Influence of Different Photoperiod and Temperature Regimes on Growth and Bulb Quality of Garlic (Allium sativum L.) Cultivars

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Cloves Sowing Site Description
2.2. Photoperiod and Temperature Treatments
2.3. Measurement of Garlic Plant Morphological Indices, Bulb Characteristics, Growth Period, and Bulbing Index
2.4. Assessment of Garlic Bulb Physiological and Nutritive Quality Indices
2.4.1. TSS (Total Soluble Solid)
2.4.2. Soluble Protein Content
2.4.3. Soluble Sugar Content
2.4.4. Total Sugar Content
2.4.5. Glucose Content
2.4.6. Sucrose Content
2.4.7. Fructose Content
2.4.8. Starch Content
2.4.9. Total Phenol Content
2.4.10. Total Flavonoid Content
2.5. Statistical Analysis
3. Results
3.1. The Main Effect of Cultivar, Photoperiod and Temperature Treatments on the Garlic Plant Morphology, Bulb Characteristics, Growth Period, and Bulbing Index
3.2. Interaction Effects of Photoperiod and Temperature Treatments on Plant Morphology of Three Garlic Cultivars
3.3. Interaction Effects of Photoperiod and Temperature Treatments on Bulb Characteristics and Growth Period of Three Garlic Cultivars
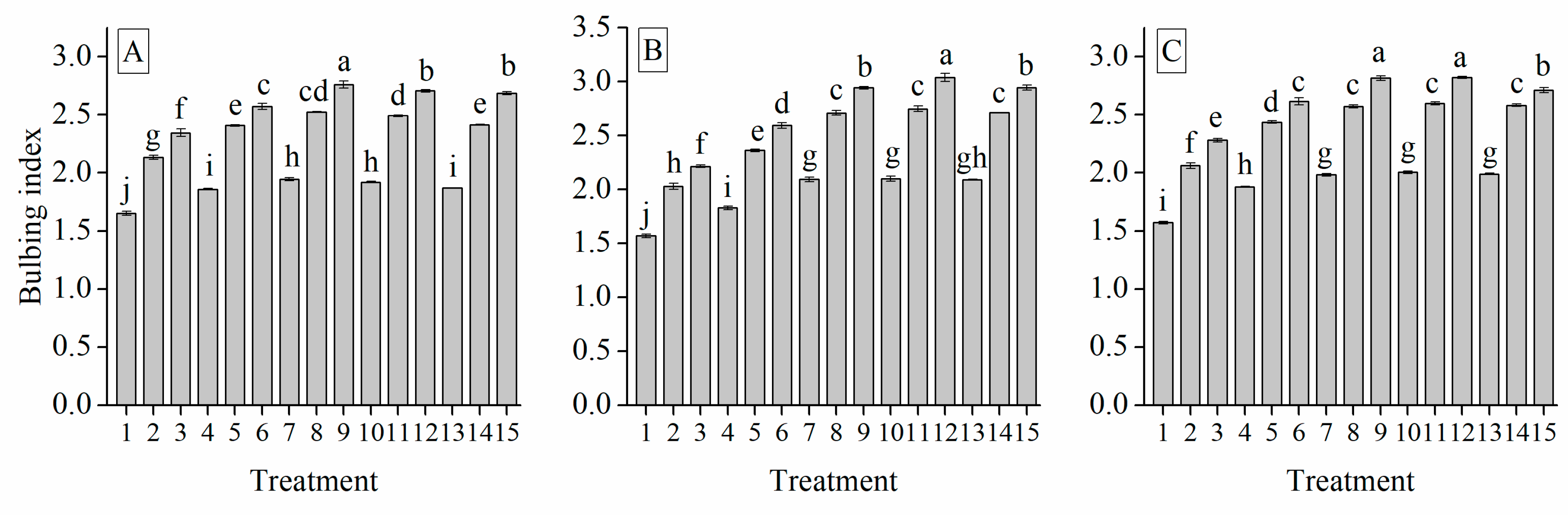
3.4. Interaction of Cultivar, Temperature and Photoperiod Treatments on Bulbing Index
3.5. The Main Effect of Cultivar, Photoperiod, and Temperature Treatments on the Garlic Bulb Physiological and Nutritive Quality Traits
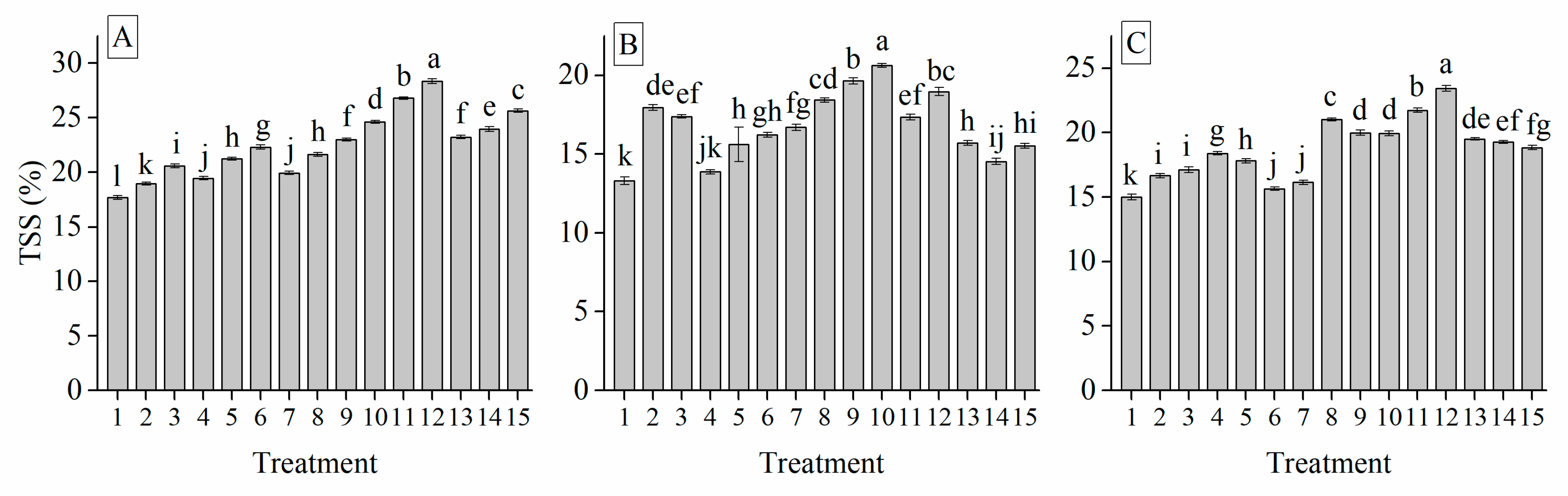
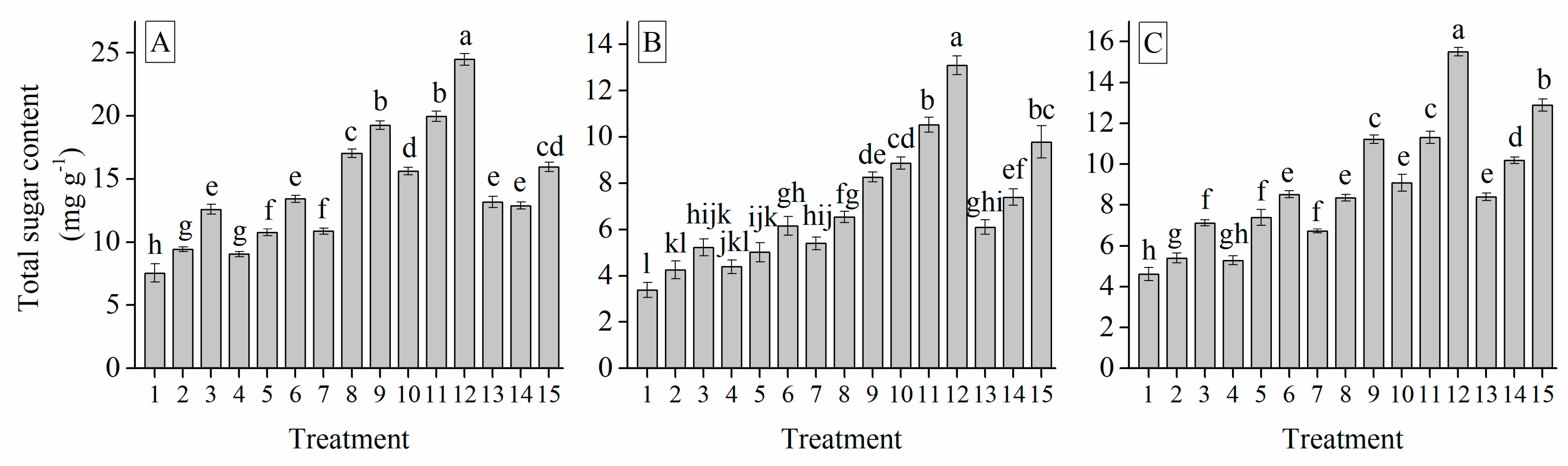
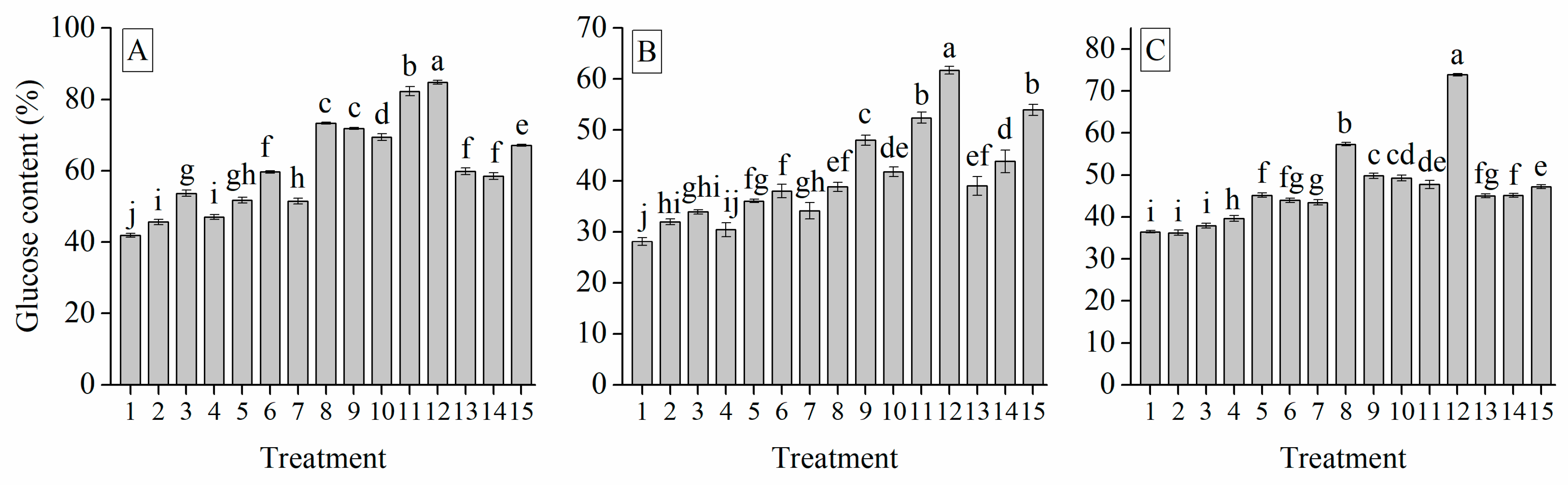
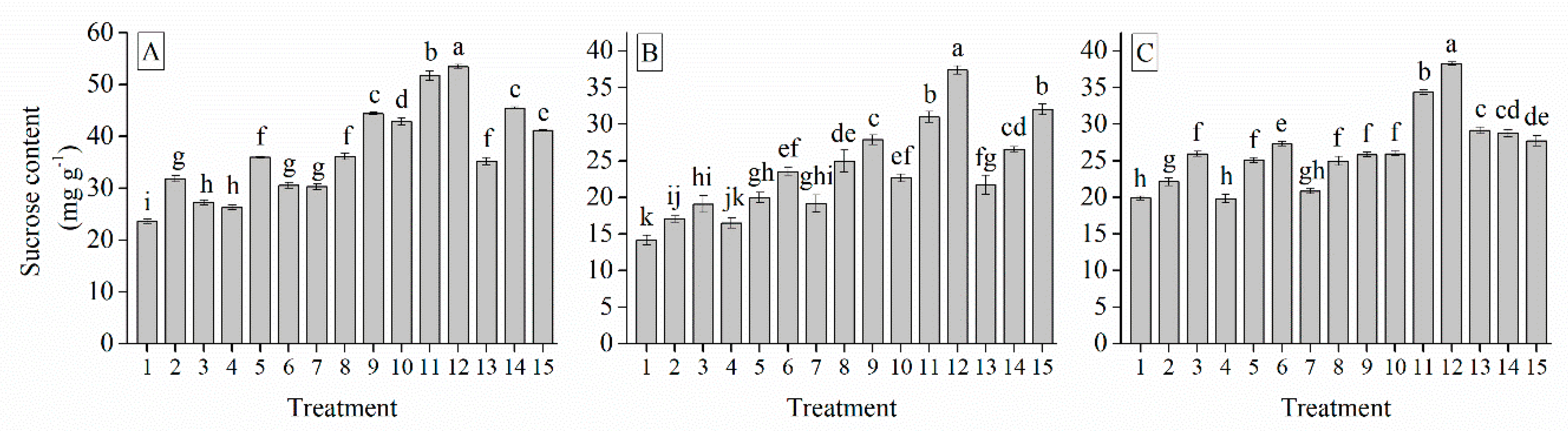
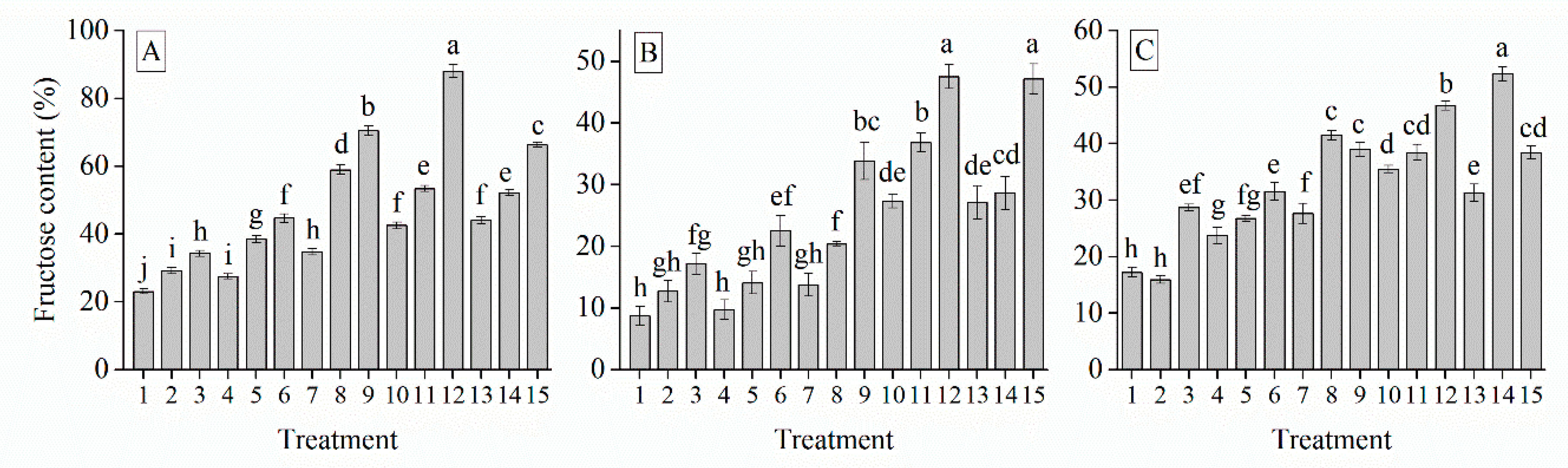
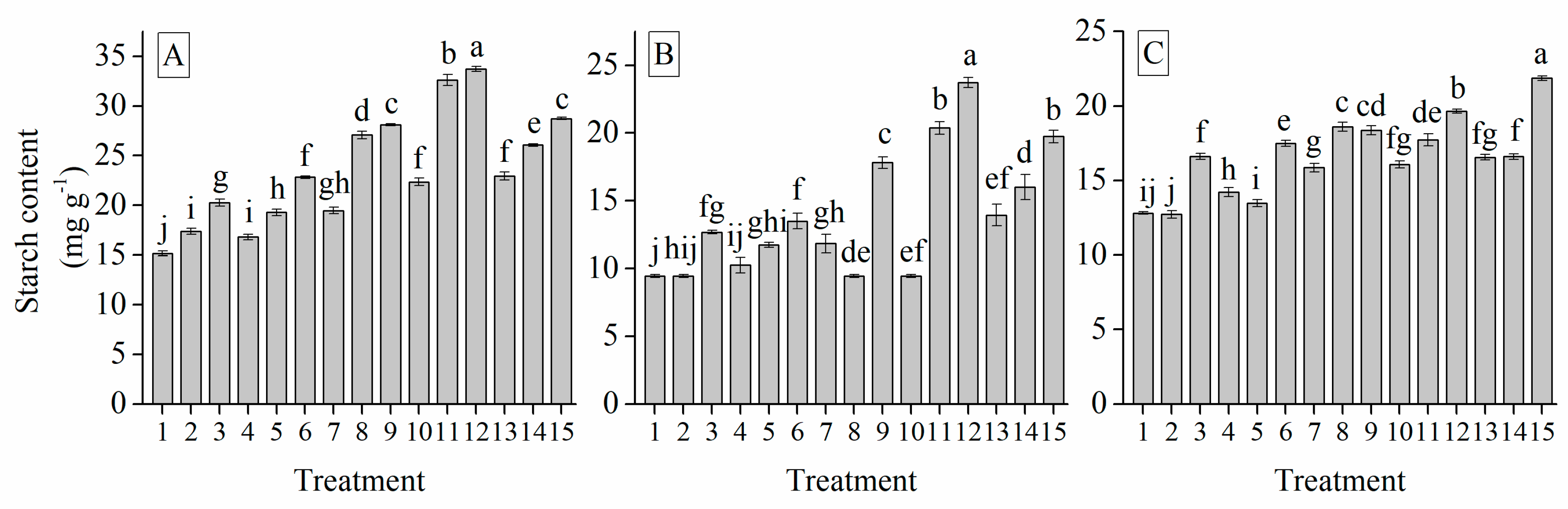
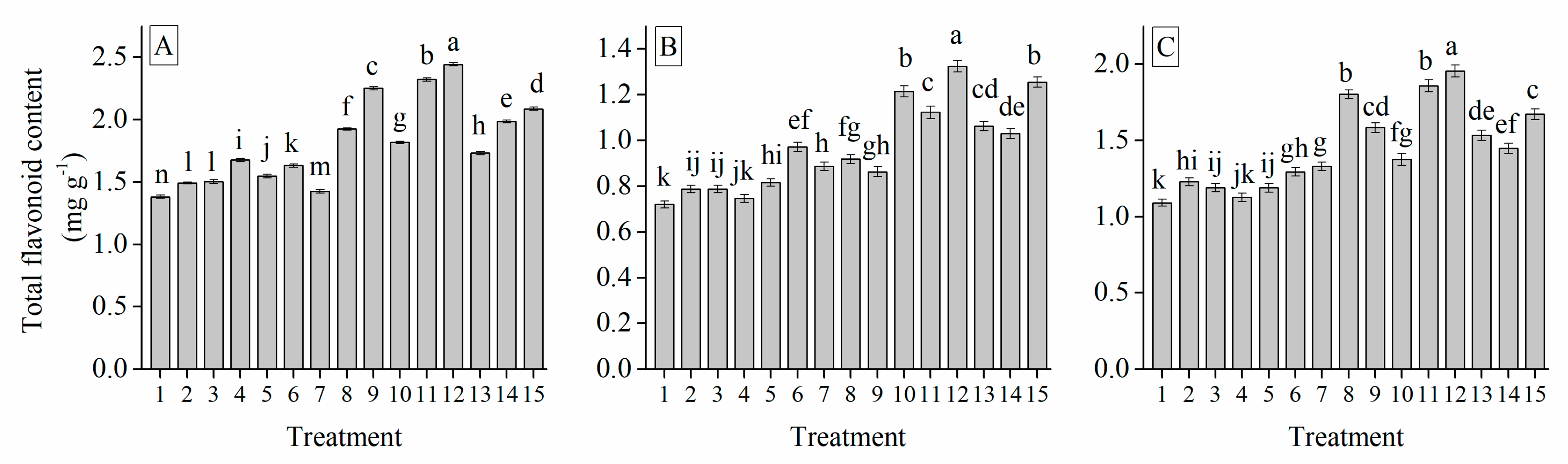
3.6. Interaction of Cultivar, Photoperiod and Temperature Treatments on the Garlic Bulb Physiology and Nutritive Quality Traits
3.7. The Main Effect of Cultivar, Photoperiod and Temperature Treatments on the Garlic Bulb Phenolic Compounds
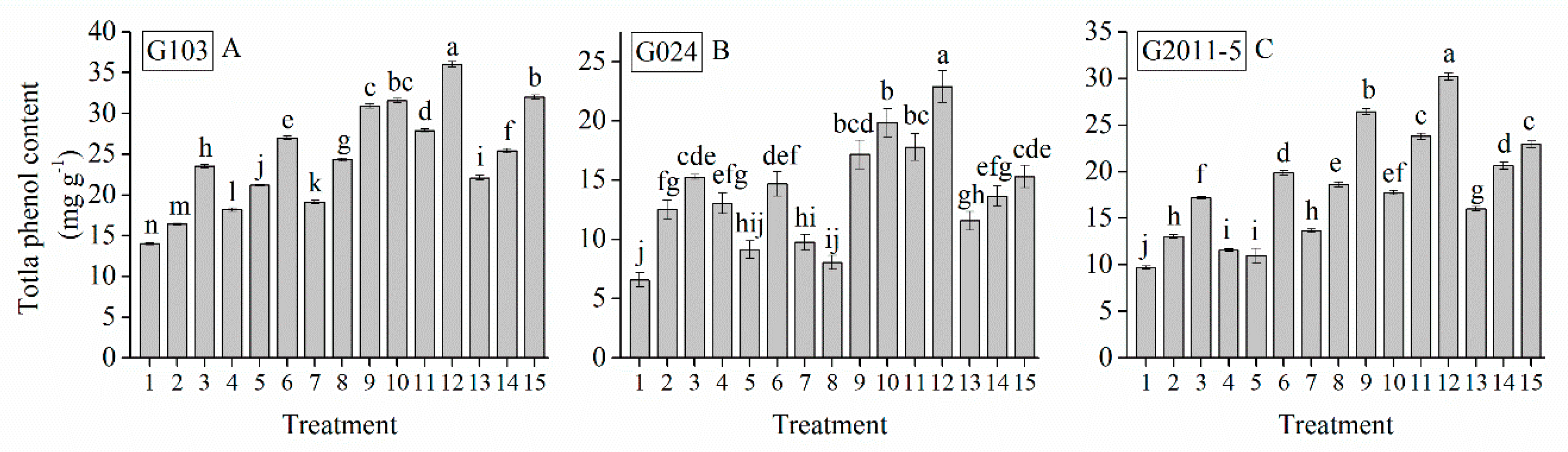
3.8. Interaction of Cultivar, Photoperiod, and Temperature Treatments on Garlic Bulb Phenolic Compounds
3.9. Correlation Analysis
4. Discussion
4.1. Effect of Photoperiod and Temperature on Garlic Plant Growth Traits and Bulbing
4.2. Role of Photoperiod and Temperature in the Garlic Bulb Quality Attributes
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wang, H.; Li, X.; Shen, D.; Oiu, Y.; Song, J. Diversity evaluation of morphological traits and allicin content in garlic (Allium sativum L.) from China. Euphytica 2014, 198, 243–254. [Google Scholar] [CrossRef]
- Grubben, G.J.H.; Denton, C.M. Plant Resources of Tropical Africa 2: Vegetables. In Plant Resources of Tropical Asia Foundation; Bakhuys Publishers: Leiden, The Netherlands, 2004; p. 59. [Google Scholar]
- Wu, C.; Wang, M.; Dong, Y.; Cheng, Z.; Meng, H. Growth, bolting and yield of garlic (Allium sativum L.) in response to clove chilling treatment. Sci. Hortic. 2015, 194, 43–52. [Google Scholar] [CrossRef]
- Chia, P.L.; Kubota, C. End-of-day far-red light quality and dose requirements for tomato rootstock hypocotyl elongation. HortScience 2010, 45, 1501–1506. [Google Scholar] [CrossRef] [Green Version]
