
Influence of Immunocastration and Diet on Meat and Fat Quality of Heavy Female and Male Pigs

 , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Pig Husbandry and Experimental Design
2.2. Feed Analyses
2.3. Slaughtering and Meat and Fat Sampling
2.4. Meat Quality Traits
2.5. Fatty Acid Profile of the Subcutaneous Fat
2.6. Boar Taint Compounds
2.7. Statistical Analyses
3. Results and Discussion
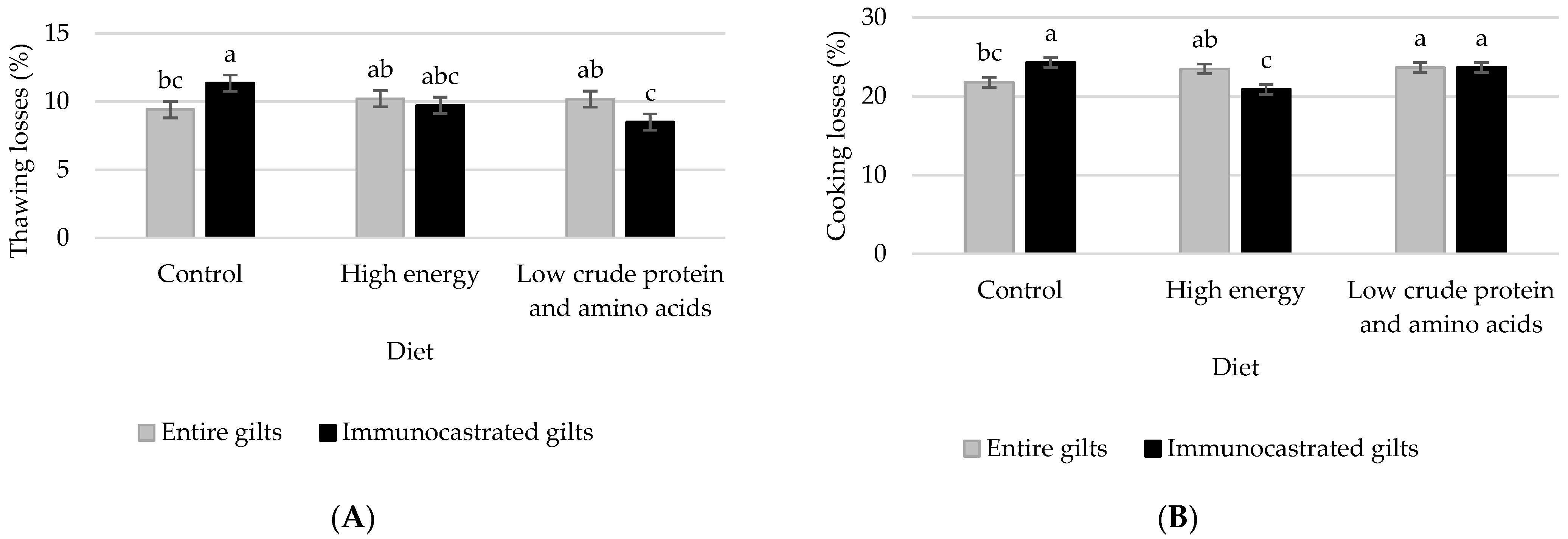
3.1. Trial 1: Meat Quality and Fat Composition in Female Pigs
3.2. Trial 2: Meat Quality, Fat Composition and Boar Taint Compounds in Male Pigs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Latorre, M.A.; Ripoll, G.; García-Belenguer, E.; Ariño, L. The increase of slaughter weight in gilts as a strategy to optimize the production of Spanish high quality dry-cured ham. J. Anim. Sci. 2009, 87, 1464–1471. [Google Scholar] [CrossRef] [Green Version]
- Bosi, P.; Russo, V. The production of the heavy pig for high quality processed products. Ital. J. Anim. Sci. 2004, 3, 309–321. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Sánchez, J.A.; Calvo, S.; Suárez-Belloch, J.; Latorre, M.A. Effect of pig slaughter weight on chemical and sensory characteristics of Teruel dry-cured ham. Ital. J. Food Sci. 2014, 26, 420–426. [Google Scholar]
- Ruiz-Carrascal, J.; Ventanas, J.; Cava, R.; Andrés, A.I.; García, C. Texture and appearance of dry cured ham as affected by fat content and fatty acid composition. Food Res. Int. 2000, 33, 91–95. [Google Scholar] [CrossRef]
- Latorre, M.A.; Ripoll, G.; García-Belenguer, E.; Ariño, L. The effect of gender and slaughter weight on loin and fat characteristics of pigs intended for Teruel dry-cured ham production. Span. J. Agric. Res. 2009, 7, 407–416. [Google Scholar] [CrossRef]
- Weatherup, R.N.; Beattie, V.E.; Moss, B.W.; Kilpatrick, D.J.; Walker, N. The effect of increasing slaughter weight on the production performance and meat quality of finishing pigs. Anim. Sci. 1998, 67, 591–600. [Google Scholar] [CrossRef]
- Peinado, J.; Medel, P.; Fuentetaja, A.; Mateos, G.G. Influence of sex and castration of females on growth performance and carcass and meat quality of heavy pigs destined for the dry-cured industry. J. Anim. Sci. 2008, 86, 1410–1417. [Google Scholar] [CrossRef] [Green Version]
- Suarez-Belloch, J.; Sanz, M.A.; Joy, M.; Latorre, M.A. Impact of increasing dietary energy level during the finishing period on growth performance, pork quality and fatty acid profile in heavy pigs. Meat Sci. 2013, 93, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Belloch, J.; Latorre, M.A.; Guada, J.A. The effect of protein restriction during the growing period on carcass, meat and fat quality of heavy barrows and gilts. Meat Sci. 2016, 112, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Council Directive 2008/120/EC of 18 December 2008 laying down minimum standards for the protection of pigs. Off. J. Eur. Union 2009, L 47, 5–13. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32008L0120 (accessed on 23 July 2021).
