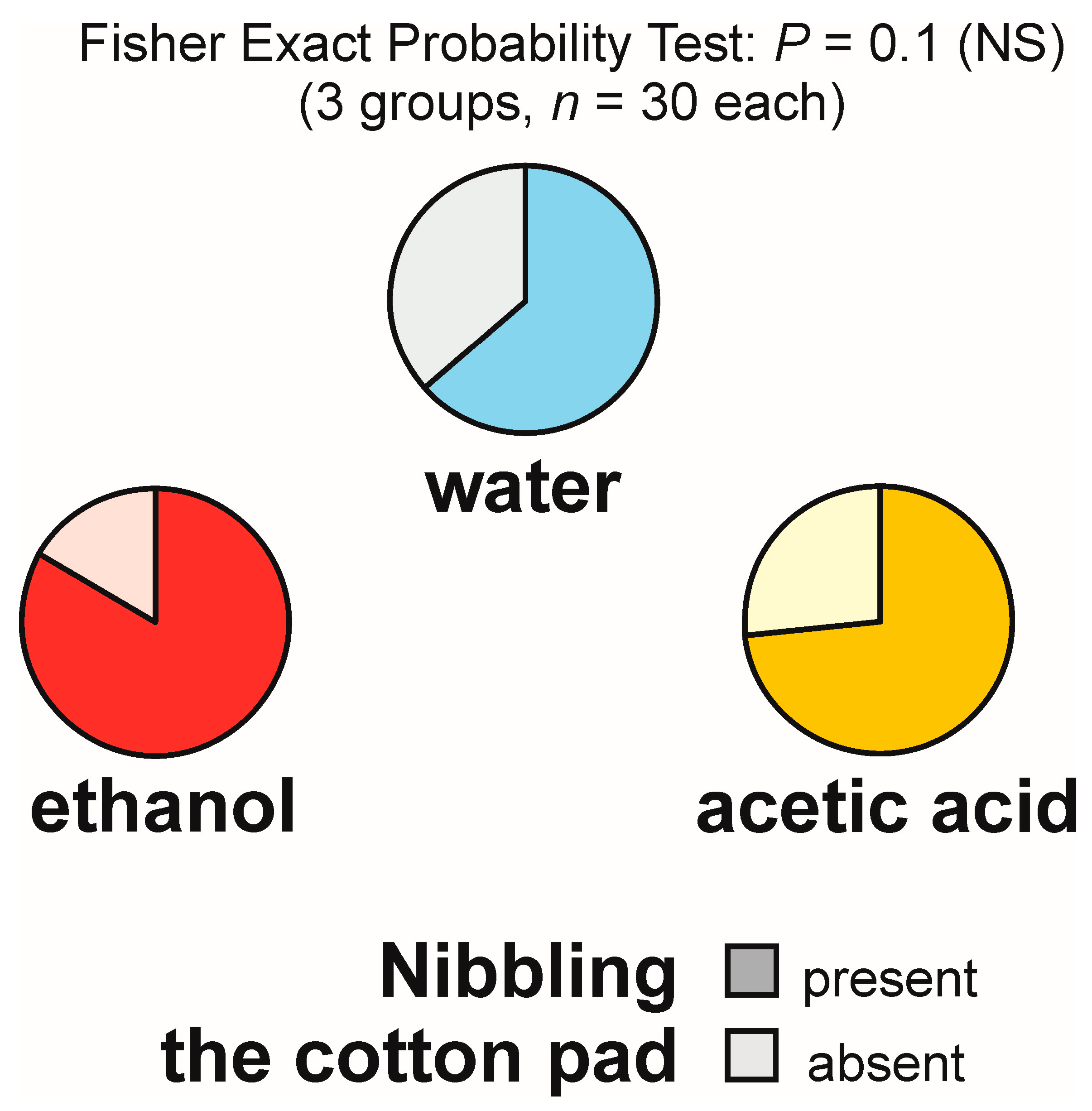
3.8. Nibbling of the Experimental Object (Not Accompanied or Accompanied by Gaster Flexing) and Nibbling of the Substrate in Its Vicinity
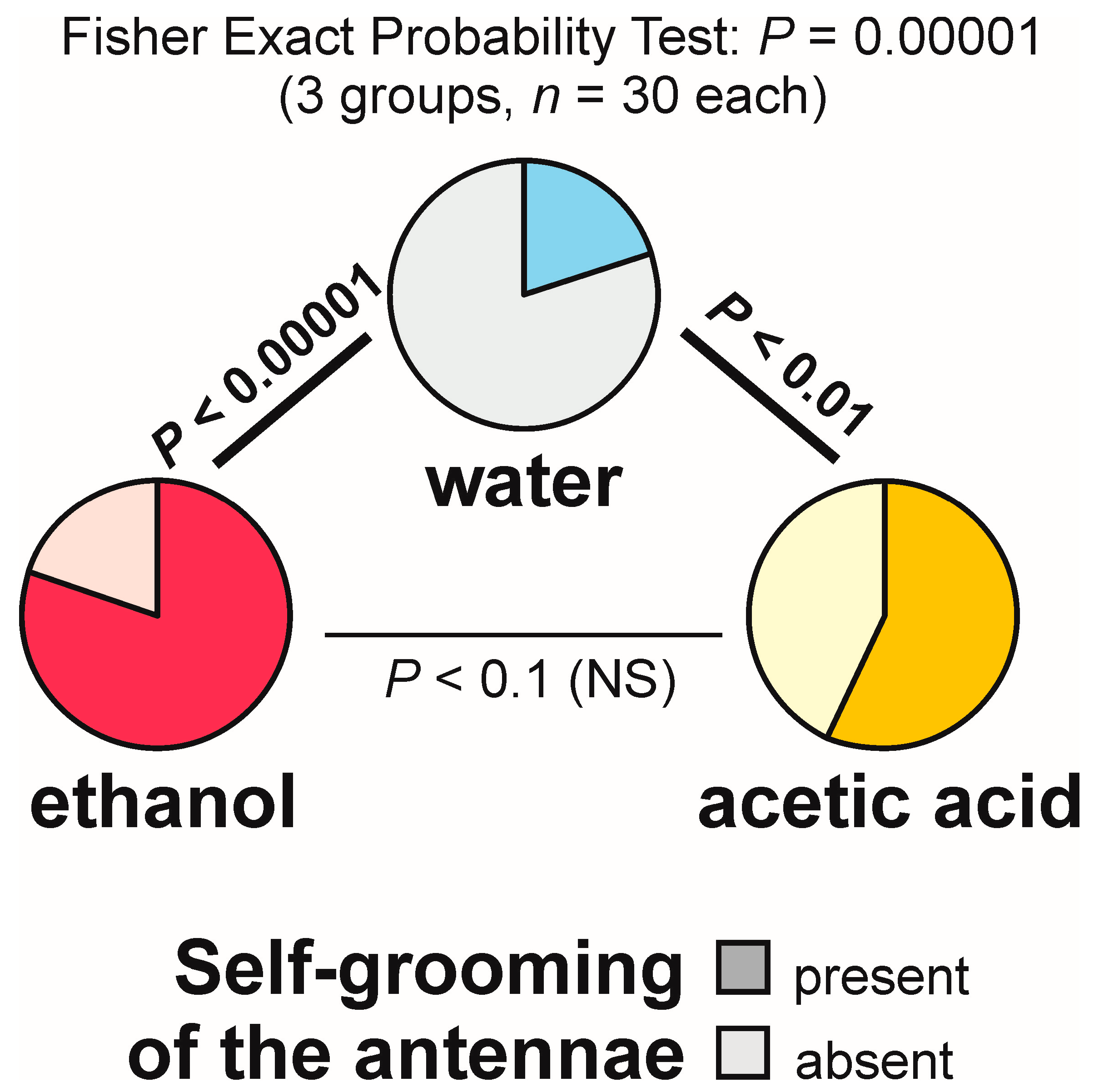
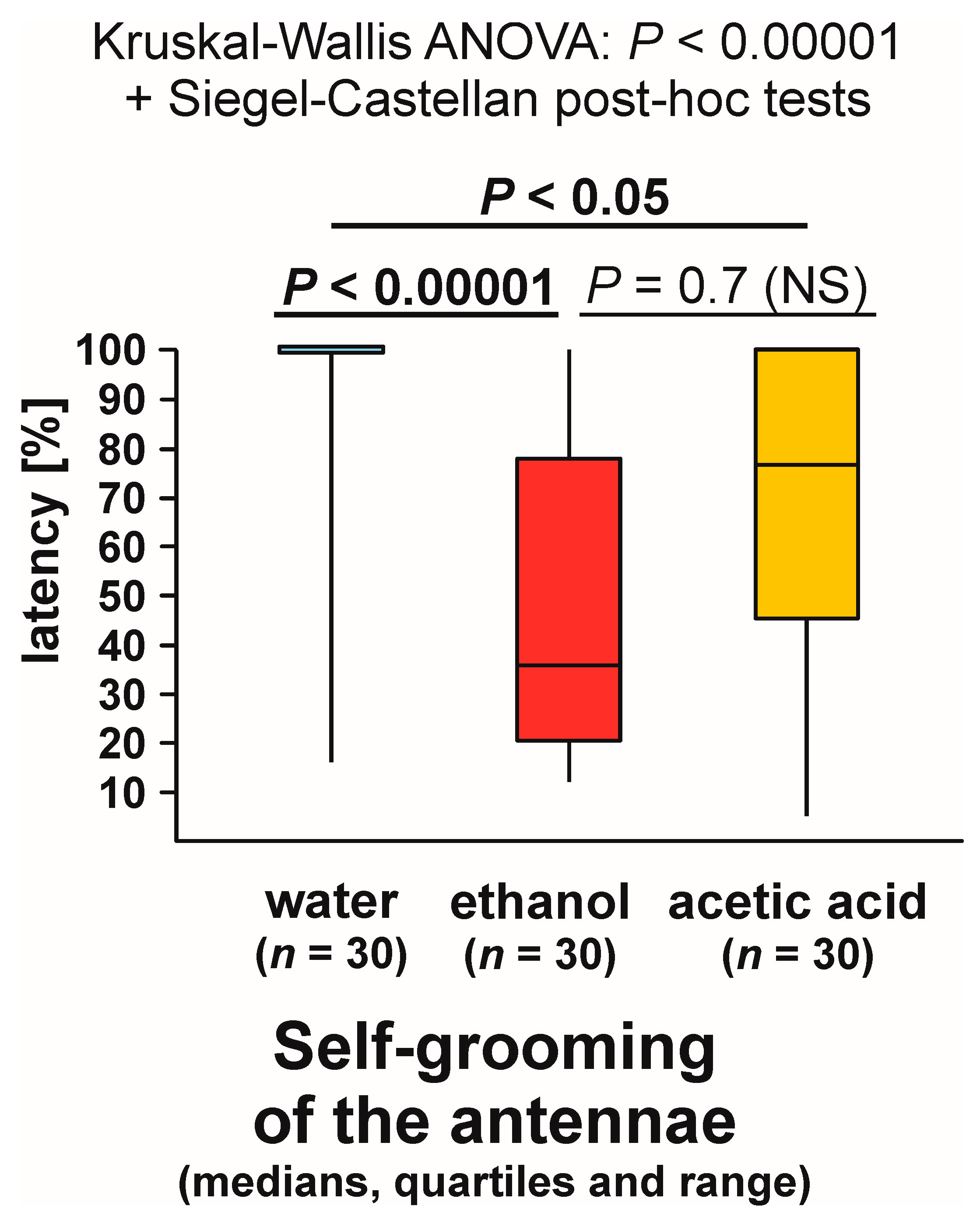
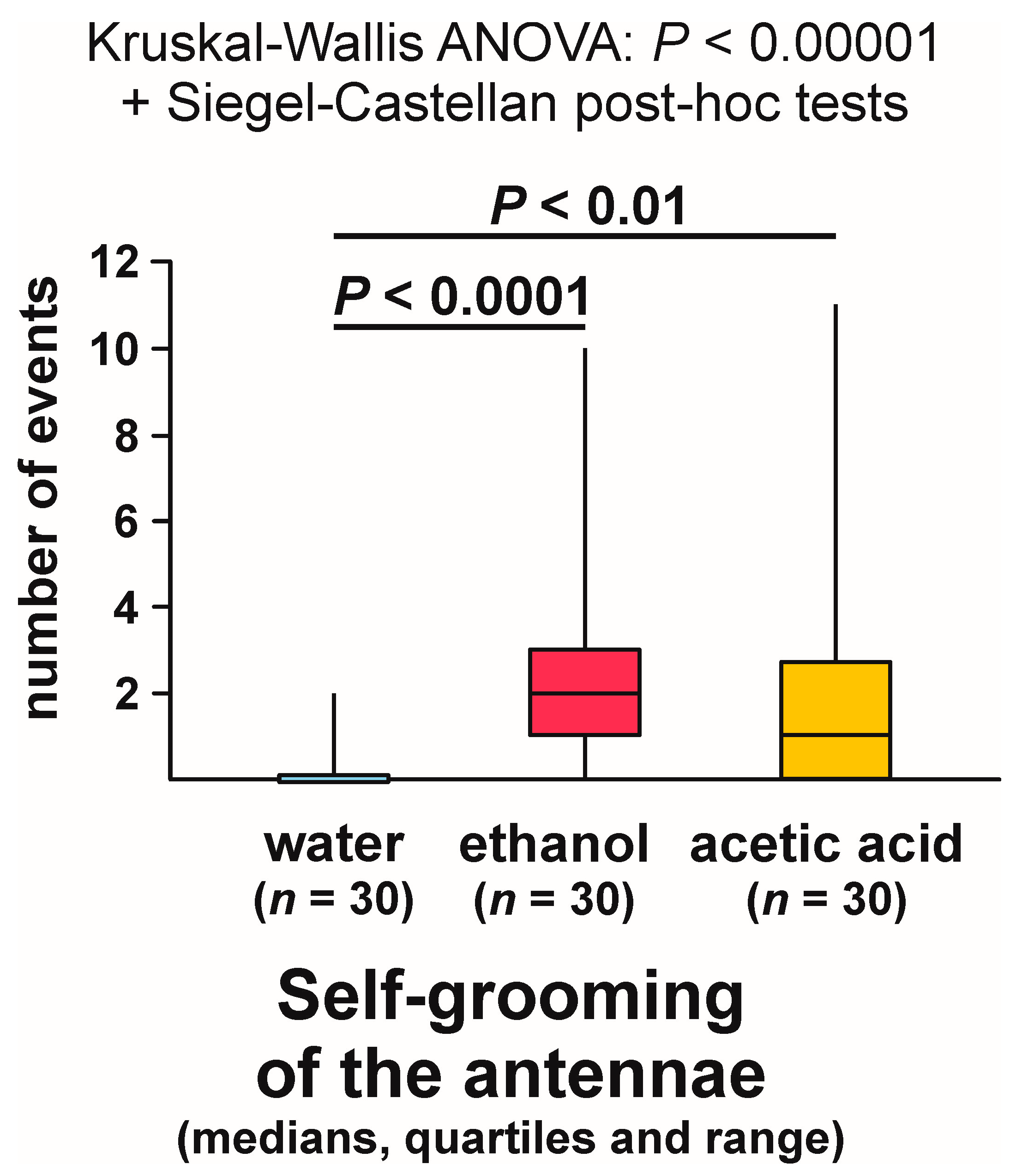
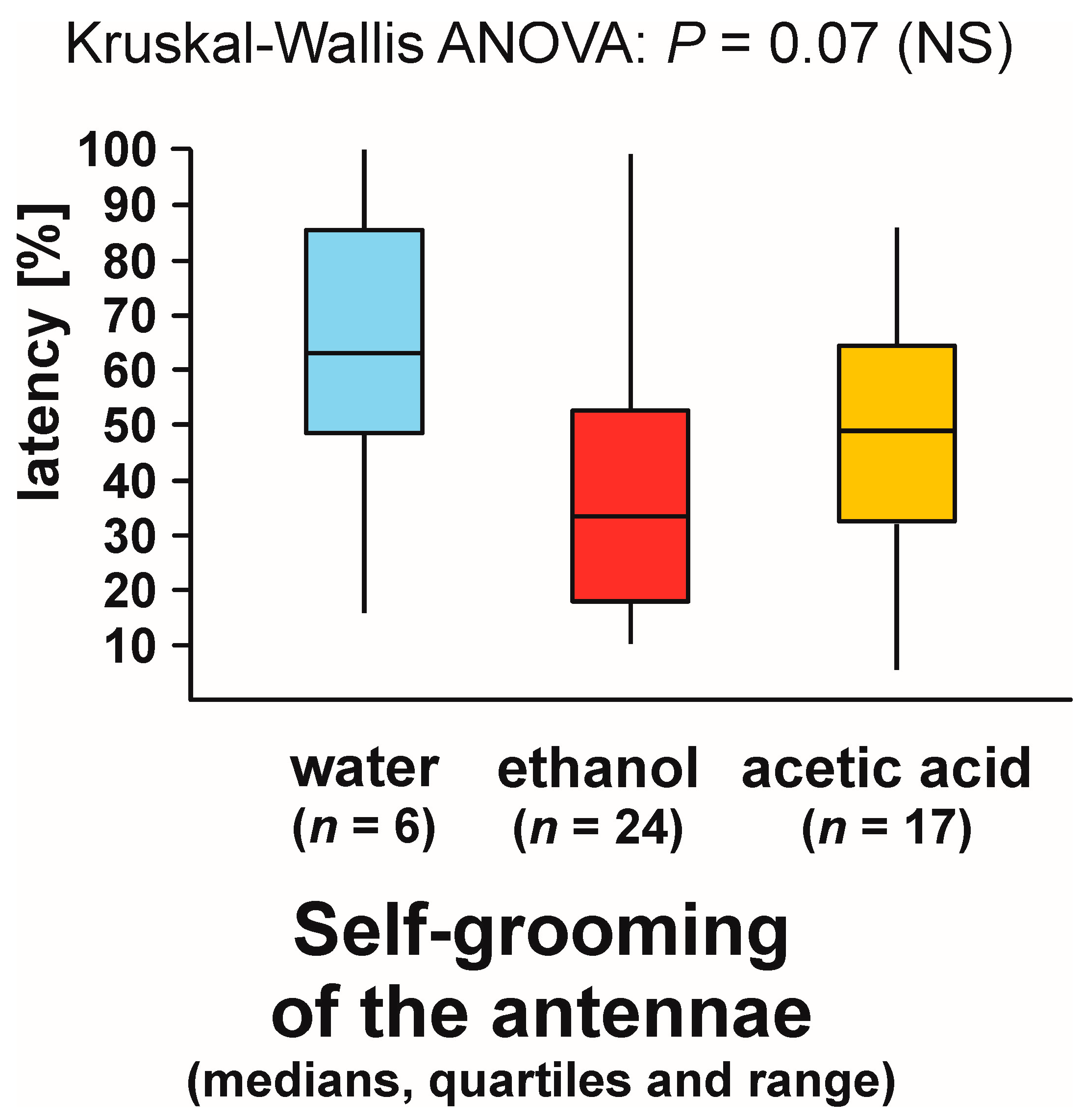
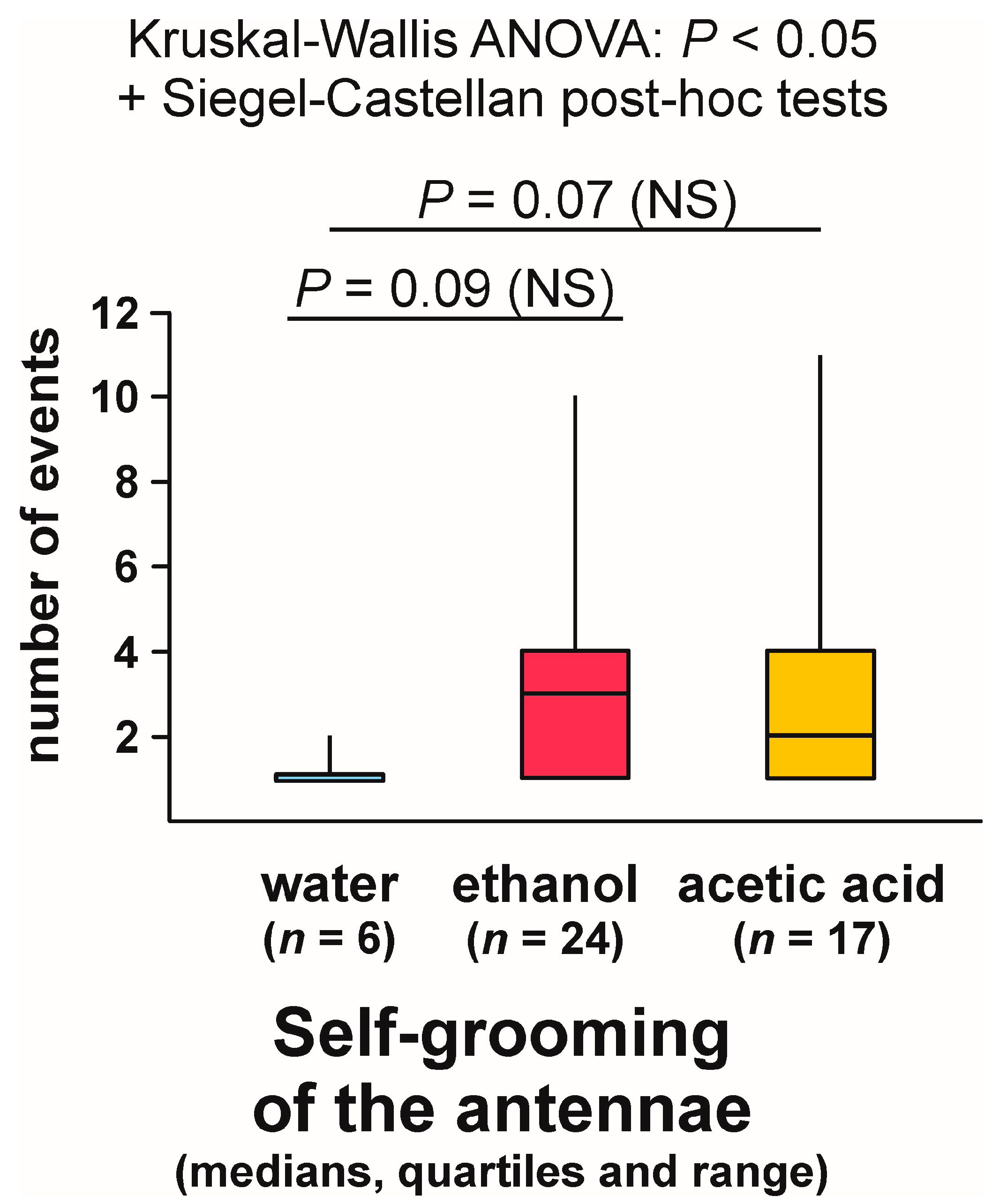
3.9. Self-grooming Behaviour Displayed Close to the Experimental Object
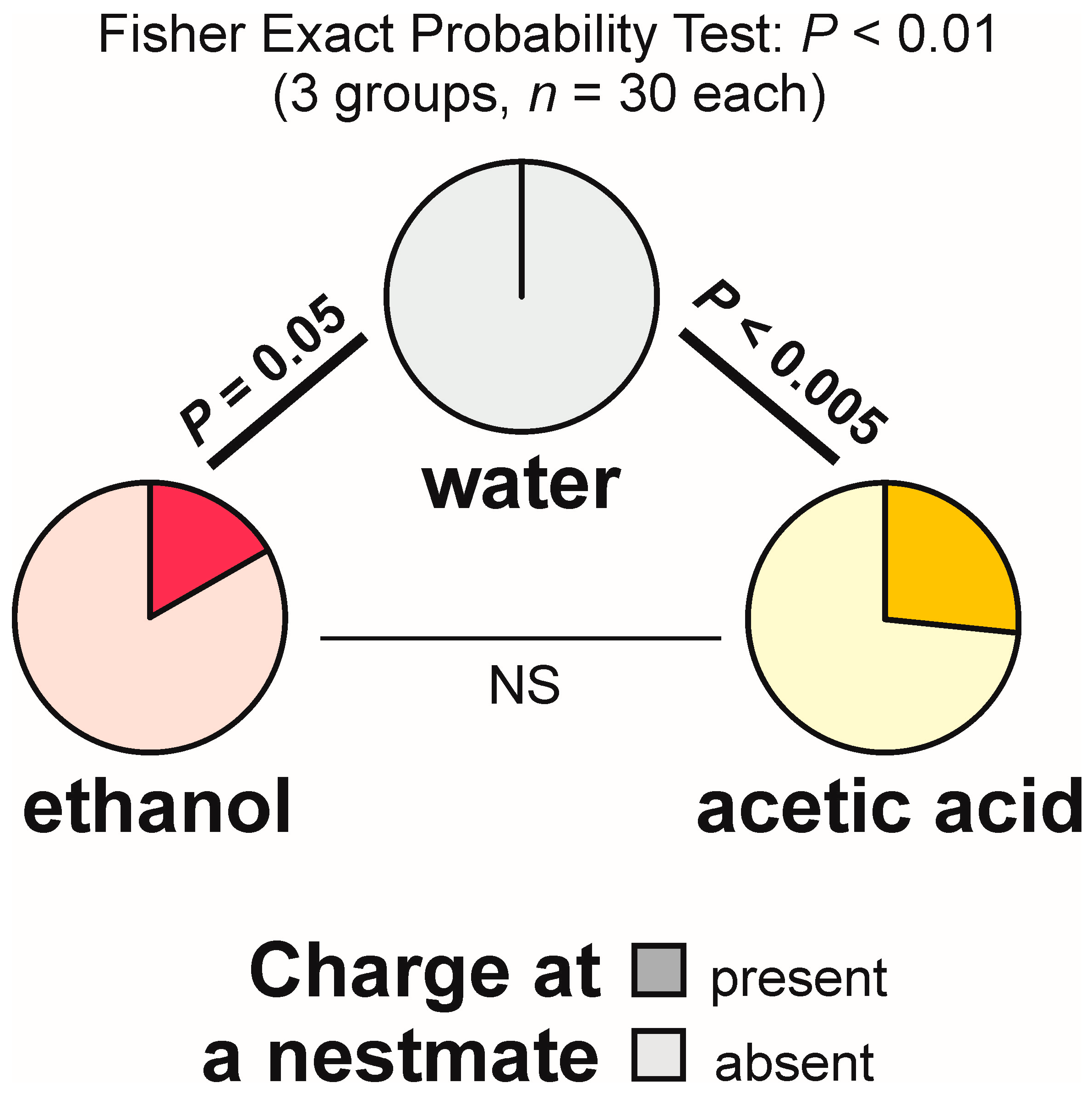
3.10. Aggressive Behaviour Directed at a Nestmate Close to the Experimental Object
3.11. Friendly Social Behaviour Directed at a Nestmate Close to the Experimental Object
4. Discussion
4.1. The Effects of Exposure to Water, Ethanol and Acetic Acid on Behaviour of the Tested Ants
4.2. Differences between the Effects of Ethanol and Acetic Acid
4.3. The Effects of Exposure to Different Experimental Compounds on Behaviour Patterns Fulfilling Similar Functions
4.4. Ethanol in Ant Exocrine Glands and Its Role in the Mediation of Ant Behaviour
4.5. Acetic Acid in Ant Exocrine Glands and Its Role in the Mediation of Ant Behaviour
4.6. Limitations of the Present Work and Perspectives of Future Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nestler, E.J.; Hyman, E.S. Animal models of neuropsychiatric disorders. Nat. Neurosci. 2010, 13, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Kramer, J.; Dick, D.M.; King, A.; Ray, A.L.; Sher, K.J.; Vena, A.; Vendruscolo, L.F.; Acion, L. Mechanisms of Alcohol Addiction: Bridging Human and Animal Studies. Alcohol Alcohol. 2020, 55, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Petković, A.; Chaudhury, D. Encore: Behavioural animal models of stress, depression and mood disorders. Front. Behav. Neurosci. 2022, 16, 931964. [Google Scholar] [CrossRef]
- Phillips, K.A.; Bales, K.L.; Capitanio, J.P.; Conley, A.; Czoty, P.W.; Hart, B.A.‘t.; Hopkins, W.D.; Hu, S.-L.; Miller, L.A.; Nader, M.A.; et al. Why primate models matter. Am. J. Primatol. 2014, 76, 801–827. [Google Scholar] [CrossRef]
- Czéh, B.; Fuchs, E.; Wiborg, O.; Simon, M. Animal models of major depression and their clinical implications. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 64, 293–310. [Google Scholar] [CrossRef]
- Moulin, T.C.; Covill, L.E.; Itskov, P.M.; Williams, M.J.; Schiöth, H.B. Rodent and fly models in behavioral neuroscience: An evaluation of methodological advances, comparative research, and future perspectives. Neurosci. Biobehav. Rev. 2021, 120, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Spence, R.; Gerlach, G.; Lawrence, C.; Smith, C. The behaviour and ecology of the zebrafish, Danio rerio. Biol. Rev. Camb. Philos. Soc. 2008, 83, 13–34. [Google Scholar] [CrossRef]
