Blooming under Mediterranean Climate: Estimating Cultivar-Specific Chill and Heat Requirements of Almond and Apple Trees Using a Statistical Approach

 ,
,
Abstract
:
1. Introduction
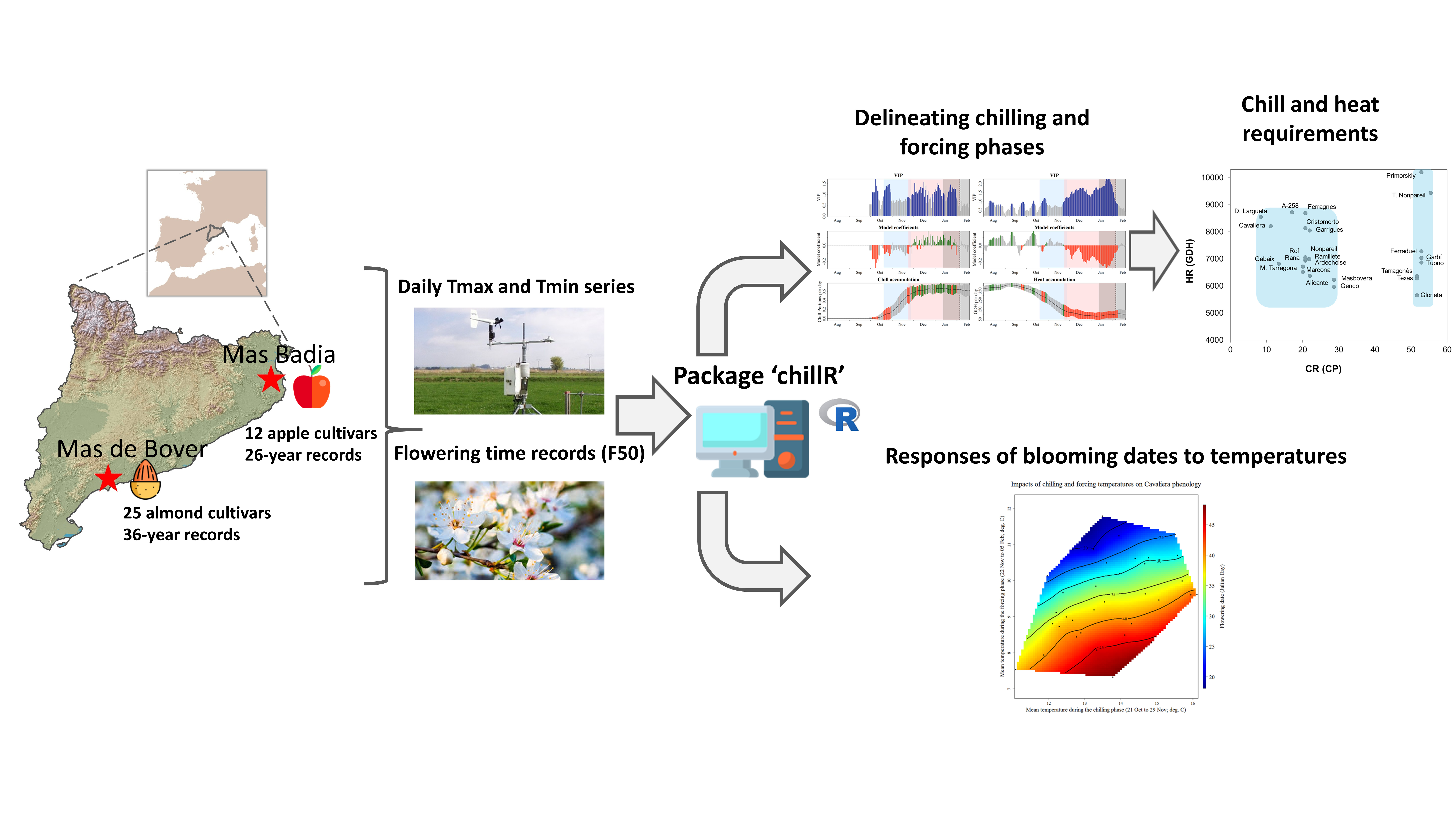
2. Materials and Methods
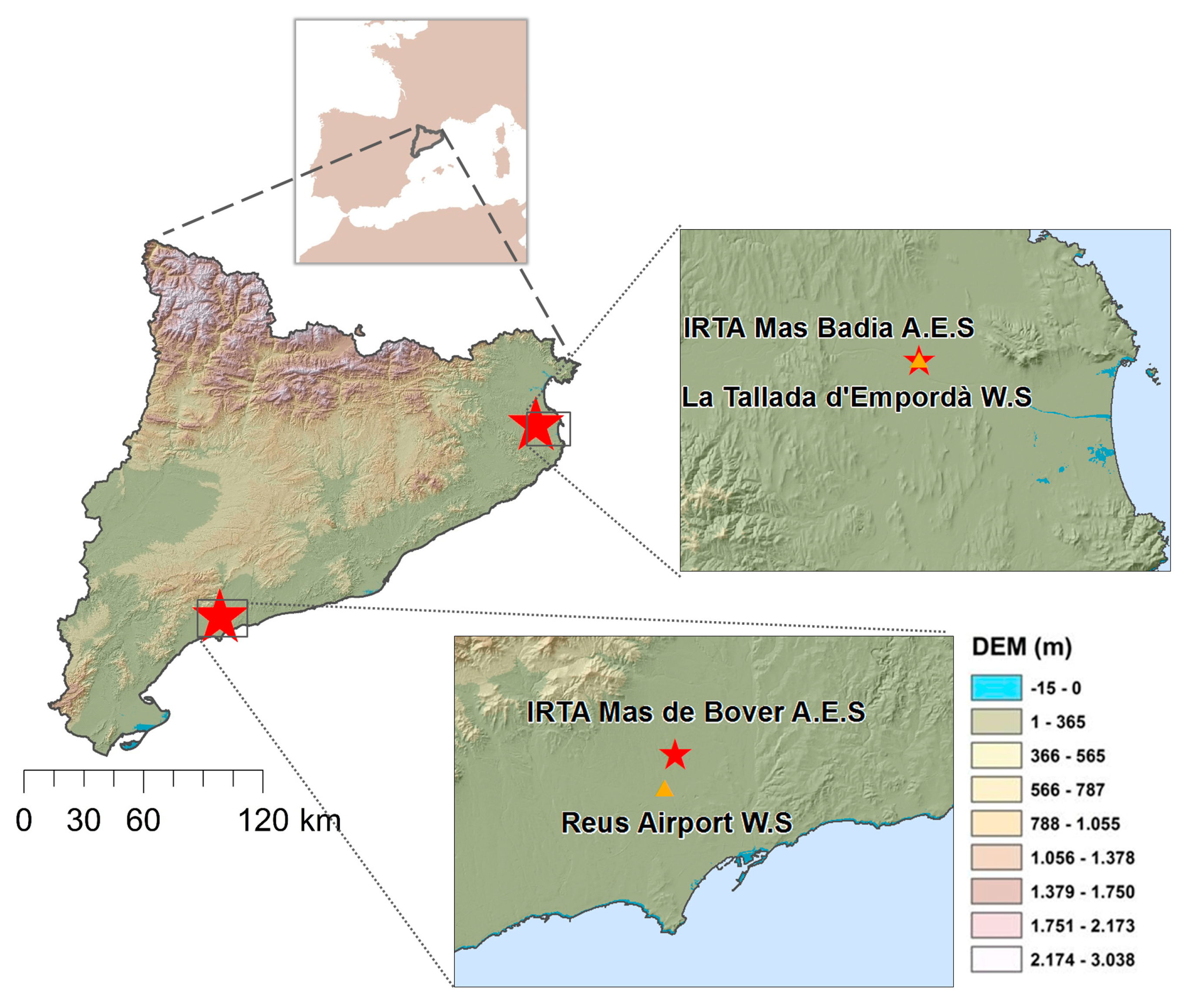
2.1. Study Area
2.2. Phenological Records
2.3. Climate Records
2.4. Temperature and Blooming Time Historical Trends
2.5. Estimating Chill and Heat Accumulation
2.6. Delineating Chilling and Forcing Phases: Cultivar-Specific Chill and Heat Requirements Estimation
2.7. Impacts of Chilling and Forcing Temperatures on Blooming Date
3. Results
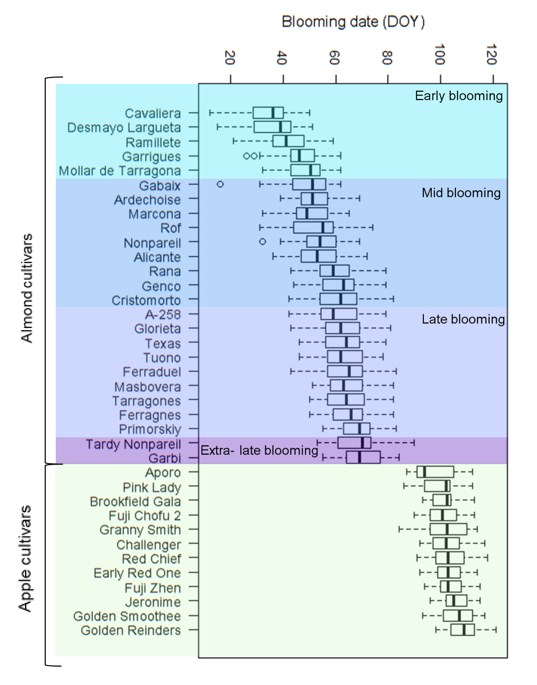
3.1. Temperature and Blooming Past Trends
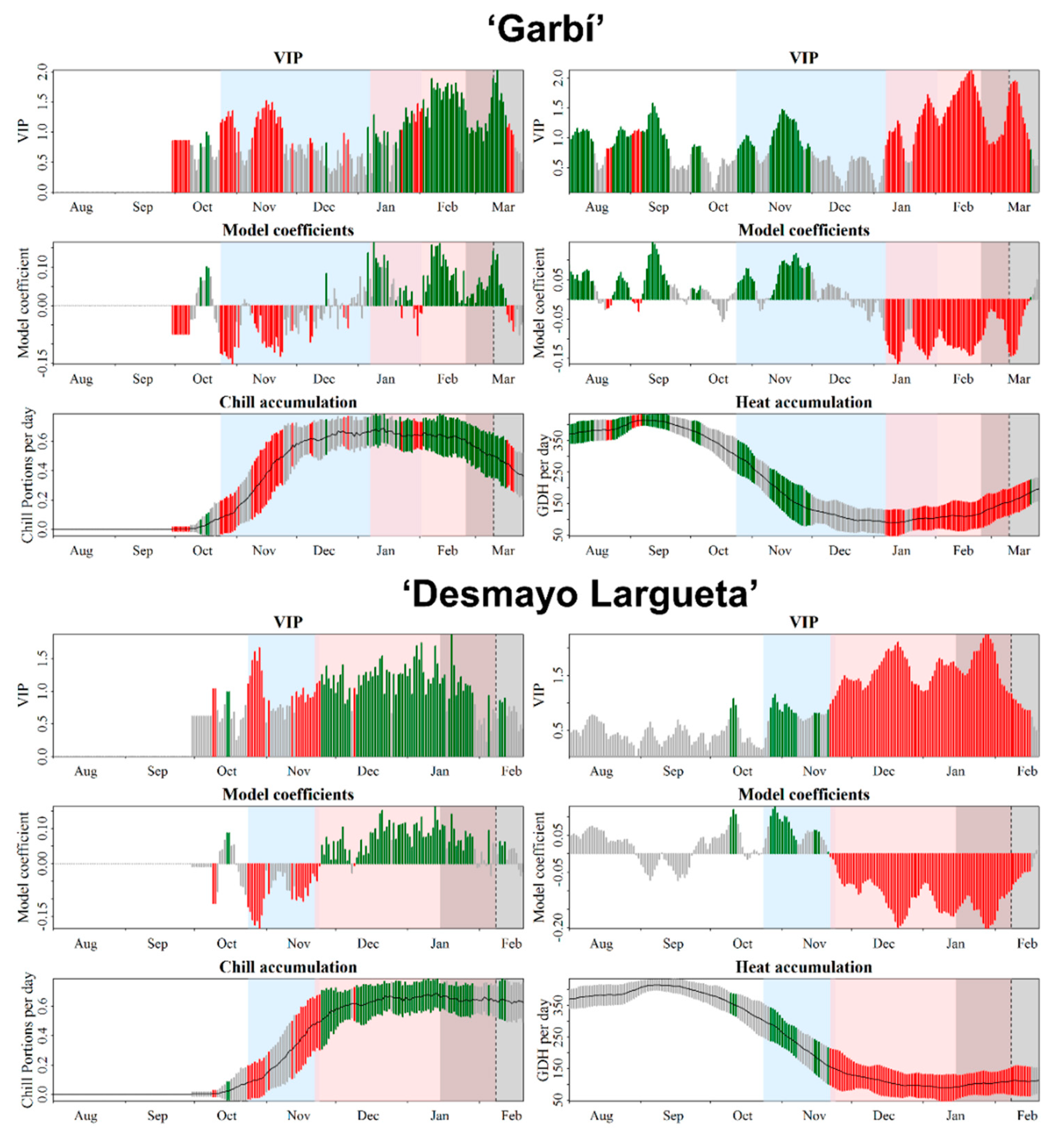
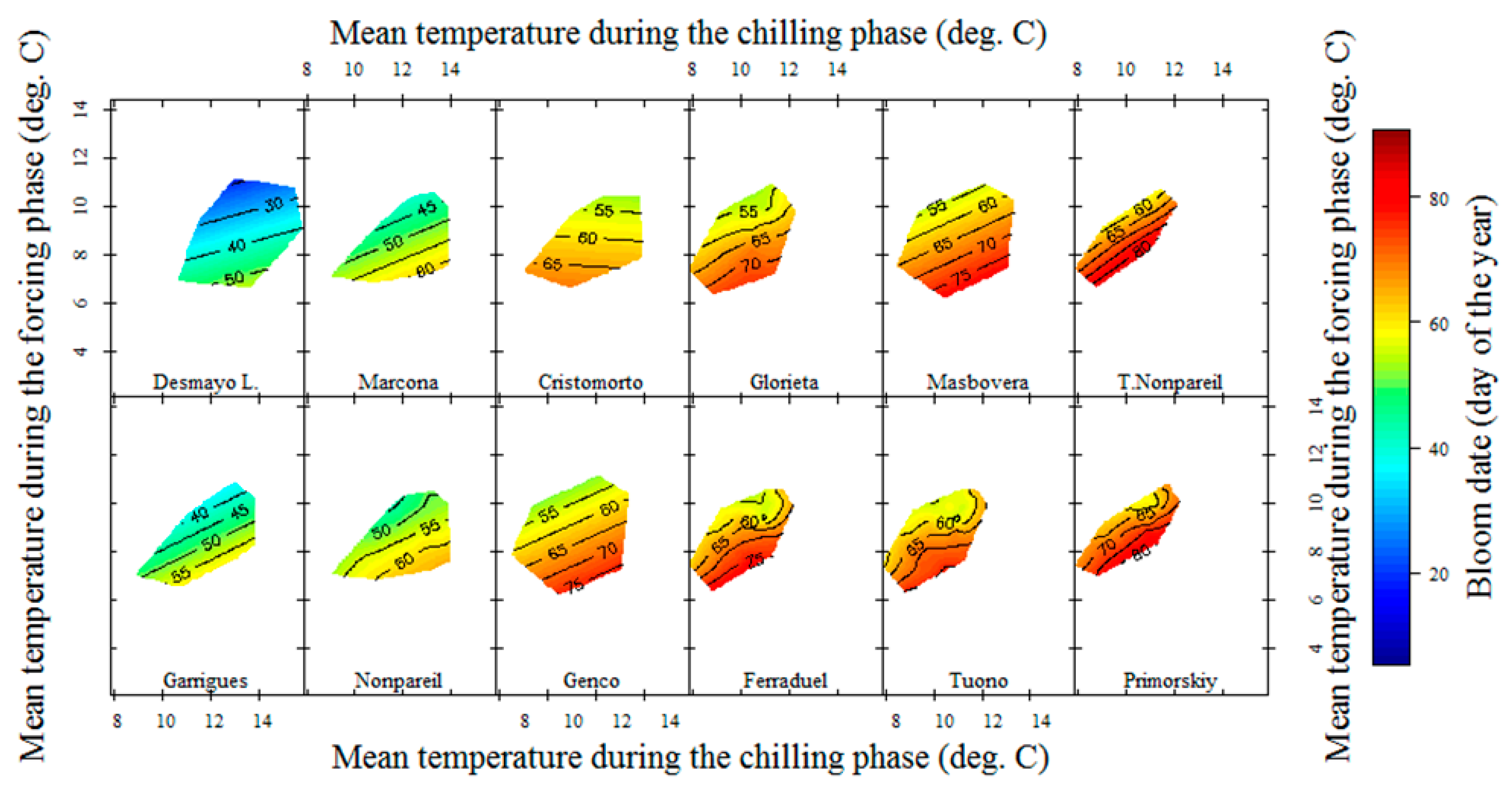
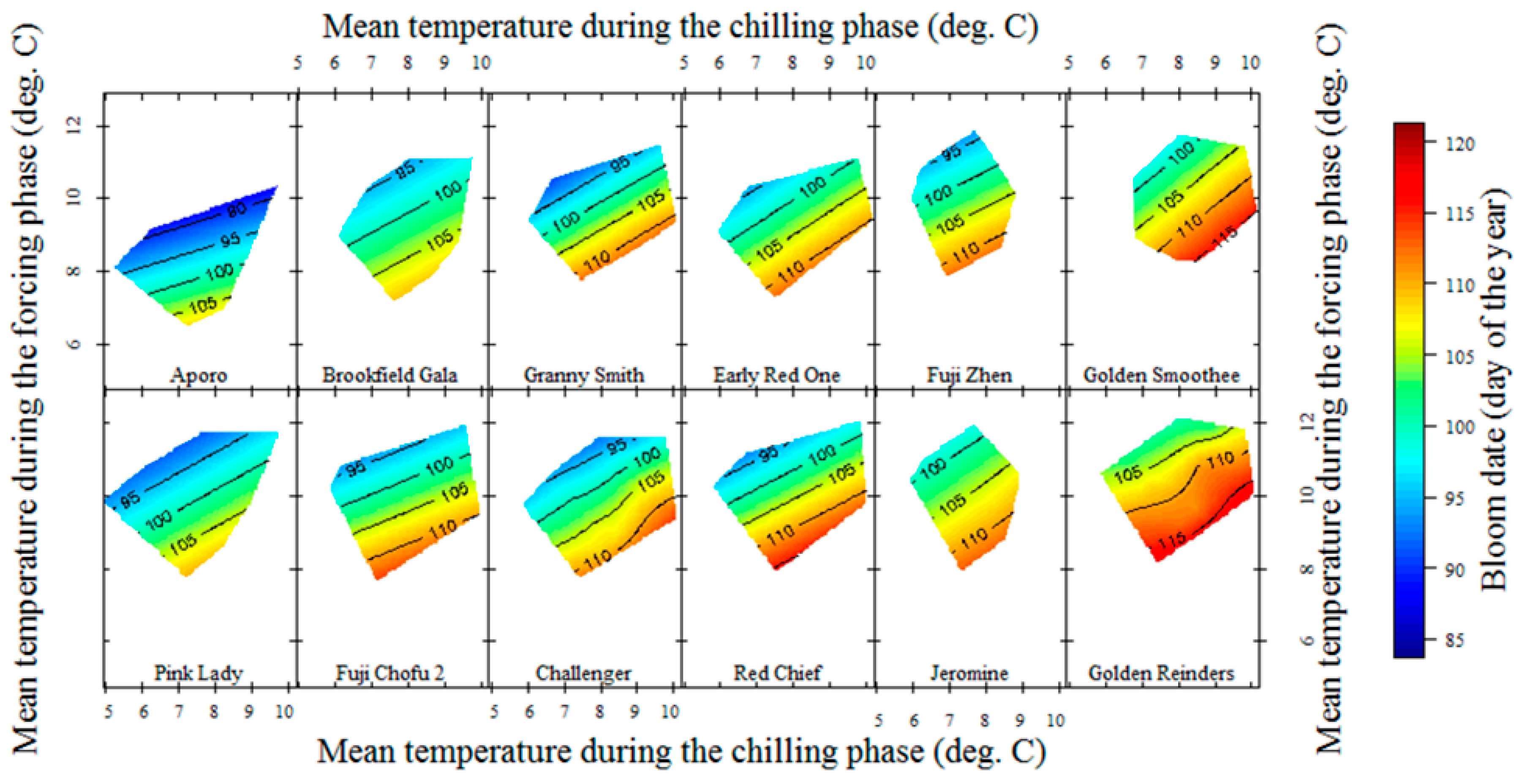
3.2. Chilling and Forcing Phases Delineation
3.2.1. Almond Cultivars
3.2.2. Apple Cultivars
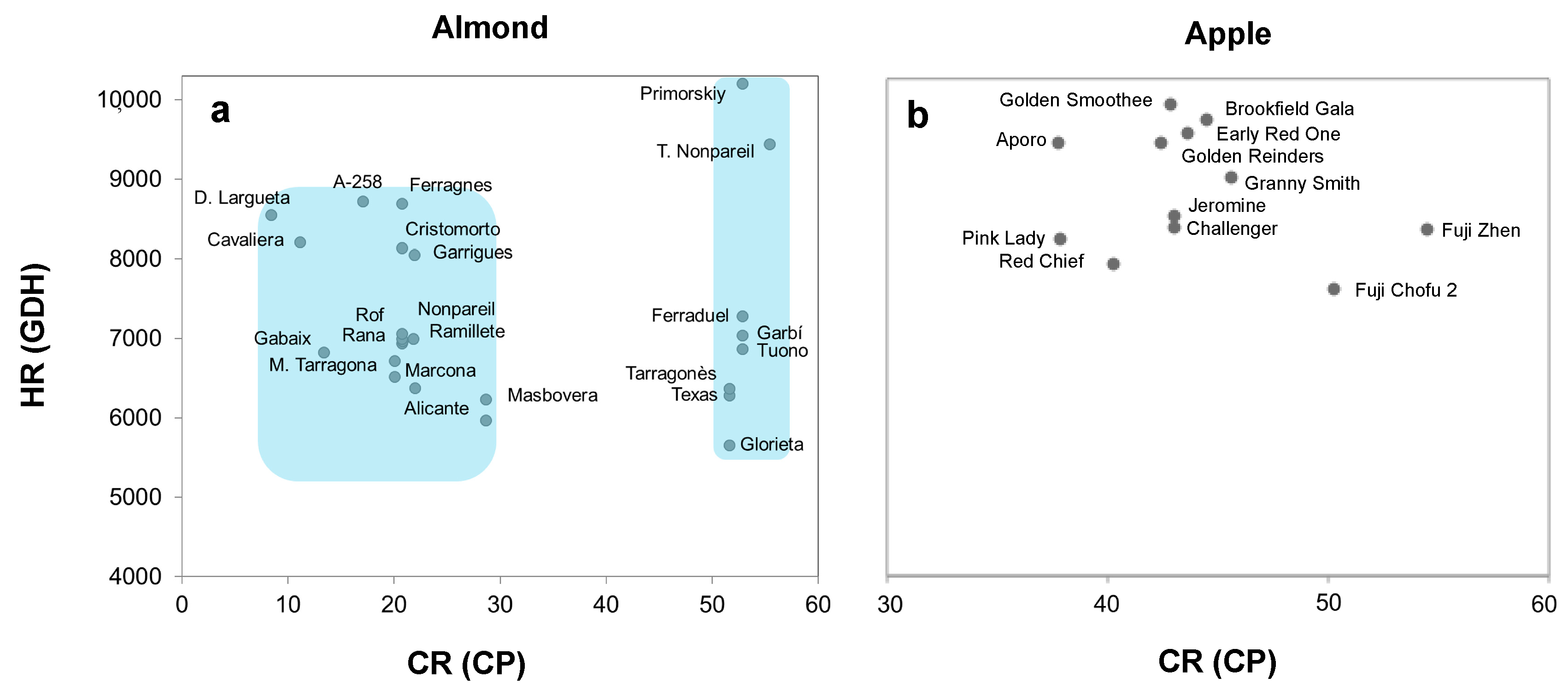
3.3. Chill and Heat Requirements
3.3.1. Almond Cultivars
3.3.2. Apple Cultivars
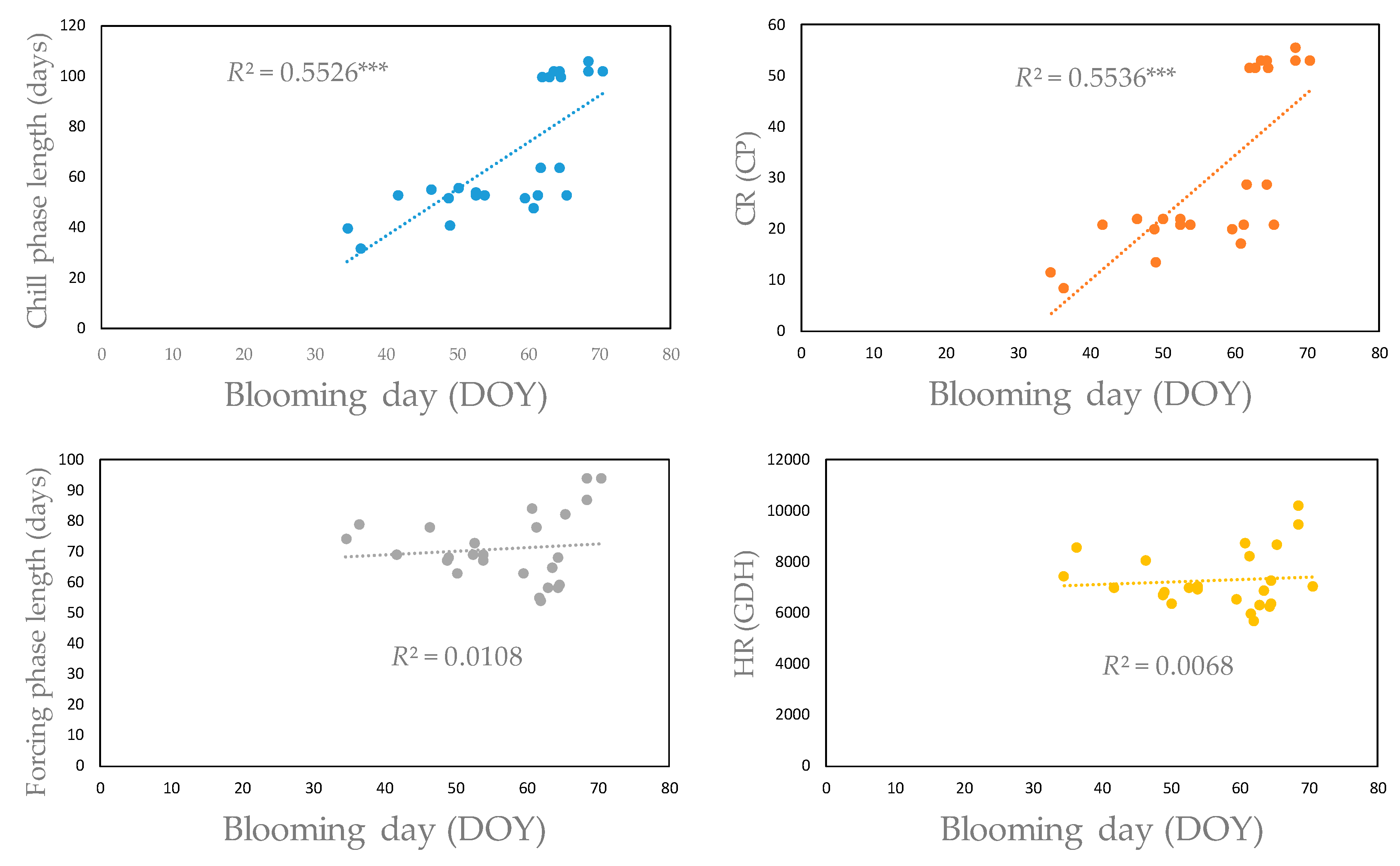
3.4. Impacts of Chilling and Forcing Temperatures on Blooming Dates
3.4.1. Almond Cultivars
3.4.2. Apple Cultivars
4. Discussion
4.1. Chilling and Forcing Phases Delineation Using a Partial Least Squares (PLS) Approach and Its Limitations
4.2. Chill and Heat Accumulation
4.2.1. Chill and Heat Requirements for Almond
4.2.2. Chill and Heat Requirements for Apple
4.3. Past Blooming Trends and Impacts of Chilling and Forcing Temperatures on Blooming Dates
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Faust, M.; Erez, A.; Rowland, L.J.; Wang, S.Y.; Norman, H.A. Bud dormancy in perennial fruit trees: Physiological basis for dormancy induction, maintenance, and release. Hortscience 1997, 32, 623–629. [Google Scholar] [CrossRef]
