1. Introduction
The global cattle industry, through both of its derivatives (dairy and beef), provides 81% of the world’s milk and 22% of the world’s meat needs [
1]. Cow milk production has more than doubled in the past 50 years, now at more than 800 million tonnes per year, and is predicted to increase with the growing population [
1]. Across commercial livestock breeding programs, breeders in both the dairy and beef industry focus their objectives on increasing output and efficiency, which, in addition to maximising profits, can also reduce waste and environmental impact per animal produced [
2]. Optimal reproductive strategies are essential to this process and, as such, sub-fertile animals can have a significant negative impact on productivity. This is especially true of males where the use of artificial insemination (AI) for reproduction in cattle has become widely adopted, particularly in the dairy segments [
3]. Because bulls can produce large quantities of semen (around 800–1200 doses per week), the industry relies on a small pool of high value animals that have been selected for beneficial genetic traits, but not necessarily for fertility [
3,
4]. This is becoming increasingly relevant with regard to not only dairy production, as the combination of AI with sexed semen is becoming more widely used, but also for beef, where the benefits of using AI over natural service are being realised [
5]. On average, a breeding bull can inseminate from around 130,000 to 200,000 cows in its lifetime and, in extreme cases, this can rise as high as 400,000 [
6].
If a bull is sub-fertile, this can impact the conception and birth rates; it can also pass on any genetic abnormalities to his offspring, affecting their fertility and thereby perpetuating further opportunities for cost and economic losses for the producer. It is generally accepted that having a proactive screening programme for genetically determined fertility issues in bulls can help prevent further losses. Nonetheless, the extent to which genetic abnormalities causing infertility are a financial burden to the modern cattle breeding industry is hitherto not calculated. Consequently, the likely benefits of a screening programme have not been fully appreciated, nor has the additional benefit of a system that identifies additional abnormalities not detected by older methods.
Balanced chromosomal rearrangements are the leading genetic cause of fertility issues in livestock, recently reassessed in the pig breeding industry as being present in around 1% of breeding boars when enhanced screening approaches are used [
7,
8]. We recently calculated the financial impact on the industry of a translocation-carrying boar as between GBP 69,802 and GBP 51,215,378 per animal, depending on its position in the breeding pyramid [
8]. In pigs, most chromosomes in the karyotype are found translocated; however, among bulls, the distribution is non-random. The karyotype of domestic cattle (both
Bos indicus and Bos taurus) comprises 29 pairs of acrocentric autosomes (the submetacentric X and the tiny Y completing the set), making Robertsonian translocations (centric fusions) very likely [
9]. Indeed, arguably the best known non-human chromosome abnormality in all of the scientific literature is a Robertsonian translocation (RobT) of the largest and smallest cattle autosomes (1:29). This specific abnormality has been observed in over 50 breeds around the world, with reported frequencies varying from up to 60% to as low as 0.4% in Holsteins [
10]. As with any balanced translocation, the phenotypic effects are rarely seen in the animal itself, but manifested in its subsequent reproductive performance, largely because of perturbations of the normal meiotic process, leading to a reduction in the number of gametes produced, and chromosomal unbalanced products that can lead to implantation failure and/or pregnancy loss [
11].
In addition to the 1:29 RobT, over 44 other similar errors (also RobTs) have been observed and described in cattle, with equivalent negative effects on fertility [
9]. These additional RobTs have a lower reported incidence rate: in general, around 6.5% in Holstein cattle; however, this number varies with other breeds [
10]. It is possible, however, given the difficulty in identifying individual chromosomes, that some RobTs are identified as 1:29, when in fact they involve other chromosomes. RobTs, in general terms, are nonetheless relatively easy to detect with standard Giemsa staining (with or without banded karyotyping), as the two fused chromosomes will appear as one larger, (sub)metacentric chromosome [
9]. Reciprocal translocations (RECTs) are, however, far more difficult to identify as not only are all the autosomes acrocentric but they are all of similar size. As such, while, with relatively sophisticated extended chromosome preparation and G-banding techniques, RECTs can be identified, it is estimated that only 16% are spotted, leaving 84% undiagnosed [
11]. In point of fact, most labs only stain chromosomes homogeneously (i.e., without banding) in order to spot 1:29 and other RobTs, and do not attempt to identify RECTs.
Finding a precise figure as to the incidence of RECTs in cattle is not an easy task, mostly because most labs do not look for them. Moreover, finding a relative incidence compared to RobTs is complicated by the fact that the incidence of RobTs is so variable, in turn depending on the extent to which screening programmes have near eliminated them. De Lorenzi et al. [
12] suggested that the rate of RECTs is around four–five times the rate of non-1:29 RobTs; however, if we accept that at best, only 16% were identified [
11], it seems clear that RECTs are an under-discovered genetic hazard, at least as significant as the 1:29 RobT, and possibly much more so. One of the purposes of this study was to estimate the relative rates of RECTs vs. RobTs.