- Klee, E.W.; Schneider, H.; Clark, K.J.; Cousin, M.A.; Ebbert, J.O.; Hooten, W.M.; Karpyak, V.M.; Warner, D.O.; Ekker, S.C. Zebrafish: A model for the study of addiction genetics. Hum. Genet. 2012, 131, 977–1008. [Google Scholar] [CrossRef]
- Abramson, C.; Wells, H.; Božič, J. A social insect model for the study of ethanol induced behavior: The honey bee. In Trends in Alcohol Abuse and Alcoholism Research; Yoshida, R., Ed.; Nova Science: New York, NY, USA, 2007; pp. 197–218. [Google Scholar]
- Kaun, K.R.; Devineni, A.V.; Heberlein, U. Drosophila melanogaster as a model to study drug addiction. Hum. Genet. 2012, 131, 959–975. [Google Scholar] [CrossRef]
- Devineni, A.V.; Heberlein, U. The Evolution of Drosophila melanogaster as a Model for Alcohol Research. Annu. Rev. Neurosci. 2013, 36, 121–138. [Google Scholar] [CrossRef]
- Scholz, H.; Mustard, J.A. Invertebrate models of alcoholism. In Behavioral Neurobiology of Alcohol Addiction. Current Topics in Behavioral Neurosciences; Sommer, W., Spanagel, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 13, pp. 433–457. [Google Scholar]
- Søvik, E.; Barron, A.B. Invertebrate Models in Addiction Research. Brain Behav. Evol. 2013, 82, 153–165. [Google Scholar] [CrossRef]
- Narayanan, A.S.; Rothenfluh, A. I Believe I Can Fly!: Use of Drosophila as a Model Organism in Neuropsychopharmacology Research. Neuropsychopharmacology 2016, 41, 1439–1446. [Google Scholar] [CrossRef] [PubMed]
- Rehm, J.; Mathers, C.; Popova, S.; Thavorncharoensap, M.; Teerawattananon, Y.; Patra, J. Global burden of disease and injury and economic cost attributable to alcohol use and alcohol-use disorders. Lancet 2009, 373, 2223–2233. [Google Scholar] [CrossRef]
- Heberlein, U. Genetics of Alcohol-Induced Behaviors in Drosophila. Alcohol Res. Health J. Natl. Inst. Alcohol Abus. Alcohol. 2000, 24, 185–188. [Google Scholar]
- Chvilicek, M.M.; Titos, I.; Rothenfluh, A. The Neurotransmitters Involved in Drosophila Alcohol-Induced Behaviors. Front. Behav. Neurosci. 2020, 14, 607700. [Google Scholar] [CrossRef] [PubMed]
- Cadieu, N.; El Ghadraoui, L.; Cadieu, J.-C. Egg-laying preference for ethanol involving learning has adaptive significance in Drosophila melanogaster. Anim. Learn. Behav. 2000, 28, 187–194. [Google Scholar] [CrossRef]
