
Influence of Insoluble Dietary Fibre on Expression of Pro-Inflammatory Marker Genes in Caecum, Ileal Morphology, Performance, and Foot Pad Dermatitis in Broiler


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Diets and Feeding
2.3. Performane and Carcass Characteristics
 and two
and two  ; n = 360), close to the pen’s average BW, were selected and applied for the analysis of carcass characteristics. Of these, four birds per pen (two and two ; n = 144) were used for gizzard examination. The chickens were slaughtered without prior feed restriction by electrically stunning and exsanguination. After removing the digestive tract, the gizzard weight was captured, and digesta pH was measured directly. The remaining birds were fasted for 12 h and killed the same way on the following day and subjected to carcass analysis. Regarding foot pad dermatitis (FPD), the feet of 30 birds per box (n = 1080) were examined, evaluated, and given a score according to the visual scale emerging out of the Welfare Quality® Assessment protocol for poultry [9]. Score 0 is to be understood as “no evidence of FPD”, score 1 and 2 as “mild FPD”, and “score 3 and 4 as “severe FPD”.
; n = 360), close to the pen’s average BW, were selected and applied for the analysis of carcass characteristics. Of these, four birds per pen (two and two ; n = 144) were used for gizzard examination. The chickens were slaughtered without prior feed restriction by electrically stunning and exsanguination. After removing the digestive tract, the gizzard weight was captured, and digesta pH was measured directly. The remaining birds were fasted for 12 h and killed the same way on the following day and subjected to carcass analysis. Regarding foot pad dermatitis (FPD), the feet of 30 birds per box (n = 1080) were examined, evaluated, and given a score according to the visual scale emerging out of the Welfare Quality® Assessment protocol for poultry [9]. Score 0 is to be understood as “no evidence of FPD”, score 1 and 2 as “mild FPD”, and “score 3 and 4 as “severe FPD”.2.4. Sample Collections and Analyses