Cattle breeding companies are often required to demonstrate that their breeding bulls are free of the 1:29 RobT [
13]. The rationale for this is clear in that the risks of letting an undiagnosed translocation into the breeding herd would have significant costs in terms of finance, loss of reputation and the environment. Pregnancy rates per insemination in UK herds are in the region of 50–60% and it takes at least 55 days to establish whether a cow is pregnant [
1], which leads to a loss of productive days lactating if she is not. The effect of RobT 1:29 are therefore profound. By extension, therefore, the same would apply to other RobTs and RECTs. Tests to detect RobT 1:29 are often (by PCR or FISH) targeted at this translocation alone, rather than using standard chromosome preparations [
13], which would at least detect other RobTs. Thus, if employing standard chromosome preparation techniques, a reasonably competent laboratory would detect a RobT that involved chromosomes other than 1 and 29; however, it would be unlikely to detect an RECT at all.
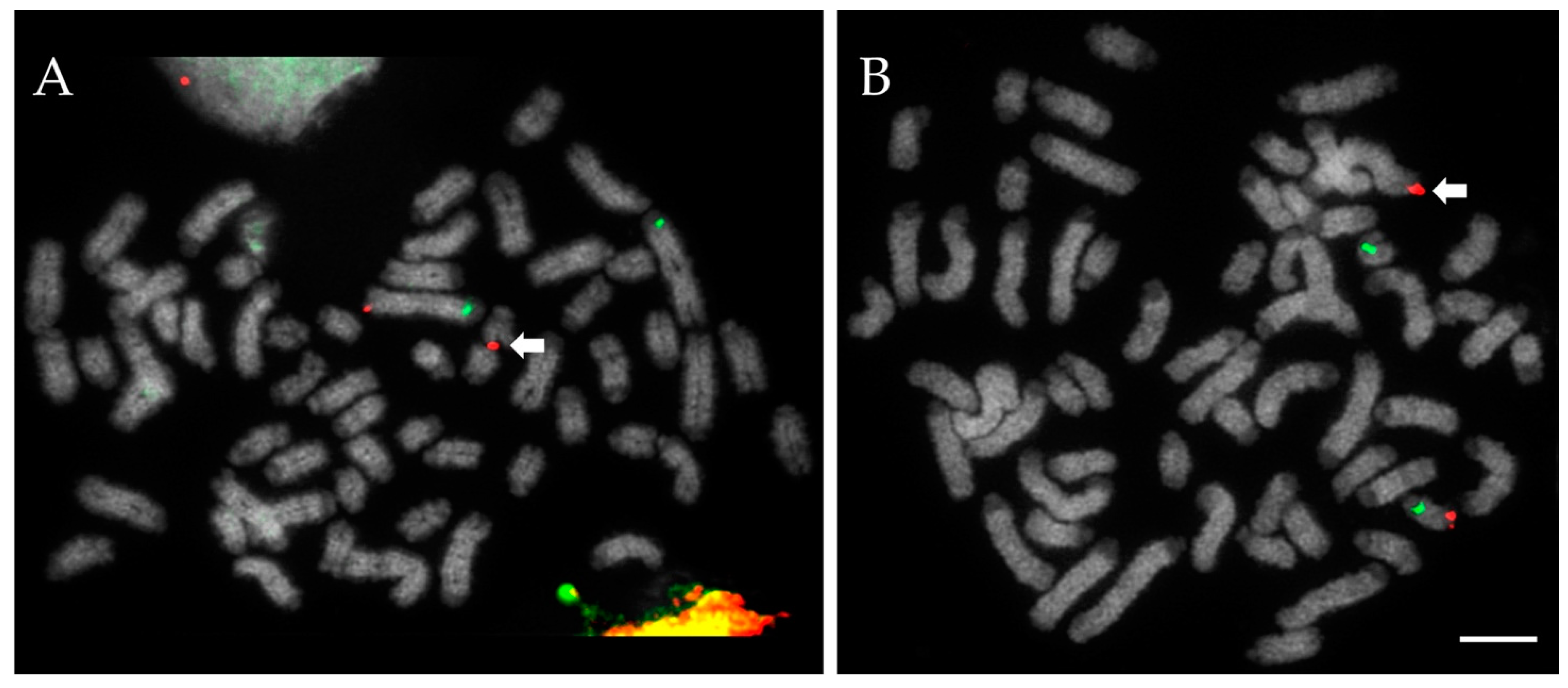
In recent years, we developed a screening protocol that can readily detect both RobTs and RECTs (collectively henceforth referred to as “RTs”) for all cattle chromosomes [
3]. The method utilises fluorescence in situ hybridisation (FISH), sub-telomeric probes and a two-slide multi-hybridization device, with probes for distal (furthest from centromere) and proximal (nearest) regions, one per chromosome, per square, highlighting the chromosome of interest. RECTs are detected by probes appearing on different chromosomes and RobT fusions are also easily spotted [
3]. This, in turn, is based on our current pig-based screening approach [
7] which has, to date, been used to screen over 2500 boars in our laboratory. While we have calculated the potential financial impact of a pig chromosome translocation entering the breeding herd [
8], we have yet to do the same for the (dairy) cattle breeding industry. The purpose of this study is to achieve this. Moreover, we provide an update of our efforts with the cattle device, improving on the prototype previously reported in [
3] and describing new, hitherto unreported chromosome abnormalities.
