Efficiency of Crude Protein Utilisation in Grazing Dairy Cows: A Case Study Comparing Two Production Systems Differing in Intensification Level in New Zealand

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Description of Production Systems
2.1.1. Low-Intensity Production System
2.1.2. High-Intensity Production System
2.2. Feed Allocation and Chemical Composition Measurements
2.3. Animal Measurements
2.4. Energy Requirements and Intake Estimates
2.5. Definitions of Nitrogen Utilisation Efficiency and Nitrogen Excreta Estimates
2.6. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Zamani, P. Chapter 7: Efficiency of Lactation. In Milk Production—An Up-to-Date Overview of Animal Nutrition, Management and Health; Chaiyabutr, N., Ed.; Intech: Rijeka, Croatia, 2012; pp. 139–150. [Google Scholar]
- Kolver, E.S.; Muller, L.D. Performance and nutrient intake of high producing Holstein cows consuming pasture or a total mixed ration. J. Dairy Sci. 1998, 81, 1403–1411. [Google Scholar] [CrossRef]
- Waghorn, G.C.; Clark, D.A. Feeding value of pastures for ruminants. N. Z. Vet. J. 2004, 52, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Kebreab, E.; France, J.; Beever, D.E.; Castillo, A.R. Nitrogen pollution by dairy cows and its mitigation by dietary manipulation. Nutr. Cycl. Agroecosyst. 2001, 60, 275–285. [Google Scholar] [CrossRef]
- Ledgard, S.F.; Schils, R.; Eriksen, J.; Luo, J. Environmental impacts of grazed clover-grass pastures. Irish J. Agricult. Food Res. 2009, 48, 209–226. [Google Scholar]
- Tamminga, S. Nutrition management of dairy cows as a contribution to pollution control. J. Dairy Sci. 1992, 75, 345–357. [Google Scholar] [CrossRef]
- Mulligan, F.J.; Dillon, P.; Callan, J.J.; Rath, M.; O’Mara, F.P. Supplementary concentrate type affects nitrogen excretion of grazing dairy cows. J. Dairy Sci. 2004, 87, 3451–3460. [Google Scholar] [CrossRef]
- Mackle, T.R.; Bryant, A.M.; Petch, S.F.; Hooper, R.J.; Auldist, M.J. Variation in the composition of milk protein from pasture-fed dairy cows in late lactation and the effect of grain and silage supplementation. N. Z. J. Agric. Res. 1999, 42, 147–154. [Google Scholar] [CrossRef]