and one ; n = 72). The wet sieving and particle size distribution was examined according to Röhe et al. (2014) [12] The un-fasted and un-plucked chickens were electrically stunned and scarified by exsanguination. After removing the digestive tract, the ileal section was dissected and gently rinsed with physiological saline solution. Samples from 2 cm cranial to the ileocaecal junction were excised and stored in a 10% buffered formaldehyde solution. After 24 h of dehydration with ethanol (70%), the samples were drained, paraffin embedded, sectioned via rotary microtome (5 µm; Leica RM 2255, Leica Biosystems GmbH, Nussloch, Germany), and stained (Leica Auto Stainer XL ST5010, Leica Biosystems GmbH, Nussloch, Germany) following the protocol with Alcian blue and periodic acid–Schiff. Six well-oriented, representative villi and crypts as well as muscular layers of each sampled animal were examined with computerised light microscopy (Leica DM 600 B, Darmstadt, Germany) using the Leica Application Suit software (Leica, Version 4.12). Caecal samples were collected for the evaluation of the gene expression of the inflammation-related genes. Gene expression analyses were carried out to reach the MIQE guidelines [13]. Samples were rinsed with physiological sodium chloride solution, embedded in cryovials (2 mL, steril, Biozym Scientific GmbH, Oldendorf, Germany) immediately, frozen in liquid nitrogen, and stored at −80 °C until RNA extraction, which was carried out with TRI Reagent© solution according to the manufacturer’s protocol (Sigma Aldrich, Steinheim, Germany). To quantify concentration of each extracted RNA, a spectral analysis was carried out using a NanoDrop (ND-100 Spectrophotometer, Thermo Fisher Scientific, Waltham, MA, USA), whereas the RNA integrity was determined with a chip-based electrophoresis system (ExperionTM Automated Electrophoresis System, Bio-Rad, Hercules, CA, USA). The total RNA was reverse-transcribed with a QuantiTect Rev Transcription Kit (Qiagen, Hilden, Germany), following the manufacturer’s instructions. PCR was performed for each gene measured in triplicate on Rotor-Gene Q (Qiagen, Hilden, Germany) with SYBR Green PCR Kit (Qiagen, Hilden, Germany), following the manufacturer´s instructions. The reference genes used for normalisation in the quantitative real-time PCR (qPCR) were glyceraldehyde-3-phosphate dehydrogenase, Actin-β, and ubiquitin; target genes were interleukin 1β (IL1β), interleukin 6 (IL6), interleukin 8 (IL8), tumour necrosis factor alpha (TNF-α), and nuclear factor kappa B (NF-κB). All primers were obtained from Eurofins Genomic, taken out of previous studies [3,14,15,16] and individually tested. Gene expression was calculated using the ΔΔCT method [17]. The characteristics of the primers are displayed in Table 2.2.5. Statistic Analyses
