
Evidence for Unknown Sarcocystis-Like Infection in Stranded Striped Dolphins (Stenella coeruleoalba) from the Ligurian Sea, Italy

 ,
,  , , , , , , and
, , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Naturally Infected Dolphins
2.2. Histology and Immunohistochemistry
2.3. PCR and Sequence Analysis
2.4. Serological, Toxicological, and Microbiological Analyses
2.5. Light Microscopy Re-Examination for Parasite Characterization: Histology and Immunohistochemistry
2.6. Electron Microscopy Examination
2.7. Molecular Analyses: Parasite Detection, Identification, and Characterization
3. Results
3.1. Naturally Infected Dolphins
3.2. Histology and Immunohistochemistry
3.3. PCR and Sequence Analysis
3.4. Serological, Toxicological, and Microbiological Analyses
3.5. Light Microscopy Re-Examination: Histology and Immunohistochemistry
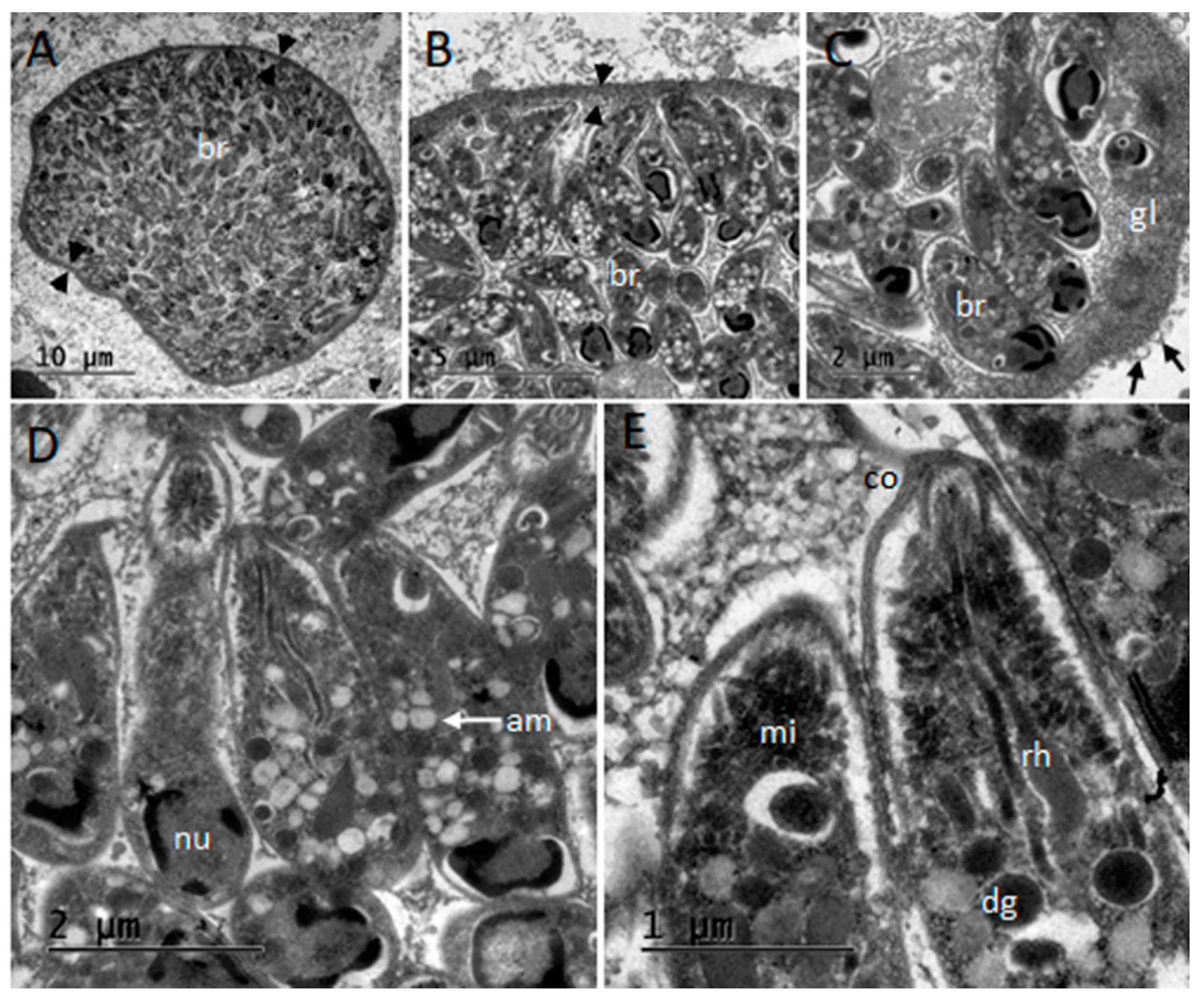
3.6. Ultrastructural Description of Tissue Cysts
3.7. Molecular Detection and Parasite Identification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, M.A.; Shapiro, K.; Murray, M.J.; Haulena, M.; Raverty, S. Protozoan Parasites of Marine Mammals. In CRC Handbook of Marine Mammals; Gulland, F.M.D., Dierauf, L.A., Whitman, K.L., Eds.; CRC Press: New York, NY, USA, 2018; pp. 425–469. ISBN 9781498796873. [Google Scholar]
- Gibson, A.K.; Raverty, S.; Lambourn, D.M.; Huggins, J.; Magargal, S.L.; Grigg, M.E. Polyparasitism is associated with increased disease severity in Toxoplasma gondii-infected marine sentinel species. PLoS Negl. Trop. Dis. 2011, 5, 14–16. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, K.; Miller, M.A.; Mazet, J. Temporal Association Between Land-Based Runoff Events and California Sea Otter (Enhydra Lutris Nereis) Protozoal Mortalities. J. Wildl. Dis. 2012, 48, 394–404. [Google Scholar] [CrossRef]
- Van Bressem, M.; Raga, A.; Di Guardo, G.; Jepson, P.; Duignan, P.; Siebert, U.; Barrett, T.; Santos, M.; Moreno, I.; Siciliano, S.; et al. Emerging infectious diseases in cetaceans worldwide and the possible role of environmental stressors. Dis. Aquat. Organ. 2009, 86, 143–157. [Google Scholar] [CrossRef]
- Barbosa, L.; Johnson, C.K.; Lambourn, D.M.; Gibson, A.K.; Haman, K.H.; Huggins, J.L.; Sweeny, A.R.; Sundar, N.; Raverty, S.A.; Grigg, M.E. A novel Sarcocystis neurona genotype XIII is associated with severe encephalitis in an unexpectedly broad range of marine mammals from the northeastern Pacific Ocean. Int. J. Parasitol. 2015, 45, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Di Guardo, G.; Proietto, U.; Di Francesco, C.E.; Marsilio, F.; Zaccaroni, A.; Scaravelli, D.; Mignone, W.; Garibaldi, F.; Kennedy, S.; Forster, F.; et al. Cerebral Toxoplasmosis in Striped Dolphins (Stenella coeruleoalba) Stranded Along the Ligurian Sea Coast of Italy. Vet. Pathol. 2010, 47, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Pintore, M.D.; Mignone, W.; Di Guardo, G.; Mazzariol, S.; Ballardini, M.; Florio, C.L.; Goria, M.; Romano, A.; Caracappa, S.; Giorda, F.; et al. Neuropathologic findings in cetaceans stranded in Italy (2002–2014). J. Wildl. Dis. 2018, 54. [Google Scholar] [CrossRef] [PubMed]