- Hristov, A.N.; Ropp, J.K.; Grandeen, K.L.; Abedi, S.; Etter, R.P.; Melgar, A.; Foley, A.E. Effect of carbohydrate source on ammonia utilization in lactating dairy cows. J. Animal Sci. 2005, 83, 408–421. [Google Scholar] [CrossRef] [Green Version]
- Broderick, G.A.; Clayton, M.K. A statistical evaluation of animal and nutritional factors influencing concentrations of milk urea nitrogen. J. Dairy Sci. 1997, 80, 2964–2971. [Google Scholar] [CrossRef]
- Jonker, J.S.; Kohn, R.A.; Erdman, R.A. Using milk urea nitrogen to predict nitrogen excretion and utilization efficiency in lactating dairy cows. J. Dairy Sci. 1998, 81, 2681–2692. [Google Scholar] [CrossRef] [Green Version]
- Nousiainen, J.; Shingfield, K.J.; Huhtanen, P. Evaluation of milk urea nitrogen as a diagnostic of protein feeding. J. Dairy Sci. 2004, 87, 386–398. [Google Scholar] [CrossRef] [Green Version]
- Bargo, F.; Muller, L.D.; Delahoy, J.E.; Cassidy, T.W. Milk response to concentrate supplementation of high producing dairy cows grazing at two pasture allowances. J. Dairy Sci. 2002, 85, 1777–1792. [Google Scholar] [CrossRef] [Green Version]
- Totty, V.K.; Greenwood, S.L.; Bryant, R.H.; Edwards, G.R. Nitrogen partitioning and milk production of dairy cows grazing simple and diverse pastures. J. Dairy Sci. 2013, 96, 141–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gourley, C.J.P.; Aarons, S.R.; Powell, J.M. Nitrogen use efficiency and manure management practices in contrasting dairy production systems. Agric. Ecosyst. Environ. 2012, 147, 73–81. [Google Scholar] [CrossRef]
- Macdonald, K.A.; Penno, J.W.; Lancaster, J.A.S.; Bryant, A.M.; Kidd, J.M.; Roche, J.R. Production and economic responses to intensification of pasture-based dairy production systems. J. Dairy Sci. 2017, 100, 6602–6619. [Google Scholar] [CrossRef]
- LIC. New Zealand Dairy Statistics 2016–17; DairyNZ: Hamilton, New Zealand, 2017. [Google Scholar]
- De Klein, C.A.M.; Monaghan, R.M.; Alfaro, M.; Gourley, C.J.P.; Oenema, O.; Powell, J.M. Nitrogen performance indicators for dairy production systems. Soil Res. 2017, 55, 479–488. [Google Scholar] [CrossRef]
- Parsons, A.; Thornley, J.H.M.; Rasmussen, S.; Rowarth, J.S. Some clarification of the impacts of grassland intensification on food production, nitrogen release, greenhouse gas emissions and carbon sequestration: Using the example of New Zealand. CAB Reviews: Perspect. Agric., Vet. Sci., Nutr. Nat. Resour. 2016, 11. [Google Scholar] [CrossRef]