- Craig, D.S.; Runkle, E.S. An intermediate phytochrome photoequilibria from night-interruption lighting optimally promotes flowering of several long-day plants. Environ. Exp. Bot. 2016, 121, 132–138. [Google Scholar] [CrossRef]
- Mathew, D.; Forer, Y.; Rabinowitch, H.D.; Kamenetsky, R. Effect of long photoperiod on the reproductive and bulbing processes in garlic (Allium sativum L.) genotypes. Environ. Exp. Bot. 2010, 71, 166–173. [Google Scholar] [CrossRef]
- Miralles, D.J.; Richards, R.A. Responses of leaf and tiller emergence and primordium initiation in wheat and barley to interchanged photoperiod. Ann. Bot. 2000, 85, 655–663. [Google Scholar] [CrossRef]
- Lefsrud, M.G.; Kopsell, D.A.; Auge, R.M.; Both, A.J. Biomass production and pigment accumulation in kale grown under increasing photoperiods. HortScience 2006, 41, 603–606. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.D.; Khandaker, L.; Oba, S. Comparative study on functional components, antioxidant activity and color parameters of selected colored leafy vegetables as affected by photoperiods. J. Food Agric. Environ. 2009, 7, 392–398. [Google Scholar]
- Xu, H.L.; Xu, Q.C.; Li, F.L.; Feng, Y.Z.; Qin, F.F.; Fang, W. Applications of xerophytophysiology in plant production-LED blue light as a stimulus improved the tomato crop. Sci. Hortic. 2012, 148, 190–196. [Google Scholar] [CrossRef]
- Steer, B.T. The role of night temperature in the bulbing of onion (Allium cepa L.). Aust. J. Agric. Res. 1980, 31, 519–523. [Google Scholar] [CrossRef]
- Bandara, M.S.; Krieger, K.; Slinkard, A.E.; Tanino, K.K. Pre-plant chilling requirements for cloving of spring-planted garlic. Can. J. Plant Sci. 2000, 80, 379–384. [Google Scholar] [CrossRef]
- Etoh, T.; Simon, P.W. Diversity, fertility and seed production in garlic. In Allium Crop Science: Recent Advances; Rabinowitch, H.D., Currah, L., Eds.; CAB International: Wallingford, UK, 2002; pp. 101–117. [Google Scholar]
- Gaskell, M.; Cantwell, M.; Nie, X.; Smith, S.; Faber, B.; Voss, R. Effects of transplant date and transplant size on production, quality and pungency of sweet onions. In Proceedings of the National Onion (and other Allium) Research Conference, Sacramento, CA, USA, 10–12 December 1998; pp. 337–341. [Google Scholar]
- Takagi, H. Garlic (Allium sativum L.). In Onions and Allied Crops, Volume 3: Biochemistry, Food Science, and Minor Crops; Rabinowitch, H.D., Brewster, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 109–157. [Google Scholar]
- Kamenetsky, R. Garlic: Botany and horticulture. Hortic. Rev. 2007, 33, 123. [Google Scholar]
- Huyskens-Keil, S.; Schreiner, M. Quality dynamics and quality assurance of fresh fruits and vegetables in pre and postharvest. In Production Practices and Quality Assessment of Food Crops; Dris, R., Jain, S.M., Eds.; Quality Handing and Evaluation; Kluwer Academic Publishers: Dordrecht, The Netherland, 2004; Volume 3, pp. 401–409. [Google Scholar]
- Filgueira, F.A.R. New Manual of Olericulture, 3rd ed.; UFV: Vicosa, Brazil, 2008; p. 421. [Google Scholar]
