
A KIT Variant Associated with Increased White Spotting Epistatic to MC1R Genotype in Horses (Equus caballus)

 ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Horses and Putative Candidate Variant Inspection
2.2. Linkage Disequilibrium Analysis with Dominant White 19 and MC1R
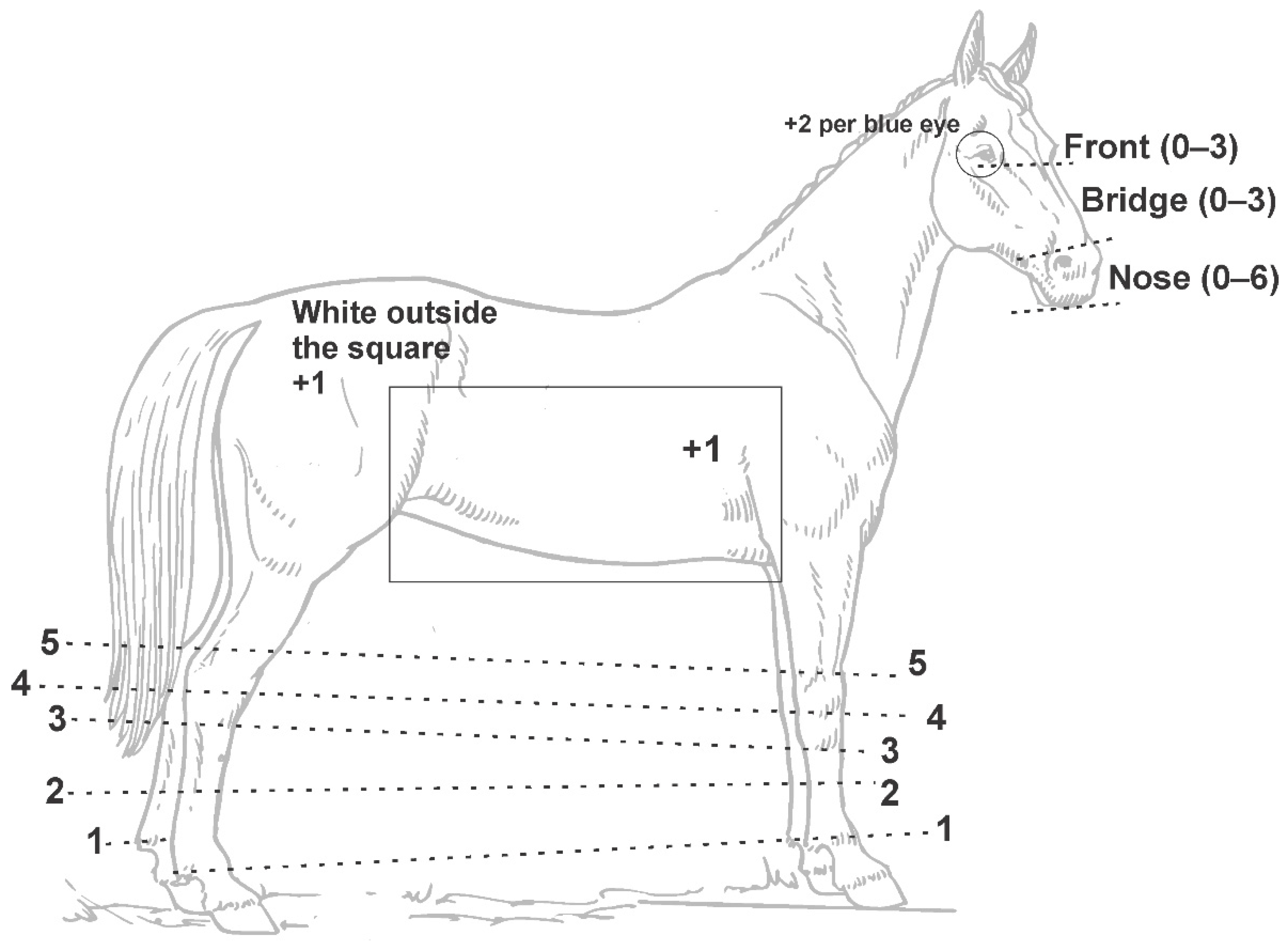
2.3. Phenotyping and Statistical Analysis
3. Results
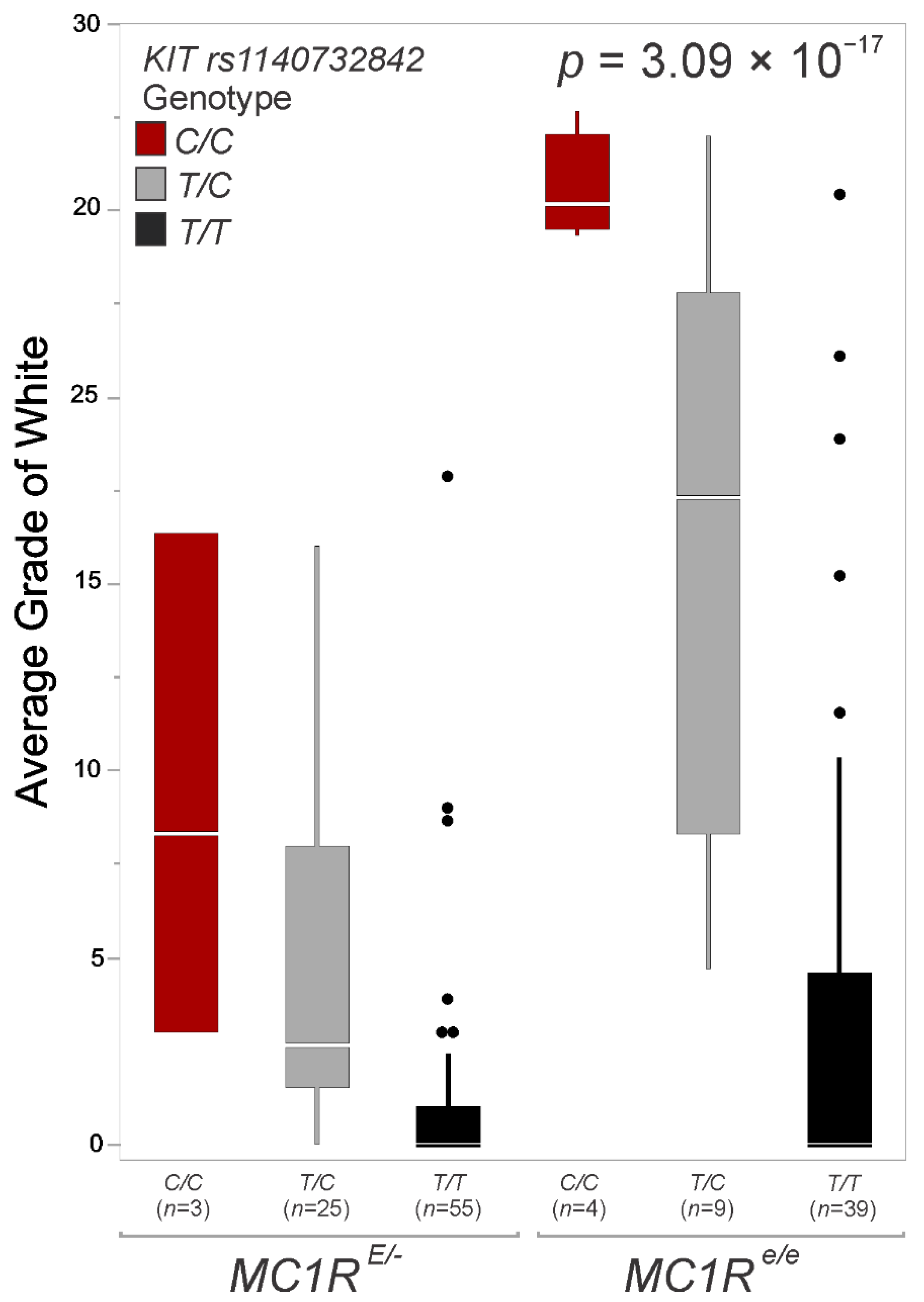
3.1. Variant Analysis Suggests a Candidate in KIT Influenced by MC1R
3.2. Linkage Disequilibrium between KIT and MC1R
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haase, B.; Brooks, S.A.; Schlumbaum, A.; Azor, P.J.; Bailey, E.; Alaeddine, F.; Mevissen, M.; Burger, D.; Poncet, P.-A.; Rieder, S. Allelic heterogeneity at the equine KIT locus in dominant white (W) horses. PLoS Genet. 2007, 3, e195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haase, B.; Brooks, S.A.; Tozaki, T.; Burger, D.; Poncet, P.A.; Rieder, S.; Hasegawa, T.; Penedo, C.; Leeb, T. Seven novel KIT mutations in horses with white coat colour phenotypes. Anim. Genet. 2009, 40, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Haase, B.; Rieder, S.; Tozaki, T.; Hasegawa, T.; Penedo, M.C.T.; Jude, R.; Leeb, T. Five novel KIT mutations in horses with white coat colour phenotypes. Anim. Genet. 2011, 42, 337–339. [Google Scholar] [CrossRef]
- Hauswirth, R.; Haase, B.; Blatter, M.; Brooks, S.A.; Burger, D.; Drögemüller, C.; Gerber, V.; Henke, D.; Janda, J.; Jude, R. Mutations in MITF and PAX3 cause “splashed white” and other white spotting phenotypes in horses. PLoS Genet. 2012, 8, e1002653. [Google Scholar] [CrossRef] [Green Version]
