Accelerating Breeding for Heat Tolerance in Tomato (Solanum lycopersicum L.): An Integrated Approach

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Types of Heat Stress Regimes
3. Screening Environments for Heat Tolerance Tomato: Towards A More Integrated Approach
4. Target Traits for Heat Tolerance Screening in Tomatoes
4.1. Male Reproductive Organ
4.2. Female Fertility
4.3. Biochemical Traits
4.4. Membrane Stability
4.5. Chlorophyll Fluorescence
4.6. Canopy Temperature as Selection Trait for Heat Tolerance
4.7. Choice of Yield Surrogate Traits
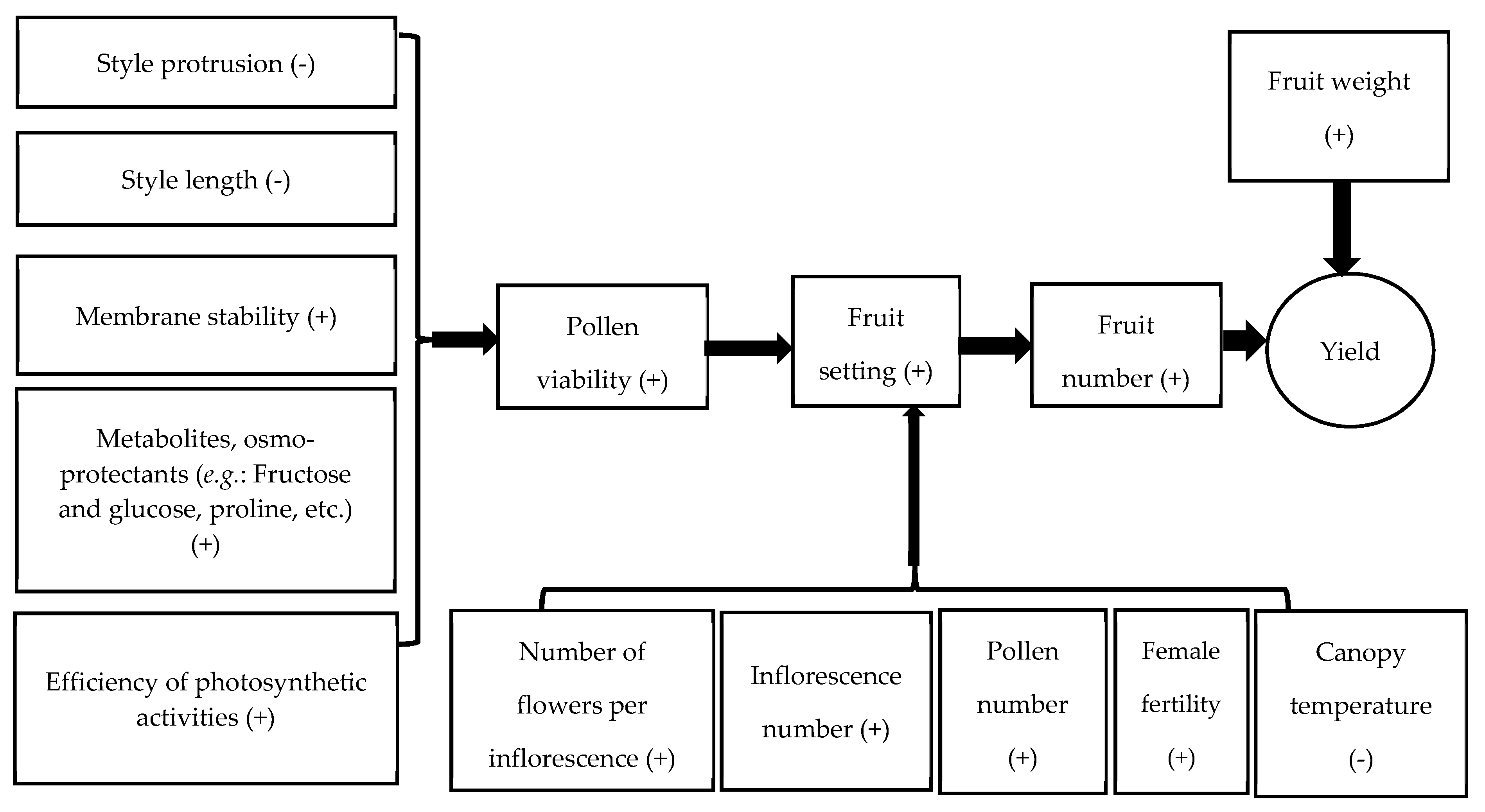
5. Association among Reproductive Traits under Heat Stress
6. Gene Action of Heat Tolerance Traits
7. Genetic Resources for Improvement of Heat Tolerance in Tomato
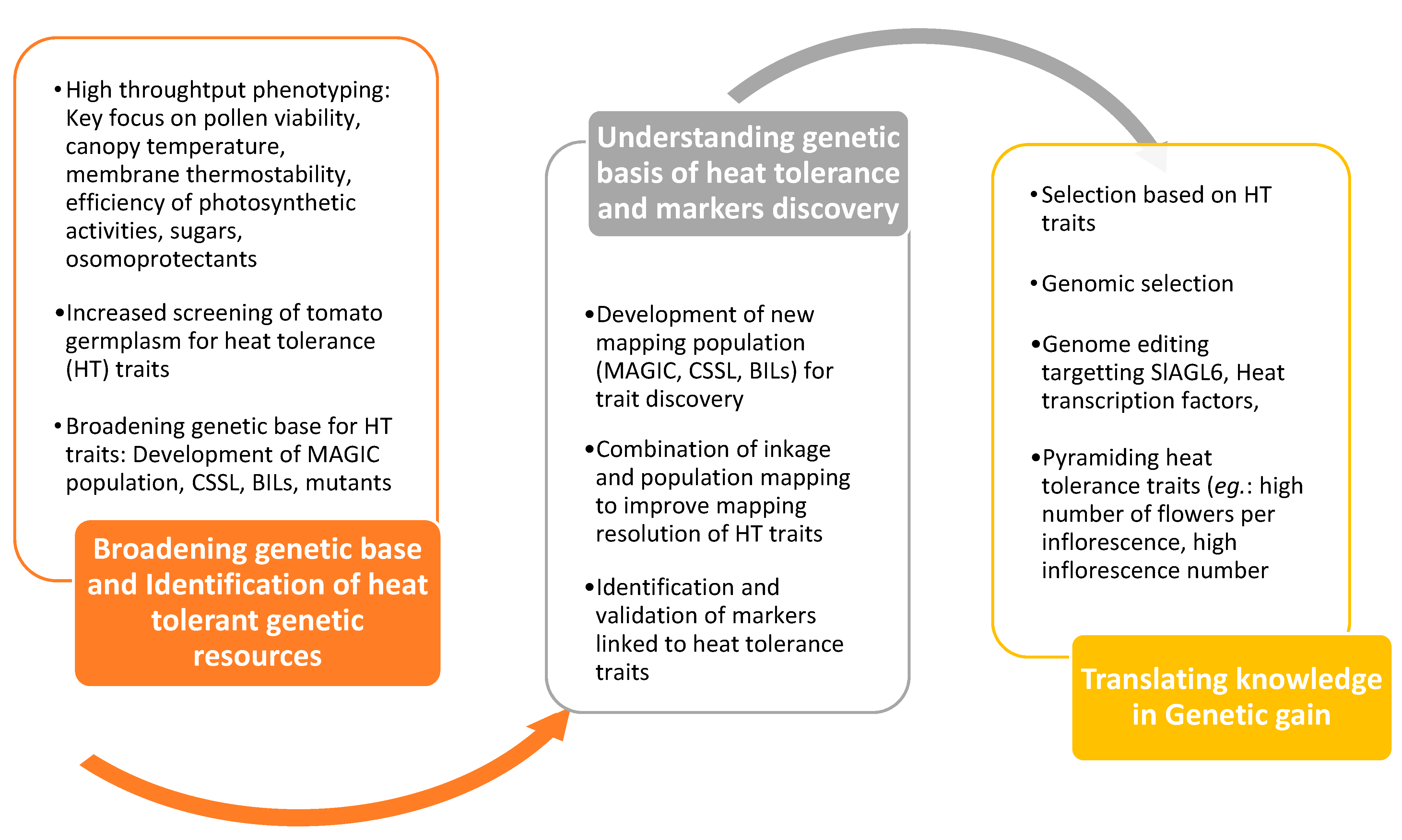
8. Better Exploitation of Tomato Genetic Resource for Heat Tolerance
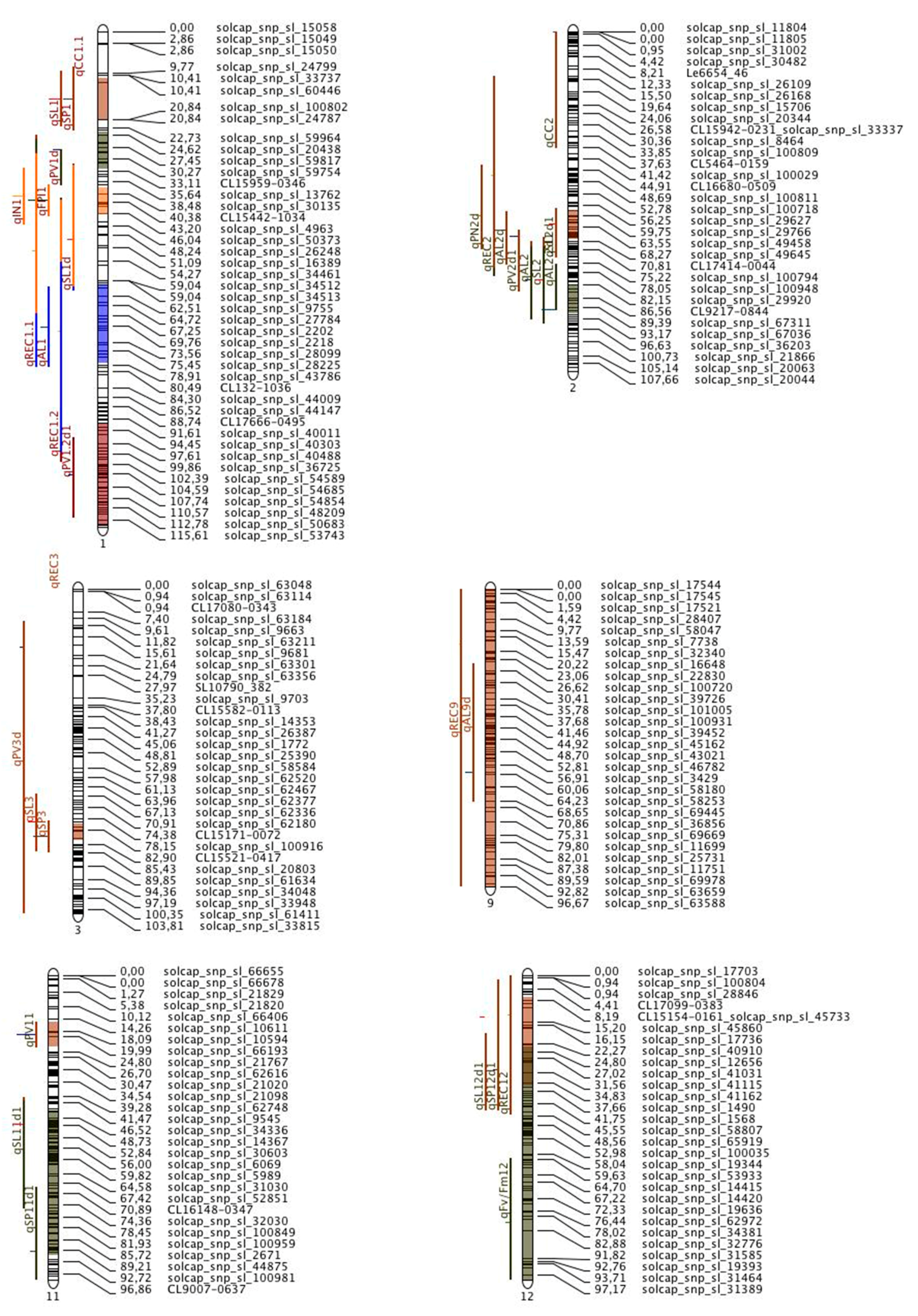
9. Genomic Resources for Improvement of Heat Tolerance in Tomato: Meta QTL Analysis
10. Taking Advantage of Emerging Breeding Techniques
10.1. Genomic Selection
10.2. Genome Editing Technologies to Improve Heat Tolerance
10.3. Application of Speed Breeding in Tomato
11. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Battisti, D.S.; Naylor, R.L. Historical Warnings of Future Food Insecurity with Uprecedented Seasonal Heat. Science 2009, 323, 403–406. [Google Scholar] [CrossRef] [PubMed]
- IPPC. Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change Core; Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; ISBN 9789291691432. [Google Scholar]
- Porter, J.R.; Xie, L.; Challinor, A.J.; Cochrane, K.; Howden, S.M.; Iqbal, M.M.; Lobell, D.B.; Travasso, M.I. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK, 2014; pp. 485–533. ISBN 9781107415379. [Google Scholar]
- Jones, B.J. Tomato Plant Culture: In the Field, Greenhouse, and Home Garden, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2008; Volume 136, ISBN 9780849373954. [Google Scholar]
- Peet, M.M.; Willits, D.H.; Gardner, R. Response of ovule development and post-pollen production processes in male-sterile tomatoes to chronic, sub-acute high temperature stress. J. Exp. Bot. 1997, 48, 101–111. [Google Scholar] [CrossRef]
- Sato, S.; Peet, M.M.; Thomas, J.F. Physiological factors limit fruit set of tomato (Lycopersicon esculentum Mill.) under chronic, mild heat stress. Plant Cell Environ. 2000, 23, 719–726. [Google Scholar] [CrossRef]
- Firon, N.; Shaked, R.; Peet, M.M.; Pharr, D.M.; Zamski, E.; Rosenfeld, K.; Althan, L.; Pressman, E. Pollen grains of heat tolerant tomato cultivars retain higher carbohydrate concentration under heat stress conditions. Sci. Hortic. 2006, 109, 212–217. [Google Scholar] [CrossRef]
- Silva, R.S.; Kumar, L.; Shabani, F.; Picanço, M.C. Assessing the impact of global warming on worldwide open field tomato cultivation through CSIRO-Mk3.0 global climate model. J. Agric. Sci. 2017, 155, 407–420. [Google Scholar] [CrossRef]
- Solh, M.; van Ginkel, M. Drought preparedness and drought mitigation in the developing world’s drylands. Weather Clim. Extrem. 2014, 3, 62–66. [Google Scholar] [CrossRef]
- De la Peña, R.C.; Ebert, A.W.; Gniffke, P.A.; Hanson, P.; Symonds, R.C. Genetic Adjustment to Changing Climates: Vegetables. In Crop Adaptation to Climate Change; Shyam, S., Robert, J., Jerry, L., Lotze-Campen, H., Anthony, E., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 396–410. ISBN 9780813820163. [Google Scholar]
- Hedhly, A. Sensitivity of flowering plant gametophytes to temperature fluctuations. Environ. Exp. Bot. 2011, 74, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Driedonks, N.; Rieu, I.; Vriezen, W.H. Breeding for plant heat tolerance at vegetative and reproductive stages. Plant Reprod. 2016, 29, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Tanger, P.; Klassen, S.; Mojica, J.P.; Lovell, J.T.; Moyers, B.T.; Baraoidan, M.; Naredo, M.E.B.; Mcnally, K.L.; Poland, J.; Bush, D.R.; et al. Field-based high throughput phenotyping rapidly identifies genomic regions controlling yield components in rice. Nat. Publ. Gr. 2017, 1–8. [Google Scholar] [CrossRef]
