
Growth Performance, Bone Development and Phosphorus Metabolism in Chicks Fed Diets Supplemented with Phytase Are Associated with Alterations in Gut Microbiota


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Housing and Experimental Design
2.2. Growth Performance and Bone Development Analysis
2.3. Blood Biochemical Parameters Analysis
2.4. Phosphorus Digestibility and Emission Analysis
2.5. Gut Microbial Analysis
2.6. Statistical Analysis
3. Results
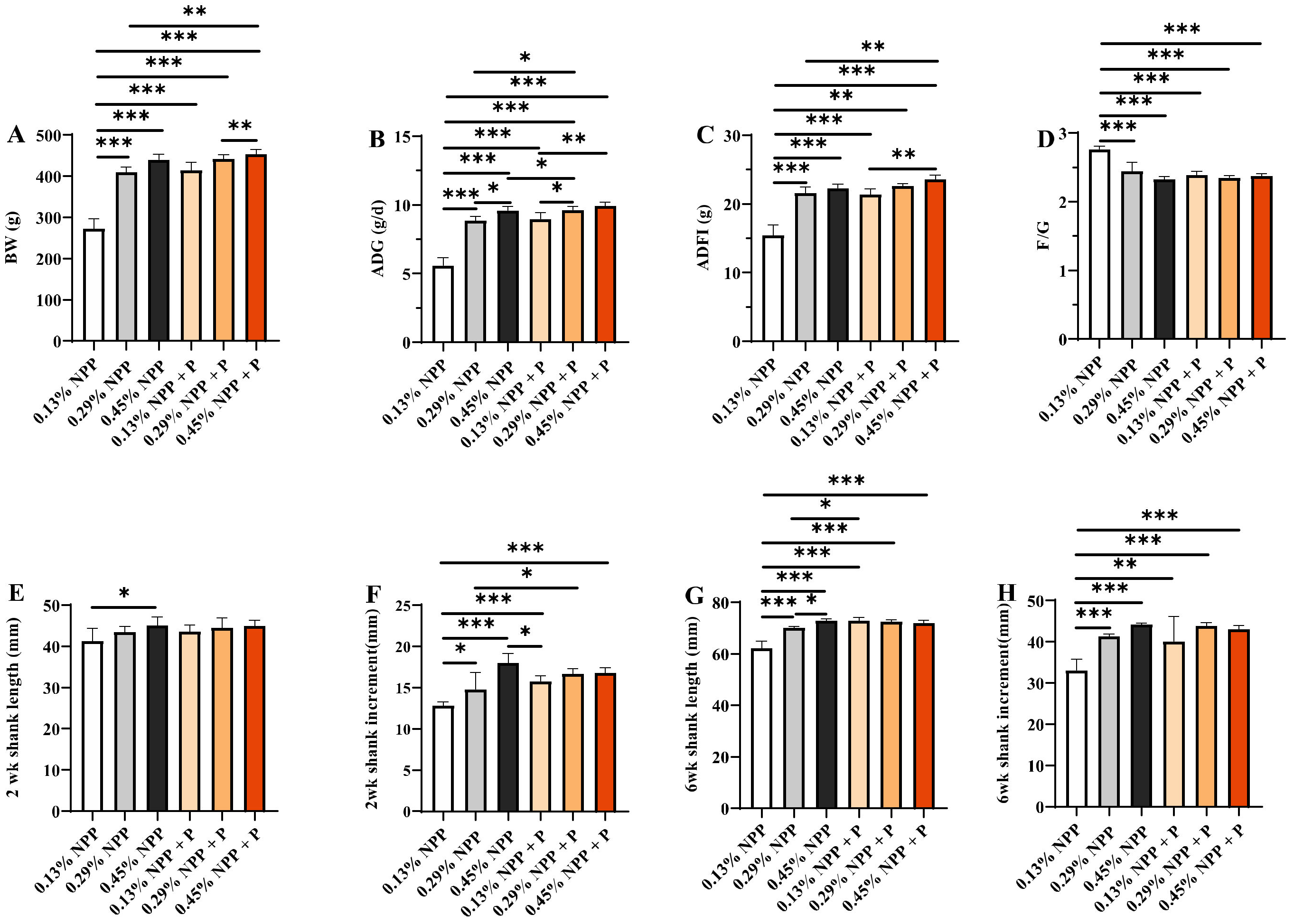
3.1. Growth Performance and Bone Development
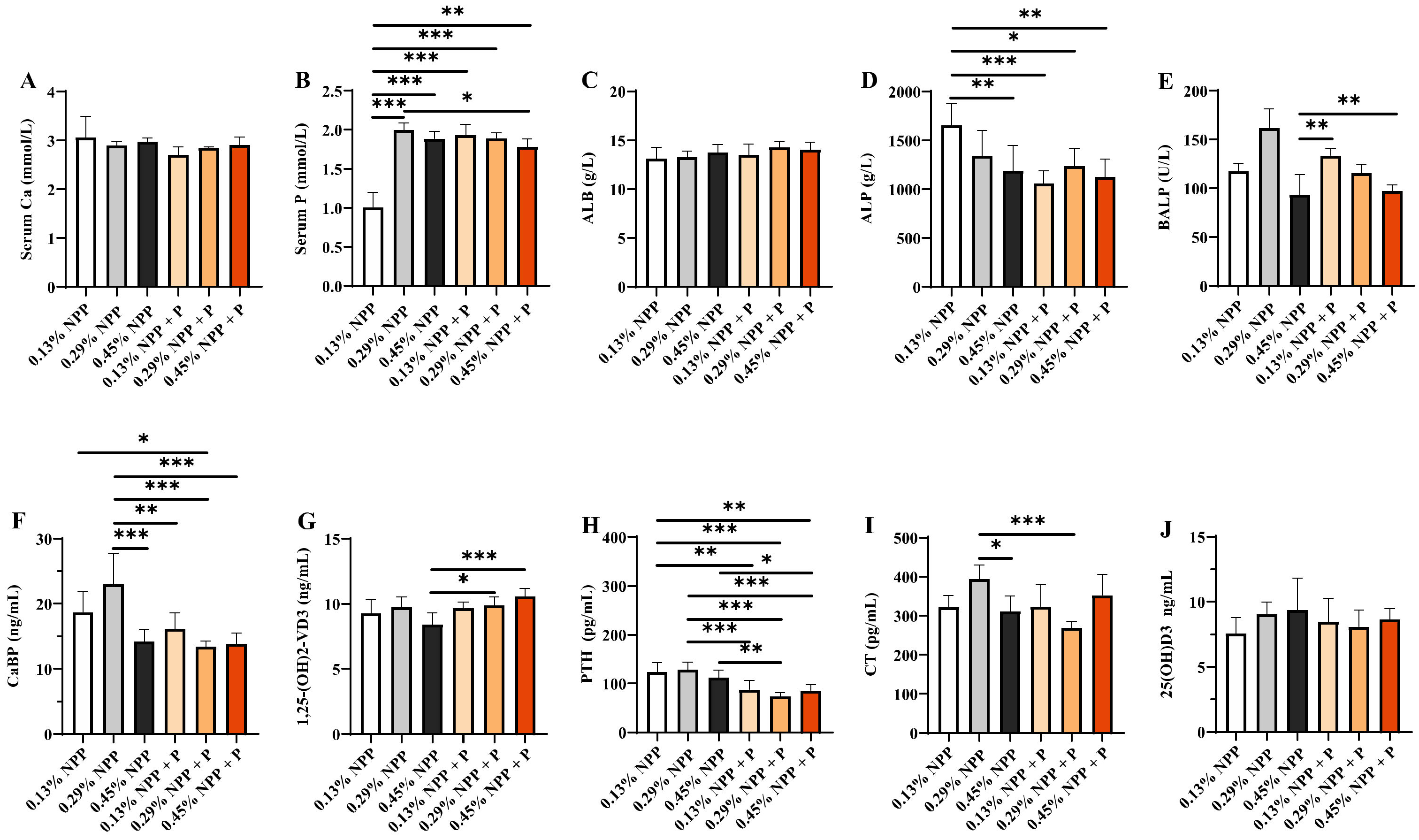
3.2. Serum Biochemical Index
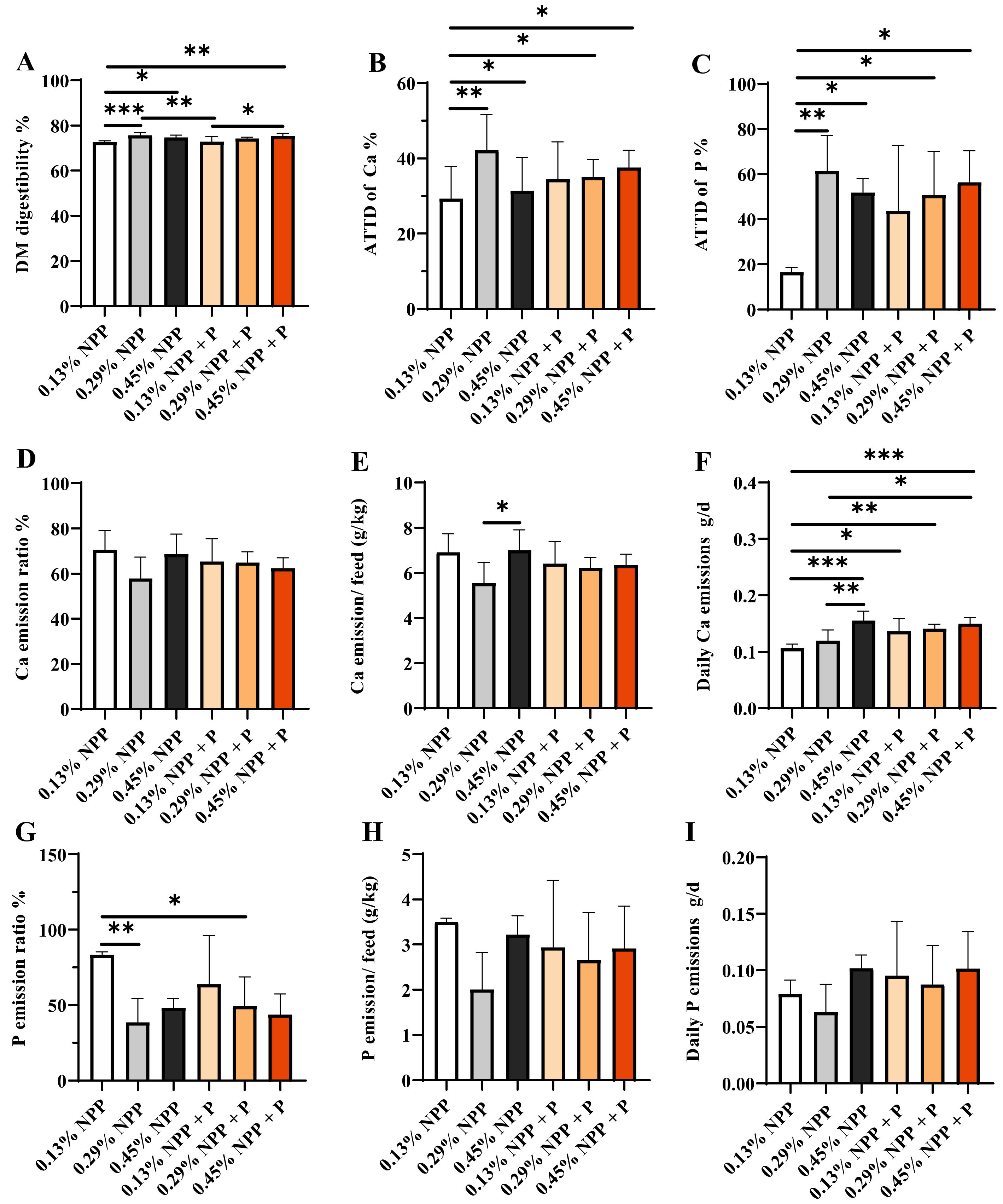
3.3. ATTD and Emission of Ca and Phosphorus
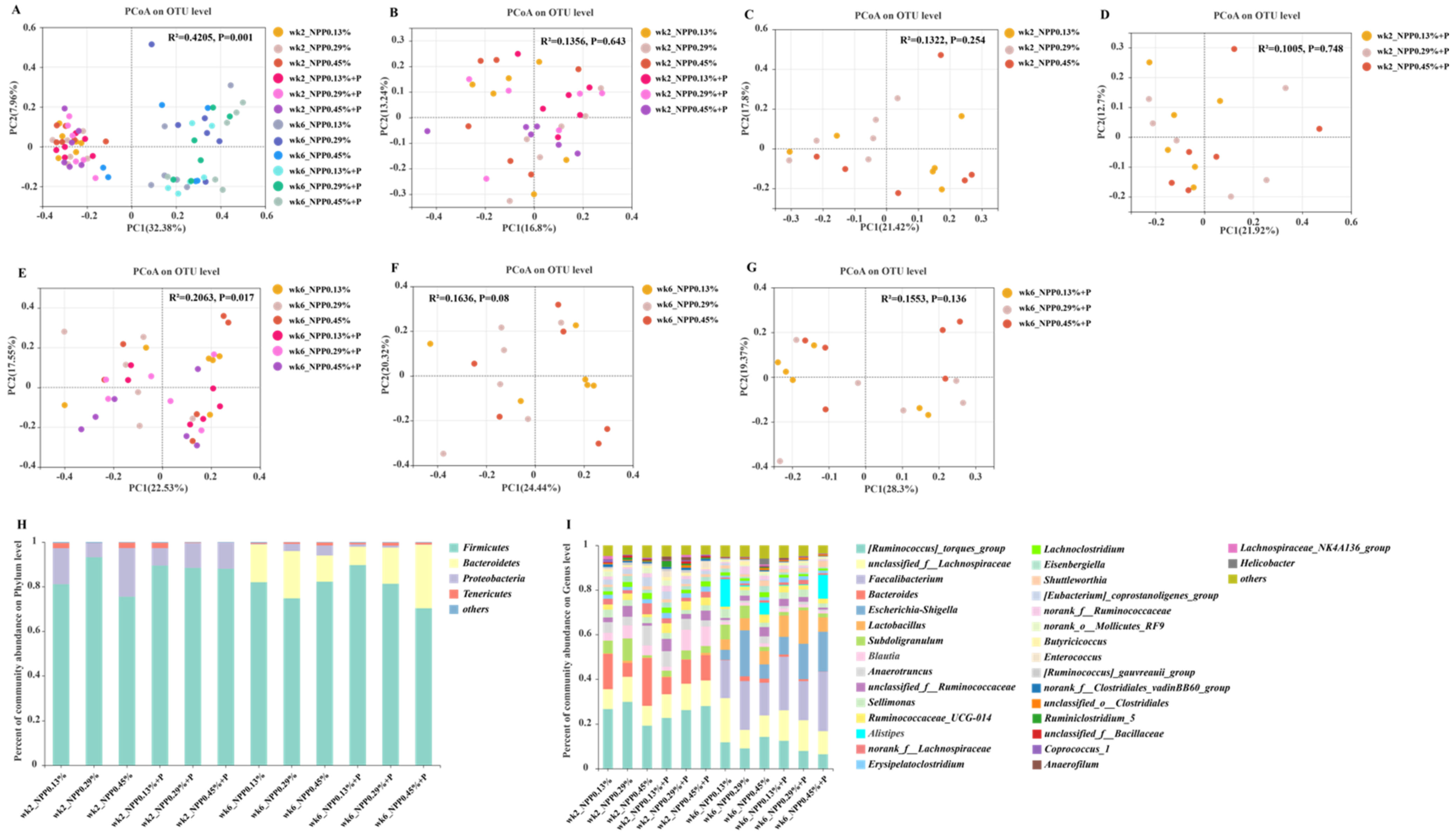
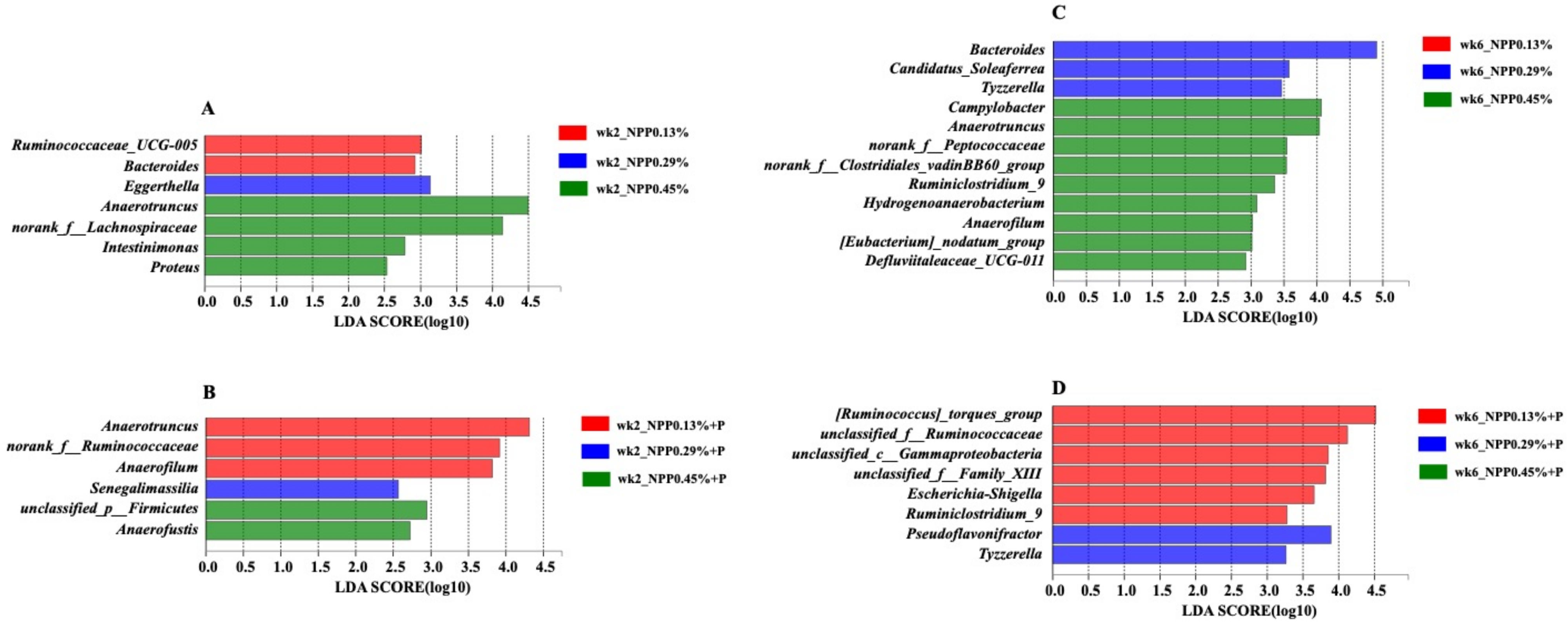
3.4. Gut Microbial Community and Structure
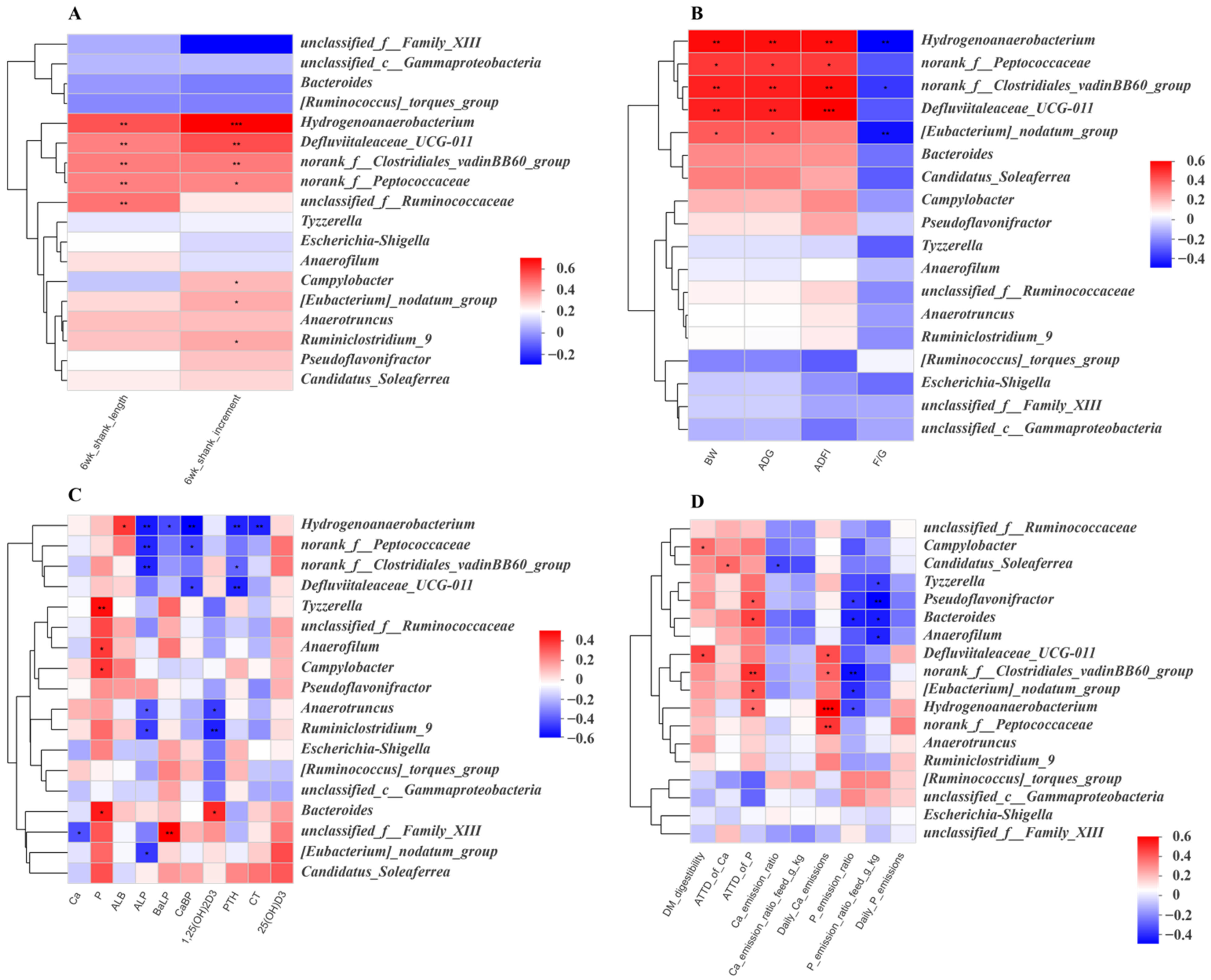
3.5. Correlation Analysis of the Gut Microbiota and Growth Performance and Phosphorus Metabolism Emission
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Soetan, K.O.; Olaiya, C.O.; Oyewole, O.E. The importance of mineral elements for humans, domestic animals and plants: A review. Afr. J. Food Sci. 2010, 4, 200–222. [Google Scholar]
- Heyer, C.M.; Weiss, E.; Schmucker, S.; Rodehutscord, M.; Hoelzle, L.E.; Mosenthin, R.; Stefanski, V. The impact of phosphorus on the immune system and the intestinal microbiota with special focus on the pig. Nutr. Res. Rev. 2015, 28, 67–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordell, D.; Drangert, J.-O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Qiugang, M.A.; Rodehutscord, M.; Novotny, M.; Lan, L.I.; Yang, L. Phytate and phosphorus utilization by broiler chickens and laying hens fed maize-based diets. Front. Agric. Sci. Eng. 2019, 6, 380–387. [Google Scholar]