- Hauswirth, R.; Jude, R.; Haase, B.; Bellone, R.R.; Archer, S.; Holl, H.; Brooks, S.A.; Tozaki, T.; Penedo, M.C.T.; Rieder, S. Novel variants in the KIT and PAX 3 genes in horses with white-spotted coat colour phenotypes. Anim. Genet. 2013, 44, 763–765. [Google Scholar] [CrossRef]
- Haase, B.; Jagannathan, V.; Rieder, S.; Leeb, T. A novel KIT variant in an Icelandic horse with white-spotted coat colour. Anim. Genet. 2015, 46, 466. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.A.; Bailey, E. Exon skipping in the KIT gene causes a Sabino spotting pattern in horses. Mamm. Genome 2005, 16, 893–902. [Google Scholar] [CrossRef]
- Brooks, S.A.; Lear, T.L.; Adelson, D.L.; Bailey, E. A chromosome inversion near the KIT gene and the Tobiano spotting pattern in horses. Cytogenet. Genome Res. 2008, 119, 225–230. [Google Scholar] [CrossRef]
- Holl, H.; Brooks, S.; Bailey, E. De novo mutation of KIT discovered as a result of a non-hereditary white coat colour pattern. Anim. Genet. 2010, 41, 196–198. [Google Scholar] [CrossRef]
- Dürig, N.; Jude, R.; Holl, H.; Brooks, S.A.; Lafayette, C.; Jagannathan, V.; Leeb, T. Whole genome sequencing reveals a novel deletion variant in the KIT gene in horses with white spotted coat colour phenotypes. Anim. Genet. 2017, 48, 483–485. [Google Scholar] [CrossRef]
- Holl, H.M.; Brooks, S.A.; Carpenter, M.L.; Bustamante, C.D.; Lafayette, C. A novel splice mutation within equine KIT and the W15 allele in the homozygous state lead to all white coat color phenotypes. Anim. Genet. 2017, 48, 497–498. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.; Vierra, M.; Foster, G.; Brooks, S.A.; Lafayette, C. De novo mutation of KIT causes extensive coat white patterning in a family of Berber horses. Anim. Genet. 2020, 52, 135–137. [Google Scholar] [CrossRef] [PubMed]