- Devineni, A.V.; Heberlein, U. Preferential Ethanol Consumption in Drosophila Models Features of Addiction. Curr. Biol. 2009, 19, 2126–2132. [Google Scholar] [CrossRef] [PubMed]
- Park, A.; Tran, T.; Gutierrez, L.; Stojanik, C.J.; Plyler, J.; Thompson, G.A.; Bohm, R.A.; Scheuerman, E.A.; Smith, D.P.; Atkinson, N.S. Alcohol-induced aggression in Drosophila. Addict. Biol. 2021, 26, e13045. [Google Scholar] [CrossRef]
- Abramson, C.I.; Stone, S.M.; Ortez, R.A.; Luccardi, A.; Vann, K.L.; Hanig, K.D.; Rice, J. The development of an ethanol model using social insects: I: Behavior studies of the honey bee (Apis mellifera L.). Alcohol. Clin. Exp. Res. 2000, 24, 1153–1166. [Google Scholar] [CrossRef]
- Abramson, C.I.; Fellows, G.W.; Browne, B.L.; Lawson, A.; Ortiz, R.A. Development of an Ethanol Model Using Social Insects: II. Effect of Antabuse® on Consumatory Responses and Learned Behavior of the Honey Bee (Apis mellifera L.). Psychol. Rep. 2003, 92, 365–378. [Google Scholar] [CrossRef]
- Abramson, C.I.; Sheridan, A.; Donohue, D.; Kandolf, A.; Božič, J.; Meyers, J.E.; Benbassat, D. Development of an Ethanol Model Using Social Insects: III. Preferences for Ethanol Solutions. Psychol. Rep. 2004, 94, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Abramson, C.I.; Place, A.J.; Aquino, I.S.; Fernandez, A. Development of an ethanol model using social insects: IV. Influence of ethanol on the aggression of Africanized honey bees (Apis mellifera L.). Psychol. Rep. 2004, 94, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Abramson, C.I.; Sanderson, C.; Painter, J.; Barnett, S.; Wells, H. Development of an ethanol model using social insects: V. Honeybee foraging decisions under the influence of alcohol. Alcohol 2005, 36, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Abramson, C.I.; Craig, D.P.A.; Varnon, C.A.; Wells, H. The effect of ethanol on reversal learning in honey bees (Apis mellifera anatolica): Response inhibition in a social insect model. Alcohol 2015, 49, 245–258. [Google Scholar] [CrossRef]
- Bozic, J.; Abramson, C.I.; Bedencic, M. Reduced ability of ethanol drinkers for social communication in honeybees (Apis mellifera carnica Poll.). Alcohol 2006, 38, 179–183. [Google Scholar] [CrossRef]
- Maze, I.S.; Wright, G.A.; Mustard, J.A. Acute ethanol ingestion produces dose-dependent effects on motor behavior in the honey bee (Apis mellifera). J. Insect Physiol. 2006, 52, 1243–1253. [Google Scholar] [CrossRef]