- Luedeling, E.; Kunz, A.; Blanke, M.M. Identification of chilling and heat requirements of cherry trees-a statistical approach. Int. J. Biometeorol. 2013, 57, 679–689. [Google Scholar] [CrossRef]
- Campoy, J.A.; Ruiz, D.; Egea, J. Dormancy in temperate fruit trees in a global warming context: A review. Sci. Hortic. 2011, 130, 357–372. [Google Scholar] [CrossRef]
- Sanchez-Perez, R.; Del Cueto, J.; Dicenta, F.; Martinez-Gomez, P. Recent advancements to study flowering time in almond and other Prunus species. Front. Plant Sci. 2014, 5, 334. [Google Scholar] [CrossRef] [PubMed]
- Erez, A. Bud dormancy; phenomenon, problems and solutions in the tropics and subtropics. In Temperate Fruit Crops in Warm Climates; Springer: Dordrecht, The Netherlands, 2000; pp. 17–48. [Google Scholar]
- Harrington, C.A.; Gould, P.J.; St Clair, J.B. Modeling the effects of winter environment on dormancy release of Douglas-fir. For. Ecol. Manag. 2010, 259, 798–808. [Google Scholar] [CrossRef]
- Kovaleski, A.P.; Reisch, B.I.; Londo, J.P. Deacclimation kinetics as a quantitative phenotype for delineating the dormancy transition and thermal efficiency for budbreak in Vitis species. Aob Plants 2018, 10, ply066. [Google Scholar] [CrossRef] [PubMed]
- Dantec, C.F.; Vitasse, Y.; Bonhomme, M.; Louvet, J.-M.; Kremer, A.; Delzon, S. Chilling and heat requirements for leaf unfolding in European beech and sessile oak populations at the southern limit of their distribution range. Int. J. Biometeorol. 2014, 58, 1853–1864. [Google Scholar] [CrossRef] [PubMed]