- Resendes, A.R.; Juan-Sallés, C.; Almeria, S.; Majó, N.; Domingo, M.; Dubey, J.P. Hepatic sarcocystosis in a striped dolphin (Stenella coeruleoalba) from the Spanish Mediterranean coast. J. Parasitol. 2002, 88, 206–209. [Google Scholar] [CrossRef]
- Geraci, J.R.; Lounsbury, V.J. Marine Mammals Ashore: A Field Guide for Strandings, 2nd ed.; National Aquarium: Baltimore, MD, USA, 2005; Volume 486, ISBN 0977460908. [Google Scholar]
- Garofolo, G.; Petrella, A.; Lucifora, G.; Di Francesco, G.; Di Guardo, G.; Pautasso, A.; Iulini, B.; Varello, K.; Giorda, F.; Goria, M.; et al. Occurrence of Brucella ceti in striped dolphins from Italian Seas. PLoS ONE 2020, 15, e0240178. [Google Scholar] [CrossRef]
- Giorda, F.; Ballardini, M.; Di Guardo, G.; Pintore, M.D.; Grattarola, C.; Iulini, B.; Mignone, W.; Goria, M.; Serracca, L.; Varello, K.; et al. Postmortem findings in cetaceans found stranded in the Pelagos sanctuary, Italy, 2007–2014. J. Wildl. Dis. 2017, 53, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Verna, F.; Giorda, F.; Miceli, I.; Rizzo, G.; Pautasso, A.; Romano, A.; Iulini, B.; Pintore, M.D.; Mignone, W.; Grattarola, C.; et al. Detection of morbillivirus infection by RT-PCR RFLP analysis in cetaceans and carnivores. J. Virol. Methods 2017, 247, 22–27. [Google Scholar] [CrossRef]
- VanDevanter, D.R.; Warrener, P.; Bennett, L.; Schultz, E.R.; Coulter, S.; Garber, R.L.; Rose, T.M. Detection and analysis of diverse herpesviral species by consensus primer PCR. J. Clin. Microbiol. 1996, 34, 1666–1671. [Google Scholar] [CrossRef] [Green Version]
- Vitale, M.; Galluzzo, P.; Currò, V.; Gozdzik, K.; Schillaci, D.; Di Marco Lo Presti, V. A high sensitive nested PCR for Toxoplasma gondii detection in animal and food samples. J. Microb. Biochem. Technol. 2013, 05, 39–41. [Google Scholar] [CrossRef] [Green Version]
- Baily, G.G.; Krahn, J.B.; Drasar, B.S.; Stoker, N.G. Detection of Brucella melitensis and Brucella abortus by DNA amplification. J. Trop. Med. Hyg. 1992, 95, 271–275. [Google Scholar]
- Hernández-Mora, G.; González-Barrientos, R.; Morales, J.A.; Chaves-Olarte, E.; Guzmán-Verri, C.; Baquero-Calvo, E.; De-Miguel, M.J.; Marín, C.M.; Blasco, J.M.; Moreno, E. Neurobrucellosis in stranded dolphins, Costa Rica. Emerg. Infect. Dis. 2008, 14, 1430–1433. [Google Scholar] [CrossRef]
- Marsili, L.; Focardi, S. Chlorinated hydrocarbon (HCB, DDTs and PCBs) levels in cetaceans stranded along the Italian coasts: An overview. Environ. Monit. Assess. 1997, 45, 129–180. [Google Scholar] [CrossRef]