- Poulsen-Nautrup, B.; Van Vlaenderen, I.; Aldaz, A.; Mah, C.K. The effect of immunization against gonadotropin-releasing factor on growth performance, carcass characteristics and boar taint relevant to pig producers and the pork packing industry: A meta-analysis. Res. Vet. Sci. 2018, 119, 182–195. [Google Scholar] [CrossRef]
- Real Decreto 53/2013, de 1 de febrero, por el que se establecen las normas básicas aplicables para la protección de los animales utilizados en experimentación y otros fines científicos, incluyendo la docencia. BOE 2013, 34, 11370–11421. Available online: https://www.boe.es/diario_boe/txt.php?id=BOE-A-2013-1337 (accessed on 23 July 2021).
- FEDNA. Necesidades Nutricionales Para Ganado Porcino: Normas FEDNA, 2nd ed.; De Blas, C., Gasa, J., Mateos, G.G., Eds.; Fundación Española para el Desarrollo de la Nutrición Animal: Madrid, Spain, 2013. [Google Scholar]
- Pérez-Ciria, L.; Miana-Mena, F.J.; Falceto, M.V.; Mitjana, O.; Latorre, M.A. Effect of immunocastration and diet on growth performance, serum metabolites and sex hormones, reproductive organ development and carcass quality of heavy gilts. Animals 2021, 11, 1900. [Google Scholar] [CrossRef] [PubMed]
- Sukhija, P.S.; Palmquist, D.L. Rapid method for determination of total fatty acid content and composition of feedstuffs and feces. J. Agric. Food Chem. 1988, 36, 1202–1206. [Google Scholar] [CrossRef]
- Lopez-Bote, C.J.; Isabel, B.; Ruiz, J.; Daza, A. Effect of vitamin E supplementation and partial substitution of poly- with mono-unsaturated fatty acids in pig diets on muscle, and microsome extract α-tocopherol concentration and lipid oxidation. Arch. Anim. Nutr. 2003, 57, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Ministério da Agricultura, Pecuária e Abastecimento. Informação Diversa no 061, de 23 de Abril de 2007; Autorização para Abate de Suínos Imunocastrados: Brasília, Brazil, 2007.
- CIE. Colorimetry, 2nd ed.; Commission Internationale de L’Eclairage: Vienna, Austria, 1986. [Google Scholar]
- Honikel, K.O. Reference methods for the assessment of physical characteristics of meat. Meat Sci. 1998, 49, 447–457. [Google Scholar] [CrossRef]
- Ripoll, G.; Alcalde, M.J.; Córdoba, M.G.; Casquete, R.; Argüello, A.; Ruiz-Moyano, S.; Panea, B. Influence of the use of milk replacers and pH on the texture profiles of raw and cooked meat of suckling kids. Foods 2019, 8, 589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orden de 31 de julio de 1979 por la que se establecen métodos oficiales de análisis de aceites y grasas, productos cárnicos, cereales y derivados, fertilizantes, productos fitosanitarios, productos lácteos, piensos, aguas y productos derivados de la uva. BOE 1979, 207, 20221–20247. Available online: https://www.boe.es/buscar/doc.php?id=BOE-A-1979-21118 (accessed on 23 July 2021).
- Hansen-Møller, J. Rapid high-performance liquid chromatographic method for simultaneous determination of androstenone, skatole and indole in back fat from pigs. J. Chromatogr. B Biomed. Sci. Appl. 1994, 661, 219–230. [Google Scholar] [CrossRef]
- Pauly, C.; Spring, P.; O’Doherty, J.V.; Ampuero Kragten, S.; Bee, G. Performances, meat quality and boar taint of castrates and entire male pigs fed a standard and a raw potato starch-enriched diet. Animal 2008, 2, 1707–1715. [Google Scholar] [CrossRef] [Green Version]
- Batorek, N.; Škrlep, M.; Prunier, A.; Louveau, I.; Noblet, J.; Bonneau, M.; Čandek-Potokar, M. Effect of feed restriction on hormones, performance, carcass traits, and meat quality in immunocastrated pigs. J. Anim. Sci. 2012, 90, 4593–4603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohrer, B.M.; Flowers, W.L.; Kyle, J.M.; Johnson, S.S.; King, V.L.; Spruill, J.L.; Thompson, D.P.; Schroeder, A.L.; Boler, D.D. Effect of gonadotropin releasing factor suppression with an immunological on growth performance, estrus activity, carcass characteristics, and meat quality of market gilts. J. Anim. Sci. 2014, 92, 4719–4724. [Google Scholar] [CrossRef] [Green Version]
- Van den Broeke, A.; Leen, F.; Aluwé, M.; Ampe, B.; Van Meensel, J.; Millet, S. The effect of GnRH vaccination on performance, carcass, and meat quality and hormonal regulation in boars, barrows, and gilts. J. Anim. Sci. 2016, 94, 2811–2820. [Google Scholar] [CrossRef]
- Pérez-Ciria, L.; Carcò, G.; Miana-Mena, F.J.; Mitjana, O.; Falceto, M.V.; Latorre, M.A. Immunocastration in gilts: A preliminary study of the effect of the second dose administration time on growth, reproductive tract development, and carcass and meat quality. Animals 2021, 11, 510. [Google Scholar] [CrossRef] [PubMed]