- Gong, H.; Meng, F.; Wang, G.; Hartmann, T.E.; Feng, G.; Wu, J.; Jiao, X.; Zhang, F. Toward the sustainable use of mineral phosphorus fertilizers for crop production in China: From primary resource demand to final agricultural use. Sci. Total Environ. 2022, 804, 150183. [Google Scholar] [CrossRef]
- Azam, H.M.; Alam, S.T.; Hasan, M.; Yameogo, D.D.S.; Kannan, A.D.; Rahman, A.; Kwon, M.J. Phosphorous in the environment: Characteristics with distribution and effects, removal mechanisms, treatment technologies, and factors affecting recovery as minerals in natural and engineered systems. Environ. Sci. Pollut. Res. Int. 2019, 26, 20183–20207. [Google Scholar] [CrossRef]
- Kundu, S.; Coumar, V.M.; Rajendiran, S.; Ajay, R.; Subba, A. Phosphates from detergents and eutrophication of surface water ecosystem in India. Curr. Sci. 2015, 108, 1320–1325. [Google Scholar]
- Düngelhoef, M.; Rodehutscord, M.; Spiekers, H.; Pfeffer, E. Effects of supplemental microbial phytase on availability of phosphorus contained in maize, wheat and triticale to pigs. Anim. Feed Sci. Technol. 1994, 49, 1–10. [Google Scholar] [CrossRef]
- Singh, P.K. Significance of phytic acid and supplemental phytase in chicken nutrition: A review. Worlds Poult. Sci. J. 2008, 64, 553–580. [Google Scholar] [CrossRef]
- Lei, X.G.; Weaver, J.D.; Mullaney, E.; Ullah, A.H.; Azain, M.J. Phytase, a new life for an “old” enzyme. Annu. Rev. Anim. Biosci. 2013, 1, 283–309. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Jan, Y.C.; Chung, T.K.; Lee, T.T.; Chiou, P.W.S. Exogenous phytase activity in the gastrointestinal tract of broiler chickens. Anim. Feed Sci. Technol. 2004, 117, 295–303. [Google Scholar] [CrossRef]