- Brewster, J.L. Physiology of crop growth and bulbing. In Onions and other Vegetable Alliums; Rabinowitch, H.D., Brewster, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 53–88. [Google Scholar]
- Lercari, B. The promoting effect of far-red light on bulb formation in the long day plant Allium cepa L. Plant Sci. Lett. 1982, 27, 243–254. [Google Scholar] [CrossRef]
- Hemm, M.R.; Rider, S.D.; Ogas, J.; Murry, D.J.; Chapple, C. Light induces phenylpropanoid metabolism in Arabidopsis roots. Plant J. 2004, 38, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Z.; Guo, C.; Wang, Y.C.; Ouyang, F. Effect of light irradiation on hairy root growth and artemisinin biosynthesis of Artemisia annua L. Process Biochem. 2002, 38, 581–585. [Google Scholar] [CrossRef]
- Xie, B.D.; Wang, H.T. Effects of light spectrum and photoperiod on contents of flavonoid and terpene in leaves of Ginkgo biloba L. Nanjing For. Univ. 2006, 30, 51–54. [Google Scholar]
- Rizzalli, R.H.; Villalobos, F.J.; Orgaz, F. Radiation interception, radiation-use efficiency and dry matter partitioning in garlic (Allium sativum L.). Eur. J. Agron. 2002, 18, 33–43. [Google Scholar] [CrossRef]
- Orcutt, D.M.; Nilsen, E.T. The Physiology of Plants under Stress: Soil and Biotic Factors; John Wiley & Sons: New York, NY, USA, 2000. [Google Scholar]
- Mohammadi, B.; Khodadadi, M.; Karami, E.; Shaaf, S. Variation in agro-morphological characters in Iranian garlic landraces. Int. J. Veg. Sci. 2014, 20, 202–215. [Google Scholar] [CrossRef]
- Gonzalez, R.E.; Soto, V.C.; Sance, M.M.; Camargo, A.B.; Galmarini, C.R. Variability of solids, organosulfur compounds, pungency and health-enhancing traits in garlic (Allium sativum L.) cultivars belonging to different ecophysiological groups. J. Agric. Food Chem. 2009, 57, 10282–10288. [Google Scholar] [CrossRef]
- Singh, L.; Kaul, V.; Gohil, R.N. Analysis of morphological variability in the Indian germplasm of Allium sativum L. Plant Syst. Evol. 2014, 300, 245–254. [Google Scholar] [CrossRef]
- Mann, L.K. Anatomy of the garlic bulb and factors affecting bulb development. Hilgardia 1952, 21, 195–251. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Fei, M.L.I.; Tong, L.I.; Wei, L.I.; De Yang, L. Changes in antioxidant capacity, levels of soluble sugar, total polyphenol, organosulfur compound and constituents in garlic clove during storage. Ind. Crops Prod. 2015, 69, 137–142. [Google Scholar] [CrossRef]
- McCready, R.M.; Guggolz, J.; Silviera, V.; Owens, H.S. Determination of starch and amylase in vegetables. Anal. Chem. 1950, 22, 1156–1158. [Google Scholar] [CrossRef]
- Miller, G.N. Use of dinitrosalicylic acid reagent for determination of reducing sugars. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Handel, E.V. Direct micro determination of sucrose. Anal. Biochem. 1968, 22, 280–283. [Google Scholar] [CrossRef]
- Ashwell, G. Methods in Enzymology. In Methods in Enzymol; Colowick, S.J., Kaplan, N.O., Eds.; Academic Press: New York, NY, USA, 1957; Volume 3, p. 75. [Google Scholar]
- Singleton, V.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Yong, S.P.; Soon, T.J.; Seong, G.K.; Buk, G.H.; Patricia, A.A.; Fernando, T. Antioxidants and proteins in ethylene-treated kiwifruits. Food Chem. 2008, 107, 640–648. [Google Scholar]