- Foote, K.J.; Joy, M.K.; Death, R.G. New Zealand dairy farming: Milking our environment for all its worth. Environ. Manag. 2015, 56, 709–720. [Google Scholar] [CrossRef]
- Jay, M. The political economy of a productivist agriculture: New Zealand dairy discourses. Food Policy 2007, 32, 266–279. [Google Scholar] [CrossRef] [Green Version]
- Capuco, A.V.; Ellis, S.E.; Hale, S.A.; Long, E.; Erdman, R.A.; Zhao, X.; Paape, M.J. Lactation persistency: Insights from mammary cell proliferation studies. J. Animal Sci. 2003, 81, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Clark, D.A.; Phyn, C.V.; Tong, M.J.; Collis, S.J.; Dalley, D.E. A systems comparison of once- versus twice-daily milking of pastured dairy cows. J. Dairy Sci. 2006, 89, 1854–1862. [Google Scholar] [CrossRef]
- Edwards, J.P. Comparison of milk production and herd characteristics in New Zealand herds milked once or twice a day. Anim. Prod. Sci. 2018. [Google Scholar] [CrossRef]
- Lynch, G.A.; Hunt, M.E.; MacKenzie, D.D.S. The effects of once daily milking as a management practice in late lactation. Proc. N. Z. Soc. Anim. Prod. 1991, 51, 191–195. [Google Scholar]
- Bewsell, D.; Clark, D.A.; Dalley, D.E. Understanding motivations to adopt once-a-day milking amongst New Zealand dairy farmers. J. Agric. Educ. Ext. 2008, 14, 69–80. [Google Scholar] [CrossRef]
- Stelwagen, K.; Phyn, C.V.C.; Davis, S.R.; Guinard-Flament, J.; Pomiès, D.; Roche, J.R.; Kay, J.K. Invited review: Reduced milking frequency: Milk production and management implications. J. Dairy Sci. 2013, 96, 3401–3413. [Google Scholar] [CrossRef]
- L’Huillier, P.J.; Thomson, N.A. Estimation of herbage mass in ryegrass/white clover dairy pastures. Proc. N. Z. Grassl. Assoc. 1988, 41, 117–122. [Google Scholar]
- Baker, R.D. Estimating herbage intake from animal performance. In Herbage Intake Handbook, 2nd ed.; British Grassland Society: London, UK, 2004; p. 191. [Google Scholar]
- Corson, D.C.; Waghorn, G.C.; Ulyatt, M.J.; Lee, J. NIRS: Forage analysis and livestock feeding. Proc. N. Z. Grassl. Assoc. 1999, 61, 127–132. [Google Scholar]