- Daza, A.; Latorre, M.A.; Olivares, A.; López-Bote, C.J. The effect of immunocastration and a diet based on granulated barley on growth performance and carcass, meat and fat quality in heavy gilts. Animal 2014, 8, 484–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamero-Negrón, R.; Sánchez del Pulgar, J.; Ventanas, J.; García, C. Immune-spaying as an alternative to surgical spaying in Iberian×Duroc females: Effect on carcass traits and meat quality characteristics. Meat Sci. 2015, 99, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Macipe, M.; Rodríguez, P.; Izquierdo, M.; Gispert, M.; Manteca, X.; Mainau, E.; Hernández, F.I.; Claret, A.; Guerrero, L.; Dalmau, A. Comparison of meat quality parameters in surgical castrated versus vaccinated against gonadotrophin-releasing factor male and female Iberian pigs reared in free-ranging conditions. Meat Sci. 2016, 111, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Zheng, W.; Zhang, F.; Rao, S.; Peng, Z.; Yao, W. Effect of immunocastration on growth performance, gonadal development and carcass and meat quality of SuHuai female pigs. Anim. Prod. Sci. 2019, 59, 794–800. [Google Scholar] [CrossRef]
- Suárez-Belloch, J.; Guada, J.A.; Latorre, M.A. Effects of sex and dietary lysine on performances and serum and meat traits in finisher pigs. Animal 2015, 9, 1731–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Sánchez, J.A.; Sanz, M.A.; Blanco, M.; Serrano, M.P.; Joy, M.; Latorre, M.A. The influence of dietary lysine restriction during the finishing period on growth performance and carcass, meat, and fat characteristics of barrows and gilts intended for dry-cured ham production. J. Anim. Sci. 2011, 89, 3651–3662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teye, G.A.; Sheard, P.R.; Whittington, F.M.; Nute, G.R.; Stewart, A.; Wood, J.D. Influence of dietary oils and protein level on pork quality. 1. Effects on muscle fatty acid composition, carcass, meat and eating quality. Meat Sci. 2006, 73, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Daza, A.; Latorre, M.A.; Olivares, A.; López Bote, C.J. The effects of male and female immunocastration on growth performances and carcass and meat quality of pigs intended for dry-cured ham production: A preliminary study. Livest. Sci. 2016, 190, 20–26. [Google Scholar] [CrossRef]
- Gandemer, G. Lipids in muscles and adipose tissues, changes during processing and sensory properties of meat products. Meat Sci. 2002, 62, 309–321. [Google Scholar] [CrossRef]
- Škrlep, M.; Tomašević, I.; Mörlein, D.; Novaković, S.; Egea, M.; Garrido, M.D.; Linares, M.B.; Peñaranda, I.; Aluwé, M.; Font-I-furnols, M. The use of pork from entire male and immunocastrated pigs for meat products—an overview with recommendations. Animals 2020, 10, 1754. [Google Scholar] [CrossRef]
- Hugo, A.; Roodt, E. Significance of porcine fat quality in meat technology: A review. Food Rev. Int. 2007, 23, 175–198. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Y.; Liang, X.; Wu, X.; Liu, J.; Yang, S.; Tao, C.; Zhang, J.; Tian, J.; Zhao, J.; et al. Stearoyl-CoA desaturase is essential for porcine adipocyte differentiation. Int. J. Mol. Sci. 2020, 21, 2446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauly, C.; Spring, P.; Odoherty, J.V.; Ampuero Kragten, S.; Bee, G. Growth performance, carcass characteristics and meat quality of group-penned surgically castrated, immunocastrated (Improvac) and entire male pigs and individually penned entire male pigs. Animal 2009, 3, 1057–1066. [Google Scholar] [CrossRef] [Green Version]
- Seiquer, I.; Palma-Granados, P.; Haro, A.; Lara, L.; Lachica, M.; Fernández-Fígares, I.; Nieto, R. Meat quality traits in longissimus lumborum and gluteus medius muscles from immunocastrated and surgically castrated Iberian pigs. Meat Sci. 2019, 150, 77–84. [Google Scholar] [CrossRef]
- Aluwé, M.; Langendries, K.C.M.; Bekaert, K.M.; Tuyttens, F.A.M.; Brabander, D.L.D.; De Smet, S.; Millet, S. Effect of surgical castration, immunocastration and chicory-diet on the meat quality and palatability of boars. Meat Sci. 2013, 94, 402–407. [Google Scholar] [CrossRef]
- Škrlep, M.; Poklukar, K.; Kress, K.; Vrecl, M.; Fazarinc, G.; Lukač, N.B.; Weiler, U.; Stefanski, V.; Čandek-Potokar, M. Effect of immunocastration and housing conditions on pig carcass and meat quality traits. Transl. Anim. Sci. 2020, 4, 1224–1237. [Google Scholar] [CrossRef] [PubMed]
- Zanardi, E.; Novelli, E.; Ghiretti, G.P.; Dorigoni, V.; Chizzolini, R. Colour stability and vitamin E content of fresh and processed pork. Food Chem. 1999, 67, 163–171. [Google Scholar] [CrossRef]