- Patterson Rosa, L.; Martin, K.; Vierra, M.; Foster, G.; Lundquist, E.; Brooks, S.A.; Lafayette, C. Two variants of KIT causing white patterning in Stock-type horses. J. Hered. 2021, 112, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Metallinos, D.L.; Bowling, A.T.; Rine, J. A missense mutation in the endothelin-B receptor gene is associated with Lethal White Foal Syndrome: An equine version of Hirschsprung disease. Mamm. Genome 1998, 9, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Esdaile, E.; Till, B.; Kallenberg, A.; Fremeux, M.; Bickel, L.; Bellone, R.R. A de novo missense mutation in KIT is responsible for dominant white spotting phenotype in a Standardbred horse. Anim. Genet. 2022, 53, 534–537. [Google Scholar] [CrossRef] [PubMed]
- Patterson Rosa, L.; Martin, K.; Vierra, M.; Foster, G.; Brooks, S.A.; Lafayette, C. Non-frameshift deletion on MITF is associated with a novel splashed white spotting pattern in horses (Equus caballus). Anim. Genet. 2022, 53, 538–540. [Google Scholar] [CrossRef]
- Stachurska, A.; Jansen, P. Crypto-tobiano horses in Hucul breed. Czech J. Anim. Sci. 2015, 60, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Haase, B.; Signer-Hasler, H.; Binns, M.M.; Obexer-Ruff, G.; Hauswirth, R.; Bellone, R.R.; Burger, D.; Rieder, S.; Wade, C.M.; Leeb, T. Accumulating mutations in series of haplotypes at the KIT and MITF loci are major determinants of white markings in Franches-Montagnes horses. PLoS ONE 2013, 8, e75071. [Google Scholar] [CrossRef] [Green Version]