- Arunvipas, P.; Van Leeuwen, J.A.; Dohoo, I.R.; Keefe, G.P. Evaluation of the reliability and repeatability of automated milk urea nitrogen testing. Can. J. Vet. Sci. 2003, 67, 60–63. [Google Scholar]
- Macdonald, K.A.; Macmillan, K.L. Condition score and liveweight in Jersey and Friesian cows. Proc. Ruakura Farmers Conf. 1993, 45, 47–50. [Google Scholar]
- Macciotta, N.P.P.; Vicario, D.; Cappio-Borlino, A. Detection of different shapes of lactation curve for milk yield in dairy cattle by empirical mathematical models. J. Dairy Sci. 2005, 88, 1178–1191. [Google Scholar] [CrossRef] [Green Version]
- Silvestre, A.M.; Petim-Batista, F.; Colaço, J. The accuracy of seven mathematical functions in modeling dairy cattle from lactation curves based on test-day records from varying sample schemes. J. Dairy Sci. 2006, 89, 1813–1821. [Google Scholar] [CrossRef] [Green Version]
- Jarrige, R.; Demarquilly, C.; Dulphy, J.P.; Hoden, A.; Robelin, J.; Beranger, C.; Geay, Y.; Journet, M.; Malterre, C.; Micol, D.; et al. The INRA "fill unit" system for predicting the voluntary intake of forage-based diets in ruminants: A review. J. Anim. Sci. 1986, 63, 1737–1758. [Google Scholar] [CrossRef]
- Berry, D.P.; Veerkamp, R.F.; Dillon, P. Phenotypic profiles for body weight, body condition score, energy intake, and energy balance across different parities and concentrate feeding levels. Livest. Sci. 2006, 104, 1–12. [Google Scholar] [CrossRef]
- Spanghero, M.; Kowalski, Z.M. Critical analysis of N balance experiments with lactating cows. Livest. Prod. Sci. 1997, 52, 113–122. [Google Scholar] [CrossRef]
- Huhtanen, P.; Cabezas-Garcia, E.H.; Krizsan, S.J.; Shingfield, K.J. Evaluation of between-cow variation in milk urea and rumen ammonia nitrogen concentrations and the association with nitrogen utilization and diet digestibility in lactating cows. J. Dairy Sci. 2015, 98, 3182–3196. [Google Scholar] [CrossRef] [Green Version]
- Peyraud, J.L.; Vérité, R.; Delaby, L. Nitrogen excretion by dairy cows: Effect of the diet and of the level of production. Fourrages. 1995, 142, 131–144. [Google Scholar]