3. Results
3.1. Performance and Carcass Characteristics
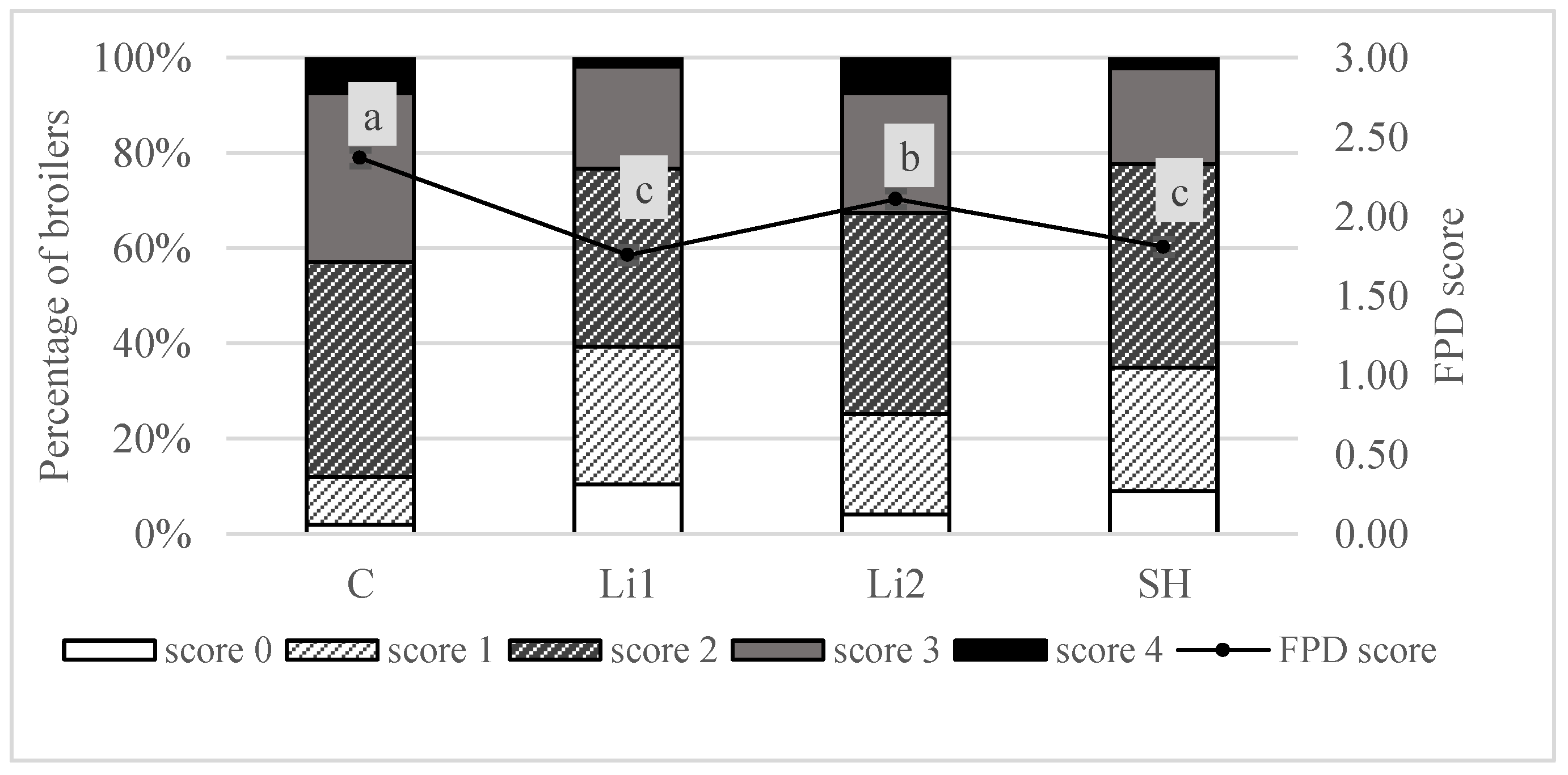
3.2. Foot Pad Health
3.3. Ileal Morphology and Caecal Inflammatory Cytokine Gene Expression
4. Discussion
4.1. Performance and Carcass Characteristics
4.2. Foot Pad Health
4.3. Ileal Morphology
4.4. Caecal Inflammatory Cytokines Genes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mateos, G.; Jiménez-Moreno, E.; Serrano, M.; Lázaro, R. Poultry response to high levels of dietary fibre sources varying in physical and chemical characteristics. J. Appl. Poult. Res. 2012, 21, 156–174. [Google Scholar] [CrossRef]
- Montagne, L.; Pluske, J.R.; Hampson, D.J. A review of interactions between dietary fibre and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Anim. Feed Sci. Technol. 2003, 108, 95–117. [Google Scholar] [CrossRef]
- Zeitz, J.O.; Neufeld, K.; Potthast, C.; Kroismayr, A.; Most, E.; Eder, K. Effects of dietary supplementation of the lignocelluloses FibreCell and OptiCell on performance, expression of inflammation-related genes and the gut microbiome of broilers. Poult. Sci. 2019, 98, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Kogut, M. The effect of microbiome modulation on the intestinal health of poultry. Anim. Feed Sci. Technol. 2019, 250, 32–40. [Google Scholar] [CrossRef]
- Tejeda, O.J.; Kim, W.K. Role of Dietary Fiber in Poultry Nutrition. Animals 2021, 11, 461. [Google Scholar] [CrossRef]
- Sarikhan, M.; Shahryar, H.A.; Gholizadeh, B. Effects of insoluble fibre on growth performance, carcass traits and ileum morphological parameters on broiler chick males. Int. J. Agric. Biol. 2010, 12, 531–536. [Google Scholar]
- Slama, J.; Schedle, K.; Wurzer, G.K.; Gierus, M. Physicochemical properties to support fibre characterization in monogastric animal nutrition. J. Sci. Food Agri. 2019, 99, 3895–3902. [Google Scholar] [CrossRef]
- Society of Nutrition Physiology. Empfehlungen zur Energie- und Nährstoffversorgung der Legehennen und Masthühner (Broiler). Ausschuss für Bedarfsnormen der Gesellschaft für Ernährungsphysiologie; DLG-Verlag: Frankfurt am Main, Germany, 1999.