- Savé, R.; de Herralde, F.; Aranda, X.; Pla, E.; Pascual, D.; Funes, I.; Biel, C. Potential changes in irrigation requirements and phenology of maize, apple trees and alfalfa under global change conditions in Fluvià watershed during XXIst century: Results from a modeling approximation to watershed-level water balance. Agric. Water Manag. 2012, 114, 78–87. [Google Scholar] [CrossRef]
- Lopez-Bustins, J.A.; Pla, E.; Nadal, M.; de Herralde, F.; Save, R. Global change and viticulture in the Mediterranean region: A case of study in north-eastern Spain. Span. J. Agric. Res. 2014, 12, 78–88. [Google Scholar] [CrossRef]
- Pope, K.S.; Dose, V.; Da Silva, D.; Brown, P.H.; Leslie, C.A.; Dejong, T.M. Detecting nonlinear response of spring phenology to climate change by Bayesian analysis. Glob. Chang. Biol. 2013, 19, 1518–1525. [Google Scholar] [CrossRef] [PubMed]
- Funes, I.; Aranda, X.; Biel, C.; Carbo, J.; Camps, F.; Molina, A.J.; de Herralde, F.; Grau, B.; Save, R. Future climate change impacts on apple flowering date in a Mediterranean subbasin. Agric. Water Manag. 2016, 164, 19–27. [Google Scholar] [CrossRef]
- Luedeling, E.; Zhang, M.H.; Girvetz, E.H. Climatic changes lead to declining winter chill for fruit and nut trees in California during 1950–2099. PLoS ONE 2009, 4, e6166. [Google Scholar] [CrossRef]
- Luedeling, E.; Girvetz, E.H.; Semenov, M.A.; Brown, P.H. Climate change affects winter chill for temperate fruit and nut trees. PLoS ONE 2011, 6, e20155. [Google Scholar] [CrossRef]