- Marsili, L.; D’Agostino, A.; Bucalossi, D.; Malatesta, T.; Fossi, M.C. Theoretical models to evaluate hazard due to organochlorine compounds (OCs) in Mediterranean striped dolphin (Stenella coeruleoalba). Chemosphere 2004, 56, 791–801. [Google Scholar] [CrossRef]
- World Organisation for Animal Health (OIE). Manual of Diagnostic Tests and Vaccines for Terrestrial Animals (Terrestrial Manual), 8th ed.; World Organisation for Animal Health (OIE): Paris, France, 2018; ISBN 978-92-95108-18-9. [Google Scholar]
- Dubey, J.P.; Garner, M.M.; Stetter, M.D.; Marsh, A.E.; Barr, B.C. Acute Sarcocystis Falcatula-Like Infection in a Carmine Bee-Eater (Merops Nubicus) and Immunohistochemical Cross Reactivity Between Sarcocystis Falcatula and Sarcocystis Neurona. J. Parasitol. 2001, 87, 824–832. [Google Scholar] [CrossRef]
- Marsh, A.; Hyun, C.; Barr, B.C.; Tindall, R.; Lakritz, J. Characterization of monoclonal antibodies developed against Sarcocystis neurona. Parasitol. Res. 2002, 88, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Marsh, A.E.; Chaney, S.B.; Howe, D.K.; Saville, W.J.; Reed, S.M. Small sarcocysts can be a feature of experimental infections with Sarcocystis neurona merozoites. Vet. Parasitol. 2017, 245, 116–118. [Google Scholar] [CrossRef] [PubMed]
- Chaney, S.B.; Marsh, A.E.; Lewis, S.; Carman, M.; Howe, D.K.; Saville, W.J.; Reed, S.M. Sarcocystis neurona manipulation using culture-derived merozoites for bradyzoite and sporocyst production. Vet. Parasitol. 2017, 238, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Calero-Bernal, R.; Rosenthal, B.M.; Speer, C.A.; Fayer, R. Unique Multiple-Host Sarcocystis Species. In Sarcocystosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 125–168. [Google Scholar]
- Hall, T.A. BIOEDIT: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/ NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Su, C.; Shwab, E.K.; Zhou, P.; Zhu, X.Q.; Dubey, J.P. Moving towards an integrated approach to molecular detection and identification of Toxoplasma gondii. Parasitology 2010, 137, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pena, H.F.J.; Gennari, S.M.; Dubey, J.P.; Su, C. Population structure and mouse-virulence of Toxoplasma gondii in Brazil. Int. J. Parasitol. 2008, 38, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution (N. Y.) 1985, 39, 783. [Google Scholar] [CrossRef]
- Masatoshi, N.; Sudhir, K. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK, 2000; ISBN 0 19 513584 9. [Google Scholar]
- Sierra, E.; Sánchez, S.; Saliki, J.T.; Blas-Machado, U.; Arbelo, M.; Zucca, D.; Fernández, A. Retrospective study of etiologic agents associated with nonsuppurative meningoencephalitis in stranded cetaceans in the canary Islands. J. Clin. Microbiol. 2014, 52, 2390–2397. [Google Scholar] [CrossRef] [Green Version]