- Butterworth, A.; Arnould, C.; van Niekerk, T.; Veissier, I.; Keeling, L. Welfare Quality® Assessment Protocol for Poultry (Broilers, Laying Hens); ASG Wageningen University and Research Centre: Wageningen, The Netherlands, 2009. [Google Scholar]
- Naumann, C.; Bassler, R. Die Chemische Untersuchung von Futtermitteln; VDLUFA-Verlag: Darmstadt, Germany, 2012; ISBN 9783941273146. [Google Scholar]
- Hansen, B. Determination of nitrogen as elementary N, an alternative to Kjeldahl. Acta Agric. Scand. 1989, 39, 113–118. [Google Scholar] [CrossRef]
- Röhe, I.; Ruhnke, I.; Knorr, R.; Mader, A.; Boroojeni, F.G.; Lowe, R.; Zentkek, J. Effects of grinding method, particle size, and physical form of the diet on gastrointestinal morphology and jejunal glucose transport in laying hens. Poult. Sci. 2014, 93, 2060–2068. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- De Boever, S.; Vangestel, C.; De Backer, P.; Croubels, S.; Sys, S.U. Identification and validation of housekeeping genes as internal control for gene expression in an intravenous LPS inflammation model in chickens. Vet. Immunol. Immunopathol. 2008, 122, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, L.; Li, L.; Tian, D.; Li, W.; Xu, L.; Yan, R.; Li, X.; Song, X. Protective immunity induced by Eimeria common antigen 14-3-3 against Eimeria tenella, Eimeria acervulina and Eimeria maxima. BMC Vet. Res. 2018, 14, 337. [Google Scholar] [CrossRef] [PubMed]
- Mountzouris, K.C.; Paraskeuas, V.; Fegeros, K. Priming of intestinal cytoprotective genes and antioxidant capacity by dietary phytogenic inclusion in broilers. Anim. Nutr. 2020, 6, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schedle, K. Sustainable Pig and Poultry Nutrition by Improvement of Nutrient Utilisation—A Review. Bodenkultur 2016, 67, 45–60. [Google Scholar] [CrossRef]
- Wanzenböck, E.; Zitz, U.; Steinbauer, C.; Kneifel, W.; Domig, K.; Schedle, K. A diet containing native or fermented wheat bran does not interfere with natural microbiota of laying hens. Animal 2020, 14, 1147–1155. [Google Scholar] [CrossRef]
- Jha, R.; Mishra, P. Dietary fibre in poultry nutrition and their effects on nutrient utilization, performance, gut health, and on the environment: A review. J. Anim. Sci. Biotechnol. 2021, 12, 51. [Google Scholar] [CrossRef]
- Oikeh, I.; Sakks, P.; Taylor, J.; Giannenas, I.; Blake, D.P.; Kyriazakis, I. Effects of reducing growth rate via diet dilution on bone mineralization, performance and carcass yield of coccidia-infected broilers. Poult. Sci. 2019, 25, 83–91. [Google Scholar] [CrossRef]
- Röhe, I.; Zentek, J. Lignocellulose as an insoluble fibre source in poultry nutrition: A review. J. Anim. Sci. Biotechnol. 2021, 12, 82. [Google Scholar] [CrossRef]
- Siegert, W.; Ganzer, C.; Kluth, H.; Rodehutscord, M. Effect of particle size distribution of maize and soybean meal on the precaecal amino acid digestibility in broiler chickens. Br. Poult. Sci. 2018, 59, 68–75. [Google Scholar] [CrossRef]