- Rabinowitch, H.D. Onions and other edible Alliums. In Handbook of Flowering; Halevy, A.H., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 398–409. [Google Scholar]
- Rabinowitch, H.D. Physiology of flowering. In Onions and Allied Crops, I: Botany, Physiology and Genetics; Rabinowitch, H.D., Brewster, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 398–409. [Google Scholar]
- Kamenetsky, R.; Rabinowitch, H.D. Florogenesis. In Allium Crop Science: Recent Advances; Rabinowitch, H.D., Currah, L., Eds.; CAB International: Wallingford, UK, 2002; pp. 31–57. [Google Scholar]
- Khokhar, K.M.; Hadley, P.; Pearson, S. Effect of photoperiod and temperature on inflorescence appearance and subsequent development towards flowering in onion raised from sets. Sci. Hortic. 2007, 112, 9–15. [Google Scholar] [CrossRef]
- Brewster, J.L. Onions and other Vegetable Alliums, 2nd ed.; CABI Publishing: Wallingford, UK, 2008; p. 432. [Google Scholar]
- Dong, Y.; Cheng, Z.; Meng, H.; Liu, H.; Wu, C.; Khan, A.R. The effect of cultivar, sowing date and transplant location in field on bolting of welsh onion (Allium fistulosum L.). BMC Plant Biol. 2013, 13, 154. [Google Scholar] [CrossRef] [Green Version]
- Benschop, M.; Kamenetsky, R.; Le Nard, M.; Okubo, H.; De Hertogh, A. The global flower bulb industry: Production, utilization, and research. Hortic. Rev. 2010, 36, 1–115. [Google Scholar]
- Wu, C.; Wang, M.; Dong, Y.; Cheng, Z.; Meng, H. Effect of plant age and vernalization on bolting, plant growth and enzyme activity of garlic (Allium sativum L.). Sci. Hortic. 2016, 201, 295–305. [Google Scholar] [CrossRef]
- Brewster, J.L. The physiology of the onion. Hortic. Abstr. 1977, 47, 17–23. [Google Scholar]
- Burba, J.L.; Riera, P.G. Characterization, adaptation and selection of garlic germplasm (Allium sativum L.) through the management of dormancy in Mendoza, Argentina. First International Symposium on Edible Alliaceae. Acta Hortic. 1997, 433, 151–164. [Google Scholar]
- Panthee, D.R.; KC, R.B.; Regmi, H.N.; Subedi, P.P.; Bhattarai, S.; Dhakal, J. Diversity analysis of garlic (Allium sativum L.) germplasms available in Nepal based on morphological characters. Genet. Resour. Crop Evol. 2006, 53, 205–212. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Gibson, S. ABA and sugar interactions regulating development: Cross-talk or voices in a crowd? Curr. Opin. Plant Biol. 2001, 5, 26–32. [Google Scholar] [CrossRef]
- Zheng, J.; Yang, B.; Toumasjukka, S.; Ou, S.; Kallio, H. Effects of latitude and weather conditions on contents of sugars, fruit acids, and ascorbic acid in black currant (Ribes nigrum L.) juice. J. Agric. Food Chem. 2009, 57, 2977–2987. [Google Scholar] [CrossRef]
- Zheng, J.; Kallio, H.; Yang, B. Effects of latitude and weather conditions on sugars, fruit acids and ascorbic acid in currant (Ribes sp.) cultivars. J. Sci. Food Agric. 2009, 89, 2011–2023. [Google Scholar] [CrossRef]
- Zheng, J.; Yang, B.; Trepanier, M.; Kallio, H. Effects of genotype, latitude, and weather conditions on the composition of sugars, sugar alcohols, fruit acids, and ascorbic acid in sea buckthorn (Hippophaerhamnoides ssp. mongolica) berry juice. J. Agric. Food Chem. 2012, 60, 3180–3189. [Google Scholar] [CrossRef]