- Boler, D.D.; Puls, C.L.; Clark, D.L.; Ellis, M.; Schroeder, A.L.; Matzat, P.D.; Killefer, J.; Mckeith, F.K.; Dilger, A.C. Effects of immunological castration (Improvest) on changes in dressing percentage and carcass characteristics of finishing pigs. J. Anim. Sci. 2014, 92, 359–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Gariépy, C.; Jin, Y.; Font i Furnols, M.; Fortin, J.; Rocha, L.M.; Faucitano, L. Effects of ractopamine administration and castration method on muscle fiber characteristics and sensory quality of the longissimus muscle in two Piétrain pig genotypes. Meat Sci. 2015, 102, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Škrlep, M.; Batorek, N.; Bonneau, M.; Prevolnik, M.; Kubale, V.; Čandek-Potokar, M. Effect of immunocastration in group-housed commercial fattening pigs on reproductive organs, malodorous compounds, carcass and meat quality. Czech J. Anim. Sci. 2012, 57, 290–299. [Google Scholar] [CrossRef] [Green Version]
- Caldara, F.R.; Moi, M.; Dos Santos, L.S.; De Lima Almeida Paz, I.C.; Garcia, R.G.; De Alencar Nääs, I.; Fernandes, A.R.M. Carcass characteristics and qualitative attributes of pork from immunocastrated animals. Asian-Australas. J. Anim. Sci. 2013, 26, 1630–1636. [Google Scholar] [CrossRef] [Green Version]
- Andreo, N.; Bridi, A.M.; Da Silva, C.A.; Peres, L.M.; De Lima Giangareli, B.; Dos Santos, É.R.; Rogel, C.P.; Vero, J.G.; Ferreira, G.A. Immunocastration and its effects on carcass and meat traits of male pigs. Semin. Agrar. 2018, 39, 2531–2540. [Google Scholar] [CrossRef]
- Mancini, R.A.; Hunt, M.C. Current research in meat color. Meat Sci. 2005, 71, 100–121. [Google Scholar] [CrossRef]
- Yamsakul, P.; Patchanee, P.; Yano, T.; Boonma, T.; Somkert, C.; Sathanawongs, A. Gonadotropin-releasing hormone immunization and castration in male pigs: Effects on growth, hormonal levels, antibody titer response, testicular function, back fat, and consumers’ sensory perceptions. Korean J. Vet. Res. 2017, 57, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Wittert, G.A.; Chapman, I.M.; Haren, M.T.; Mackintosh, S.; Coates, P.; Morley, J.E. Oral testosterone supplementation increases muscle and decreases fat mass in healthy elderly males with low-normal gonadal status. J. Gerontol. A Biol. Sci. Med. Sci. 2003, 58, 618–625. [Google Scholar] [CrossRef] [Green Version]
- Font-I-Furnols, M.; Gispert, M.; Soler, J.; Diaz, M.; Garcia-Regueiro, J.A.; Diaz, I.; Pearce, M.C. Effect of vaccination against gonadotrophin-releasing factor on growth performance, carcass, meat and fat quality of male Duroc pigs for dry-cured ham production. Meat Sci. 2012, 91, 148–154. [Google Scholar] [CrossRef]
- Huff-Lonergan, E.; Baas, T.J.; Malek, M.; Dekkers, J.C.M.; Prusa, K.; Rothschild, M.F. Correlations among selected pork quality traits. J. Anim. Sci. 2002, 80, 617–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, J.O.; Higbie, A.D.; Southern, L.L.; Coombs, D.F.; Bidner, T.D.; Odgaard, R.L. Effect of chromium propionate and metabolizable energy on growth, carcass traits, and pork quality of growing-finishing pigs. J. Anim. Sci. 2003, 81, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Sirtori, F.; Crovetti, A.; Acciaioli, A.; Pugliese, C.; Bozzi, R.; Campodoni, G.; Franci, O. Effect of dietary protein level on carcass traits and meat properties of Cinta Senese pigs. Animal 2014, 8, 1987–1995. [Google Scholar] [CrossRef] [PubMed]
- Tejeda, J.F.; Hernández-Matamoros, A.; Paniagua, M.; González, E. Effect of free-range and low-protein concentrated diets on growth performance, carcass traits, and meat composition of Iberian pig. Animals 2020, 10, 273. [Google Scholar] [CrossRef] [Green Version]
- Millet, S.; Ongenae, E.; Hesta, M.; Seynaeve, M.; De Smet, S.; Janssens, G.P.J. The feeding of ad libitum dietary protein to organic growing-finishing pigs. Vet. J. 2006, 171, 483–490. [Google Scholar] [CrossRef]
- Liu, Z.H.; Yang, F.Y.; Kong, L.J.; Lai, C.H.; Piao, X.S.; Gu, Y.H.; Ou, X.Q. Effects of dietary energy density on growth, carcass quality and mRNA expression of fatty acid synthase and hormone-sensitive lipase in finishing pigs. Asian-Australas. J. Anim. Sci. 2007, 20, 1587–1593. [Google Scholar] [CrossRef]
- Mackay, J.; Pearce, M.C.; Thevasagayam, S.; Doran, O. Fatty acid composition and lipogenic enzyme protein expression in subcutaneous adipose tissue of male pigs vaccinated against boar taint, barrows, and entire boars. J. Anim. Sci. 2013, 91, 395–404. [Google Scholar] [CrossRef] [Green Version]