- El Yaacoubi, A.; Malagi, G.; Oukabli, A.; Citadin, I.; Hafidi, M.; Bonhomme, M.; Legave, J.M. Differentiated dynamics of bud dormancy and growth in temperate fruit trees relating to bud phenology adaptation, the case of apple and almond trees. Int. J. Biometeorol. 2016, 60, 1695–1710. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Jacob, D.; Taylor, M.; Bindi, M.; Brown, S.; Camilloni, I.; Diedhiou, A.; Djalante, R.; Ebi, K.L.; Engelbrecht, F.; et al. 2018: Impacts of 1.5 °C Global Warming on Natural and Human Systems. In Global Warming of 1.5 °C; An IPCC Special Report on the impacts of global warming of 1.5 °C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty; Masson-Delmotte, V., Zhai, P., Pörtner, H.-O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Moufouma-Okia, W., Péan, C., Pidcock, R., et al., Eds.; 2019; in press. [Google Scholar]
- TICCC; Government of Catalonia; Institute of Catalan Studies. Climate Change in Catalonia: Executive Summary of the Third Report on Climate Change in Catalonia. 2016. Writing Team: Xavier Duran, M. Josep Picó and Lluís Reales. Edited by Arnau Queralt, Barcelona. Available online: http://cads.gencat.cat/ca/detalls/detallarticle/Tercer-informe-sobre-el-canvi-climatic-a-Catalunya-00003 (accessed on 14 November 2019).
- MAPA. Encuesta Sobre Superficies y Rendimientos de Cultivos. ESYRCE. Resultados Nacionales y Autonómicos. 2018. Ministerio de Agricultura, Pesca y Alimentación del Gobierno de España. Subsecretaría de Agricultura, Pesca y Alimentación. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/agricultura/esyrce/ (accessed on 13 February 2018).
- MAPA. Superficies y Producciones Anuales de Cultivo. Datos Avances de Frutales Cítricos y Frutales Secos año 2018. 2018. Ministerio de Agricultura, Pesca y Alimentación del Gobierno de España. Subsecretaría de Agricultura, Pesca y Alimentación. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/agricultura/superficies-producciones-anuales-cultivos/ (accessed on 20 February 2019).