- Araus, J.L.; Kefauver, S.C. Breeding to adapt agriculture to climate change: Affordable phenotyping solutions. Curr. Opin. Plant Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Crain, J.; Mondal, S.; Rutkoski, J.; Singh, R.P.; Poland, J. Combining High-Throughput Phenotyping and Genomic Information to Increase Prediction and Selection Accuracy in Wheat Breeding. Plant Genome 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Halperin, O.; Gebremedhin, A.; Wallach, R.; Moshelion, M. High-throughput physiological phenotyping and screening system for the characterization of plant—Environment interactions. Plant J. 2017, 89, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Araus, J.L.; Kefauver, S.C.; Zaman-Allah, M.; Olsen, M.S.; Cairns, J.E. Translating High-Throughput Phenotyping into Genetic Gain. Trends Plant Sci. 2018, 23, 451–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesihovic, A.; Iannacone, R.; Firon, N.; Fragkostefanakis, S. Heat stress regimes for the investigation of pollen thermotolerance in crop plants. Plant Reprod. 2016, 29, 93–105. [Google Scholar] [CrossRef]
- Driedonks, N. From Flower to Fruit in the Heat Reproductive Thermotolerance in Tomato and Its Wild Relatives. Ph.D. Thesis, Radboud University, Nijmegen, The Netherlands, 2018. [Google Scholar]
- Paupière, M.J.; van Haperen, P.; Rieu, I.; Visser, R.G.F.; Tikunov, Y.M.; Bovy, A.G. Screening for pollen tolerance to high temperatures in tomato. Euphytica 2017, 213, 130. [Google Scholar] [CrossRef]
- Shaheen, M.R.; Ayyub, C.M.; Amjad, M.; Waraich, E.A. Morpho-physiological evaluation of tomato genotypes under high temperature stress conditions. J. Sci. Food Agric. 2016, 96, 2698–2704. [Google Scholar] [CrossRef]
- Xu, J.; Wolters-Arts, M.; Mariani, C.; Huber, H.; Rieu, I. Heat stress affects vegetative and reproductive performance and trait correlations in tomato (Solanum lycopersicum). Euphytica 2017, 213, 1–12. [Google Scholar] [CrossRef]
- Xu, J.; Driedonks, N.; Rutten, M.J.M.; Vriezen, W.H.; de Boer, G.-J.; Rieu, I. Mapping quantitative trait loci for heat tolerance of reproductive traits in tomato (Solanum lycopersicum). Mol. Breed. 2017, 37, 58. [Google Scholar] [CrossRef]
- Paupière, M.J.; van Heusden, A.W.; Bovy, A.G. The metabolic basis of pollen thermo-tolerance: Perspectives for breeding. Metabolites 2014, 4, 889–920. [Google Scholar] [CrossRef]
- Bokszczanin, K. Perspectives on deciphering mechanisms underlying plant heat stress response and thermotolerance. Front. Plant Sci. 2013, 4, 1–20. [Google Scholar] [CrossRef] [PubMed]
- De Storme, N.; Geelen, D. The impact of environmental stress on male reproductive development in plants: Biological processes and molecular mechanisms. Plant. Cell Environ. 2014, 37, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Zinn, K.E.; Tunc-Ozdemir, M.; Harper, J.F. Temperature stress and plant sexual reproduction: Uncovering the weakest links. J. Exp. Bot. 2010, 61, 1959–1968. [Google Scholar] [CrossRef] [PubMed]
- Jha, U.C.; Bohra, A.; Singh, N.P. Heat stress in crop plants: Its nature, impacts and integrated breeding strategies to improve heat tolerance. Plant Breed. 2014, 133, 679–701. [Google Scholar] [CrossRef]
- Hazra, P.; Samsul, A.; Sikder, D.; Peter, K.V. Breeding Tomato (Lycopersicon esculentum Mill.) Resistant to High Temperature Stress. Int. J. Plant Breed. 2007, 1, 31–40. [Google Scholar]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Yeh, C.H.; Kaplinsky, N.J.; Hu, C.; Charng, Y. Some like it hot, some like it warm: Phenotyping to explore thermotolerance diversity. Plant Sci. 2012, 195, 10–23. [Google Scholar] [CrossRef] [Green Version]
- Driedonks, N.; Wolters-Arts, M.; Huber, H.; de Boer, G.J.; Vriezen, W.; Mariani, C.; Rieu, I. Exploring the natural variation for reproductive thermotolerance in wild tomato species. Euphytica 2018, 214, 67. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Yu, X.; Kjær, K.H.; Rosenqvist, E.; Ottosen, C.O.; Wu, Z. Screening and validation of tomato genotypes under heat stress using Fv/Fm to reveal the physiological mechanism of heat tolerance. Environ. Exp. Bot. 2015, 118, 1–11. [Google Scholar] [CrossRef]
- Camejo, D.; Rodríguez, P.; Morales, M.A.; Dell’Amico, J.M.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J. Plant Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef]