- Mustard, J.A.; Edgar, E.A.; Mazade, R.E.; Wu, C.; Lillvis, J.L.; Wright, G.A. Acute ethanol ingestion impairs appetitive olfactory learning and odor discrimination in the honey bee. Neurobiol. Learn. Mem. 2008, 90, 633–643. [Google Scholar] [CrossRef]
- Wright, G.A.; Lillvis, J.L.; Bray, H.J.; Mustard, J.A. Physiological State Influences the Social Interactions of Two Honeybee Nest Mates. PLoS ONE 2012, 7, e32677. [Google Scholar] [CrossRef]
- Giannoni-Guzmán, M.A.; Giray, T.; Agosto-Rivera, J.L.; Stevison, B.K.; Freeman, B.; Ricci, P.; Brown, E.A.; Abramson, C.I. Ethanol-Induced Effects on Sting Extension Response and Punishment Learning in the Western Honey Bee (Apis mellifera). PLoS ONE 2014, 9, e100894. [Google Scholar] [CrossRef]
- Ostap-Chec, M.; Opalek, M.; Stec, D.; Miler, K. Discontinued alcohol consumption elicits withdrawal symptoms in honeybees. Biol. Lett. 2021, 17, 20210182. [Google Scholar] [CrossRef]
- Stephenson, L.; Chicas-Mosier, A.; Black, T.; Wells, H.; Abramson, C. Inducing Ethanol Tolerance in Free-Flying Honey Bees (Apis mellifera L.). Int. J. Comp. Psychol. 2021, 34, 1–13. [Google Scholar] [CrossRef]
- Ahmed, I.; Abramson, C.I.; Faruque, I.A. Honey bee flights near hover under ethanol-exposure show changes in body and wing kinematics. PLoS ONE 2022, 17, e0278916. [Google Scholar] [CrossRef]
- Cohan, F.M.; Graf, J.D. Latitudinal cline in Drosophila melanogaster for knockdown resistance to ethanol fumes and for rates of response to selection for further resistance. Evolution 1985, 39, 278–293. [Google Scholar] [CrossRef]
- Cohan, F.M.; Hoffmann, A.A. Genetic Divergence under Uniform Selection. II. Different Responses to Selection for Knockdown Resistance to Ethanol among Drosophila Melanogaster Populations and Their Replicate Lines. Genetics 1986, 114, 145–164. [Google Scholar] [CrossRef]
- Weber, K.E. An apparatus for measurement of resistance to gas-phase agents. Drosoph. Inf. Serv. 1988, 67, 91–93. [Google Scholar]
- Weber, K.E.; Diggins, L.T. Increased selection response in larger populations. II. Selection for ethanol vapor resistance in Drosophila melanogaster at two population sizes. Genetics 1990, 125, 585–597. [Google Scholar] [CrossRef]
- Singh, C.M.; Heberlein, U. Genetic control of acute ethanol-induced behaviors in Drosophila. Alcohol. Clin. Exp. Res. 2000, 24, 1127–1136. [Google Scholar] [CrossRef]
- Moore, M.S.; DeZazzo, J.; Luk, A.Y.; Tully, T.; Singh, C.M.; Heberlein, U. Ethanol Intoxication in Drosophila: Genetic and Pharmacological Evidence for Regulation by the cAMP Signaling Pathway. Cell 1998, 93, 997–1007. [Google Scholar] [CrossRef]