- FAO. Statistical Data Base Food and Agricultural Organization, FAOSTAT. 2018. Available online: http://www.fao.org/faostat/es/#data (accessed on 22 March 2018).
- Fishman, S.; Erez, A.; Couvillon, G.A. The temperature-dependence of dormancy breaking in plants—Computer-simulation of processes studied under controlled temperatures. J. Theor. Biol. 1987, 126, 309–321. [Google Scholar] [CrossRef]
- Anderson, J.; Richardson, E.; Kesner, C. Validation of chill unit and flower bud phenology models for’Montmorency’sour cherry. Acta Horticulturae 1986, 184, 71–78. [Google Scholar] [CrossRef]
- Benmoussa, H.; Ghrab, M.; Ben Mimoun, M.; Luedeling, E. Chilling and heat requirements for local and foreign almond (Prunus dulcis Mill.) cultivars in a warm Mediterranean location based on 30 years of phenology records. Agric. For. Meteorol. 2017, 239, 34–46. [Google Scholar] [CrossRef]
- Benmoussa, H.; Luedeling, E.; Ghrab, M.; Ben Yahmed, J.; Ben Mimoun, M. Performance of pistachio (Pistacia vera L.) in warming Mediterranean orchards. Environ. Exp. Bot. 2017, 140, 76–85. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Hadley, P.; Ordidge, M.; Xu, X.; Luedeling, E. Delayed chilling appears to counteract flowering advances of apricot in southern UK. Agric. For. Meteorol. 2017, 237, 209–218. [Google Scholar] [CrossRef]
- Guo, L.; Dai, J.H.; Ranjitkar, S.; Yu, H.Y.; Xu, J.C.; Luedeling, E. Chilling and heat requirements for flowering in temperate fruit trees. Int. J. Biometeorol. 2014, 58, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Dai, J.; Wang, M.; Xu, J.; Luedeling, E. Responses of spring phenology in temperate zone trees to climate warming: A case study of apricot flowering in China. Agric. For. Meteorol. 2015, 201, 1–7. [Google Scholar] [CrossRef]