- Zhou, R.; Wu, Z.; Wang, X.; Rosenqvist, E.; Wang, Y.; Zhao, T.; Ottosen, C.O. Evaluation of temperature stress tolerance in cultivated and wild tomatoes using photosynthesis and chlorophyll fluorescence. Hortic. Environ. Biotechnol. 2018, 59, 499–509. [Google Scholar] [CrossRef]
- Sato, S.; Kamiyama, M.; Iwata, T.; Makita, N.; Furukawa, H.; Ikeda, H. Moderate increase of mean daily temperature adversely affects fruit set of Lycopersicon esculentum by disrupting specific physiological processes in male reproductive development. Ann. Bot. 2006, 97, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Dane, F.; Hunter, A.G.; Chambliss, O.L. Fruit Set, Pollen Fertility, and Combining Ability of Selected Tomato Genotypes under High-Temperature Field Conditions. J. Am. Soc. Hort. Sci. 1991, 116, 906–910. [Google Scholar] [CrossRef]
- Poudyal, D.; Rosenqvist, E.; Ottosen, C.-O. Phenotyping from lab to field -tomato lines screened for heat stress using Fv/Fm maintain high fruit yield during thermal stress in the field. Funct. Plant Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hall, A. Breeding for heat tolerance-an approach based on whole-plant physiology. Hort Sci. 1990, 25, 17–19. [Google Scholar] [CrossRef]
- Kugblenu, Y.O.; Oppong Danso, E.; Ofori, K.; Andersen, M.N.; Abenney-Mickson, S.; Sabi, E.B.; Plauborg, F.; Abekoe, M.K.; Ofosu-Anim, J.; Ortiz, R.; et al. Screening tomato genotypes for adaptation to high temperature in West Africa. Acta Agric. Scand. Sect. B Soil Plant Sci. 2013, 63, 516–522. [Google Scholar] [CrossRef]
- Singh, U.; Patel, P.K.; Singh, A.K.; Tiwari, V.; Kumar, R.; Rai, N.; Bahadur, A.; Tiwari, S.K.; Singh, M.; Singh, B. Screening of tomato genotypes underhigh temperature stress for reproductive traits. Veg. Sci. 2015, 42, 52–55. [Google Scholar]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Bai, Y.; Kissoudis, C.; Yan, Z.; Visser, R.G.F.; van der Linden, G. Plant behaviour under combined stress: Tomato responses to combined salinity and pathogen stress. Plant J. 2018, 93. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Ottosen, C.O.; Rosenqvist, E.; Zhao, L.; Wang, Y.; Yu, W.; Zhao, T.; Wu, Z. Drought stress had a predominant effect over heat stress on three tomato cultivars subjected to combined stress. BMC Plant Biol. 2017, 17, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Rivero, R.M.; Mestre, T.C.; Mittler, R.; Rubio, F.; Garcia-Sanchez, F.; Martinez, V. The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants. Plant Cell Environ. 2014, 37, 1059–1073. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.C.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R.M. Tolerance to stress combination in tomato plants: New insights in the protective role of melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef] [PubMed]
- Leke, W.N.; Mignouna, D.B.; Brown, J.K.; Kvarnheden, A. Begomovirus disease complex: Emerging threat to vegetable production systems of West and Central Africa. Agric. Food Secur. 2015, 4, 1. [Google Scholar] [CrossRef]
- Sikirou, R.; Beed, F.; Ezin, V.; Hoteigni, J.; Miller, S.A. Distribution, pathological and biochemical characterization of Ralstonia solanacearum in Benin. Ann. Agric. Sci. 2017, 62, 83–88. [Google Scholar] [CrossRef]
- Yan, C.; Ding, Y.; Wang, Q.; Liu, Z.; Li, G.; Muhammad, I.; Wang, S. The impact of relative humidity, genotypes and fertilizer application rates on panicle, leaf temperature, fertility and seed setting of rice. J. Agric. Sci. 2010, 148, 329–339. [Google Scholar] [CrossRef]
- Weerakoon, W.M.W.; Maruyama, A.; Ohba, K. Impact of Humidity on Temperature-Induced Grain Sterility in Rice (Oryza sativa L). J. Agron. Crop Sci. 2008, 194, 135–140. [Google Scholar] [CrossRef]
- Hanson, P.M.; Chen, J.T.; Kuo, G. Gene action and heritability of high-temperature fruit set in tomato line CL5915. Hort. Sci. 2002, 37, 172–175. [Google Scholar] [CrossRef]
- Kakani, V.G.; Reddy, K.R.; Koti, S.; Wallace, T.P.; Prasad, P.V.V.; Reddy, V.R.; Zhao, D. Differences in in vitro pollen germination and pollen tube growth of cotton cultivars in response to high temperature. Ann. Bot. 2005, 96, 59–67. [Google Scholar] [CrossRef]