- Rothenfluh, A.; Threlkeld, R.J.; Bainton, R.J.; Tsai, L.T.-Y.; Lasek, A.W.; Heberlein, U. Distinct Behavioral Responses to Ethanol Are Regulated by Alternate RhoGAP18B Isoforms. Cell 2006, 127, 199–211. [Google Scholar] [CrossRef]
- Neckameyer, W.S.; Nieto-Romero, A.R. Response to stress in Drosophila is mediated by gender, age and stress paradigm. Stress 2015, 18, 254–266. [Google Scholar] [CrossRef]
- Ammons, A.D.; Hunt, G.J. Characterization of honey bee sensitivity to ethanol vapor and its correlation with aggression. Alcohol 2008, 42, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Miler, K.; Kuszewska, K.; Privalova, V.; Woyciechowski, M. Honeybees show adaptive reactions to ethanol exposure. Sci. Rep. 2018, 8, 8707. [Google Scholar] [CrossRef]
- Miler, K.; Opalek, M.; Ostap-Chec, M.; Stec, D. Diel rhythmicity of alcohol-induced intoxication in the honeybee workers. J. Zool. 2021, 314, 96–103. [Google Scholar] [CrossRef]
- Miler, K.; Stec, D.; Kamińska, A.; Pardyak, L.; Kuszewska, K. Alcohol intoxication resistance and alcohol dehydrogenase levels differ between the honeybee castes. Apidologie 2021, 52, 230–241. [Google Scholar] [CrossRef]
- Miler, K.; Stec, D.; Pardyak, L.; Kamińska, A.; Kuszewska, K. No increase in alcohol dehydrogenase levels following repeated ethanol exposure in young honeybee workers. Physiol. Entomol. 2022, 47, 110–116. [Google Scholar] [CrossRef]
- Robinson, B.G.; Khurana, S.; Pohl, J.B.; Li, W.-K.; Ghezzi, A.; Cady, A.M.; Najjar, K.; Hatch, M.M.; Shah, R.R.; Bhat, A.; et al. A Low Concentration of Ethanol Impairs Learning but Not Motor and Sensory Behavior in Drosophila Larvae. PLoS ONE 2012, 7, e37394. [Google Scholar] [CrossRef] [PubMed]
- Shohat-Ophir, G.; Kaun, K.R.; Azanchi, R.; Mohammed, H.; Heberlein, U. Sexual Deprivation Increases Ethanol Intake in Drosophila. Science 2012, 335, 1351–1355. [Google Scholar] [CrossRef] [PubMed]
- Meurville, M.-P.; LeBoeuf, A.C. Trophallaxis: The functions and evolution of social fluid exchange in ant colonies (Hymenoptera: Formicidae). Myrmecol. News 2021, 31, 1–30. [Google Scholar] [CrossRef]
- Kacsoh, B.Z.; Lynch, Z.R.; Mortimer, N.T.; Schlenke, T.A. Fruit Flies Medicate Offspring After Seeing Parasites. Science 2013, 339, 947–950. [Google Scholar] [CrossRef]
- Bozler, J.; Kacsoh, B.Z.; Bosco, G.; Geisel School of Medicine at Dartmouth, United States. Transgenerational inheritance of ethanol preference is caused by maternal NPF repression. eLife 2019, 8, e45391. [Google Scholar] [CrossRef]