- Ravindran, V. Feed enzymes: The science, practice, and metabolic realities. J. Appl. Poult. Res. 2013, 22, 628–636. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V. Microbial phytase in poultry nutrition. Anim. Feed Sci. Technol. 2007, 135, 1–41. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Fuhren, J.; Schwalbe, M.; Boekhorst, J.; Rösch, C.; Schols, H.A.; Kleerebezem, M. Dietary calcium phosphate strongly impacts gut microbiome changes elicited by inulin and galacto-oligosaccharides consumption. Microbiome 2021, 9, 218. [Google Scholar] [CrossRef]
- Ptak, A.; Bedford, M.R.; Świątkiewicz, S.; Żyła, K.; Józefiak, D. Phytase modulates ileal microbiota and enhances growth performance of the broiler chickens. PLoS ONE 2015, 10, e0119770. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Huang, S.; Zhao, L.; Zhang, J.; Ji, C.; Ma, Q. Pine (Pinus massoniana Lamb.) needle extract supplementation improves performance, egg quality, serum parameters, and the gut microbiome in laying hens. Front. Nutr. 2022, 9, 810462. [Google Scholar] [CrossRef]
- Sales, J.; Janssens, G. Acid-insoluble ash as a marker in digestibility studies: A review. J. Anim. Feed Sci. 2003, 12, 383–401. [Google Scholar] [CrossRef]
- Spowart, P.R.; Michael, G.; Easom, J.A.; Fritzler, L.D.; Lust, D.G.; Samuelson, K.L. PSI-13 Evaluation of acid insoluble ash as a digestibility marker in feedlot diets containing corn-milling byproducts. J. Anim. Sci. 2020, 98 (Suppl. 4), 273. [Google Scholar] [CrossRef]
- Geng, S.; Huang, S.; Ma, Q.; Li, F.; Gao, Y.; Zhao, L.; Zhang, J. Alterations and correlations of the gut microbiome, performance, egg quality, and serum biochemical indexes in laying hens with low-protein amino acid-deficient diets. ACS Omega 2021, 6, 13094–13104. [Google Scholar] [CrossRef]
- Ma, M.; Geng, S.; Liu, M.; Zhao, L.; Zhang, J.; Huang, S.; Ma, Q. Effects of different methionine levels in low protein diets on production performance, reproductive system, metabolism, and gut microbiota in laying hens. Front. Nutr. 2021, 8, 739676. [Google Scholar] [CrossRef]