- Reed, K.F.; Moraes, L.E.; Casper, D.P.; Kebreab, E. Predicting nitrogen excretion from cattle. J. Dairy Sci. 2015, 98, 3025–3035. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, H.; Berthiaume, R.; Raggio, G.; Thivierge, M.C.; Doepel, L.; Pacheco, D.; Dubreuil, P.; Lobley, G.E. The route of absorbed nitrogen into milk protein. Anim. Sci. 2005, 80, 11–22. [Google Scholar] [CrossRef]
- Daniel, J.B.; Friggens, N.C.; Van Laar, H.; Ferris, C.P.; Sauvant, D. A method to estimate cow potential and subsequent responses to energy and protein supply according to stage of lactation. J. Dairy Sci. 2017, 100, 3641–3657. [Google Scholar] [CrossRef] [PubMed]
- Spek, J.W.; Dijkstra, J.; Van Duinkerken, G.; Bannink, A. A review of factors influencing milk urea concentration and its relationship with urinary urea excretion in lactating dairy cattle. J. Agricult. Sci. 2013, 151, 407–423. [Google Scholar] [CrossRef] [Green Version]
- Friggens, N.C.; Rasmussen, M.D. Milk quality assessment in automatic milking systems: Accounting for the effects of variable intervals between milkings on milk composition. Livest. Prod. Sci. 2001, 73, 45–54. [Google Scholar] [CrossRef]
- Nielsen, N.I.; Larsen, T.; Bjerring, M.; Ingvartsen, K.L. Quarter health, milking interval, and sampling time during milking affect the concentration of milk constituents. J. Dairy Sci. 2005, 88, 3186–3200. [Google Scholar] [CrossRef] [Green Version]
- Barros, T.; Quaassdorff, M.A.; Aguerre, M.J.; Olmos Colmenero, J.J.; Bertics, S.J.; Crump, P.M.; Wattiaux, M.A. Effects of dietary crude protein concentration on late-lactation dairy cow performance and indicators of nitrogen utilization. J. Dairy Sci. 2017, 100, 5434–5448. [Google Scholar] [CrossRef] [PubMed]
- Mutsvangwa, T.; Davies, K.L.; McKinnon, J.J.; Christensen, D.A. Effects of dietary crude protein and rumen-degradable protein concentrations on urea recycling, nitrogen balance, omasal nutrient flow, and milk production in dairy cows. J. Dairy Sci. 2016, 99, 6298–6310. [Google Scholar] [CrossRef] [PubMed]
- De Campeneere, S.; De Brabander, D.L.; Vanacker, J.M. Milk urea concentration as affected by the roughage type offered to dairy cattle. Livest. Sci. 2006, 103, 30–39. [Google Scholar] [CrossRef]