- Schumann, I.; Berger, M.; Nowag, N.; Schäfer, Y.; Saumweber, J.; Scholz, H.; Thum, A.S. Ethanol-guided behavior in Drosophila larvae. Sci. Rep. 2021, 11, 12307. [Google Scholar] [CrossRef]
- Godzińska, E.J.; Korczyńska, J.; Szczuka, A. Dyadic Nestmate Reunion Test in The Research on Ant Social Behavior. Kosmos 2020, 68, 561–574. [Google Scholar] [CrossRef]
- Szczuka, A.; Godzińska, E.J.; Korczyńska, J. Factors Mediating Ant Social Behavior: Interplay of Neuromodulation and Social Context. Kosmos 2019, 68, 575–589. [Google Scholar] [CrossRef]
- Godzińska, E.J. Human and ant social behavior should be compared in a very careful way to draw valid parallels. Behav. Brain Sci. 2016, 39, e98. [Google Scholar] [CrossRef]
- Pisarski, B. La structure des colonies polycaliques de Formica (Coptoformica) exsecta Nyl. Ekol. Pol. 1972, 20, 111–116. [Google Scholar]
- Pisarski, B. Structure et organisation des sociétés de fourmis de l’espèce Formica (Coptoformica) exsecta Nyl. (Hymenoptera, Formicidae). Memorab. Zool. 1982, 38, 1–281. [Google Scholar]
- Dobrzanska, J. Ethological studies on polycalic colonies of the ants Formica exsecta Nyl. Acta Neurobiol. Exp. 1973, 33, 597–622. [Google Scholar]
- Czechowski, W. Competition between Formica exsecta Nyl. and Formica pressilabris Nyl. (Hymenoptera, Formicidae). Ann. Zool. 1976, 33, 273–285. [Google Scholar]
- Chudzicka, E. Développement des colonies de Formica (Coptoformica) exsecta Nyl. Memorab. Zool. 1982, 38, 205–237. [Google Scholar]
- Chudzicka, E. Les échanges d ’ouvrières dans les colonies polycaliques de Formica (Coptoformica) exsecta Nyl. Memorab. Zool. 1982, 38, 239–260. [Google Scholar]
- Czechowski, W.; Radchenko, A.; Czechowska, W.; Vepsäläinen, K. The Ants of Poland with Reference to the Myrmecofauna of Europe; Fauna Poloniae Vol. 4 New series; Natura Optima Dux Foundation: Warsaw, Poland, 2012. [Google Scholar]
- Csata, E.; Markó, B.; Erős, K.; Gál, C.; Szász-Len, A.-M.; Czekes, Z. Outstations as stable meeting points for workers from different nests in a polydomous nest system of Formica exsecta Nyl. (Hymenoptera: Formicidae). Pol. J. Ecol. 2012, 60, 177–186. [Google Scholar]
- Markó, B.; Czekes, Z.; Erős, K.; Csata, E.; Szász-Len, A.-M. The largest polydomous system of Formica ants (Hymenoptera: Formicidae) in Europe discovered thus far in Romania. North-West. J. Zool. 2012, 8, 287–291. [Google Scholar]
- Siegel, S.; Castellan, N.J. Nonparametric Statistics for the Behavioral Sciences; McGraw-Hill: New York, NY, USA, 1988. [Google Scholar]