4. Discussion
So called “clinical” cytogenetics of domestic species have a long and distinguished history [
12]. In pigs, activity was it its highest in the late the 20th century, with numerous cytogenetic screening programmes (notably the National Veterinary School of Toulouse in France) routinely screening samples. An unfortunate decline in the number and activity of screening laboratories, however, has not been accompanied by a decline in the problem. We recently suggested that around 1% of AI breeding boars currently carry an RT, twice that reported in the literature [
3]. In sheep, goats, camels, chicken, duck and turkey, screening programmes largely do not exist, presumably due to the difficulty in interpreting the karyotype. While the cattle karyotype is similarly intractable (large number of similar looking chromosomes), the much higher value of breeding bulls (especially for dairy) compared to other species means that attempts to screen, at least for RobTs, remains paramount. In Sweden, the fertility of the whole cattle breeding population has demonstrably been increased through systematic screening and eradication of Rob1:29 and, in the UK and Australia, cytogenetic evaluation is a requirement for imported cattle [
13]. RTs are heritable and transmitted to around half of the surviving offspring of the carrier. Therefore, the continuation of screening programmes is essential, even if the problem is near eradicated, as de novo abnormalities will inevitably occur, accumulate and ultimately be inherited.
Despite the clear association between hypo-prolificacy and all RTs, RECTs are not screened for in cattle, as a readily workable solution to screen for them has not hitherto been developed. In this paper, we provide a solution to redress this anomaly. As mentioned in the results section, RECTs, theoretically, carry a greater chance of reduction in productivity than RobTs, as the quadrivalent formed at meiosis impedes the process more, and leads to a greater proportion of chromosomally unbalanced products than the trivalent formed via a RobT [
11]. For this reason, our calculations of financial loss were based on established figures of reduction in productivity of 5% established through the study of Rob1:29 (the most conservative end of the 5–10% figure quoted), and 10% for RECTs, although the effect is most probably greater still. Indeed, RECTs typically lead to a 50% reduction in litter size in pigs [
7,
8].
Even in animals such as humans and pigs where the karyotype is relatively easy to analyse, many translocations (regardless of whether they involve exchange of large blocks of chromatin) are difficult to identify by classical cytogenetics [
7]. Moreover, some (cryptic) translocations cannot be detected by banding, regardless of preparation quality and tractability of karyotype [
7]. The screening approach described herein, however, detects all RobTs and RECTs and analysis is both simple and straightforward to implement. Under the stated assumptions, our figures are consistently conservative and they do not take into account any potential loss in reputation or customer base that a breeder might face as a result, nor the costs involved with any attempt to replace the “missing” animals (if at all possible, due to the likely higher genetic merit of the new generation). It is also interesting to note that our calculations highlight how the more efficient a breeding business becomes (i.e., improved birth rates, less miscarriage, lower mortality) the more severe the economic impact of an RT will be. Across the global cattle breeding industry, breeding practices, average herd performance and monetary values inevitably display a large variability. As such, calculations such as the ones presented here can only give an idea of the order of magnitude of a particular problem and a tailored scenario calculation that will accurately reflect the economic impact of RTs for any specific breeder will require imputation of the producer’s specific needs into our algorithm. In our previous porcine study [
8] we calculated likely losses of five scenarios in the pig breeding industry that, broadly speaking, encompassed a wide range of possible real-life cases that could be adapted to model any actual occurrence. Here, for cattle, we provide the framework through which the financial benefits of RT screening can be calculated, taking into account individual circumstances. We encourage companies to do this for their own particular purposes and all calculation spreadsheets are freely available by contacting the authors.
In pigs, the reported incidence of RTs is 0.47% [
16]; however, our efforts with a multi-probe screening device suggests that the true incidence is probably around twice that number (around 1%) [
8]. In bulls, the reported incidence of a single (Rob1:29) translocation is between 0.4% in Holstein cattle (in herds where there have been active efforts to eliminate it) to 60% (where they have not) [
9,
10]. In the results section, we make an attempt to perform the near impossible task of calculating the relative contribution of RECTs. A figure of twice the frequency of RECTs vs. RobTs (so our screening approach detects 3x as many abnormalities overall) does not seem unreasonable. In reality, it probably is much higher (and of course infinitely if all the RobTs have been eradicated). Therefore, the added ability to detect all RECTs, including the cryptic ones, suggests that a switch from the classical Giemsa staining method (that detects RobTs only) or PCR-based “Rob1:29 only” to widespread adoption of our multi-FISH approach should become a priority.
Finally, while this paper has concentrated on the financial impact of genetic abnormalities, the environmental consequences deserve consideration. A saving in wasted animals and resources also has a corresponding saving in methane or other greenhouse gas emissions. As we develop more sophisticated financial models to map the impact of RTs, future studies will concentrate on their environmental impact also likely incorporating methodologies such as lifecycle analysis.


 ,
, 
{kind=link}