- Asmus, M.D.; Tavarez, M.A.; Tokach, M.D.; Dritz, S.S.; Schroeder, A.L.; Nelssen, J.L.; Goodband, R.D.; DeRouchey, J.M. The effects of immunological castration and corn dried distillers grains with solubles withdrawal on growth performance, carcass characteristics, fatty acid analysis, and iodine value of pork fat depots. J. Anim. Sci. 2014, 92, 2116–2132. [Google Scholar] [CrossRef] [Green Version]
- Costa E Silva, L.C.; Barbosa, R.D.; Silveira, E.T.F. Effects of ractopamine hydrochloride and immunological castration in pigs. Part 2: Belly quality characteristics and fatty acid composition. Food Sci. Technol. 2017, 37, 404–410. [Google Scholar] [CrossRef] [Green Version]
- Madsen, A.; Jakobsen, K.; Mortensen, H.P. Influence of dietary fat on carcass fat quality in pigs. A review. Acta Agric. Scand. A Anim. Sci. 1992, 42, 220–225. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M. Monounsaturated fatty acids and risk of cardiovascular disease. Circulation 1999, 100, 1253–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, G.L. Dietary n-6 and n-3 polyunsaturated fatty acids: From biochemistry to clinical implications in cardiovascular prevention. Biochem. Pharmacol. 2009, 77, 937–946. [Google Scholar] [CrossRef]
- Madeira, M.S.; Pires, V.M.R.; Alfaia, C.M.; Costa, A.S.H.; Luxton, R.; Doran, O.; Bessa, R.J.B.; Prates, J.A.M. Differential effects of reduced protein diets on fatty acid composition and gene expression in muscle and subcutaneous adipose tissue of Alentejana purebred and Large White × Landrace × Pietrain crossbred pigs. Br. J. Nutr. 2013, 110, 216–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walstra, P.; Chevillon, P.; Von Seth, G.; Diestre, A.; Matthews, K.R.; Homer, D.B.; Bonneau, M. An international study on the importance of androstenone and skatole for boar taint: Levels of androstenone and skatole by country and season. Livest. Prod. Sci. 1999, 62, 15–28. [Google Scholar] [CrossRef]
- Zamaratskaia, G.; Andersson, H.K.; Chen, G.; Andersson, K.; Madej, A.; Lundström, K. Effect of a gonadotropin-releasing hormone vaccine (ImprovacTM) on steroid hormones, boar taint compounds and performance in entire male pigs. Reprod. Domest. Anim. 2008, 43, 351–359. [Google Scholar] [CrossRef]
- Weiler, U.; Götz, M.; Schmidt, A.; Otto, M.; Müller, S. Influence of sex and immunocastration on feed intake behavior, skatole and indole concentrations in adipose tissue of pigs. Animal 2013, 7, 300–308. [Google Scholar] [CrossRef]
- Čandek-Potokar, M.; Škrlep, M.; Zamaratskaia, G. Immunocastration as alternative to surgical castration in pigs. In Theriogenology; Payan-Carreira, R., Ed.; IntechOpen: London, UK, 2017; pp. 109–126. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Zhou, M.; Cao, X.; Du, X.; Meng, F.; Bu, G.; Kong, F.; Huang, A.; Zeng, X. Mechanistic insight into the role of immunocastration on eliminating skatole in boars. Theriogenology 2019, 131, 32–40. [Google Scholar] [CrossRef]
- Jaros, P.; Bürgi, E.; Stärk, K.D.C.; Claus, R.; Hennessy, D.; Thun, R. Effect of active immunization against GnRH on androstenone concentration, growth performance and carcass quality in intact male pigs. Livest. Prod. Sci. 2005, 92, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.Y.; Turkstra, J.A.; Meloen, R.H.; Liu, X.Y.; Chen, F.Q.; Schaaper, W.M.M.; Oonk, H.B.; Guo, D.Z.; Van de Wiel, D.F.M. Active immunization against gonadotrophin-releasing hormone in Chinese male pigs: Effects of dose on antibody titer, hormone levels and sexual development. Anim. Reprod. Sci. 2002, 70, 223–233. [Google Scholar] [CrossRef]
- Kress, K.; Millet, S.; Labussière, É.; Weiler, U.; Stefanski, V. Sustainability of Pork Production with Immunocastration in Europe. Sustainability 2019, 11, 3335. [Google Scholar] [CrossRef] [Green Version]
- Claus, R.; Weiler, U.; Herzog, A. Physiological aspects of androstenone and skatole formation in the boar-A review with experimental data. Meat Sci. 1994, 38, 289–305. [Google Scholar] [CrossRef]
- Lundström, K.; Malmfors, B.; Malmfors, G.; Stern, S.; Petersson, H.; Mortensen, A.B.; Rensen, S.E.S. Skatole, androstenone and taint in boars fed two different diets. Livest. Prod. Sci. 1988, 18, 55–67. [Google Scholar] [CrossRef]
- Wesoly, R.; Weiler, U. Nutritional influences on skatole formation and skatole metabolism in the pig. Animals 2012, 2, 221–242. [Google Scholar] [CrossRef] [PubMed]
- Malmfors, B.; Lundström, K.; Andresen, Ø.; Bonneau, M.; Kempster, A.J.; Patterson, R.L. Boars for meat production- Report from the EAAP working group, Spain 1989. Livest. Prod. Sci. 1990, 26, 319–326. [Google Scholar] [CrossRef]