- González-Alvarado, J.M.; Jiménez-Moreno, E.; Lázaro, R.; Mateos, G.G. Effect of type of cereal, heat processing of the cereal, and inclusion of fibre in the diet on productive performance and digestive traits of broilers. Poult. Sci. 2007, 86, 1705–1715. [Google Scholar] [CrossRef] [PubMed]
- Makivic, L.; Glisic, M.; Boskovic, M.; Djordjevic, J.; Markovic, R.; Baltic, M.; Sefe, D. Performances, ileal and cecal microbial populations and histological characteristics in broilers fed diets supplemented with lignocellulose. Kafkas Univ. Vet. Fak. Derg. 2019, 25, 83–91. [Google Scholar] [CrossRef]
- Humer, E.; Rohrer, E.; Windisch, W.; Wetscherek, W.; Schwarz, C.; Jungbauer, L.; Schedle, K. Gender-specific effects of a phytogenic feed additive on performance, intestinal physiology and morphology in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2014, 99, 788–800. [Google Scholar] [CrossRef]
- Kheravii, S.K.; Swick, R.A.; Choct, M.; Wu, S.B. Coarse particle inclusion and lignocellulose-rich fibre addition in feed benefit performance and health of broiler chickens. Poult. Sci. 2017, 96, 3272–3281. [Google Scholar] [CrossRef] [PubMed]
- Mateos, G.; Guzman, P.; Saldana, B.; Bonilla, A.; Lazar, R.; Jimenez-Moreno, E. Relevance of dietary fibre in poultry feeding. In Proceedings of the 19th European Symposium on Poultry Nutrition, Potsdam, Germany, 26–29 August 2013; pp. 1–11. [Google Scholar]
- Amerah, A.M.; Lentle, R.G.; Ravindran, V. Influence of feed form on gizzard morphology, particle size spectra of duodenal digesta in broiler chickens. J. Poult Sci. 2007, 44, 175–181. [Google Scholar] [CrossRef]
- Morgan, N.; Bhuiyan, M.M.; Nguyen, T.N.A.; Middlebrook, T.; Hopcroft, R. Dietary soluble non-starch polysaccharide level and composition influences grower and finisher phase performance, excreta moisture content and total tract nutrient digestibility in broilers. Br. Poult. Sci. 2021, 62, 759–770. [Google Scholar] [CrossRef]
- Ducatelle, R.; Goossens, E.; de Meyer, F.; Eeckhaut, V.; Antonissen, G.; Haesebrouck, F.; van Immerseel, F. Biomarkers for monitoring intestinal health in poultry: Present status and future perspectives. Vet. Res. 2018, 49, 43. [Google Scholar] [CrossRef]
- Rezaei, M.; Karimi Torshizi, M.A.; Rouzbehan, Y. The influence of different levels of micronized insoluble fibre on broiler performance and litter moisture. Poult. Sci. 2011, 90, 12. [Google Scholar] [CrossRef]
- Shepherd, E.M.; Fairchild, B.D. Footpad dermatitis in poultry. Poult. Sci. 2010, 89, 2043–2051. [Google Scholar] [CrossRef]
- Tejeda, O.J.; Kim, W.K. The effects of cellulose and soybean hulls as sources of dietary fibre on the growth performance, organ growth, gut histomorphology, and nutrient digestibility of broiler chickens. Poult. Sci. 2020, 99, 6828–6836. [Google Scholar] [CrossRef]