- Luedeling, E.; Gassner, A. Partial Least Squares Regression for analyzing walnut phenology in California. Agric. For. Meteorol. 2012, 158, 43–52. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Sakar, E.H.; El Yamani, M.; Boussakouran, A.; Rharrabti, Y. Codification and description of almond (Prunus dulcis) vegetative and reproductive phenology according to the extended BBCH scale. Sci. Hortic. 2019, 247, 224–234. [Google Scholar] [CrossRef]
- Felipe, A. Estados fenológicos del almendro. Inf. Tec. Econ. Agrar. 1977, 8, 8–9. [Google Scholar]
- SAS. Version 9.4; SAS Institute Inc.: Cary, NC, USA, 2014.
- Hamed, K.H.; Rao, A.R. A modified Mann-Kendall trend test for autocorrelated data. J. Hydrol. 1998, 204, 182–196. [Google Scholar] [CrossRef]
- Ramos, M.; Jones, G.; Martínez-Casasnovas, J. Structure and trends in climate parameters affecting winegrape production in northeast Spain. Clim. Res. 2008, 38, 1–15. [Google Scholar] [CrossRef]
- Fishman, S.; Erez, A.; Couvillon, G. The temperature dependence of dormancy breaking in plants: Mathematical analysis of a two-step model involving a cooperative transition. J. Theor. Biol. 1987, 124, 473–483. [Google Scholar] [CrossRef]
- Darbyshire, R.; Webb, L.; Goodwin, I.; Barlow, S. Winter chilling trends for deciduous fruit trees in Australia. Agric. For. Meteorol. 2011, 151, 1074–1085. [Google Scholar] [CrossRef]
- Luedeling, E. chillR. Statistical Methods for Phenology Analysis in Temperate Fruit Trees. R Package Version 0.70.2. 2018. Available online: https://CRAN.R-project.org/package=chillR (accessed on 27 June 2016).