- Nold, R.A.; Romans, J.R.; Costello, W.J.; Henson, J.A.; Libal, G.W. Sensory characteristics and carcass traits of boars, barrows, and gilts fed high- or adequate-protein diets and slaughtered at 100 or 110 kilograms. J. Anim. Sci. 1997, 75, 2641–2651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neupert, B.; Claus, R.; Herbert, E.; Weiler, U. Influence of sex, energy, supply and light on fattening traits and carcass composition and their relation to androstenone and skatole concentrations in adipose-tissue of pigs. Züchtungskunde 1995, 67, 317–331. [Google Scholar]
- Claus, R.; Raab, S. Influences on Skatole Formation from Tryptophan in the Pig Colon. In Tryptophan, Serotonin and Melatonin. Basic Aspects and Applications. Advances in Experimental Medicine and Biology; Huether, G., Kochen, W., Simat, T.J., Steinhart, H., Eds.; Springer: Boston, MA, USA, 1999; Volume 467, pp. 679–684. [Google Scholar]
- Wesoly, R. Exogenous Influences on Skatole Formation in the Pig. PhD. Thesis, University of Hohenheim, Stuttgart, Germany, 16 July 2015. [Google Scholar]
- Chuzel, F.; Clark, A.M.; Avallet, O.; Saez, J.M. Transcriptional regulation of the lutropin/human choriogonadotropin receptor and three enzymes of steroidogenesis by growth factors in cultured pig Leydig cells. Eur. J. Biochem. 1996, 239, 8–16. [Google Scholar] [CrossRef]
- Zeng, X.Y.; Turkstra, J.A.; Jongbloed, A.W.; Van Diepen, J.T.M.; Meloen, R.H.; Oonk, H.B.; Guo, D.Z.; Van De Wiel, D.F.M. Performance and hormone levels of immunocastrated, surgically castrated and intact male pigs fed ad libitum high- and low-energy diets. Livest. Prod. Sci. 2002, 77, 1–11. [Google Scholar] [CrossRef]


{kind=link}
| Ingredient | Grower Diet (78 to 106 kg Body Weight) | Finisher Diet (106 to 136 kg Body Weight) | ||||
|---|---|---|---|---|---|---|
| Control | High Energy | Low CP and AA | Control | High Energy | Low CP and AA | |
| Corn | 35.0 | 33.9 | 35.0 | 35.0 | 32.5 | 35.0 |
| Wheat | 18.0 | 18.0 | 18.4 | 17.0 | 18.0 | 18.1 |
| Barley | 17.6 | 15.0 | 21.0 | 21.1 | 21.8 | 25.0 |
| Oat | 9.00 | 8.72 | 11.0 | 11.0 | 8.00 | 12.0 |
| Soybean meal 47% CP | 17.8 | 18.7 | 11.9 | 13.6 | 14.4 | 7.69 |
| Palm oil | 0.53 | 3.65 | 0.34 | 0.36 | 3.36 | 0.08 |
| Calcium carbonate | 0.79 | 0.78 | 0.80 | 0.85 | 0.85 | 0.86 |
| Sodium chloride | 0.45 | 0.45 | 0.45 | 0.45 | 0.45 | 0.45 |
| Monocalcium phospate | 0.26 | 0.27 | 0.31 | 0.13 | 0.13 | 0.18 |
| L-Lysine 50% | 0.23 | 0.21 | 0.30 | 0.14 | 0.12 | 0.23 |
| L-Threonine | 0.02 | 0.02 | 0.02 | - | - | 0.01 |
| DL-Methionine | 0.02 | 0.02 | 0.01 | - | - | - |
| L-Tryptophan | - | - | - | 0.01 | 0.01 | 0.01 |
| Premix 1 | 0.40 | 0.40 | 0.40 | 0.40 | 0.40 | 0.40 |
| Nutrient | Grower Diet (78 to 106 kg Body Weight) | Finisher Diet (106 to 136 kg Body Weight) | ||||
|---|---|---|---|---|---|---|
| Control | High Energy | Low CP and AA | Control | High Energy | Low CP and AA | |
| Net energy, Mcal/kg | 2.33 | 2.48 | 2.33 | 2.33 | 2.48 | 2.33 |
| CP | 16.0 | 16.0 | 14.0 | 14.5 | 14.5 | 12.5 |
| Digestible AA | ||||||
| Lysine | 0.77 | 0.77 | 0.67 | 0.63 | 0.63 | 0.54 |
| Methionine | 0.24 | 0.24 | 0.21 | 0.21 | 0.20 | 0.18 |
| Methionine + Cysteine | 0.49 | 0.49 | 0.44 | 0.44 | 0.43 | 0.39 |
| Threonine | 0.50 | 0.50 | 0.43 | 0.43 | 0.43 | 0.36 |
| Tryptophan | 0.16 | 0.16 | 0.14 | 0.15 | 0.15 | 0.13 |
| Nutrient | Grower Diet (78 to 106 kg Body Weight) | Finisher Diet (106 to 136 kg Body Weight) | ||||
|---|---|---|---|---|---|---|
| Control | High Energy | Low CP and AA | Control | High Energy | Low CP and AA | |
| Gross energy, Mcal/kg | 3.99 | 4.12 | 3.92 | 3.91 | 4.12 | 3.95 |
| Dry matter | 88.7 | 88.2 | 88.0 | 88.0 | 89.4 | 88.1 |
| Ash | 4.18 | 4.19 | 4.17 | 3.85 | 3.98 | 3.65 |
| Starch | 42.1 | 40.3 | 44.0 | 44.5 | 47.8 | 49.0 |
| Ether extract | 3.55 | 5.88 | 3.44 | 3.00 | 5.65 | 3.73 |
| Neutral detergent fiber | 10.9 | 10.2 | 10.5 | 10.5 | 8.96 | 10.2 |
| CP | 16.2 | 15.9 | 14.4 | 14.5 | 15.1 | 12.7 |
| Total AA | ||||||
| Lysine | 0.98 | 0.98 | 0.79 | 0.76 | 0.77 | 0.71 |
| Methionine | 0.28 | 0.27 | 0.25 | 0.24 | 0.25 | 0.23 |
| Threonine | 0.62 | 0.60 | 0.59 | 0.56 | 0.58 | 0.51 |
| FA, % of total FA | ||||||
| C12:0 | 3.45 | 1.66 | 3.35 | 3.26 | 1.72 | 2.50 |
| C14:0 | 0.58 | 0.65 | 0.41 | 0.34 | 0.84 | 0.83 |
| C16:0 | 23.3 | 30.2 | 25.5 | 25.0 | 34.6 | 27.9 |
| C16:1n-7 | 0.18 | 0.19 | 0.20 | 0.18 | 0.17 | 0.20 |
| C16:1n-9 | 0.05 | 0.05 | 0.06 | 0.06 | 0.04 | 0.06 |
| C18:0 | 3.20 | 3.61 | 3.18 | 3.18 | 4.05 | 3.49 |
| C18:1n-7 | 0.70 | 0.61 | 0.73 | 0.71 | 0.62 | 0.65 |
| C18:1n-9 | 30.9 | 33.6 | 31.1 | 31.6 | 34.2 | 32.9 |
| C18:2n-6 | 34.9 | 27.4 | 32.8 | 32.6 | 21.6 | 28.8 |
| C18:3n-3 | 1.89 | 1.46 | 1.66 | 1.62 | 1.05 | 1.29 |
| C18:4n-3 | 0.24 | 0.12 | 0.28 | 0.43 | 0.32 | 0.44 |