- Bogusławska-Tryk, M.; Bogucka, J.; Dankowiakowska, A.; Walasik, K. Small intestine morphology and ileal biogenic amines content in broiler chickens fed diets supplemented with lignocellulose. Livest. Sci. 2020, 241, 104189. [Google Scholar] [CrossRef]
- Verdal, H.; Mignon-Grasteau, S.; Jeulin, C.; le Bihan-Duval, E.; Leconte, M.; Mallet, S.; Martin, C.; Narcy, A. Digestive tract measurements and histological adaptation in broiler lines divergently selected for digestive efficiency. Poult. Sci. 2010, 89, 1955–1961. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Starter | Grower | Finisher | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ingredients % FM | C | Li1 | Li2 | SH | C | Li1 | Li2 | SH | C | Li1 | Li2 | SH |
| Corn | 50.0 | 49.6 | 49.6 | 49.0 | 50.0 | 49.6 | 49.6 | 48.9 | 52.5 | 52.1 | 52.1 | 51.4 |
| Soybean meal, hp | 28.6 | 28.4 | 28.4 | 28.0 | 27.8 | 27.6 | 27.6 | 27.3 | 23.6 | 23.4 | 23.4 | 23.1 |
| Wheat | 12.2 | 12.1 | 12.1 | 11.9 | 14.4 | 14.3 | 14.3 | 14.1 | 16.7 | 16.5 | 16.5 | 16.3 |
| Corn gluten | 2.5 | 2.5 | 2.5 | 2.5 | ||||||||
| Plant-based oil | 2.2 | 2.2 | 2.2 | 2.6 | 4.0 | 4.0 | 4.0 | 4.4 | 3.9 | 3.9 | 3.9 | 4.3 |
| Premix 1 | 1.2 | 1.2 | 1.2 | 1.2 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Monocalcium phosphate | 1.1 | 1.1 | 1.1 | 1.0 | 0.8 | 0.8 | 0.8 | 0.8 | 0.6 | 0.6 | 0.6 | 0.6 |
| Calcium carbonate | 0.9 | 0.9 | 0.9 | 0.9 | 0.8 | 0.8 | 0.8 | 0.8 | 0.9 | 0.9 | 0.9 | 0.8 |
| DL-methionine | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.3 | 0.2 |
| Zootechnical additives 2 | 0.3 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| L-Lysin | 0.2 | 0.2 | 0.2 | 0.2 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| Sodium chloride | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Coccidiostats 3 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | ||||
| L-Threonine | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| Lignocellulose product I | 0.8 | 0.8 | 0.8 | |||||||||
| Lignocellulose product II | 0.8 | 0.8 | 0.8 | |||||||||
| Soybean hulls | 1.6 | 1.6 | 1.6 | |||||||||
| Chemical composition %FM | ||||||||||||
| Dry matter | 90.1 | 90.1 | 90.4 | 89.9 | 89.0 | 89.6 | 88.6 | 88.9 | 88.7 | 89.0 | 88.4 | 88.3 |
| Crude protein | 21.2 | 21.4 | 22.0 | 21.1 | 19.4 | 19.0 | 19.4 | 18.8 | 18.9 | 18.4 | 19.2 | 18.1 |
| Ether extract | 4.6 | 4.5 | 4.5 | 4.5 | 6.3 | 6.3 | 6.1 | 6.7 | 6.3 | 6.3 | 6.0 | 6.6 |
| Crude fibre | 2.6 | 2.4 | 2.7 | 2.6 | 1.8 | 2.2 | 2.0 | 2.2 | 2.3 | 2.1 | 2.4 | 2.2 |
| IDF | 13.5 | 13.9 | 13.8 | 14 | 13.3 | 13.5 | 13.5 | 14.1 | 11.6 | 11.7 | 12.1 | 11.8 |
| SDF | >1 | >1 | >1 | >1 | >1 | >1 | >1 | >1 | >1 | >1 | >1 | >1 |
| Crude ash | 5.5 | 5.3 | 5.4 | 5.3 | 5.2 | 4.7 | 4.6 | 4.8 | 4.7 | 4.4 | 4.5 | 4.2 |
| Starch | 42.8 | 43.4 | 42.9 | 43.2 | 43.3 | 43.9 | 43.0 | 42.7 | 44.1 | 45.6 | 44.0 | 45.1 |
| Sugar | 4.5 | 4.3 | 4.6 | 4.6 | 4.4 | 4.4 | 4.5 | 4.2 | 4.1 | 4.0 | 4.0 | 4.1 |