- Elkin, R.G. A review of duck nutrition research. World’s Poult. Sci. J. 1987, 43, 84–106. [Google Scholar] [CrossRef]
- Lei, X.; Ku, P.; Miller, E.; Yokoyama, M. Supplementing corn-soybean meal diets with microbial phytase linearly improves phytate phosphorus utilization by weanling pigs. J. Anim. Sci. 1993, 71, 3359–3367. [Google Scholar] [CrossRef]
- Perney, K.M.; Cantor, A.H.; Straw, M.L.; Herkelman, K.L. The effect of dietary phytase on growth performance and phosphorus utilization of broiler chicks. Poult. Sci. 1993, 72, 2106–2114. [Google Scholar] [CrossRef]
- Ravindran, V.; Cowieson, A.J.; Selle, P.H. Influence of dietary electrolyte balance and microbial phytase on growth performance, nutrient utilization, and excreta quality of broiler chickens. Poult. Sci. 2008, 87, 677–688. [Google Scholar] [CrossRef]
- Li, L.; Zhang, X.; Zhao, L.; Zhang, J.; Ji, C.; Ma, Q. Phosphorus restriction in brooding stage has continuous effects on growth performance and early laying performance of layers. Animals 2021, 11, 3546. [Google Scholar] [CrossRef]
- Létourneau-Montminy, M.P.; Jondreville, C.; Sauvant, D.; Narcy, A. Meta-analysis of phosphorus utilization by growing pigs: Effect of dietary phosphorus, calcium and exogenous phytase. Animal 2012, 6, 1590–1600. [Google Scholar] [CrossRef]
- Farhadi, D.; Karimi, A.; Sadeghi, G.; Rostamzadeh, J.; Bedford, M.R. Effects of a high dose of microbial phytase and myo-inositol supplementation on growth performance, tibia mineralization, nutrient digestibility, litter moisture content, and foot problems in broiler chickens fed phosphorus-deficient diets. Poult. Sci. 2017, 96, 3664–3675. [Google Scholar] [CrossRef]
- Jiang, X.R.; Luo, F.H.; Qu, M.R.; Valentino, B. Effects of non-phytate phosphorus levels and phytase sources on growth performance, serum biochemical and tibia parameters of broiler chickens. Ital. J. Anim. Sci. 2013, 12, e60. [Google Scholar] [CrossRef]