- Van Bressem, M.-F.; Duignan, P.; Banyard, A.; Barbieri, M.; Colegrove, K.; De Guise, S.; Di Guardo, G.; Dobson, A.; Domingo, M.; Fauquier, D.; et al. Cetacean Morbillivirus: Current Knowledge and Future Directions. Viruses 2014, 6, 5145–5181. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Calero-Bernal, R.; Rosenthal, B.M.; Speer, C.A.; Fayer, R. Sarcocystosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2015; ISBN 9780429183188. [Google Scholar]
- Sundermann, C.A.; Estridge, B.H.; Branton, M.S.; Bridgman, C.R.; Lindsay, D.S. Immunohistochemical diagnosis of Toxoplasma gondii: Potential for cross- reactivity with Neospora caninum. J. Parasitol. 1997, 83, 440–443. [Google Scholar] [CrossRef]
- Miller, M.A.; Barr, B.C.; Nordhausen, R.; James, E.R.; Magargal, S.L.; Murray, M.; Conrad, P.A.; Toy-Choutka, S.; Jessup, D.A.; Grigg, M.E. Ultrastructural and molecular confirmation of the development of Sarcocystis neurona tissue cysts in the central nervous system of southern sea otters (Enhydra lutris nereis). Int. J. Parasitol. 2009, 39, 1363–1372. [Google Scholar] [CrossRef] [Green Version]
- Dubey, J.P.; Calero-Bernal, R.; Rosenthal, B.M.; Speer, C.A.; Fayer, R. Techniques. In Sarcocystosis of Animals and Humans; CRC Press: Boca Raton, FL, USA, 2015; pp. 121–122. [Google Scholar]
- Gjerde, B.; Hilali, M.; Abbas, I.E. Molecular differentiation of Sarcocystis buffalonis and Sarcocystis levinei in water buffaloes (Bubalus bubalis) from Sarcocystis hirsuta and Sarcocystis cruzi in cattle (Bos taurus). Parasitol. Res. 2016, 115, 2459–2471. [Google Scholar] [CrossRef]
- Gjerde, B. Phylogenetic relationships among Sarcocystis species in cervids, cattle and sheep inferred from the mitochondrial cytochrome c oxidase subunit I gene. Int. J. Parasitol. 2013, 43, 579–591. [Google Scholar] [CrossRef]
- Hassanin, A.; Delsuc, F.; Ropiquet, A.; Hammer, C.; Jansen Van Vuuren, B.; Matthee, C.; Ruiz-Garcia, M.; Catzeflis, F.; Areskoug, V.; Nguyen, T.T.; et al. Pattern and timing of diversification of Cetartiodactyla (Mammalia, Laurasiatheria), as revealed by a comprehensive analysis of mitochondrial genomes. Comptes Rendus Biol. 2012, 335, 32–50. [Google Scholar] [CrossRef]
- Seilacher, A.; Reif, W.-E.; Wenk, P. The parasite connection in ecosystems and macroevolution. Naturwissenschaften 2007, 94, 155–169. [Google Scholar] [CrossRef]
- Amouei, A.; Sarvi, S.; Sharif, M.; Aghayan, S.A.; Javidnia, J.; Mizani, A.; Moosazadeh, M.; Shams, N.; Hosseini, S.A.; Hosseininejad, Z.; et al. A systematic review of Toxoplasma gondii genotypes and feline: Geographical distribution trends. Transbound. Emerg. Dis. 2020, 67, 46–64. [Google Scholar] [CrossRef]
- Fernández-Escobar, M.; Calero-Bernal, R.; Benavides, J.; Regidor-Cerrillo, J.; Guerrero-Molina, M.C.; Gutiérrez-Expósito, D.; Collantes-Fernández, E.; Ortega-Mora, L.M. Isolation and genetic characterization of Toxoplasma gondii in Spanish sheep flocks. Parasites Vectors 2020, 13, 1–14. [Google Scholar] [CrossRef]