- Rieder, S.; Hagger, C.; Obexer-Ruff, G.; Leeb, T.; Poncet, P.-A. Genetic Analysis of White Facial and Leg Markings in the Swiss Franches-Montagnes Horse Breed. J. Hered. 2008, 99, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Rieder, S.; Taourit, S.; Mariat, D.; Langlois, B.; Guérin, G. Mutations in the agouti (ASIP), the extension (MC1R), and the brown (TYRP1) loci and their association to coat color phenotypes in horses (Equus caballus). Mamm. Genome 2001, 12, 450–455. [Google Scholar] [CrossRef]
- Kalbfleisch, T.S.; Rice, E.S.; DePriest, M.S.; Walenz, B.P.; Hestand, M.S.; Vermeesch, J.R.; O’Connell, B.L.; Fiddes, I.T.; Vershinina, A.O.; Petersen, J.L.; et al. EquCab3, an Updated Reference Genome for the Domestic Horse. bioRxiv 2018, 306928. [Google Scholar] [CrossRef]
- Beeson, S.K.; Mickelson, J.R.; McCue, M.E. Exploration of fine-scale recombination rate variation in the domestic horse. Genome Res. 2019, 29, 1744–1752. [Google Scholar] [CrossRef] [PubMed]
- Beeson, S.K.; Mickelson, J.R.; McCue, M.E. Equine recombination map updated to EquCab3.0. Anim. Genet. 2020, 51, 341–342. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.A.; Palermo, K.M.; Kahn, A.; Hein, J. Impact of white-spotting alleles, including W20, on phenotype in the American Paint Horse. Anim. Genet. 2020, 51, 707–715. [Google Scholar] [CrossRef]
- Choi, Y.; Sims, G.E.; Murphy, S.; Miller, J.R.; Chan, A.P. Predicting the functional effect of amino acid substitutions and indels. PLoS ONE 2012, 7, e46688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, P.C.; Henikoff, S. SIFT: Predicting amino acid changes that affect protein function. Nucleic Acids Res. 2003, 31, 3812–3814. [Google Scholar] [CrossRef] [Green Version]
- Diagnostics, E. Etalon DNA Test Panel Comparison Chart. Available online: https://www.etalondx.com/compare-tests (accessed on 31 January 2022).