- Wagner-Ziemka, A.; Szczuka, A.; Korczyńska, J.; Kieruzel, M.; Godzińska, E.J. Behavior of ant-workers of Aphaenogaster senilis (Hymenoptera: Formicidae) during dyadic nestmate reunion tests carried out after a period of social isolation. Sociobiology 2006, 48, 281–308. [Google Scholar]
- Szczuka, A.; Korczyńska, J.; Wnuk, A.; Symonowicz, B.; Szwacka, A.G.; Mazurkiewicz, P.; Kostowski, W.; Godzińska, E.J. The effects of serotonin, dopamine, octopamine and tyramine on behavior of workers of the ant Formica polyctena during dyadic aggression tests. Acta Neurobiol. Exp. 2013, 73, 495–520. [Google Scholar]
- Korczyńska, J.; Szczuka, A.; Symonowicz, B.; Wnuk, A.; Anna, G.S.; Mazurkiewicz, P.J.; Studnicki, M.; Godzińska, E.J. The effects of age and past and present behavioral specialization on behavior of workers of the red wood ant Formica polyctena Först. during nestmate reunion tests. Behav. Process. 2014, 107, 29–41. [Google Scholar] [CrossRef]
- Szczuka, A.; Godzińska, E.J. The effect of past and present group size on responses to prey in the ant Formica polyctena Först. Acta Neurobiol. Exp. 1997, 57, 135–150. [Google Scholar]
- Lubbock, J. Ants, Bees and Wasps: A Record of Observations on the Habits of the Social Hymenoptera; Kegan Paul: London, UK; Appleton: New York, NY, USA, 1884. [Google Scholar]
- Cammaerts-Tricot, M.-C.; Morgan, E.D.; Tyler, R.C.; Braekman, J.-C. Dufour’s gland secretion of Myrmica rubra: Chemical, electrophysiological, and ethological studies. J. Insect Physiol. 1976, 22, 927–932. [Google Scholar] [CrossRef]
- Cammaerts, M.-C.; Evershed, R.P.; Morgan, E.D. Comparative study of the mandibular gland secretion of four species of Myrmica ants. J. Insect Physiol. 1981, 27, 225–231. [Google Scholar] [CrossRef]
- Morgan, E.; Tyler, R.; Cammaerts, M. Identification of the components of Dufour gland secretion of the ant Myrmica rubra and responses to them. J. Insect Physiol. 1977, 23, 511–515. [Google Scholar] [CrossRef]
- Morgan, E.D.; Inwood, M.R.; Cammaerts, M.-C. The mandibular gland secretion of the ant, Myrmica scabrinodis. Physiol. Entomol. 1978, 3, 107–114. [Google Scholar] [CrossRef]
- Jackson, B.D.; Cammaerts, M.-C.; Morgan, E.D.; Attygalle, A.B. Chemical and behavioral studies on dufour gland contents of Manica rubida (Hymenoptera: Formicidae). J. Chem. Ecol. 1990, 16, 827–840. [Google Scholar] [CrossRef]
- Bot, A.N.M.; Ortius-Lechner, D.; Finster, K.; Maile, R.; Boomsma, J.J.; Bot, A.N.M.; Ortius-Lechner, D.; Finster, K.; Maile, R.; Boomsma, J.J.; et al. Variable sensitivity of fungi and bacteria to compounds produced by the metapleural glands of leaf-cutting ants. Insectes Sociaux 2002, 49, 363–370. [Google Scholar] [CrossRef]