- Di Guardo, G.; Di Cesare, A.; Otranto, D.; Casalone, C.; Iulini, B.; Mignone, W.; Tittarelli, C.; Meloni, S.; Castagna, G.; Forster, F.; et al. Genotyping of Toxoplasma gondii isolates in meningo-encephalitis affected striped dolphins (Stenella coeruleoalba) from Italy. Vet. Parasitol. 2011, 183, 31–36. [Google Scholar] [CrossRef]
- Marcer, F.; Marchiori, E.; Centelleghe, C.; Ajzenberg, D.; Gustinelli, A.; Meroni, V.; Mazzariol, S. Parasitological and pathological findings in fin whales Balaenoptera physalus stranded along Italian coastlines. Dis. Aquat. Organ. 2019, 133, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Terracciano, G.; Fichi, G.; Comentale, A.; Ricci, E.; Mancusi, C.; Perrucci, S. Dolphins stranded along the tuscan coastline (Central Italy) of the “pelagos sanctuary”: A parasitological investigation. Pathogens 2020, 9, 612. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Fair, P.A.; Sundar, N.; Velmurugan, G.; Kwok, O.C.H.H.; McFee, W.E.; Majumdar, D.; Su, C. Isolation of Toxoplasma gondii From Bottlenose Dolphins (Tursiops truncatus). J. Parasitol. 2008, 94, 821–823. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Mergl, J.; Gehring, E.; Sundar, N.; Velmurugan, G.V.; Kwok, O.C.H.H.; Grigg, M.E.; Su, C.; Martineau, D. Toxoplasmosis in captive dolphins (Tursiops truncatus) and walrus (Odobenus rosmarus). J. Parasitol. 2009, 95, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Roe, W.D.; Howe, L.; Baker, E.J.; Burrows, L.; Hunter, S.A. An atypical genotype of Toxoplasma gondii as a cause of mortality in Hector’s dolphins (Cephalorhynchus hectori). Vet. Parasitol. 2013, 192, 67–74. [Google Scholar] [CrossRef]
- Dubey, J.P.; Morales, J.A.; Sundar, N.; Velmurugan, G.V.; González-Barrientos, C.R.; Hernández-Mora, G.; Su, C. Isolation and genetic characterization of Toxoplasma gondii from striped dolphin (Stenella coeruleoalba) from Costa Rica. J. Parasitol. 2007, 93, 710–711. [Google Scholar] [CrossRef]
- Calero-Bernal, R.; Mauroo, N.F.; Hui, S.W.; Kuiken, T.; van de Bildt, M.W.G.; de Jong, A.W.; Osterhaus, A.D.M.E.; Sims, L.; Gendron-Fitzpatrick, A.; Carmena, D.; et al. Acute fatal sarcocystosis hepatitis in an Indo-Pacific bottlenose dolphin (Tursiops aduncus) in Hong Kong. Vet. Parasitol. 2017, 235, 64–68. [Google Scholar] [CrossRef]
- Dubey, J.P.; Zarnke, R.; Thomas, N.J.; Wong, S.K.; Van Bonn, W.; Briggs, M.; Davis, J.W.; Ewing, R.; Mense, M.; Kwok, O.C.H.; et al. Toxoplasma gondii, Neospora caninum, Sarcocystis neurona, and Sarcocystis canis-like infections in marine mammals. Vet. Parasitol. 2003, 116, 275–296. [Google Scholar] [CrossRef]