- Rosa, E.; David, M.; Gomes, M. Glucose, fructose and sucrose content in broccoli, white cabbage and Portuguese cabbage grown in early and late seasons. J. Sci. Food Agric. 2001, 81, 1145–1149. [Google Scholar] [CrossRef]
- Beato, V.M.; Orgaz, F.; Mansilla, F.; Montano, A. Changes in phenolic compounds in garlic (Allium sativum L.) owing to the cultivar and location of growth. Plant Foods Hum. Nutr. 2011, 66, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Younis, M.E.; Hasaneen, M.N.A.; Abdel-Aziz, H.M.M. An enhancing effect of visible light and UV radiation on phenolic compounds and various antioxidants in broad bean seedlings. Plant Signal. Behav. 2010, 5, 1197–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swieca, M.; Gawlik-Dziki, U.; Kowalczyk, D.; Zlotek, U. Impact of germination time and type of illumination on the antioxidant compounds and antioxidant capacity of Lens culinaris sprouts. Sci. Hortic. 2012, 140, 87–95. [Google Scholar] [CrossRef]
- Lanzotti, V.; Scala, F.; Bonanomi, G. Compounds from Allium species with cytotoxic and antimicrobial activity. Phytochem. Rev. 2014, 13, 769–791. [Google Scholar] [CrossRef]
- Volk, G.M.; Stern, D. Phenotypic characteristics of ten garlic cultivars grown at different North American locations. HortScience 2009, 44, 1238–1247. [Google Scholar] [CrossRef] [Green Version]
- Pardo, J.E.; Escribano, J.; Gomez, R.; Alvarruiz, A. Physical-chemical and sensory quality evaluation of garlic cultivars. J. Food Qual. 2007, 30, 609–622. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Plant Standing Height (cm) | Fresh Weight (g) | Pseudostem Diameter (mm) | Bulb Diameter (mm) | Bulb Weight (g) | Bulb Height (mm) | Growth Period (day) | Bulbing Index |
|---|---|---|---|---|---|---|---|---|
| Grand mean | 72.8 | 41.36 | 13.2 | 30.5 | 25.83 | 44.1 | 74.5 | 2.34 |
| Cultivar | ||||||||
| G103 | 76.7 a | 43.41 a | 14.5 a | 32.7 a | 28.08 a | 46.5 a | 73.8 b | 2.29 c |
| G024 | 68.9 c | 39.55 c | 11.8 c | 28.2 c | 24.03 c | 41.6 c | 73.0 c | 2.39 a |
| G2011-5 | 72.7 b | 41.11 b | 13.3 b | 30.6 b | 25.36 b | 44.1 b | 76.5 a | 2.33 a |
| Photoperiod (light/dark) | ||||||||
| 8/16 hr | 70.9 e | 40.34 d | 12.3 d | 24.2 e | 24.93 e | 42.8 d | 92.9 a | 1.98 d |
| 10/14hr | 72.1 d | 40.13 e | 12.6 c | 28.2 d | 25.72 c | 43.5 c | 82.1 b | 2.28 c |
| 12/12hr | 72.9 c | 41.40 c | 13.3 b | 32.5 c | 27.19 a | 44.0 b | 72.1 c | 2.48 a |
| 14/10hr | 74.3 a | 42.86 a | 13.9 a | 34.0 a | 25.82 b | 45.0 a | 59.1 e | 2.49 a |
| 16/8hr | 73.7 b | 42.05 b | 13.9 a | 33.5 b | 25.47 d | 45.0 a | 66.00 d | 2.44 b |
| Temperature (light/dark) | ||||||||
| 20/15 °C | 82.2 a | 38.02 c | 15.0 a | 28.3 c | 26.00 b | 41.00 c | 86.0 a | 1.89 c |
| 25/18 °C | 67.7 c | 39.10 b | 12.5 b | 30.7 b | 26.37 a | 41.5 b | 72.3 b | 2.45 b |
| 30/20 °C | 68.5 b | 46.95 a | 12.1 c | 32.4 a | 25.11 c | 49.75 a | 65.1 c | 2.67 a |
| F-test | ||||||||
| Cultivar (C) | *** | *** | *** | *** | *** | *** | *** | *** |
| Photoperiod (L) | *** | *** | *** | *** | *** | *** | *** | *** |