- Hynes, D.N.; Stergiadis, S.; Gordon, A.; Yan, T. Effects of crude protein level in concentrate supplements on animal performance and nitrogen utilization of lactating dairy cows fed fresh-cut perennial grass. J. Dairy Sci. 2016, 99, 8111–8120. [Google Scholar] [CrossRef] [Green Version]
- Trevaskis, L.M.; Fulkerson, W.J. The relationship between various animal and management factors and milk urea, and its association with reproductive performance of dairy cows grazing pasture. Livest. Prod. Sci. 1999, 57, 255–265. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Item | Production System | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Low Intensity | High Intensity | ||||||||||||
| Pasture 1 | Pasture Silage | Brassica | Herb-Mix 2 | Lucerne | Total | Pasture 1 | Pasture Silage | Brassica | Maize Silage | Concentrate 3 | DDG 4 | Total | |
| Feed allocation, % DM | 92 | 3 | 1 | 3 | 1 | 100 | 60 | 8 | 1 | 15 | 15 | 1 | 100 |
| ME, MJ ME/kg DM | 11.0 | 10.7 | 11.7 | 12.1 | 9.9 | 11.1 | 10.5 | 10.9 | 7.8 | 10.4 | 12.6 | 9.5 | 10.3 |
| CP, % of DM | 20 | 14 | 16 | 20 | 25 | 19 | 19 | 12 | 22 | 7 | 17 | 21 | 16 |
| ADF, % of DM | 22 | 34 | 22 | 15 | 21 | 22.8 | 24 | 40 | 18 | 28 | 2 | 11 | 21 |
| NDF, % of DM | 44 | 50 | 34 | 29 | 34 | 38.2 | 46 | 56 | 31 | 37 | 23 | 28 | 37 |
| OMD, % of DM | 75 | 67 | 79 | 83 | 68 | 74.4 | 73 | 68 | 81 | - | - | - | 74 |
| Variable | Production System | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Low Intensity | High Intensity | |||||||||
| N | Mean | SD | Min | Max | N | Mean | SD | Min | Max | |
| Lactation length, days | 257 | 269 | 33 | 127 | 319 | 207 | 272 | 36 | 110 | 321 |
| Milk yield, kg/day | 2284 | 15.7 | 6.0 | 0.9 | 36.6 | 1217 | 21.2 | 5.5 | 3.0 | 37.7 |
| Fat yield, kg/day | 2284 | 0.8 | 0.3 | 0.1 | 2.5 | 1217 | 1.0 | 0.2 | 0.2 | 1.5 |
| Protein yield, kg/day | 2284 | 0.6 | 0.2 | 0.1 | 1.3 | 1217 | 0.8 | 0.2 | 0.2 | 1.4 |
| Lactose yield, kg/day | 726 | 0.8 | 0.3 | 0.1 | 1.9 | 558 | 1.1 | 0.3 | 0.5 | 1.9 |
| Somatic cell score 1 | 2276 | 5.76 | 1.59 | 0.01 | 12.43 | 1217 | 5.13 | 1.45 | 1.58 | 12.02 |
| MUN 2, mg/dL day | 726 | 13.11 | 4.70 | 4.25 | 28.83 | 557 | 9.72 | 2.58 | 2.84 | 18.42 |
| MUNY 3, g/day | 726 | 1.9 | 0.8 | 0.3 | 4.7 | 557 | 2.1 | 0.6 | 0.4 | 3.9 |
| Body weight, kg | 2359 | 487 | 70 | 320 | 684 | 1999 | 502 | 62 | 352 | 770 |