- Ortius-Lechner, D.; Maile, R.; Morgan, E.D.; Boomsma, J.J. Metapleural Gland Secretion of the Leaf-cutter Ant Acromyrmex octospinosus: New Compounds and Their Functional Significance. J. Chem. Ecol. 2000, 26, 1667–1683. [Google Scholar] [CrossRef]
- Van der Heyde, H.C. Quelques observations sur la psychologie des fourmis. Arch. Néerlandaises Physiol. L’homme Animaux 1920, 4, 259–282. [Google Scholar]
- Honda, K. Defensive potential of components of the larval osmeterial secretion of papilionid butterflies against ants. Physiol. Entomol. 1983, 8, 173–179. [Google Scholar] [CrossRef]
- Pasteels, J.M.; Daloze, D.; Boeve, J.L. Aldehydic contact poisons and alarm pheromone of the ant Crematogaster scutellaris (Hymenoptera: Myrmicinae): Enzyme-mediated production from acetate precursors. J. Chem. Ecol. 1989, 15, 1501–1511. [Google Scholar] [CrossRef]
- How to Get Rid of Black Ants Inside the House. Available online: https://www.positivepest.net/how-to-get-rid-of-black-ants-inside-the-house/ (accessed on 22 July 2023).
- Natural Methods to Get Rid of Ants without Killing Them. Available online: https://medium.com/@dounyaz/natural-methods-to-get-rid-of-ants-without-killing-them-e687d5cf4efd (accessed on 22 July 2023).














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korczyńska, J.; Szczuka, A.; Urzykowska, J.; Kochanowski, M.; Andrzejczyk, N.G.; Piwowarek, K.J.; Godzińska, E.J. The Effects of Ethanol and Acetic acid on Behaviour of Extranidal Workers of the Narrow-Headed Ant Formica exsecta (Hymenoptera, Formicidae) during a Field Experiment. Animals 2023, 13, 2734. https://doi.org/10.3390/ani13172734
Korczyńska J, Szczuka A, Urzykowska J, Kochanowski M, Andrzejczyk NG, Piwowarek KJ, Godzińska EJ. The Effects of Ethanol and Acetic acid on Behaviour of Extranidal Workers of the Narrow-Headed Ant Formica exsecta (Hymenoptera, Formicidae) during a Field Experiment. Animals. 2023; 13(17):2734. https://doi.org/10.3390/ani13172734
Chicago/Turabian StyleKorczyńska, Julita, Anna Szczuka, Julia Urzykowska, Michał Kochanowski, Neptun Gabriela Andrzejczyk, Kacper Jerzy Piwowarek, and Ewa Joanna Godzińska. 2023. "The Effects of Ethanol and Acetic acid on Behaviour of Extranidal Workers of the Narrow-Headed Ant Formica exsecta (Hymenoptera, Formicidae) during a Field Experiment" Animals 13, no. 17: 2734. https://doi.org/10.3390/ani13172734