- Singh, S.K.; Kakani, V.G.; Brand, D.; Baldwin, B.; Reddy, K.R. Assessment of cold and heat tolerance of winter-grown canola (Brassica napus L.) cultivars by pollen-based parameters. J. Agron. Crop Sci. 2008, 194, 225–236. [Google Scholar] [CrossRef]
- Reddy, R.K.; Kakani, V.G. Screening Capsicum species of different origins for high temperature tolerance by in vitro pollen germination and pollen tube length. Sci. Hortic. 2007, 112, 130–135. [Google Scholar] [CrossRef]
- Dreccer, M.F.; Molero, G.; Rivera-Amado, C.; John-Bejai, C.; Wilson, Z. Yielding to the image: How phenotyping reproductive growth can assist crop improvement and production. Plant Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Heidmann, I.; Schade-Kampmann, G.; Lambalk, J.; Ottiger, M.; Di Berardino, M. Impedance flow cytometry: A novel technique in pollen analysis. PLoS ONE 2016, 11, e0165531. [Google Scholar] [CrossRef] [PubMed]
- Tello, J.; Montemayor, M.I.; Forneck, A.; Ibáñez, J. A new image-based tool for the high throughput phenotyping of pollen viability: Evaluation of inter- and intra-cultivar diversity in grapevine. Plant Methods 2018, 14, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Obata, T.; Witt, S.; Lisec, J.; Palacios-Rojas, N.; Florez-Sarasa, I.; Araus, J.L.; Cairns, J.E.; Yousfi, S.; Fernie, A.R. Metabolite profiles of maize leaves in drought, heat and combined stress field trials reveal the relationship between metabolism and grain yield. Plant Physiol. 2015, 169, 01164. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Nada, K.; Tachibana, S. Ameliorative effect of polyamines on the high temperature inhibition of in vitro pollen germination in tomato (Lycopersicon esculentum Mill.). Sci. Hortic. 1999, 80, 203–212. [Google Scholar] [CrossRef]
- Song, J.; Nada, K.; Tachibana, S. Suppression of S-adenosylmethionine decarboxylase activity is a major cause for high-temperature inhibition of pollen germination and tube growth in tomato (Lycopersicon esculentum Mill.). Plant Cell Physiol. 2002, 43, 619–627. [Google Scholar] [CrossRef]
- Cheng, L.; Zou, Y.; Ding, S.; Zhang, J.; Yu, X.; Cao, J.; Lu, G. Polyamine accumulation in transgenic tomato enhances the tolerance to high temperature stress. J. Integr. Plant Biol. 2009, 51, 489–499. [Google Scholar] [CrossRef]
- Din, J.U.; Khan, S.U.; Khan, A.; Qayyum, A.; Abbasi, K.S.; Jenks, M.A. Evaluation of potential morpho-physiological and biochemical indicators in selecting heat-tolerant tomato (Solanum lycopersicum Mill.) genotypes. Hortic. Environ. Biotechnol. 2015, 56, 769–776. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Dobra, J.; Motyka, V.; Dobrev, P.; Malbeck, J.; Prasil, I.T.; Haisel, D.; Gaudinova, A.; Havlova, M.; Gubis, J.; Vankova, R. Comparison of hormonal responses to heat, drought and combined stress in tobacco plants with elevated proline content. J. Plant Physiol. 2010, 167, 1360–1370. [Google Scholar] [CrossRef]
- Schijlen, E.G.W.M.; de Vos, C.H.R.; Martens, S.; Jonker, H.H.; Rosin, F.M.; Molthoff, J.W.; Tikunov, Y.M.; Angenent, G.C.; van Tunen, A.J.; Bovy, A.G. RNA Interference Silencing of Chalcone Synthase, the First Step in the Flavonoid Biosynthesis Pathway, Leads to Parthenocarpic Tomato Fruits. Plant Physiol. 2007, 144, 1520–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, C.; Yang, D.; Zhao, X.; Jiao, C.; Yan, Y.; Lamin-Samu, A.T.; Wang, Q.; Xu, X.; Fei, Z.; Lu, G. Tomato stigma exsertion induced by high temperature is associated with the jasmonate signaling pathway. Plant. Cell Environ. 2018, 1–17. [Google Scholar] [CrossRef]
- Saeed, A.; Hayat, K.; Khan, A.A.; Iqbal, S. Heat Tolerance Studies in Tomato (Lycopersicon esculentum Mill.). Int. J. Agric. Biol. 2007, 9, 649–652. [Google Scholar]
- Srinivasan, A.; Takeda, H.; Senboku, T. Heat tolerance in food legumes as evaluated by cell membrane thermostability and chlorophyll fluorescence techniques. Euphytica 1996, 88, 35–45. [Google Scholar] [CrossRef]
- Ismail, A.M.; Hall, A.E. Reproductive-Stage Heat Tolerance, Leaf Membrane Thermostability and Plant Morphology in Cowpea. Crop Sci. 1999, 39, 1762–1768. [Google Scholar] [CrossRef]
- Čajánek, M.; Štroch, M.; Lachetová, I.; Kalina, J.; Špunda, V. Characterization of the photosystem II inactivation of heat-stressed barley leaves as monitored by the various parameters of chlorophyll a fluorescence and delayed fluorescence. J. Photochem. Photobiol. B Biol. 1998, 47, 39–45. [Google Scholar] [CrossRef]