| Body condition score | 2060 | 4.6 | 0.4 | 3.0 | 6.5 | 1405 | 4.2 | 0.4 | 3.0 | 5.5 |
| Trait | Overall | Production System | |
|---|---|---|---|
| LIPS | HIPS | ||
| Milk yield, kg/day | 0.997 ± 0.003 | 0.996 ± 0.004 | 0.995 ± 0.005 |
| Fat yield, kg/day | 0.992 ± 0.004 | 0.992 ± 0.006 | 0.990 ± 0.007 |
| Protein yield, kg/day | 0.994 ± 0.004 | 0.995 ± 0.005 | 0.991 ± 0.007 |
| Somatic cell score 2 | 0.923 ± 0.005 | 0.922 ± 0.006 | 0.916 ± 0.008 |
| Milk urea nitrogen, mg/dL day | 0.989 ± 0.003 | 0.991 ± 0.004 | 0.968 ± 0.008 |
| Milk urea nitrogen yield, g/day | 0.941 ± 0.007 | 0.960 ± 0.007 | 0.898 ± 0.014 |
| Body weight, kg | 0.988 ± 0.002 | 0.995 ± 0.001 | 0.976 ± 0.003 |
| Body condition score 3 | 0.868 ± 0.006 | 0.846 ± 0.009 | 0.804 ± 0.012 |
| Trait | Herd | α0 | α1 | α2 | α3 |
|---|---|---|---|---|---|
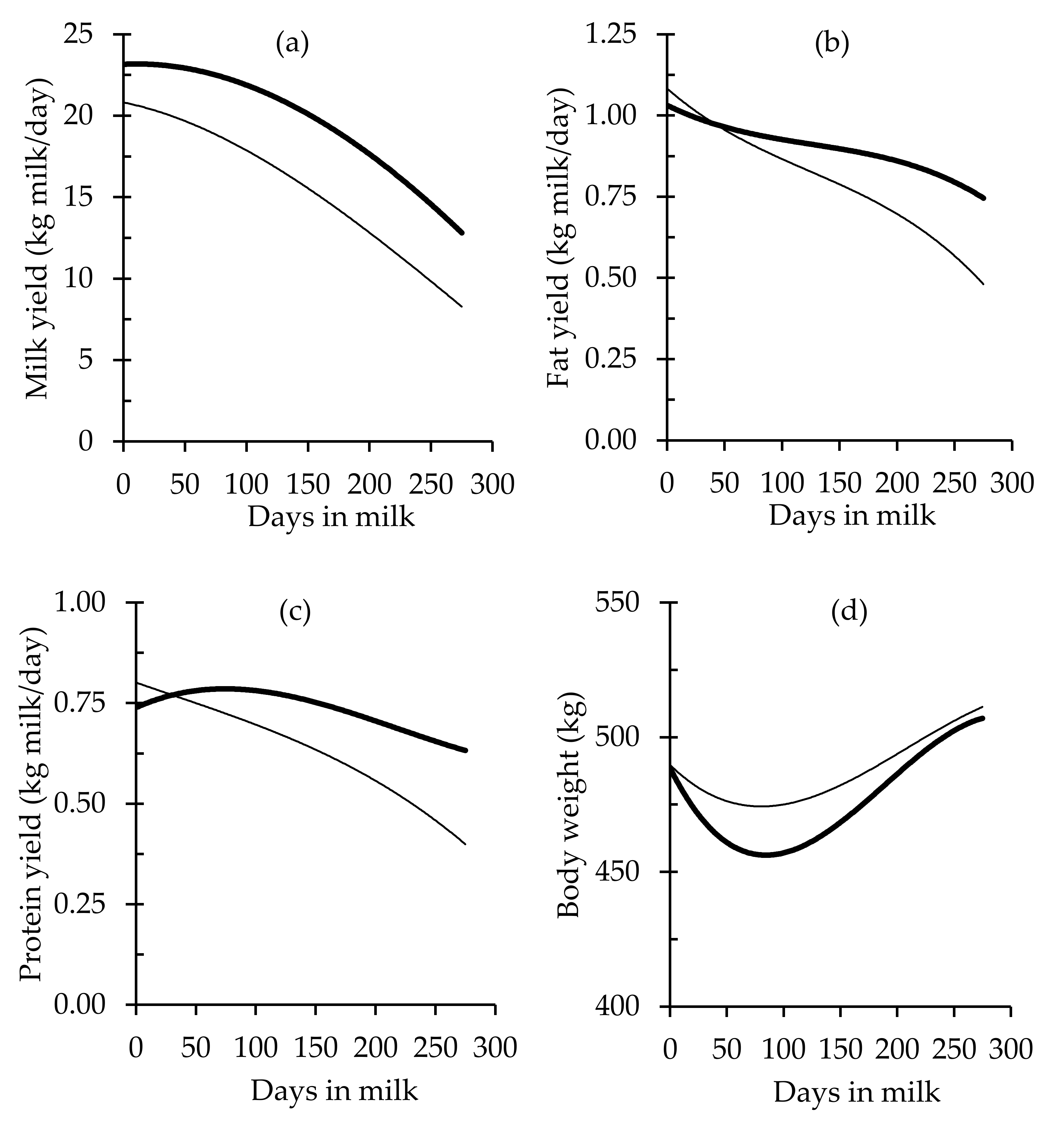
| Milk yield, kg/day | LIPS | 15.20 b ± 0.16 | −6.94 b ± 0.12 | −1.13 b ± 0.06 | 0.21 b ± 0.02 |
| HIPS | 19.33 a ± 0.22 | −5.83 a ± 0.17 | −1.90 a ± 0.09 | 0.10 a ± 0.02 | |
| Fat yield, kg/day | LIPS | 0.78 b ± 0.01 | −0.29 b ± 0.01 | −0.03 b ± 0.00 | −0.04 b ± 0.00 |
| HIPS | 0.89 a ± 0.01 | −0.13 a ± 0.01 | −0.02 a ± 0.00 | −0.03 a ± 0.00 | |
| Protein yield, kg/day | LIPS | 0.62 b ± 0.01 | −0.21 b ± 0.01 | −0.04 ± 0.00 | −0.01 b ± 0.00 |
| HIPS | 0.73 a ± 0.01 | −0.08 a ± 0.01 | −0.05 ± 0.00 | 0.02 a ± 0.00 | |
| Lactose yield, kg/day | LIPS | 0.77 b ± 0.01 | −0.37 ± 0.01 | 0.01 b ± 0.00 | 0.05 a ± 0.00 |
| HIPS | 1.07 a ± 0.01 | −0.39 ± 0.01 | 0.02 a ± 0.00 | −0.05 b ± 0.00 | |
| Somatic cell score 1 | LIPS | 5.80 a ± 0.07 | 1.12 a ± 0.05 | 0.04 b ± 0.04 | 0.08 ± 0.02 |
| HIPS | 5.02 b ± 0.10 | 0.71 b ± 0.07 | 0.66 a ± 0.05 | 0.03 ± 0.03 | |
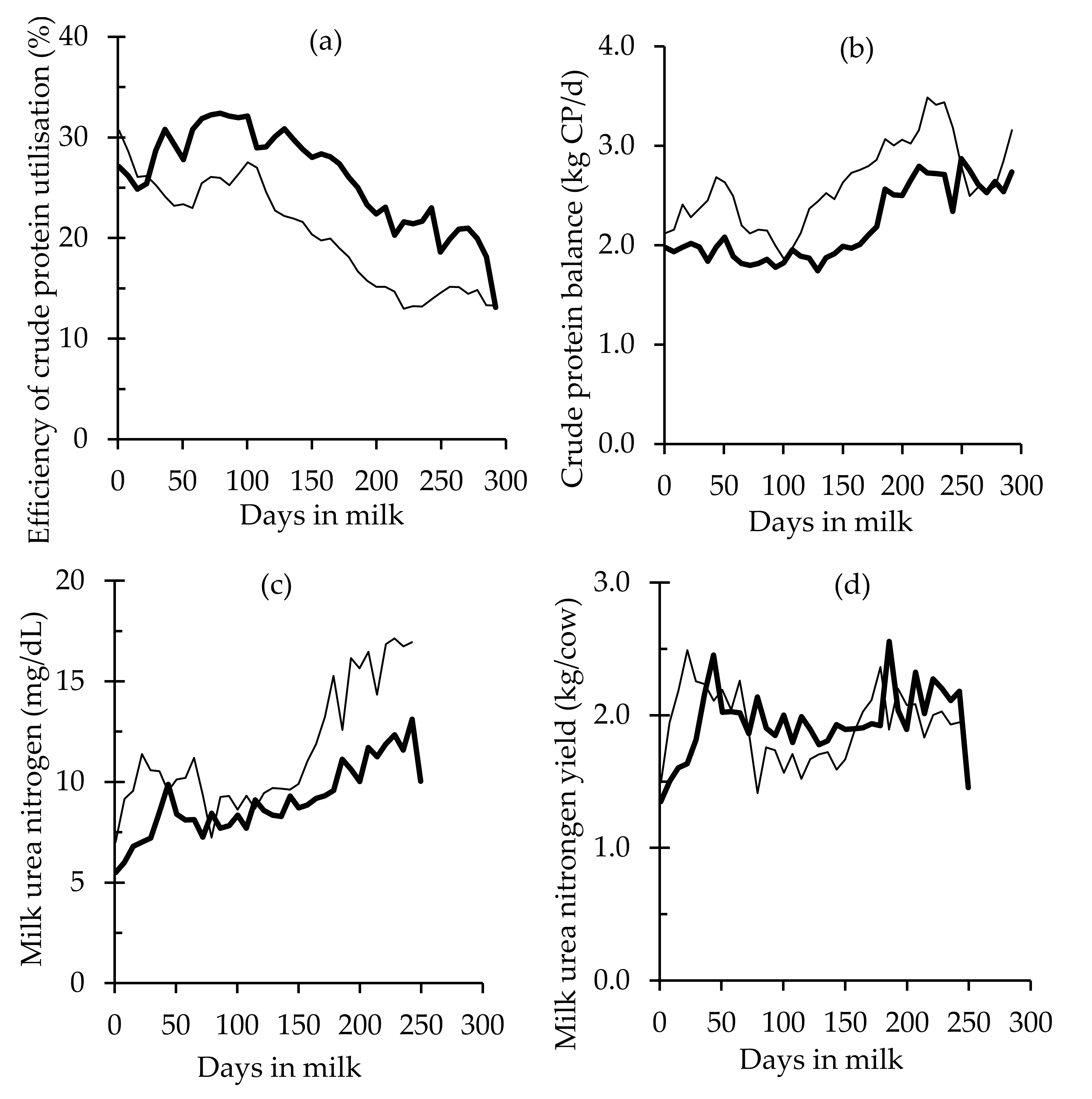
| Milk urea, mg/dL day | LIPS | 14.49 a ± 0.14 | 7.64 a ± 0.14 | 6.00 a ± 0.15 | −0.59 ± 0.05 |
| HIPS | 10.94 b ± 0.20 | 3.91 b ± 0.19 | 3.26 b ± 0.21 | −0.49 ± 0.08 | |
| Milk urea yield, g/day | LIPS | 1.99 b ± 0.03 | −0.17 ± 0.02 | 0.48 b ± 0.01 | −0.33 ± 0.01 |
| HIPS | 2.10 a ± 0.04 | −0.14 ± 0.03 | 0.50 a ± 0.01 | −0.33 ± 0.02 | |
| Live weight, kg | LIPS | 488.06 a ± 2.71 | 18.35 b ± 0.94 | 13.48 b ± 0.71 | −6.15 a ± 0.91 |