- Shao, Y.; Sun, G.; Cao, S.; Lu, L.; Zhang, L.; Liao, X.; Luo, X. Bone phosphorus retention and bone development of broilers at different ages. Poult. Sci. 2019, 98, 2114–2121. [Google Scholar] [CrossRef] [PubMed]
- Brenes, A.; Viveros, A.; Arija, I.; Centeno, C.; Pizarro, M.; Bravo, C. The effect of citric acid and microbial phytase on mineral utilization in broiler chicks. Anim. Feed Sci. Technol. 2003, 110, 201–219. [Google Scholar] [CrossRef] [Green Version]
- Tilgar, V.; Kilgas, P.; Viitak, A.; Reynolds, S.J. The rate of bone mineralization in birds is directly related to alkaline phosphatase activity. Physiol. Biochem. Zool. 2008, 81, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Shao, P.; Ohtsuka-Isoya, M.; Shinoda, H. Circadian rhythms in serum bone markers and their relation to the effect of etidronate in rats. Chronobiol. Int. 2003, 20, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.S.; Kumar, R.; Heath, H. Persistently elevated parathyroid hormone secretion and action in young women after four weeks of ingesting high phosphorus, low calcium diets. J. Clin. Endocrinol. Metab. 1990, 70, 1334–1340. [Google Scholar] [CrossRef]
- Krajmalnik-Brown, R.; Ilhan, Z.E.; Kang, D.W.; Dibaise, J.K. Effects of gut microbes on nutrient absorption and energy regulation. Nutr. Clin. Pract. Off. Publ. Am. Soc. Parenter. Enter. Nutr. 2015, 27, 201–214. [Google Scholar] [CrossRef] [Green Version]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Re: Role of the gut microbiota in nutrition and health. BMJ 2018, 361, 17–22. [Google Scholar]
- Yoo, J.Y.; Groer, M.; Dutra, S.V.O.; Sarkar, A.; McSkimming, D.I. Gut microbiota and immune system interactions. Microorganisms 2020, 8, 1587. [Google Scholar] [CrossRef]
- Wu, H.J.; Wu, E. The role of gut microbiota in immune homeostasis and autoimmunity. Gut Microbes 2012, 3, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef]