| Gross energy MJ/kg | 17.2 | 17.1 | 17.3 | 17.2 | 17.2 | 17.5 | 17.2 | 17.6 | 17.4 | 17.5 | 17.3 | 17.4 |
| Calculated composition | ||||||||||||
| AMEN MJ/kg 4 | 12.6 | 12.7 | 12.7 | 12.6 | 13.0 | 13.0 | 12.9 | 12.9 | 13.0 | 13.1 | 12.9 | 13.1 |
| Wet sieve analysis | ||||||||||||
| >1 mm, % | 9.9 | 6.2 | 9.2 | 7.5 | 19.5 | 17.6 | 13.7 | 12.7 | 9.8 | 11.4 | 9.8 | 8.4 |
| ≥0.5–≤ 1 mm, % | 30.9 | 30.6 | 33.3 | 34.8 | 34.1 | 31.4 | 31.5 | 32.9 | 35.7 | 37.1 | 35.4 | 36.2 |
| <0.5 mm, % | 59.2 | 63.2 | 57.5 | 57.8 | 46.4 | 51.1 | 54.8 | 54.5 | 54.6 | 51.2 | 54.8 | 55.4 |
| dMean, mm | 0.6 | 0.5 | 0.6 | 0.5 | 0.8 | 0.8 | 0.7 | 0.7 | 0.6 | 0.7 | 0.6 | 0.6 |
| Gene | Sequence 5′ to 3′ | Product Length, bp | Annealing Temperature, °C | NCBI Accession | |
|---|---|---|---|---|---|
| GAPDH | for. | GGTGGTGCTAAGCGTGTTAT | 264 | 57.3 | K01458 |
| rev. | ACCTCTGTCATCTCTCCACA | 57.3 | |||
| ACTB | for. | ATGAAGCCCAGAGCAAAAGA | 223 | 55.3 | NM_205518 |
| rev. | GGGGTGTTGAAGGTCTCAAA | 57.3 | |||
| Ubiquitin | for. | GGGATGCAGATCTTCGTGAAA | 147 | 57.9 | M11100 |
| rev. | CTT GCC AGC AAA GAT CAA CCT T | 58.4 | |||
| IL 1β | for. | CATTACCGTCCCGTTGCTTT | 105 | 57.3 | NM 204524.1 |
| rev. | AGTCACAATAAATACCTCCACCC | 58.9 | |||
| IL6 | for. | CCAGAAATCCCTCCTCGCCAATC | 222 | 64.2 | NM 204628.1 |
| rev. | TGAAACGGAACAACACTGCCATC | 60.6 | |||
| IL8 | for. | TGCTGTGGGATTCACTGTCCA | 93 | 59.8 | HM179639.1 |
| rev. | ACTGAAGTGGCTTCCAAGGGA | 59.8 | |||
| TNF-α | for. | CAGGACAGCCTATGCCAACA | 95 | 59.4 | NM 204628.1 |
| rev. | CATCTGAACTGGGCGGTCAT | 59.4 | |||
| NF-kB | for. | GAAGGAATCGTACCGGGAACA | 131 | 59.8 | NM_205134.1 |
| rev. | CTCAGAGGGCCTTGTGACAGTAA | 62.4 | |||
| C | Li1 | Li2 | SH | SEM | p-Value | |
|---|---|---|---|---|---|---|
| Animals day 1, n | 1260 | 1260 | 1260 | 1260 | ||
| Animals day 10, n | 1260 | 1260 | 1260 | 1260 | ||
| Animals day 28, n | 1229 | 1229 | 1230 | 1231 | ||
| Animals day 36, n | 1223 | 1225 | 1229 | 1226 | ||
| Ratio male/female at day 36, n | 610/613 | 616/609 | 618/611 | 625/601 | ||
| Body weight (BW), g | ||||||
| Day 1 1 | 39 | 39 | 39 | 39 | 0.1 | 0.994 |
| Day 10 1 | 240 | 243 | 242 | 244 | 1.3 | 0.791 |
| Day 28 2 | 1436 b | 1517 a | 1490 a | 1498 a | 9.0 | 0.005 |
| Day 36 3 | 2028 | 2083 | 2095 | 2109 | 13.7 | 0.178 |
| Average daily weight gain 1 (ADG), g | ||||||
| Starter | 20 | 20 | 20 | 21 | 0.1 | 0.785 |
| Grower | 66 b | 70 a | 69 ab | 69 a | 0.5 | 0.006 |
| Finisher | 85 | 81 | 87 | 87 | 1.6 | 0.541 |
| All | 57 | 58 | 59 | 59 | 0.4 | 0.176 |
| Daily feed intake 1 (dFI), g | ||||||
| Starter | 27 | 27 | 27 | 27 | 0.2 | 0.906 |
| Grower | 97 | 99 | 98 | 98 | 0.5 | 0.777 |
| Finisher | 170 | 170 | 171 | 172 | 2.0 | 0.990 |
| All | 92 | 93 | 93 | 93 | 0.6 | 0.955 |
| Feed conversion rate 1 (FCR), kg/kg | ||||||
| Starter | 1.35 | 1.35 | 1.35 | 1.32 | 0.01 | 0.686 |
| Grower | 1.48 | 1.41 | 1.43 | 1.43 | 0.01 | 0.101 |
| Finisher | 2.02 | 2.12 | 2.00 | 1.99 | 0.03 | 0.533 |