- Mathur, S.; Agrawal, D.; Jajoo, A. Photosynthesis: Response to high temperature stress. J. Photochem. Photobiol. B Biol. 2014, 137, 116–126. [Google Scholar] [CrossRef]
- Herzog, H.; Chai-Arree, W. Gas Exchange of Five Warm-Season Grain Legumes and their Susceptibility to Heat Stress. J. Agron. Crop Sci. 2012, 198, 466–474. [Google Scholar] [CrossRef]
- Sharma, D.K.; Fernández, J.O.; Rosenqvist, E.; Ottosen, C.O.; Andersen, S.B. Genotypic response of detached leaves versus intact plants for chlorophyll fluorescence parameters under high temperature stress in wheat. J. Plant Physiol. 2014, 171, 576–586. [Google Scholar] [CrossRef]
- Sharma, D.K.; Torp, A.M.; Rosenqvist, E.; Ottosen, C.-O.; Andersen, S.B. QTLs and Potential Candidate Genes for Heat Stress Tolerance Identified from the Mapping Populations Specifically Segregating for Fv/Fm in Wheat. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Baker, N.R.; Rosenqvist, E. Applications of chlorophyll fluorescence can improve crop production strategies: An examination of future possibilities. J. Exp. Bot. 2004, 55, 1607–1621. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, H.; Eslick, R.F.; Aase, J.K. Canopy temperatures of barley as influenced by morphological characteristics. Agron. J. 1973, 65, 425–428. [Google Scholar] [CrossRef]
- Pask, A.; Pietragalla, J.; Mullan, D.; Reynolds, M. Physiological Breeding II: A Field Guide to Wheat Phenotyping; Pask, A., Pietragalla, J., Mullan, D., Reynolds, M., Eds.; CIMMYT: Texcoco, Mexico, 2012. [Google Scholar]
- Lopes, M.S.; Reynolds, M.P. Partitioning of assimilates to deeper roots is associated with cooler canopies and increased yield under drought in wheat. Funct. Plant Biol. 2010, 37, 147–156. [Google Scholar] [CrossRef]
- Gautam, A.; Sai Prasad, S.V.; Jajoo, A.; Ambati, D. Canopy Temperature as a Selection Parameter for Grain Yield and Its Components in Durum Wheat Under Terminal Heat Stress in Late Sown Conditions. Agric. Res. 2015, 4, 238–244. [Google Scholar] [CrossRef]
- Webber, H.; Ewert, F.; Kimball, B.A.; Siebert, S.; White, J.W.; Wall, G.W.; Ottman, M.J.; Trawally, D.N.A.; Gaiser, T. Simulating canopy temperature for modelling heat stress in cereals. Environ. Model. Softw. 2016, 77, 143–155. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, S.; Acevedo, E.; Grando, S. Breeding for yield stability in unpredictable environments: Single traits, interaction between traits, and architecture of genotypes. Euphytica 1991, 56, 169–185. [Google Scholar] [CrossRef]
- Levy, A.; Rabinowitch, H.D.; Kedar, N. Morphological and physiological characters affecting flower drop and fruit set of tomatoes at high temperatures. Euphytica 1978, 27, 211–218. [Google Scholar] [CrossRef]
- Wessel-Beaver, L.; Scott, J.W. Genetic Variability of Fruit Set, Fruit Weight, and Yield in a Tomato Population Grown in Two High-temperature Environments. Am. Soc. Hortic. Sci. 1992, 117, 867–870. [Google Scholar] [CrossRef] [Green Version]
- Scott, J.W.; Volin, R.B.; Bryan, H.H.; Olson, S.M. Use of Hybrids to Develop Heat Tolerant Tomato Cultivars. Proc. Fla. State Hortic. Soc. 1986, 99, 311–314. [Google Scholar]
- Bertin, N. Competition for assimilates and fruit position affect fruit set in indeterminate greenhouse tomato. Ann. Bot. 1995, 75, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Peet, M.M.; Gardner, R.G. Formation of parthenocarpic fruit, undeveloped flowers and aborted flowers in tomato under moderately elevated temperatures. Sci. Hortic. 2001, 90, 243–254. [Google Scholar] [CrossRef]
- Giorno, F.; Wolters-Arts, M.; Mariani, C.; Rieu, I. Ensuring Reproduction at High Temperatures: The Heat Stress Response during Anther and Pollen Development. Plants 2013, 2, 489–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.; Quamruzzaman, A.K.M.; Uddin, M.N. Combining ability estimates of tomato (Solanum lycopersicum) in late Summer. SAARC J. Agric. 2009, 7, 43–56. [Google Scholar]
- El-Ahmadi, A.B.; Stevens, M.A. Reproductive responses of heat tolerant tomatoes to high temperatures. J. Am. Soc. Hortic. Sci. 1979, 686–691. [Google Scholar]