- Wood, H.G.; Clark, J.E. Biological aspects of inorganic polyphosphates. Annu. Rev. Biochem. 1988, 57, 235–260. [Google Scholar] [CrossRef]
- Francis, G.L.; Gawthorne, J.M.; Storer, G.B. Factors affecting the activity of cellulases isolated from the rumen digesta of sheep. Appl. Environ. Microbiol. 1978, 36, 643–649. [Google Scholar] [CrossRef] [Green Version]
- Montagne, L.; Pluske, J.R.; Hampson, D.J. A review of interactions between dietary fibre and the intestinal mucosa, and their consequences on digestive health in young non-ruminant animals. Anim. Feed Sci. Technol. 2003, 108, 95–117. [Google Scholar] [CrossRef]
- Joat, N.; Van, T.T.H.; Stanley, D.; Moore, R.J.; Chousalkar, K. Temporal dynamics of gut microbiota in caged laying hens: A field observation from hatching to end of lay. Appl. Microbiol. Biotechnol. 2021, 105, 4719–4730. [Google Scholar] [CrossRef]
- Li, W.; Ma, Z.S. FBA Ecological guild: Trio of firmicutes-bacteroidetes alliance against actinobacteria in human oral microbiome. Sci. Rep. 2020, 10, 287. [Google Scholar] [CrossRef] [Green Version]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The influence of probiotics on the firmicutes/bacteroidetes ratio in the treatment of obesity and inflammatory bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Liao, X.; Shao, Y.; Sun, G.; Yang, Y.; Zhang, L.; Guo, Y.; Luo, X.; Lu, L. The relationship among gut microbiota, short-chain fatty acids, and intestinal morphology of growing and healthy broilers. Poult. Sci. 2020, 99, 5883–5895. [Google Scholar] [CrossRef]
- Den Besten, G.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Emiola, A.; Akinremi, O.; Slominski, B.; Nyachoti, C.M. Nutrient utilization and manure P excretion in growing pigs fed corn-barley-soybean based diets supplemented with microbial phytase. Anim. Sci. J. 2009, 80, 19–26. [Google Scholar] [CrossRef]