| HIPS | 477.34 b ± 3.72 | 23.81 a ± 1.29 | 20.61 a ± 0.98 | −13.66 b ± 1.25 | |
| Body condition score 2 | LIPS | 4.59 a ± 0.02 | −0.13 ± 0.01 | 0.23 b ± 0.01 | 0.05 a ± 0.00 |
| HIPS | 4.25 b ± 0.02 | −0.12 ± 0.01 | 0.27 a ± 0.01 | −0.01 b ± 0.01 |
| Item (per Cow) | Production System | ||
|---|---|---|---|
| Low Intensity | High Intensity | p – Value 1 | |
| N | 257 | 207 | |
| Lactation length, days | 272 ± 2 | 271 ± 3 | 0.719 |
| Total milk yield, kg | 4232.40 ± 55.3 | 5332.10 ± 75.9 | <0.001 |
| Total milk solids yield, kg | 387.6 ± 4.4 | 443.7 ± 6.1 | <0.001 |
| Total fat yield, kg | 217.4 ± 2.5 | 243.9 ± 3.5 | <0.001 |
| Total protein yield, kg | 170.5 ± 2 | 199.7 ± 2.8 | <0.001 |
| Total lactose yield, kg | 214.7 ± 3 | 296.3 ± 4.1 | <0.001 |
| Somatic cell score 2 | 5.73 ± 0.07 | 4.95 ± 0.1 | <0.001 |
| Body weight, kg | 487 ± 3 | 475 ± 4 | 0.012 |
| Body condition score 3 | 4.6 ± 0.02 | 4.26 ± 0.02 | <0.001 |
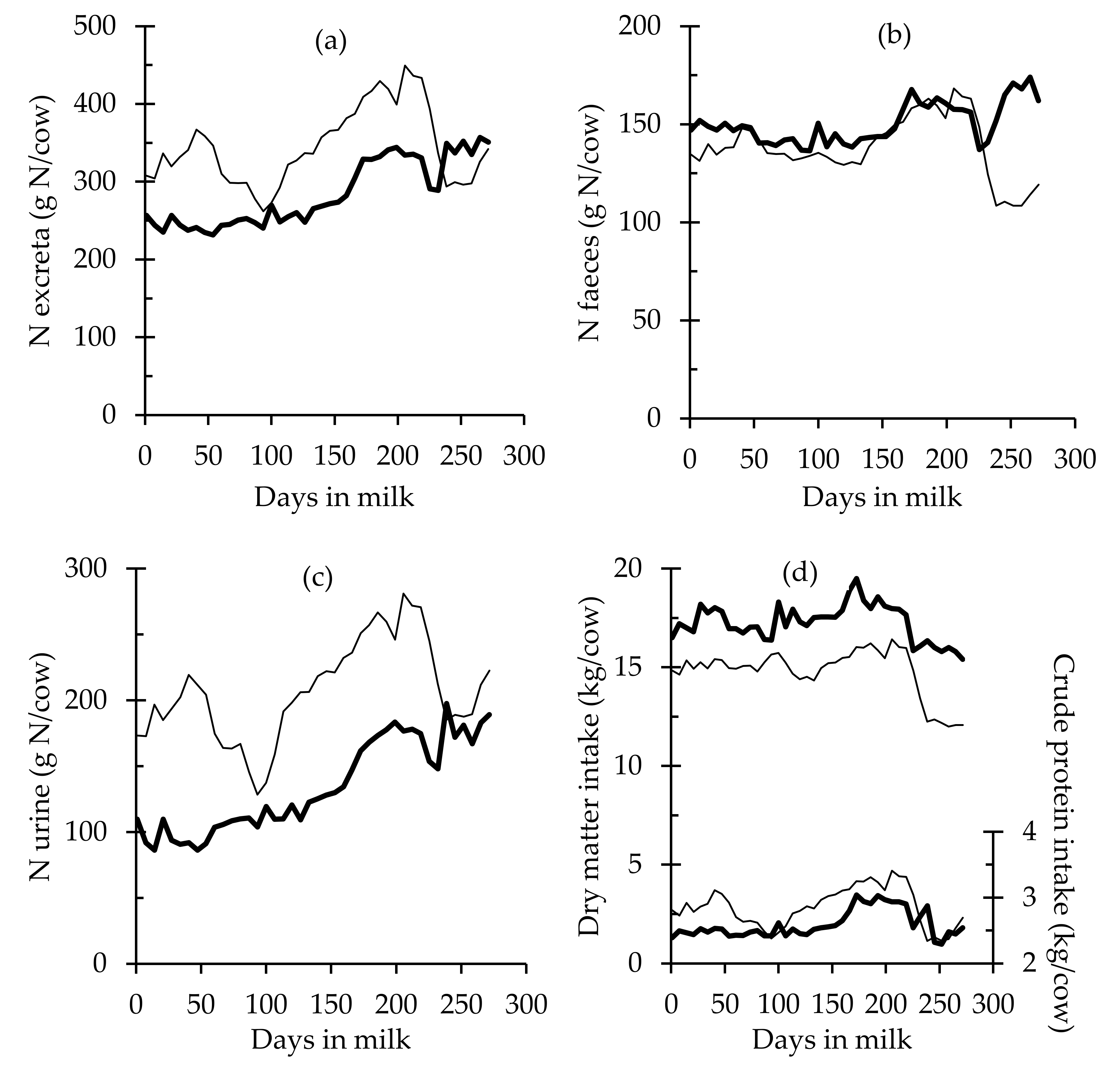
| DMI, kg/day | 16.21 ± 0.07 | 18.49 ± 0.13 | <0.001 |
| CPI, kg/day | 3.13 ± 0.01 | 2.8 ± 0.03 | <0.001 |
| MUN, mg/dL | 13.20 ± 0.34 | 9.99 ± 0.23 | <0.001 |
| MUNY, g | 535.06 ± 19.6 | 565.01 ± 12.57 | 0.058 |
| ECPU, % | 20.13 ± 0.13 | 27.65 ± 0.33 | <0.001 |
| CPB, kg CP/day | 2.51 ± 0.01 | 2.03 ± 0.03 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Correa-Luna, M.; Donaghy, D.; Kemp, P.; Schutz, M.; López-Villalobos, N. Efficiency of Crude Protein Utilisation in Grazing Dairy Cows: A Case Study Comparing Two Production Systems Differing in Intensification Level in New Zealand. Animals 2020, 10, 1036. https://doi.org/10.3390/ani10061036
Correa-Luna M, Donaghy D, Kemp P, Schutz M, López-Villalobos N. Efficiency of Crude Protein Utilisation in Grazing Dairy Cows: A Case Study Comparing Two Production Systems Differing in Intensification Level in New Zealand. Animals. 2020; 10(6):1036. https://doi.org/10.3390/ani10061036
Chicago/Turabian StyleCorrea-Luna, Martín, Daniel Donaghy, Peter Kemp, Michael Schutz, and Nicolas López-Villalobos. 2020. "Efficiency of Crude Protein Utilisation in Grazing Dairy Cows: A Case Study Comparing Two Production Systems Differing in Intensification Level in New Zealand" Animals 10, no. 6: 1036. https://doi.org/10.3390/ani10061036