| Temperature (T) | *** | *** | *** | *** | *** | *** | *** | *** |
| C × L | *** | *** | *** | *** | *** | *** | *** | *** |
| C × T | *** | *** | *** | *** | *** | *** | *** | *** |
| L × T | *** | *** | *** | *** | *** | *** | *** | *** |
| C × L × T | * | *** | *** | *** | *** | *** | *** | *** |
| Treatment | G103 | G024 | G2011-5 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Photoperiod (light/dark) | Temperature (light/dark) | Plant Standing Height (cm) | Fresh Weight (g) | Pseudostem Diameter (mm) | Plant Standing Height (cm) | Fresh Weight (g) | Pseudostem Diameter (mm) | Plant Standing Height (cm) | Fresh Weight (g) | Pseudostem Diameter (mm) |
| 8/16 hr | 20/15 °C | 83.0 e | 38.26 m | 15.4 d | 76.0 c | 35.35 l | 12.3 d | 80.6 d | 36.54 n | 14.4 c |
| 25/18 °C | 68.2 l | 39.58 k | 12.8 i | 63.8 ef | 36.87 i | 10.2 h | 66.3 k | 38.47 i | 11.8 g | |
| 30/20 °C | 69.1 k | 47.50 e | 12.3 j | 63.4 f | 44.33 d | 9.9 i | 67.2 i | 46.19 c | 11.2 h | |
| 10/14 hr | 20/15 °C | 86.2 d | 38.81 l | 15.5 d | 77.2 bc | 35.93 k | 13.1 c | 80.9 d | 36.90 m | 14.3 c |
| 25/18 °C | 71.1 j | 39.91 jk | 12.9 i | 63.3 f | 36.31 j | 10.9 g | 66.8 j | 37.21 m | 14.3 g | |
| 30/20 °C | 71.9 i | 47.91 d | 12.7 i | 64.3 ef | 43.58 e | 10.4 h | 67.5 hi | 44.62 e | 11.7 g | |
| 12/12 hr | 20/15 °C | 87.3 c | 40.24 j | 16.6 c | 77.5 bc | 36.35 j | 13.5 b | 82.0 c | 37.92 k | 15.4 b |
| 25/18 °C | 72.0 i | 41.21 h | 13.8 g | 64.0 ef | 37.82 g | 11.3 f | 67.7 h | 38.21 j | 12.8 e | |
| 30/20 °C | 72.6 h | 49.58 c | 13.3 h | 64.8 def | 45.41 b | 10.9 g | 68.4 g | 45.81 d | 12.3 f | |
| 14/10 hr | 20/15 °C | 88.6 a | 41.22 h | 17.3 b | 79.6 a | 37.22 h | 14.4 a | 83.8 a | 39.22 h | 15.9 a |
| 25/18 °C | 73.2 g | 42.62 f | 14.4 f | 64.9 def | 38.67 f | 11.9 e | 68.9 f | 40.51 f | 13.3 d | |
| 30/20 °C | 73.8 f | 51.31 a | 13.9 g | 66.4 d | 46.38 a | 11.3 f | 69.8 e | 48.62 a | 12.8 e | |
| 16/8 hr | 20/15 °C | 88.1 b | 40.83 i | 17.6 a | 78.4 ab | 36.89 i | 14.3 a | 83.0 b | 38.63 i | 15.5 b |
| 25/18 °C | 72.5 h | 41.90 g | 14.7 e | 64.6 ef | 37.30 h | 11.9 e | 68.5 g | 39.90 g | 12.9 e | |
| 30/20 °C | 73.4 g | 50.32 b | 13.9 g | 65.4 de | 44.81 c | 11.5 f | 69.2 f | 47.88 b | 12.9 e | |
| Treatment | G103 | G024 | G2011-5 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Photoperiod (h) (light/dark) | Temperature (°C) (light/dark) | Bulb Diameter (mm) | Bulb Weight (g) | Bulb Height (mm) | Growth Period (day) | Bulb Diameter (mm) | Bulb Weight (g) | Bulb Height (mm) | Growth Period (day) | Bulb Diameter (mm) | Bulb Weight (g) | Bulb Height (mm) | Growth Period (day) |
| 8/16 | 20/15 | 25.3 m | 27.91 ef | 42.0 l | 109.5 a | 19.2 l | 23.93 fg | 37.6 l | 103.9 a | 22.6 m | 25.91 e | 39.8 l | 99.2 a |
| 25/18 | 27.4 l | 28.43 cd | 42.5 k | 76.3 ef | 20.8 k | 24.28 ef | 38.1 jk | 100.0 b | 24.4 l | 26.26 d | 40.3 k | 97.9 a | |
| 30/20 | 28.7 k | 23.72 i | 51.0 d | 76.6 ef | 24.0 i | 21.55 l | 45.7 d | 84.1 cd | 25.6 k | 22.36 k | 48.3 e | 89.1 b | |
| 10/14 | 20/15 | 28.9 k | 27.73 g | 42.8 j | 100.2 b | 21.8 j | 23.69 gh | 38.3 j | 86.3 c | 26.9 j | 25.71 ef | 40.5 k | 90.9 b |