- Plumstead, P.W.; Romero-Sanchez, H.; Maguire, R.O.; Gernat, A.G.; Brake, J. Effects of phosphorus level and phytase in broiler breeder rearing and laying diets on live performance and phosphorus excretion. Poult. Sci. 2007, 86, 225–231. [Google Scholar] [CrossRef]
- Yitbarek, A.; López, S.; Tenuta, M.; Asgedom, H.; France, J.; Nyachoti, C.M.; Kebreab, E. Effect of dietary phytase supplementation on greenhouse gas emissions from soil after swine manure application. J. Clean. Prod. 2017, 166, 1122–1130. [Google Scholar] [CrossRef]
- Müller, C.B.M.; Kuhla, B. Holstein dairy cows with high phosphorus utilization efficiency fed a low phosphorous diet secreted less phosphorus with urine but more with milk and feces. Sci. Total Environ. 2021, 788, 147813. [Google Scholar] [CrossRef]
- Chang, C.L.; Chung, C.Y.; Kuo, C.H.; Kuo, T.F.; Yang, C.W.; Yang, W.C. Beneficial effect of bidens pilosa on body weight gain, food conversion ratio, gut bacteria and coccidiosis in chickens. PLoS ONE 2016, 11, e0146141. [Google Scholar] [CrossRef]
- Ponsuksili, S.; Reyer, H.; Hadlich, F.; Weber, F.; Trakooljul, N.; Oster, M.; Siengdee, P.; Muráni, E.; Rodehutscord, M.; Camarinha-Silva, A.; et al. Identification of the key molecular drivers of phosphorus utilization based on host miRNA-mRNA and gut microbiome interactions. Int J. Mol. Sci 2020, 21, 2818. [Google Scholar] [CrossRef]
- Bosman, E.S.; Albert, A.Y.; Lui, H.; Dutz, J.P.; Vallance, B.A. Skin exposure to narrow band ultraviolet (UVB) light modulates the human intestinal microbiome. Front. Microbiol. 2019, 10, 2410. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Treatment Groups | ||
|---|---|---|---|
| 0.13% NPP | 0.29% NPP | 0.45% NPP | |
| Ingredients | |||
| Corn | 65.20 | 65.20 | 65.20 |
| Soybean meal | 29.40 | 29.40 | 29.40 |