- Legave, J.M.; Blanke, M.; Christen, D.; Giovannini, D.; Mathieu, V.; Oger, R. A comprehensive overview of the spatial and temporal variability of apple bud dormancy release and blooming phenology in Western Europe. Int. J. Biometeorol. 2013, 57, 317–331. [Google Scholar] [CrossRef]
- Alonso, J.M.; Anson, J.M.; Espiau, M.T.; Company, R.S.I. Determination of endodormancy break in almond flower buds by a correlation model using the average temperature of different day intervals and its application to the estimation of chill and heat requirements and blooming date. J. Am. Soc. Hortic. Sci. 2005, 130, 308–318. [Google Scholar] [CrossRef]
- Wold, S.; Sjostrom, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Luedeling, E.; Guo, L.; Dai, J.; Leslie, C.; Blanke, M.M. Differential responses of trees to temperature variation during the chilling and forcing phases. Agric. For. Meteorol. 2013, 181, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Oliver, M.A.; Webster, R. A tutorial guide to geostatistics: Computing and modelling variograms and kriging. Catena 2014, 113, 56–69. [Google Scholar] [CrossRef]
- Vargas, F.; Romero, M. Blooming time in almond progenies. Options Méditerranéennes 2001, 56, 29–34. [Google Scholar]
- Campoy, J.A.; Ruiz, D.; Cook, N.; Allderman, L.; Egea, J. High temperatures and time to budbreak in low chill apricot ‘Palsteyn’. Towards a better understanding of chill and heat requirements fulfilment. Sci. Hortic. 2011, 129, 649–655. [Google Scholar] [CrossRef]
- Pope, K.S.; da Silva, D.; Brown, P.H.; DeJong, T.M. A biologically based approach to modeling spring phenology in temperate deciduous trees. Agric. For. Meteorol. 2014, 198, 15–23. [Google Scholar] [CrossRef]
- Darbyshire, R.; Farrera, I.; Martinez-Lüscher, J.; Leite, G.B.; Mathieu, V.; El Yaacoubi, A.; Legave, J.-M. A global evaluation of apple flowering phenology models for climate adaptation. Agric. For. Meteorol. 2017, 240, 67–77. [Google Scholar] [CrossRef]
- Leida, C.; Conesa, A.; Llácer, G.; Badenes, M.L.; Ríos, G. Histone modifications and expression of DAM6 gene in peach are modulated during bud dormancy release in a cultivar-dependent manner. New Phytol. 2012, 193, 67–80. [Google Scholar] [CrossRef]
- Egea, J.; Ortega, E.; Marínez-Gómez, P.; Dicenta, F. Chilling and heat requirements of almond cultivars for flowering. Environ. Exp. Bot. 2003, 50, 79–85. [Google Scholar] [CrossRef]
- Ramirez, L.; Sagredo, K.X.; Reginato, G.H. Prediction models for chilling and heat requirements to estimate full bloom of almond cultivars in the Central Valley of Chile. Acta Horticulturae 2010, 872, 107–112. [Google Scholar] [CrossRef]
- Gaeta, L.; Stellacci, A.M.; Losciale, P. Evaluation of three modelling approaches for almond blooming in Mediterranean climate conditions. Eur. J. Agron. 2018, 97, 1–10. [Google Scholar] [CrossRef]
- Prudencio, A.S.; Martinez-Gomez, P.; Dicenta, F. Evaluation of breaking dormancy, flowering and productivity of extra-late and ultra-late flowering almond cultivars during cold and warm seasons in South-East of Spain. Sci. Hortic. 2018, 235, 39–46. [Google Scholar] [CrossRef]
- Socias i Company, R.; Felipe, A.J. Self-compatibility and autogamy in ‘Guara’ almond. J. Hortic. Sci. 1992, 67, 313–317. [Google Scholar] [CrossRef]
- Alonso Segura, J.M.; Ansón, J.; Espiau, M.; Socias i Company, R. Stability of the Almond Blooming Date in a Changing Climate. Acta Horticulturae 2011, 912, 337–342. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Chilling Phase | Forcing Phase | Chill-Force Overlap | % Chill-Force Gap | CR (CP) | HR (GDH) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Start | End | N° days | Start | End | N° days | % CR | % period | ||||
| Cavaliera | 20 Oct | 28 Nov | 40 | 24 Nov | 5 Feb | 74 | 81% | 5% | 11.56 ± 4.03 | 7452 ± 1601 | |
| Desmayo Largueta | 24 Oct | 24 Nov | 32 | 22 Nov | 8 Feb | 79 | 88% | 3% | 8.40 ± 3.77 | 8552 ± 1741 | |
| Ramillete | 24 Oct | 15 Dec | 53 | 4 Dec | 10 Feb | 69 | 68% | 11% | 20.71 ± 4.72 | 6998 ± 1540 | |
| Garrigues | 24 Oct | 17 Dec | 55 | 30 Nov | 15 Feb | 78 | 53% | 16% | 21.91 ± 4.66 | 8054 ± 1811 | |
| Gabaix | 24 Oct | 3 Dec | 41 | 15 Dec | 20 Feb | 68 | - | - | 10% | 13.35 ± 4.35 | 6824 ± 1421 |
| Mollar de Tarragona | 24 Oct | 14 Dec | 52 | 16 Dec | 20 Feb | 67 | - | - | 1% | 20.04 ± 4.68 | 6718 ± 1378 |
| Marcona | 23 Oct | 17 Dec | 56 | 17 Dec | 18 Feb | 63 | - | - | 21.96 ± 4.66 | 6378 ± 1341 | |