- Srivastava, K.; Kumar, S.; Bhandari, H.R.; Vaishampayan, A. Genetics of heat stress tolerance traits in tomato (Solanum lycopersicum L.). Vegetos Int. J. Plant Res. 2017, 30, 397. [Google Scholar] [CrossRef]
- Wen, J.; Jiang, F.; Weng, Y.; Sun, M.; Shi, X.; Zhou, Y.; Yu, L.; Wu, Z. Identification of heat-tolerance QTLs and high-temperature stress-responsive genes through conventional QTL mapping, QTL-seq and RNA-seq in tomato. BMC Plant Biol. 2019, 19, 1–17. [Google Scholar] [CrossRef]
- Scott, J.W.; Olson, S.M.; Bryan, H.H.; Bartz, J.A.; Maynard, D.N.; Stoffella, P.J. “Solar Fire” hybrid tomato: Fla. 7776 tomato breeding line. Hort. Sci. 2006, 41, 1504–1505. [Google Scholar] [CrossRef]
- Scott, J.W. Fla. 7771, a medium-large, heat- tolerant, jointless-pedicel tomato. Hort. Sci. 2000, 35, 968–969. [Google Scholar] [CrossRef]
- Melchinger, A.E.; Gumber, R.K. Overview of Heterosis and Heterotic Groups in Agronomic Crops. In Concepts and Breeding of Heterosis in Crop Plants; CSSA: Madison, WI, USA, 1998; pp. 29–44. [Google Scholar] [CrossRef]
- Zhang, H.; Mittal, N.; Leamy, L.J.; Barazani, O.; Song, B.H. Back into the wild—Apply untapped genetic diversity of wild relatives for crop improvement. Evol. Appl. 2017, 10, 5–24. [Google Scholar] [CrossRef]
- De la Peña, R.; Hughes, J. Improving Vegetable Productivity in a Variable Changing Climate. ICRISAT 2007, 4, 1–22. [Google Scholar]
- Bedinger, P.A.; Chetelat, R.T.; McClure, B.; Moyle, L.C.; Rose, J.K.C.; Stack, S.M.; van der Knaap, E.; Baek, Y.S.; Lopez-Casado, G.; Covey, P.A.; et al. Interspecific reproductive barriers in the tomato clade: Opportunities to decipher mechanisms of reproductive isolation. Sex. Plant Reprod. 2011, 24, 171–187. [Google Scholar] [CrossRef] [PubMed]
- Klap, C.; Yeshayahou, E.; Bolger, A.M.; Arazi, T.; Gupta, S.K.; Shabtai, S.; Usadel, B.; Salts, Y.; Barg, R. Tomato facultative parthenocarpy results from Slagamous-like 6 loss of function. Plant Biotechnol. J. 2017, 15, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Tanksley, S.D.; Nelson, J.C. Advanced backcross QTL analysis: A method for the simultaneous discovery and transfer of valuable QTLs from unadapted germplasm into elite breeding lines. Theor. Appl. Genet. 1996, 92, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.L.; Sanchez, P.L.; Yu, S.; Lorieux, M.; Eizenga, G.C. Chromosome segment substitution lines: A powerful tool for the introgression of valuable genes from Oryza wild species into cultivated rice (O. sativa). Rice 2010, 3, 218–234. [Google Scholar] [CrossRef]
- Bessho-Uehara, K.; Furuta, T.; Masuda, K.; Yamada, S.; Angeles-Shim, R.B.; Ashikari, M.; Takashi, T. Construction of rice chromosome segment substitution lines harboring Oryza genome and evaluation of yield-related traits. Breed. Sci. 2017, 67, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, L.; Bai, Y.; Zhang, P.; Finkers, R.; Du, Y.; Visser, R.G.F.; van Heusden, A.W. Seedling salt tolerance in tomato. Euphytica 2011, 178, 403–414. [Google Scholar] [CrossRef]
- Uozumi, A.; Ikeda, H.; Hiraga, M.; Kanno, H.; Nanzyo, M.; Nishiyama, M.; Kanahama, K.; Kanayama, Y. Tolerance to salt stress and blossom-end rot in an introgression line, IL8-3, of tomato. Sci. Hortic. 2012, 138, 1–6. [Google Scholar] [CrossRef]
- Barrantes, W.; López-Casado, G.; García-Martínez, S.; Alonso, A.; Rubio, F.; Ruiz, J.J.; Fernández-Muñoz, R.; Granell, A.; Monforte, A.J. Exploring New Alleles Involved in Tomato Fruit Quality in an Introgression Line Library of Solanum pimpinellifolium. Front. Plant Sci. 2016, 7, 1–12. [Google Scholar] [CrossRef]
- Celik, I.; Gurbuz, N.; Uncu, A.T.; Frary, A.; Doganlar, S. Genome-wide SNP discovery and QTL mapping for fruit quality traits in inbred backcross lines (IBLs) of Solanum pimpinellifolium using genotyping by sequencing. BMC Genom. 2017, 18, 1–10. [Google Scholar] [CrossRef]
- Haggard, J.E.; Johnson, E.B.; St. Clair, D.A. Multiple QTL for Horticultural Traits and Quantitative Resistance to Phytophthora infestans Linked on Solanum habrochaites Chromosome 11. G3 Genes Genom. Genet. 2015, 5, 219–233. [Google Scholar] [CrossRef]