| C20:0 | 0.16 | 0.07 | 0.18 | 0.38 | 0.34 | 0.40 |
| C20:1n-9 | 0.46 | 0.32 | 0.50 | 0.61 | 0.40 | 0.57 |
| Trait | Type of Gilt | SEM 1 (n = 45) | Diet 2 | SEM 1 (n = 30) | p-Value 3 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Entire | Immunocastrated | Control | High Energy | Low CP and AA | Gilt | Diet | |||
| Color traits 4 | |||||||||
| Lightness, L* | 35.0 | 35.0 | 0.69 | 35.2 | 34.8 | 35.0 | 0.85 | 0.982 | 0.936 |
| Redness, a* | 2.58 | 3.13 | 0.279 | 2.83 | 2.70 | 3.02 | 0.342 | 0.168 | 0.793 |
| Yellowness, b* | 14.8 | 15.1 | 0.27 | 15.1 | 14.8 | 15.0 | 0.32 | 0.389 | 0.784 |
| Chroma, | 15.1 | 15.5 | 0.27 | 15.4 | 15.1 | 15.4 | 0.33 | 0.281 | 0.722 |
| Hue angle, hab | 80.2 | 78.4 | 1.06 | 79.5 | 79.7 | 78.7 | 1.30 | 0.216 | 0.860 |
| Maximum stress 4, N/cm2 | 40.9 | 41.2 | 1.61 | 40.4 | 42.8 | 40.0 | 1.97 | 0.887 | 0.566 |
| Chemical composition 5, % | |||||||||
| Moisture | 72.2 | 71.6 | 0.19 | 72.0 | 72.0 | 71.8 | 0.24 | 0.034 | 0.798 |
| Protein | 23.6 | 23.4 | 0.11 | 23.3 | 23.5 | 23.7 | 0.14 | 0.204 | 0.145 |
| Intramuscular fat 6 | 2.70 | 3.57 | 0.239 | 3.29 | 3.18 | 2.93 | 0.293 | 0.018 | 0.494 |
| Trait | Type of Gilt | SEM 1 (n = 24) | Diet 2 | SEM 1 (n = 16) | p-Value 3 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Entire | Immunocastrated | Control | High Energy | Low CP and AA | Gilt | Diet | |||
| C14:0 | 1.13 | 1.16 | 0.023 | 1.17 | 1.10 | 1.16 | 0.028 | 0.327 | 0.154 |
| C14:1 4 | 0.013 | 0.012 | 0.0007 | 0.012 | 0.012 | 0.013 | 0.0009 | 0.434 | 0.738 |
| C15:0 | 0.054 | 0.048 | 0.0023 | 0.050 | 0.055 | 0.048 | 0.0028 | 0.062 | 0.240 |
| C15:1 4 | 0.012 | 0.010 | 0.0008 | 0.011 | 0.013 | 0.010 | 0.0010 | 0.039 | 0.223 |
| C16:0 | 22.0 | 22.6 | 0.22 | 22.2 | 22.2 | 22.6 | 0.27 | 0.051 | 0.478 |
| C16:1n-7 | 1.57 | 1.55 | 0.042 | 1.53 ab | 1.47 b | 1.67 a | 0.051 | 0.654 | 0.026 |
| C16:1n-9 | 0.337 | 0.294 | 0.0113 | 0.299 | 0.342 | 0.306 | 0.0138 | 0.011 | 0.067 |
| C17:0 | 0.307 | 0.284 | 0.0142 | 0.301 | 0.295 | 0.290 | 0.0174 | 0.259 | 0.911 |
| C17:1 | 0.271 | 0.243 | 0.0118 | 0.270 | 0.244 | 0.255 | 0.0145 | 0.105 | 0.449 |
| C18:0 | 11.8 | 12.7 | 0.19 | 12.3 | 11.9 | 12.5 | 0.23 | 0.001 | 0.160 |
| C18:1n-7 | 1.73 | 1.65 | 0.030 | 1.75 a | 1.60 b | 1.73 a | 0.036 | 0.063 | 0.013 |
| C18:1n-9 | 42.8 | 42.8 | 0.25 | 42.8 | 43.3 | 42.3 | 0.31 | 0.968 | 0.092 |
| C18:2n-6 | 15.6 | 14.3 | 0.34 | 14.9 | 15.2 | 14.8 | 0.41 | 0.007 | 0.777 |
| C18:3n-3 | 0.728 | 0.670 | 0.0165 | 0.714 | 0.701 | 0.683 | 0.0202 | 0.017 | 0.556 |
| C18:3n-6 5 | 0.040 ± 0.017 | 0.031 ± 0.008 | - | 0.032 ± 0.006 | 0.040 ± 0.022 | 0.036 ± 0.009 | - | 0.015 | 0.236 |
| C18:4n-3 | 0.058 | 0.049 | 0.0029 | 0.053 | 0.053 | 0.055 | 0.0036 | 0.024 | 0.873 |
| C20:0 5 | 0.248 ± 0.038 | 0.261 ± 0.031 | - | 0.267 ± 0.025 | 0.246 ± 0.047 | 0.251 ± 0.026 | - | 0.138 | 0.086 |
| C20:1n-9 | 0.857 | 0.913 | 0.0202 | 0.956 a | 0.852 b | 0.848 b | 0.0247 | 0.057 | 0.005 |
| C20:3n-6 4 | 0.113 | 0.099 | 0.0049 | 0.110 | 0.107 | 0.101 | 0.0060 | 0.054 | 0.566 |
| C20:4n-6 | 0.271 | 0.238 | 0.0086 | 0.258 | 0.253 | 0.252 | 0.0105 | 0.010 | 0.899 |
| C20:5n-3 4 | 0.033 | 0.031 | 0.0033 | 0.027 | 0.034 | 0.035 | 0.0041 | 0.549 | 0.273 |
| Total SFA | 35.5 | 37.1 | 0.36 | 36.3 | 35.8 | 36.8 | 0.44 | 0.003 | 0.233 |
| Total MUFA | 47.6 | 47.5 | 0.25 | 47.6 | 47.8 | 47.2 | 0.31 | 0.771 | 0.303 |
| Total PUFA | 16.9 | 15.4 | 0.36 | 16.1 | 16.4 | 16.0 | 0.44 | 0.006 | 0.807 |
| PUFA/SFA | 0.477 | 0.418 | 0.0136 | 0.445 | 0.462 | 0.437 | 0.0166 | 0.004 | 0.533 |
| Total n-3 | 0.820 | 0.749 | 0.0173 | 0.793 | 0.787 | 0.773 | 0.0212 | 0.007 | 0.784 |
| Total n-6 | 16.1 | 14.7 | 0.35 | 15.3 | 15.6 | 15.2 | 0.43 | 0.006 | 0.804 |
| n-6/n-3 | 19.6 | 19.6 | 0.20 | 19.3 | 19.8 | 19.8 | 0.24 | 0.905 | 0.275 |
| Trait | Type of Castration | SEM 1 (n = 45) | Diet 2 | SEM 1 (n = 30) | p-Value 3 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Surgical | Immunological | Control | High Energy | Low CP and AA | Castration | Diet | |||
| Thawing losses 4, 5, % | 10.09 | 9.47 | 0.389 | 9.30 | 9.90 | 10.14 | 0.476 | 0.187 | 0.459 |
| Cooking losses 4, 5, % | 23.7 | 24.0 | 0.39 | 23.2 b | 23.2 b | 25.2 a | 0.48 | 0.491 | 0.002 |
| Color traits 5 | |||||||||
| Lightness, L* | 34.8 | 32.2 | 0.82 | 33.6 | 33.4 | 33.5 | 1.01 | 0.027 | 0.990 |
| Redness, a* | 3.83 | 4.37 | 0.299 | 3.89 | 3.97 | 4.44 | 0.367 | 0.207 | 0.505 |
| Yellowness, b* | 14.8 | 14.2 | 0.33 | 14.5 | 14.3 | 14.6 | 0.40 | 0.182 | 0.817 |
| Chroma, | 15.4 | 14.9 | 0.31 | 15.1 | 15.0 | 15.4 | 0.38 | 0.302 | 0.775 |
| Hue angle 4, hab | 75.5 | 72.5 | 1.22 | 74.9 | 73.9 | 73.2 | 1.49 | 0.084 | 0.695 |