- Howe, K.L.; Achuthan, P.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J.; et al. Ensembl 2021. Nucleic Acids Res. 2021, 49, D884–D891. [Google Scholar] [CrossRef]
- Choi, Y.; Chan, A.P. PROVEAN web server: A tool to predict the functional effect of amino acid substitutions and indels. Bioinformatics 2015, 31, 2745–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires, D.E.V.; Ascher, D.B.; Blundell, T.L. DUET: A server for predicting effects of mutations on protein stability using an integrated computational approach. Nucleic Acids Res. 2014, 42, W314–W319. [Google Scholar] [CrossRef]
- Woolf, C.M. Common white facial markings in bay and chestnut Arabian horses and their hybrids. J. Hered. 1991, 82, 167–169. [Google Scholar] [CrossRef] [Green Version]
- Woolf, C.M. Common white facial markings in Arabian horses that are homozygous and heterozygous for alleles at the A and E loci. J. Hered. 1992, 83, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Hirobe, T.; Abe, H.; Wakamatsu, K.; Ito, S.; Kawa, Y.; Soma, Y.; Mizoguchi, M. Excess tyrosine rescues the reduced activity of proliferation and differentiation of cultured recessive yellow melanocytes derived from neonatal mouse epidermis. Eur. J. Cell Biol. 2007, 86, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Wakamatsu, K. Human hair melanins: What we have learned and have not learned from mouse coat color pigmentation. Pigment Cell Melanoma Res. 2011, 24, 63–74. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, S.A.N.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AICc | MC1R Extension | ASIP Agouti | KIT rs1140732842 | |
|---|---|---|---|---|
| AGW | 918.99 | |||
| AGW + MC1R Extension | 903.95 | 17.99 | ||
| AGW + ASIP Agouti | 915.54 | 5.57 | ||
| AGW + KIT rs1140732842 | 869.13 | 32.51 | ||
| AGW + MC1R Extension + KIT rs1140732842 | 843.23 | 30.26 | 40.51 | |
| AGW + MC1R Extension + ASIP Agouti | 904.51 | 13.51 | 1.54 | |
| AGW + MC1R Extension + ASIP Agouti + KIT rs1140732842 | 845.29 | 28.14 | 0.13 | 39.09 |
| p-value (full model) | 4.71 × 10−7 | 0.7199 | 5.09 × 10−14 | |
| p-values are given for the full model incorporating AGW and genetic effects. | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patterson Rosa, L.; Martin, K.; Vierra, M.; Lundquist, E.; Foster, G.; Brooks, S.A.; Lafayette, C. A KIT Variant Associated with Increased White Spotting Epistatic to MC1R Genotype in Horses (Equus caballus). Animals 2022, 12, 1958. https://doi.org/10.3390/ani12151958
Patterson Rosa L, Martin K, Vierra M, Lundquist E, Foster G, Brooks SA, Lafayette C. A KIT Variant Associated with Increased White Spotting Epistatic to MC1R Genotype in Horses (Equus caballus). Animals. 2022; 12(15):1958. https://doi.org/10.3390/ani12151958
Chicago/Turabian StylePatterson Rosa, Laura, Katie Martin, Micaela Vierra, Erica Lundquist, Gabriel Foster, Samantha A. Brooks, and Christa Lafayette. 2022. "A KIT Variant Associated with Increased White Spotting Epistatic to MC1R Genotype in Horses (Equus caballus)" Animals 12, no. 15: 1958. https://doi.org/10.3390/ani12151958