| Ardechoise | 24 Oct | 15 Dec | 53 | 10 Dec | 20 Feb | 73 | 85% | 4% | 21.82 ± 4.50 | 6994 ± 1546 | |
| Rof | 23 Oct | 15 Dec | 54 | 18 Dec | 24 Feb | 69 | - | - | 2% | 20.76 ± 4.72 | 6965 ± 1355 |
| Alicante | 24 Oct | 15 Dec | 53 | 16 Dec | 22 Feb | 69 | - | - | 20.71 ± 4.72 | 6940 ± 1400 | |
| Nonpareil | 24 Oct | 15 Dec | 53 | 16 Dec | 20 Feb | 67 | - | - | 20.71 ± 4.72 | 7062 ± 1399 | |
| Rana | 24 Oct | 14 Dec | 52 | 28 Dec | 28 Feb | 63 | - | - | 11% | 20.04 ± 4.68 | 6518 ± 1292 |
| A-258 | 23 Oct | 9 Dec | 48 | 7 Dec | 28 Feb | 84 | 93% | 2% | 17.05 ± 4.52 | 8725 ± 1712 | |
| Cristomorto | 24 Oct | 15 Dec | 53 | 16 Dec | 3 Mar | 78 | - | - | 20.71 ± 4.72 | 8236 ± 1482 | |
| Genco | 24 Oct | 27 Dec | 64 | 9 Jan | 4 Mar | 55 | - | - | 10% | 28.68 ± 4.90 | 5971 ± 1189 |
| Glorieta | 24 Oct | 31 Jan | 100 | 6 Jan | 28 Feb | 54 | 68% | 20% | 51.58 ± 5.90 | 5654 ± 1177 | |
| Texas | 24 Oct | 31 Jan | 100 | 7 Jan | 5 Mar | 58 | 69% | 19% | 51.58 ± 5.90 | 6280 ± 1225 | |
| Ferraduel | 24 Oct | 2 Feb | 102 | 29 Dec | 6 Mar | 68 | 56% | 27% | 52.85 ± 5.95 | 7285 ± 1362 | |
| Masbovera | 24 Oct | 27 Dec | 64 | 6 Jan | 4 Mar | 58 | - | - | 8% | 28.61 ± 4.90 | 6232 ± 1221 |
| Tuono | 24 Oct | 2 Feb | 102 | 29 Dec | 3 Mar | 65 | 56% | 28% | 52.85 ± 5.95 | 6870 ± 1319 | |
| Ferragnes | 24 Oct | 15 Dec | 53 | 16 Dec | 7 Mar | 82 | - | - | 20.71 ± 4.72 | 8696 ± 1543 | |
| Tarragones | 24 Oct | 31 Jan | 100 | 6 Jan | 5 Mar | 59 | 68% | 20% | 51.58 ± 5.90 | 6370 ± 1238 | |
| Tardy Nonpareil | 24 Oct | 6 Feb | 106 | 15 Dec | 11 Mar | 87 | 37% | 39% | 55.41 ± 5.91 | 9444 ± 1658 | |
| Primorskiy | 24 Oct | 2 Feb | 102 | 7 Dec | 10 Mar | 94 | 30% | 42% | 52.85 ± 5.95 | 10,201 ± 1834 | |
| Garbí | 24 Oct | 2 Feb | 102 | 7 Jan | 10 Mar | 94 | 68% | 20% | 52.85 ± 5.95 | 7040 ± 1312 | |
| CV | 0.53 | 0.15 | |||||||||
| Cultivar | Chilling Phase | Forcing Phase | Chill-Force Overlap | % Chill-Force Gap | CR (CP) | HR (GDH) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Start | End | N° days | Start | End | N° days | % CR | % period | ||||
| Aporo | 16 Nov | 12 Jan | 58 | 4 Jan | 4 Apr | 91 | 86% | 6% | 37.79 ± 2.72 | 9232 ± 1557 | |
| Pink Lady | 21 Nov | 16 Jan | 57 | 11 Feb | 12 Apr | 61 | - | - | 18% | 37.81 ± 2.25 | 8065 ± 1332 |
| Brookfield Gala | 8 Nov | 16 Jan | 70 | 24 Jan | 13 Apr | 80 | - | - | 6% | 44.48 ± 3.38 | 9501 ± 1556 |
| Fuji Chofu 2 | 11 Nov | 27 Jan | 78 | 16 Feb | 11 Apr | 55 | - | - | 13% | 50.26 ± 3.05 | 7471 ± 1191 |
| Granny Smith | 10 Nov | 19 Jan | 71 | 3 Feb | 13 Apr | 70 | - | - | 9% | 45.57 ± 3.15 | 8813 ± 1455 |
| Challenger | 11 Nov | 16 Jan | 67 | 9 Feb | 12 Apr | 63 | - | - | 15% | 42.98 ± 2.90 | 8202 ± 1347 |
| Early Red One | 10 Nov | 16 Jan | 68 | 26 Jan | 13 Apr | 78 | - | - | 6% | 43.58 ± 3.13 | 9349 ± 1530 |
| Red Chief | 11 Nov | 12 Jan | 63 | 17 Feb | 13 Apr | 56 | - | - | 23% | 40.24 ± 3.01 | 7767 ± 1175 |
| Fuji Zhen | 11 Nov | 2 Feb | 84 | 12 Feb | 13 Apr | 61 | - | - | 6% | 54.44 ± 3.04 | 8190 ± 1319 |
| Jeromine | 11 Nov | 25 Jan | 75 | 15 Feb | 15 Apr | 60 | - | - | 13% | 42.98 ± 2.90 | 8349 ± 1263 |
| Golden Smoothee | 10 Nov | 15 Jan | 67 | 2 Feb | 17 Apr | 75 | - | - | 11% | 42.81 ± 3.21 | 9690 ± 1530 |
| Golden Reinders | 11 Nov | 15 Jan | 66 | 14 Feb | 19 Apr | 65 | - | - | 18% | 42.38 ± 3.06 | 9239 ± 1315 |
| CV | 0.11 | 0.09 | |||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díez-Palet, I.; Funes, I.; Savé, R.; Biel, C.; de Herralde, F.; Miarnau, X.; Vargas, F.; Àvila, G.; Carbó, J.; Aranda, X. Blooming under Mediterranean Climate: Estimating Cultivar-Specific Chill and Heat Requirements of Almond and Apple Trees Using a Statistical Approach. Agronomy 2019, 9, 760. https://doi.org/10.3390/agronomy9110760
Díez-Palet I, Funes I, Savé R, Biel C, de Herralde F, Miarnau X, Vargas F, Àvila G, Carbó J, Aranda X. Blooming under Mediterranean Climate: Estimating Cultivar-Specific Chill and Heat Requirements of Almond and Apple Trees Using a Statistical Approach. Agronomy. 2019; 9(11):760. https://doi.org/10.3390/agronomy9110760
Chicago/Turabian StyleDíez-Palet, Isabel, Inmaculada Funes, Robert Savé, Carmen Biel, Felicidad de Herralde, Xavier Miarnau, Francisco Vargas, Glòria Àvila, Joaquim Carbó, and Xavier Aranda. 2019. "Blooming under Mediterranean Climate: Estimating Cultivar-Specific Chill and Heat Requirements of Almond and Apple Trees Using a Statistical Approach" Agronomy 9, no. 11: 760. https://doi.org/10.3390/agronomy9110760