| Maximum stress 4, 5, N/cm2 | 40.4 | 37.6 | 1.91 | 38.0 | 41.3 | 37.7 | 2.33 | 0.233 | 0.332 |
| Chemical composition 6, % | |||||||||
| Moisture | 71.4 | 72.2 | 0.17 | 71.7 | 71.6 | 72.1 | 0.21 | 0.004 | 0.246 |
| Protein | 23.0 | 23.1 | 0.08 | 23.0 | 23.0 | 23.1 | 0.10 | 0.856 | 0.876 |
| Intramuscular fat | 4.44 | 3.40 | 0.220 | 4.02 | 4.15 | 3.58 | 0.269 | 0.001 | 0.293 |
| Diet 1 | Control | High Energy | Low CP and AA | SEM 2 (n = 8) | p-Value | |||
|---|---|---|---|---|---|---|---|---|
| Type of Castration | Surgical | Immunological | Surgical | Immunological | Surgical | Immunological | ||
| C16:0 | 23.3 abc | 22.6 c | 22.8 bc | 23.8 a | 22.7 bc | 23.5 ab | 0.31 | 0.019 |
| C16:1n-9 | 0.259 b | 0.318 a | 0.290 ab | 0.277 ab | 0.296 ab | 0.268 b | 0.0150 | 0.012 |
| C18:2n-6 | 13.8 bc | 15.7 a | 14.1 bc | 13.8 bc | 15.1 ab | 13.4 c | 0.47 | 0.002 |
| C18:3n-3 | 0.660 bc | 0.754 a | 0.651 bc | 0.632 bc | 0.687 b | 0.617 c | 0.0232 | 0.003 |
| C18:3n-6 | 0.025 c | 0.033 a | 0.032 ab | 0.030 abc | 0.028 abc | 0.026 bc | 0.0022 | 0.046 |
| C18:4n-3 | 0.040 bc | 0.045 ab | 0.052 a | 0.034 c | 0.039 bc | 0.037 bc | 0.0039 | 0.010 |
| C20:1n-9 | 0.967 a | 0.881 bc | 0.927 abc | 0.875 c | 0.874 c | 0.962 ab | 0.0351 | 0.042 |
| C20:4n-6 | 0.211 b | 0.263 a | 0.230 ab | 0.226 b | 0.233 ab | 0.218 b | 0.0123 | 0.021 |
| Total SFA | 38.0 ab | 37.0 b | 37.1 b | 38.6 ab | 37.2 b | 39.0 a | 0.60 | 0.039 |
| Total PUFA | 14.9 bc | 16.9 a | 15.2 bc | 14.8 bc | 16.2 ab | 14.4 c | 0.50 | 0.002 |
| PUFA/SFA | 0.392 bc | 0.459 a | 0.411 abc | 0.384 c | 0.436 ab | 0.371 c | 0.0181 | 0.002 |
| Total n-3 | 0.724 b | 0.829 a | 0.728 b | 0.691 b | 0.751 b | 0.682 b | 0.0259 | 0.004 |
| Total n-6 | 14.1 bc | 16.1 a | 14.5 bc | 14.1 bc | 15.4 ab | 13.7 c | 0.48 | 0.001 |
| Type of Castration | SEM 1 (n = 24) | Diet 2 | SEM 1 (n = 16) | p-Value 3 | |||||
|---|---|---|---|---|---|---|---|---|---|
| Surgical | Immunological | Control | High Energy | Low CP and AA | Castration | Diet | |||
| C14:0 | 1.22 | 1.20 | 0.022 | 1.22 | 1.18 | 1.23 | 0.027 | 0.664 | 0.488 |
| C14:1 | 0.012 | 0.012 | 0.0006 | 0.012 | 0.012 | 0.011 | 0.0007 | 0.786 | 0.546 |
| C15:0 4 | 0.047 | 0.051 | 0.0021 | 0.055 a | 0.050 a | 0.041 b | 0.0025 | 0.195 | 0.0005 |
| C15:1 4 | 0.008 | 0.008 | 0.0003 | 0.009 | 0.008 | 0.007 | 0.0004 | 0.284 | 0.105 |
| C16:1n-7 4 | 1.59 | 1.61 | 0.045 | 1.66 | 1.54 | 1.60 | 0.055 | 0.648 | 0.266 |
| C17:0 | 0.282 | 0.313 | 0.0118 | 0.342 a | 0.284 b | 0.267 b | 0.0145 | 0.064 | 0.002 |
| C17:1 | 0.250 | 0.264 | 0.0097 | 0.308 a | 0.233 b | 0.231 b | 0.0118 | 0.329 | <0.0001 |
| C18:0 | 12.7 | 13.1 | 0.24 | 12.7 | 12.8 | 13.2 | 0.29 | 0.235 | 0.418 |
| C18:1n-7 4 | 1.70 | 1.67 | 0.034 | 1.76 a | 1.58 b | 1.73 a | 0.042 | 0.478 | 0.009 |
| C18:1n-9 | 42.4 | 41.7 | 0.24 | 41.6 | 42.6 | 41.9 | 0.29 | 0.028 | 0.066 |
| C20:0 | 0.263 | 0.252 | 0.0087 | 0.254 | 0.263 | 0.256 | 0.0106 | 0.407 | 0.809 |
| C20:3n-6 | 0.096 | 0.097 | 0.0031 | 0.102 | 0.093 | 0.095 | 0.0039 | 0.768 | 0.235 |
| C20:5n-3 | 0.025 | 0.027 | 0.0014 | 0.027 | 0.025 | 0.027 | 0.0018 | 0.219 | 0.688 |
| Total MUFA | 47.2 | 46.4 | 0.26 | 46.6 | 47.2 | 46.7 | 0.32 | 0.041 | 0.403 |
| n-6/n-3 | 20.0 | 20.0 | 0.17 | 19.5 b | 20.2 a | 20.3 a | 0.21 | 0.920 | 0.024 |
| Type of Castration | Diet 1 | p-Value 2 | |||||
|---|---|---|---|---|---|---|---|
| Surgical | Immunological | Control | High Energy | Low CP and AA | Castration | Diet | |
| n | 18 | 18 | 12 | 12 | 12 | ||
| Androstenone 3 | bd | bd | bd | bd | bd | - | - |
| Skatole 4 | 0.028 ± 0.014 | 0.031 ± 0.018 | 0.037 ± 0.014 | 0.027 ± 0.019 | 0.025 ± 0.012 | 0.593 | 0.059 |
| Indole 4 | 0.021 ± 0.011 | 0.027 ± 0.018 | 0.031 ± 0.019 | 0.018 ± 0.008 | 0.022 ± 0.013 | 0.267 | 0.149 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Ciria, L.; Miana-Mena, F.J.; López-Mendoza, M.C.; Álvarez-Rodríguez, J.; Latorre, M.A. Influence of Immunocastration and Diet on Meat and Fat Quality of Heavy Female and Male Pigs. Animals 2021, 11, 3355. https://doi.org/10.3390/ani11123355
Pérez-Ciria L, Miana-Mena FJ, López-Mendoza MC, Álvarez-Rodríguez J, Latorre MA. Influence of Immunocastration and Diet on Meat and Fat Quality of Heavy Female and Male Pigs. Animals. 2021; 11(12):3355. https://doi.org/10.3390/ani11123355
Chicago/Turabian StylePérez-Ciria, Leticia, Francisco Javier Miana-Mena, María Carmen López-Mendoza, Javier Álvarez-Rodríguez, and Maria Angeles Latorre. 2021. "Influence of Immunocastration and Diet on Meat and Fat Quality of Heavy Female and Male Pigs" Animals 11, no. 12: 3355. https://doi.org/10.3390/ani11123355