| Dicalcium phosphate | 0.00 | 0.95 | 1.90 |
| Limestone | 2.35 | 1.82 | 1.30 |
| Zeolite powder | 2.01 | 1.59 | 1.16 |
| Salt | 0.30 | 0.30 | 0.30 |
| Choline chloride (50%) | 0.10 | 0.10 | 0.10 |
| L-Lysine HCl (98%) | 0.11 | 0.11 | 0.11 |
| DL-methionine | 0.19 | 0.19 | 0.19 |
| Premix 1 | 0.34 | 0.34 | 0.34 |
| Total | 100.00 | 100.00 | 100.00 |
| Nutrient levels 2 | |||
| Metabolizable energy (MJ/kg) | 11.72 | 11.72 | 11.72 |
| Crude protein | 18.00 | 18.00 | 18.00 |
| Available methionine | 0.45 | 0.45 | 0.45 |
| Available methionine and cystine | 0.74 | 0.74 | 0.74 |
| Available Lysine | 1.00 | 1.00 | 1.00 |
| Available Tryptophan | 0.20 | 0.20 | 0.20 |
| Available Threonine | 0.68 | 0.68 | 0.68 |
| Calcium | 0.98 | 0.96 | 1.02 |
| Total phosphorus | 0.42 | 0.51 | 0.67 |
| Non-phytate phosphorus | 0.13 | 0.29 | 0.45 |
| Phytate phosphorus | 0.29 | 0.22 | 0.22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Zhang, X.; Zhang, J.; Liu, M.; Zhao, L.; Ji, C.; Zhang, J.; Huang, S.; Ma, Q. Growth Performance, Bone Development and Phosphorus Metabolism in Chicks Fed Diets Supplemented with Phytase Are Associated with Alterations in Gut Microbiota. Animals 2022, 12, 940. https://doi.org/10.3390/ani12070940
Li L, Zhang X, Zhang J, Liu M, Zhao L, Ji C, Zhang J, Huang S, Ma Q. Growth Performance, Bone Development and Phosphorus Metabolism in Chicks Fed Diets Supplemented with Phytase Are Associated with Alterations in Gut Microbiota. Animals. 2022; 12(7):940. https://doi.org/10.3390/ani12070940
Chicago/Turabian StyleLi, Lan, Xiaoyi Zhang, Jiatu Zhang, Meiling Liu, Lihong Zhao, Cheng Ji, Jianyun Zhang, Shimeng Huang, and Qiugang Ma. 2022. "Growth Performance, Bone Development and Phosphorus Metabolism in Chicks Fed Diets Supplemented with Phytase Are Associated with Alterations in Gut Microbiota" Animals 12, no. 7: 940. https://doi.org/10.3390/ani12070940