| All | 1.61 | 1.57 | 1.57 | 1.56 | 0.01 | 0.368 |
| C | Li1 | Li2 | SH | SEM | p-Value | ||
|---|---|---|---|---|---|---|---|
| Diet | Sex | ||||||
| Body weight (BW) 1, g | 2171 | 2249 | 2220 | 2222 | 13 | 0.177 | <0.001 |
| Eviscerated carcass 1, g | 1753 | 1815 | 1792 | 1795 | 10 | 0.173 | <0.001 |
| % Eviscerated carcass 1 | |||||||
| Abdominal fat | 0.91 b | 1.05 a | 0.94 ab | 1.00 ab | 0.02 | 0.004 | <0.001 |
| Heart | 0.64 | 0.62 | 0.59 | 0.61 | 0.01 | 0.133 | 0.569 |
| Liver | 2.40 | 2.40 | 2.39 | 2.41 | 0.02 | 0.962 | 0.683 |
| Head and neck | 4.80 | 4.75 | 4.83 | 4.85 | 0.03 | 0.644 | 0.638 |
| Wings | 8.98 | 8.93 | 8.98 | 8.93 | 0.03 | 0.881 | 0.001 |
| Legs | 4.34 a | 4.03 b | 4.13 b | 4.18 ab | 0.03 | 0.001 | <0.001 |
| Breast | 29.54 | 29.39 | 29.71 | 29.31 | 0.09 | 0.408 | 0.047 |
| Thigh | 26.00 | 25.79 | 26.18 | 26.15 | 0.07 | 0.204 | 0.522 |
| Gizzard 2, g/kg BW | 9.73 | 10.09 | 9.44 | 9.69 | 0.16 | 0.562 | <0.001 |
| pH gizzard 2 | 2.95 | 3.02 | 2.98 | 3.10 | 0.05 | 0.769 | 0.485 |
| C | Li1 | Li2 | SH | SEM | p-Value | ||
|---|---|---|---|---|---|---|---|
| Villus | |||||||
| Height | µm | 671.07 b | 742.35 a | 724.00 a | 753.38 a | 6.75 | 0.001 |
| Villus area calculated 2 | mm² | 0.32 b | 0.38 a | 0.34 ab | 0.35 ab | 0.01 | 0.053 |
| Goblet cells | n/100 µm villus height | 19.75 | 20.98 | 19.64 | 17.69 | 0.47 | 0.149 |
| Crypt | |||||||
| Depth | µm | 160.00 b | 189.22 a | 177.99 a | 192.93 a | 2.56 | 0.001 |
| Goblet cells | n/100 µm crypt depth | 20.83 | 22.98 | 22.83 | 20.64 | 0.54 | 0.290 |
| VH/CD | 4.41 | 4.24 | 4.48 | 4.26 | 0.07 | 0.552 | |
| Muscularis | µm | 170.88 b | 168.00 b | 185.79 a | 179.08 ab | 1.93 | 0.002 |
| C | Li1 | Li2 | SH | SEM | p-Value | |
|---|---|---|---|---|---|---|
| IL 1β | 1.00 | 0.76 | 1.49 | 1.93 | 0.24 | 0.337 |
| IL6 | 1.00 | 1.38 | 0.99 | 0.99 | 0.14 | 0.716 |
| IL8 | 1.00 | 0.74 | 1.56 | 1.11 | 0.27 | 0.619 |
| TNF-α | 1.00 | 0.77 | 1.84 | 1.63 | 0.25 | 0.392 |
| NF-kB | 1.00 | 0.88 | 0.74 | 0.86 | 0.12 | 0.907 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liebl, M.; Gierus, M.; Potthast, C.; Schedle, K. Influence of Insoluble Dietary Fibre on Expression of Pro-Inflammatory Marker Genes in Caecum, Ileal Morphology, Performance, and Foot Pad Dermatitis in Broiler. Animals 2022, 12, 2069. https://doi.org/10.3390/ani12162069
Liebl M, Gierus M, Potthast C, Schedle K. Influence of Insoluble Dietary Fibre on Expression of Pro-Inflammatory Marker Genes in Caecum, Ileal Morphology, Performance, and Foot Pad Dermatitis in Broiler. Animals. 2022; 12(16):2069. https://doi.org/10.3390/ani12162069
Chicago/Turabian StyleLiebl, Mariella, Martin Gierus, Christine Potthast, and Karl Schedle. 2022. "Influence of Insoluble Dietary Fibre on Expression of Pro-Inflammatory Marker Genes in Caecum, Ileal Morphology, Performance, and Foot Pad Dermatitis in Broiler" Animals 12, no. 16: 2069. https://doi.org/10.3390/ani12162069