- Grandillo, S.; Tanksley, S.D. Advanced backcross QTL analysis: Results and perspectives. In Proceedings of the International Congress “In the wake of the double helix: From the green revolution to the Gene Revolution”, Bologna, Italy, 27–31 May 2003; Avenue media: Bologna, Italy, 2005; pp. 115–132. [Google Scholar]
- Grilli, G.V.G.; Braz, L.T.; Lemos, E.G.M. QTL identification for tolerance to fruit set in tomato by FAFLP markers. Crop Breed. Appl. Biotechnol. 2007, 7, 234–241. [Google Scholar] [CrossRef]
- Lin, K.H.; Yeh, W.L.; Chen, H.M.; Lo, H.F. Quantitative trait loci influencing fruit-related characteristics of tomato grown in high-temperature conditions. Euphytica 2010, 174, 119–135. [Google Scholar] [CrossRef]
- Darvasi, A.; Soller, M. A simple method to calculate resolving power and confidence interval of QTL map location. Behav. Genet. 1997, 27, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Sosnowski, O.; Charcosset, A.; Joets, J. Biomercator V3: An upgrade of genetic map compilation and quantitative trait loci meta-analysis algorithms. Bioinformatics 2012, 28, 2082–2083. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.C.; Durstewitz, G.; Plieske, J.; Wieseke, R.; Ganal, M.W.; van Deynze, A.; Hamilton, J.P.; Buell, C.R.; Causse, M.; Wijeratne, S.; et al. Development of a large SNP genotyping array and generation of high-density genetic maps in tomato. PLoS ONE 2012, 7, e40563. [Google Scholar] [CrossRef] [PubMed]
- Veyrieras, J.B.; Goffinet, B.; Charcosset, A. MetaQTL: A package of new computational methods for the meta-analysis of QTL mapping experiments. BMC Bioinf. 2007, 8. [Google Scholar] [CrossRef] [PubMed]
- Crossa, J.; Pérez-Rodríguez, P.; Cuevas, J.; Montesinos-López, O.; Jarquín, D.; de los Campos, G.; Burgueño, J.; González-Camacho, J.M.; Pérez-Elizalde, S.; Beyene, Y.; et al. Genomic Selection in Plant Breeding: Methods, Models, and Perspectives. Trends Plant Sci. 2017, 22, 961–975. [Google Scholar] [CrossRef]
- Ruggieri, V.; Calafiore, R.; Schettini, C.; Rigano, M.M.; Olivieri, F.; Frusciante, L.; Barone, A. Exploiting Genetic and Genomic Resources to Enhance Heat-Tolerance in Tomatoes. Agronomy 2019, 9, 22. [Google Scholar] [CrossRef]
- Alsamir, M.; Ahmad, N.; Arief, V.; Mahmood, T.; Trethowan, R. Phenotypic diversity and marker-trait association studies under heat stress in tomato (Solanum lycopersicum L.). Aust. J. Crop Sci. 2019, 13, 578–587. [Google Scholar] [CrossRef]
- Wilson, L.M. Dissection of Maize Kernel Composition and Starch Production by Candidate Gene Association. Plant Cell Online 2004, 16, 2719–2733. [Google Scholar] [CrossRef] [Green Version]
- Myles, S.; Peiffer, J.; Brown, P.J.; Ersoz, E.S.; Zhang, Z.; Costich, D.E.; Buckler, E. Association mapping: Critical considerations shift from genotyping to experimental design. Plant Cell 2009, 21, 2194–2202. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.H.; Gouker, F.E.; Crowell, C.R.; Evans, L.; DiFazio, S.P.; Smart, C.D.; Smart, L.B. Joint linkage and association mapping of complex traits in shrub willow (Salix purpurea L.). Ann. Bot. 2019, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Pascual, L.; Desplat, N.; Huang, B.E.; Desgroux, A.; Bruguier, L.; Bouchet, J.P.; Le, Q.H.; Chauchard, B.; Verschave, P.; Causse, M. Potential of a tomato magic population to decipher the genetic control of quantitative traits and detect causal variants in the resequencing era. Plant Biotechnol. J. 2015, 13, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, C.; Morell, M.; Mackay, I.; Powell, W. From mutations to magic: Resources for gene discovery, validation and delivery in crop plants. Curr. Opin. Plant Biol. 2008, 11, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar]
- Lorenz, A.J.; Chao, S.; Asoro, F.G.; Heffner, E.L.; Hayashi, T.; Iwata, H.; Smith, K.P.; Sorrells, M.E.; Jannink, J.L. Genomic Selection in Plant Breeding. Knowledge and Prospects; Elsevier: Amsterdam, The Netherlands, 2011; Volume 110, ISBN 9780123855312. [Google Scholar]
- Duangjit, J.; Causse, M.; Sauvage, C. Efficiency of genomic selection for tomato fruit quality. Mol. Breed. 2016, 36, 29. [Google Scholar] [CrossRef]
- Yamamoto, E.; Matsunaga, H.; Onogi, A.; Ohyama, A.; Miyatake, K.; Yamaguchi, H.; Nunome, T.; Iwata, H.; Fukuoka, H. Efficiency of genomic selection for breeding population design and phenotype prediction in tomato. Heredity 2017, 118, 202–209. [Google Scholar] [CrossRef]