| 25/18 | 31.2 j | 28.19 fg | 43.5 h | 82.9 cd | 25.8 h | 23.92 g | 38.0 k | 76.8 f | 29.1 i | 25.93 e | 41.0 j | 83.2 c | |
| 30/20 | 32.7 h | 33.78 a | 52.2 c | 79.0 de | 27.1 g | 20.53 m | 45.6 d | 65.8 h | 30.5 h | 21.97 l | 49.2 d | 74.2 d | |
| 12/12 | 20/15 | 32.3 i | 27.29 g | 43.1 i | 84.2 c | 28.2 f | 23.24 ij | 38.7 i | 80.4 ef | 30.5 h | 25.22 g | 40.9 j | 88.9 b |
| 25/18 | 34.9 f | 27.53 fg | 43.7 gh | 69.2 gh | 31.0 d | 23.52 hi | 39.1 h | 63.7 h | 33.0 e | 25.61 f | 41.3 i | 74.8 d | |
| 30/20 | 36.6 c | 33.36 a | 52.5 b | 68.0 h | 32.0 c | 28.21 a | 46.9 c | 53.5 j | 34.6 c | 30.71 a | 49.6 c | 66.3 ef | |
| 14/10 | 20/15 | 33.2 g | 28.30 cde | 43.9 g | 72.5 fg | 30.2 de | 24.33 e | 39.4 g | 70.5 g | 31.9 f | 26.80 b | 41.7 h | 68.7 e |
| 25/18 | 35.9 d | 29.41 b | 44.5 e | 50.0 j | 32.6 b | 27.41 b | 40.4 e | 58.3 i | 34.4 c | 24.64 h | 42.2 g | 60.2 g | |
| 30/20 | 37.7 a | 24.89 h | 53.4 a | 49.5 j | 34.2 a | 22.99 jk | 48.6 a | 48.8 k | 36.2 a | 23.63 i | 51.0 a | 53.6 h | |
| 16/8 | 20/15 | 32.9 gh | 28.70 c | 44.2 f | 77.6 e | 29.9 e | 24.91 d | 39.8 f | 81.4 de | 30.9 g | 26.32 cd | 42.0 g | 76.4 d |
| 25/18 | 35.6 e | 28.54 cd | 44.7 e | 58.3 i | 32.3 bc | 25.34 c | 40.0 f | 66.3 h | 33.4 d | 26.54 c | 42.5 f | 65.8 f | |
| 30/20 | 37.4 b | 23.46 i | 53.6 a | 53.8 j | 33.9 a | 22.68 k | 48.0 b | 55.8 ij | 35.0 b | 22.78 j | 50.6 b | 58.4 g | |
| Treatment | TSS (%) | Soluble Protein (mg g−1) | Soluble Sugar (%) | Total Sugar (mg g−1) | Glucose (%) | Sucrose (mg g−1) | Fructose (%) | Starch (mg g−1) | Total Phenol (mg g−1) | Total Flavonoid (mg g−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| Grand mean | 19.33 | 7.62 | 26.28 | 9.97 | 49.53 | 29.03 | 34.90 | 18.28 | 18.88 | 1.41 |
| Cultivar | ||||||||||
| G103 | 22.50 a | 7.91 a | 27.45 a | 14.94 a | 61.21 a | 37.09 a | 47.21 a | 23.52 a | 24.66 a | 1.82 a |
| G024 | 16.79 c | 7.31 c | 25.14 c | 6.96 c | 40.82 c | 23.57 c | 24.51 c | 14.74 c | 13.82 c | 0.97 c |
| G2011-5 | 18.70 b | 7.63 b | 26.23 b | 8.80 b | 46.55 b | 26.42 b | 32.98 b | 16.57 b | 18.16 b | 1.45 b |
| Photoperiod (light/dark) | ||||||||||
| 8/16 hr | 17.19 d | 3.54 e | 11.16 e | 6.62 e | 38.42 e | 22.43 e | 20.78 e | 14.23 e | 14.25 e | 1.11 e |
| 10/14 hr | 17.85 c | 7.11 d | 24.64 d | 7.78 d | 43.54 d | 24.90 d | 26.56 d | 15.51 d | 16.18 d | 1.21 d |
| 12/12 hr | 19.61 b | 8.41 c | 28.82 c | 10.41 c | 52.02 b | 28.03 c | 37.81 c | 19.14 c | 18.68 c | 1.35 c |
| 14/10 hr | 22.42 a | 10.00 a | 35.93 a | 14.27 a | 62.59 a | 36.16 a | 46.27 a | 22.24 a | 25.32 a | 1.83 a |
| 16/8 hr | 19.58 b | 9.02 b | 30.82 b | 10.75 b | 51.07 c | 33.61 b | 43.08 b | 20.26 b | 19.96 b | 1.56 b |
| Temperature (light/dark) | ||||||||||
| 20/15 °C | 18.28 c | 6.39 c | 23.03 c | 7.90 c | 43.80 c | 25.02 b | 26.27 c | 15.44 c | 15.63 c | 1.35 c |
| 25/18 °C | 19.54 b | 7.86 b | 27.20 b | 9.76 b | 49.75 b | 30.98 a | 34.66 b | 18.38 b | 17.57 b | 1.40 b |
| 30/20 °C | 20.18 a | 8.61 a | 28.60 a | 12.23 a | 55.03 a | 31.07 a | 43.77 a | 21.00 a | 23.44 a | 1.47 a |
| F-test | ||||||||||