- De Guise, S.; Lagacé, A.; Girard, C.; Béland, P. Intramuscular Sarcocystis in two beluga whales and an Atlantic white-sided dolphin from the St. Lawrence estuary, Quebec, Canada. J. VET Diagn. Investig. 1993, 5, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Akao, S. A New Species of Sarcocystis Parasitic in the Whale Balaenoptera borealis. J. Protozool. 1970, 17, 290–294. [Google Scholar] [CrossRef]
- Ewing, R.; Zaias, J.; Stamper, M.A.; Bossart, G.D.; Dubey, J.P. Prevalence of Sarcocystis sp. in stranded Atlantic white-sided dolphins (Lagenorhynchus acutus). J. Wildl. Dis. 2002, 38, 291–296. [Google Scholar] [CrossRef]
- Sierra, E.; Espinosa de los Monteros, A.; Fernández, A.; Díaz-Delgado, J.; Suárez-Santana, C.; Arbelo, M.; Sierra, M.A.; Herráez, P. Muscle Pathology in Free-Ranging Stranded Cetaceans. Vet. Pathol. 2017, 54, 298–311. [Google Scholar] [CrossRef]
- Munday, B.L.; Mason, R.W.; Hartley, W.J.; Presidente, P.J.; Obendorf, D. Sarcocystis and related organisms in Australian wildlife: I. Survey findings in mammals. J. Wildl. Dis. 1978, 14, 417–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | YS | DC | NuS | Age/ Sex | Main Lesions (Gross and Microscopic) | Detected Pathogens | Cause of Death | Reference |
|---|---|---|---|---|---|---|---|---|
| SD1 | 2011 | 3 | Poor | Adult M | Severe granulomatous pneumonia; fibrinous peritonitis; splenomegaly associated to chronic granulomatous splenitis; cholangiohepatitis; generalized lymphadenitis associated to lymphoid depletion; severe NS meningoencephalitis | Photobacterium damselae subsp. damselae (isolated from blowhole and lungs); Campula palliata (pancreas and liver); Monorygma grimaldi (musculature and peritoneum); Skrjabinalius guevarai (lung); anti-DMV antibodies (1:16) in blood serum (VN) | Infectious disease (parasitic and unknown agent) | [11] |
| SD2 | 2017 | 2 | Moderate | Juvenile M | Skin ulcers; ulcerative glossitis; subcutaneous parasitic cysts; bronchointerstitial pneumonia; multifocal necrotizing hepatitis; cholangiohepatitis; splenomegaly and generalized lymphadenomegaly associated to multicentric lymphoid necrosis; interstitial nephritis; lymphadenitis; severe NS meningoencephalitis | Phyllobotrium spp (blubber); Monorygma grimaldii (musculature); Brucella ceti (isolation from CNS, lung, and spleen; PCR from brain, liver, lung spleen, and lymph nodes); DMV (PCR, IHC from CNS, spleen, kidney, tonsils, and lymph nodes, bladder and muscle); Toxoplasma gondii (PCR, IHC from CNS, lymph nodes, spleen, liver, and muscle); anti-DMV antibodies (1:8) in blood serum (VN); anti-T. gondii antibodies (>1:640) in blood serum, (1:160) in CSF, (1:80) in aqueous humor (IFAT). Severe immunosuppression (CAN = 0.688) [18] | Infectious disease (viral, bacterial, and parasitic) | [10] |
| Isolate/ Sample | SAG1 | 3′-SAG2 | 5′-SAG2 | Alt. SAG2 | SAG3 | BTUB | GRA6 | c22-8 | C29-2 | L358 | PK1 | Apico | ToxoDB PCR-RFLP Genotype # |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RH (ref. type I) | I | I/III | I/II | I | I | I | I | I | I | I | I | I | #1 |
| Me-49 (ref. type II) | II/III | II | I/II | II | II | II | II | II | II | II | II | II | #1 |
| NED (ref. type III) | II/III | I/III | III | III | III | III | III | III | III | III | III | III | #2 |
| SD2 (muscle) | II/III | II | I/II | II | II | II | II | II | II | II | II | I | #3 |
| SD2 (CNS) | II/III | II | I/II | II | II | II | II | II | II | II | II | I | #3 |
| SD2 (Heart) | II/III | II | I/II | II | II | II | II | II | II | II | II | I | #3 |
| Host | Location | N°. of Individuals | Condition | Genotype (n) | Method (Markers) | Isolate ID | Reference |
|---|---|---|---|---|---|---|---|
| Bottlenose dolphin (Tursiops truncatus) | South Carolina (USA) | 3 | Stranded | #1 (2/3); Unique (1/3) | PCR-RFLP (SAG1, SAG2, SAG3, BTUB, GRA6, c22-8, c29-2, L358, PK1, and Apico) | TgDoUs1-3 | [46] |
| Canada (born in Russia) | 1 | Captivity | #3 (1/1) | PCR-RFLP (B1, SAG1, SAG2, SAG3, BTUB, GRA6, c22-8, c29-2, L358, PK1, and Apico) +PCR-Seq (B1, SAG1) | TgDoCA1 | [47] | |
| Hector’s dolphins (Cephalorhynchus hectori) | New Zealand | 8 | By caught/ stranded | #3 (7/8); Type II variant (Type I + II at L358 and Type I at Apico) | PCR-RFLP (SAG1, SAG2 (5‘ + 3‘), SAG3, GRA6, L358, PK1, and Apico) | No isolation | [48] |
| Striped dolphin (Stenella coeruleoalba) | Costa Rica | 1 | Stranded | #1 (1/1) | PCR-RFLP (SAG1, SAG2, SAG3, BTUB, GRA6, c22-8, c29-2, L358, PK1, and Apico) | TgSdCo1 | [49] |
| Italy | 3 | Stranded | Type II (2/3); Unique (1/3) | PCR-seq (B1, gra6 a nd uprt1) | TSL2, TSL3, and TSL6 | [43] | |
| Italy | 1 | Stranded | #1 (1/1) | RFLP-PCR (SAG1, SAG2 (5‘ + 3‘), alt SAG2, SAG3, BTUB, GRA6) | No isolation | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giorda, F.; Romani-Cremaschi, U.; Marsh, A.E.; Grattarola, C.; Iulini, B.; Pautasso, A.; Varello, K.; Berio, E.; Gazzuola, P.; Marsili, L.; et al. Evidence for Unknown Sarcocystis-Like Infection in Stranded Striped Dolphins (Stenella coeruleoalba) from the Ligurian Sea, Italy. Animals 2021, 11, 1201. https://doi.org/10.3390/ani11051201
Giorda F, Romani-Cremaschi U, Marsh AE, Grattarola C, Iulini B, Pautasso A, Varello K, Berio E, Gazzuola P, Marsili L, et al. Evidence for Unknown Sarcocystis-Like Infection in Stranded Striped Dolphins (Stenella coeruleoalba) from the Ligurian Sea, Italy. Animals. 2021; 11(5):1201. https://doi.org/10.3390/ani11051201
Chicago/Turabian StyleGiorda, Federica, Umberto Romani-Cremaschi, Antoinette E. Marsh, Carla Grattarola, Barbara Iulini, Alessandra Pautasso, Katia Varello, Enrica Berio, Paola Gazzuola, Letizia Marsili, and et al. 2021. "Evidence for Unknown Sarcocystis-Like Infection in Stranded Striped Dolphins (Stenella coeruleoalba) from the Ligurian Sea, Italy" Animals 11, no. 5: 1201. https://doi.org/10.3390/ani11051201