| Cultivar (C) | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Photoperiod (L) | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Temperature (T) | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| C × L | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| C × T | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| L × T | *** | *** | *** | *** | *** | *** | *** | *** | *** | * |
| C × L × T | *** | *** | *** | *** | *** | *** | *** | *** | *** | ns |
| Index | PD | BD | BW | BH | GP | BI | TSS | SPC | SSC | TSC | GC | SC | FC | SC | TP | TF |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PD | 1 | |||||||||||||||
| BD | 0.276 ** | 1 | ||||||||||||||
| BW | 0.415 *** | 0.325 *** | 1 | |||||||||||||
| BH | −0.089 | 0.597 *** | 0.121 | 1 | ||||||||||||
| GP | 0.128 | −0.807 *** | 0.021 | −0.514 *** | 1 | |||||||||||
| BI | −0.513 *** | 0.673 *** | −0.031 | 0.586 *** | −0.823 *** | 1 | ||||||||||
| TSS | 0.403 *** | 0.716 *** | 0.437 *** | 0.545 *** | −0.512 *** | 0.288 *** | 1 | |||||||||
| SPC | 0.098 | 0.879 *** | 0.163 | 0.448 *** | −0.814 *** | 0.721 *** | 0.440 *** | 1 | ||||||||
| SSC | 0.216 * | 0.865 *** | 0.094 | 0.333 *** | −0.806 *** | 0.616 *** | 0.571 *** | 0.357 *** | 1 | |||||||
| TSC | 0.307 *** | 0.814 *** | 0.399 *** | 0.694 *** | −0.633 *** | 0.463 *** | 0.557 *** | 0.260 *** | 0.823 *** | 1 | ||||||
| GC | 0.325 *** | 0.822 *** | 0.413 *** | 0.629 *** | −0.629 *** | 0.464 *** | 0.650 *** | 0.584 *** | 0.700 *** | 0.628 *** | 1 | |||||
| SC | 0.298 *** | 0.801 *** | 0.383 *** | 0.596 *** | −0.666 *** | 0.464 *** | 0.748 *** | 0.542 *** | 0.661 *** | 0.743 *** | 0.767 *** | 1 | ||||
| FC | 0.246 ** | 0.863 *** | 0.357 *** | 0.705 *** | −0.669 *** | 0.555 *** | 0.824 *** | 0.534 *** | 0.712 *** | 0.670 *** | 0.824 *** | 0.776 *** | 1 | |||
| SC | 0.283 *** | 0.816 *** | 0.397 *** | 0.686 *** | −0.641 *** | 0.485 *** | 0.853 *** | 0.501 *** | 0.639 *** | 0.651 *** | 0.718 *** | 0.753 *** | 0.816 *** | 1 | ||
| TP | 0.275 ** | 0.774 *** | 0.370 *** | 0.772 *** | −0.588 *** | 0.451 *** | 0.888 *** | 0.386 *** | 0.540 *** | 0.508 *** | 0.671 *** | 0.705 *** | 0.759 *** | 0.840 *** | 1 | |
| TF | 0.593 *** | 0.766 *** | 0.441 *** | 0.539 *** | −0.469 *** | 0.197 * | 0.828 *** | 0.590 *** | 0.506 *** | 0.423 *** | 0.683 *** | 0.741 *** | 0.767 *** | 0.767 *** | 0.824 *** | 1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atif, M.J.; Amin, B.; Ghani, M.I.; Hayat, S.; Ali, M.; Zhang, Y.; Cheng, Z. Influence of Different Photoperiod and Temperature Regimes on Growth and Bulb Quality of Garlic (Allium sativum L.) Cultivars. Agronomy 2019, 9, 879. https://doi.org/10.3390/agronomy9120879
Atif MJ, Amin B, Ghani MI, Hayat S, Ali M, Zhang Y, Cheng Z. Influence of Different Photoperiod and Temperature Regimes on Growth and Bulb Quality of Garlic (Allium sativum L.) Cultivars. Agronomy. 2019; 9(12):879. https://doi.org/10.3390/agronomy9120879
Chicago/Turabian StyleAtif, Muhammad Jawaad, Bakht Amin, Muhammad Imran Ghani, Sikandar Hayat, Muhammad Ali, Yumeng Zhang, and Zhihui Cheng. 2019. "Influence of Different Photoperiod and Temperature Regimes on Growth and Bulb Quality of Garlic (Allium sativum L.) Cultivars" Agronomy 9, no. 12: 879. https://doi.org/10.3390/agronomy9120879