- Van Eck, J. Genome editing and plant transformation of solanaceous food crops. Curr. Opin. Biotechnol. 2018, 49, 35–41. [Google Scholar] [CrossRef]
- Kim, Y.-G.; Cha, J.; Srinivasan, C. Engineering the xylan utilization system in Bacillus subtilis for production of acidic xylooligosaccharides. Proc. Natl. Acad. Sci. 1996, 93, 1156–1160. [Google Scholar] [CrossRef]
- Bogdanove, A.J.; Voytas, D.F. TAL effectors: Customizable proteins for DNA targeting. Science 2011, 333, 1843–1846. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Zhang, B.; Wona, D.; Xiaodong, L.; Dong-Lei, Y.; Pengliang, W.; Fengqiu, C.; Shihua, Z.; Feng, Z.; Mao, Y.; et al. Efficient genome editing in zebrafish using a CRISPR-Cas system. Cell Res. 2013, 23, 1229–1232. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.S.; Ding, J.; Li, Y. Genome-editing technologies and their potential application in horticultural crop breeding. Hortic. Res. 2015, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tomato Genome Consortium. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef] [Green Version]
- Zsögön, A.; Čermák, T.; Naves, E.R.; Notini, M.M.; Edel, K.H.; Weinl, S.; Freschi, L.; Voytas, D.F.; Kudla, J.; Peres, L.E.P. De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 2018. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Fragkostefanakis, S.; Mesihovic, A.; Simm, S.; Paupière, M.J.; Hu, Y.; Paul, P.; Mishra, S.K.; Tschiersch, B.; Theres, K.; Bovy, A.; et al. HsfA2 controls the activity of developmentally and stress-regulated heat stress protection mechanisms in tomato male reproductive tissues. Plant Physiol. 2016, 170, 2461–2477. [Google Scholar] [CrossRef]
- Fragkostefanakis, S.; Simm, S.; El-Shershaby, A.; Hu, Y.; Bublak, D.; Mesihovic, A.; Darm, K.; Mishra, S.K.; Tschiersch, B.; Theres, K.; et al. The repressor and co-activator HsfB1 regulates the major heat stress transcription factors in tomato. Plant Cell Environ. 2018. [Google Scholar] [CrossRef]
- Berz, J.; Simm, S.; Schuster, S.; Scharf, K.-D.; Schleiff, E.; Ebersberger, I. Heatster: A Database and Web Server for Identification and Classification of Heat Stress Transcription Factors in Plants. Bioinform. Biol. Insights 2019, 13. [Google Scholar] [CrossRef]
- O’Connor, D.J.; Wright, G.C.; Dieters, M.J.; George, D.L.; Hunter, M.N.; Tatnell, J.R.; Fleischfresser, D.B. Development and Application of Speed Breeding Technologies in a Commercial Peanut Breeding Program. Peanut Sci. 2013, 40, 107–114. [Google Scholar] [CrossRef]
- Alahmad, S.; Dinglasan, E.; Leung, K.M.; Riaz, A.; Derbal, N.; Voss-Fels, K.P.; Able, J.A.; Bassi, F.M.; Christopher, J.; Hickey, L.T. Speed breeding for multiple quantitative traits in durum wheat. Plant Methods 2018, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.D.; Asyraf Md Hatta, M.; Hinchliffe, A.; Steed, A.; Reynolds, D.; et al. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Chr | Number of Initial QTL Projected on the Map and Associated Traits | Number of MQTL Detected | Traits Clustered under Meta | CI (95%) | Position of Meta QTL (cM) | Closet Marker at the Left of MQTL with Genetic Distance from EXPEN2012 and Physical Position on SL4.0 in Bracket | Closest Marker at the Right of MQTL with Genetic Distance from EXPEN2012 and Physical Position on SL4.0 in Bracket | R2 (Average R2 of the of QTL Contained in the Meta QTL) |
|---|---|---|---|---|---|---|---|---|
| 1 | 13 (qSL1, qSP1, qFPI1, qIN1, qAL1, qPV1d, qSL1d, qPV1.2d1, qREC1.1, qREC1.2, qCC1) | 1 | qSL1, qSP1, qREC1.1 | 9.85 | 16.02 | solcap_snp_sl_60446 (10.41 cM, 0.885 Mb) | solcap_snp_sl_100802 (20.84 cM, 2.423 Mb) | 16.3 |
| 2 | qPV1d, qSL1d, qREC1.1, qREC1.2 | 8.99 | 28.19 | solcap_snp_sl_59944 (23.36 cM, 3.638 Mb) | solcap_snp_sl_18635 (33.74 cM, 67.407 Mb) | 9.8 | ||
| 3 | qFPI1, qIN1, qSL1d qREC1.1, qREC1.2 | 6.53 | 40.37 | solcap_snp_sl_50542 (36.27 cM, 68.647 Mb) | solcap_snp_sl_50427 (43.2 cM, 70,150 Mb) | 16.9 | ||
| 4 | qAL1, qSL1d, qREC1.1, qREC1.2 | 18.78 | 69.39 | solcap_snp_sl_34545 (59.99 cM, 77.219 Mb) | solcap_snp_sl_43796 (79.54 cM, 80.345 Mb) | 9.85 | ||
| 5 | qPV1.2d1, qREC1.2 | 23.93 | 104.97 | solcap_snp_sl_40177 (92.57 cM, 84.161 Mb) | solcap_snp_sl_53692 (116.87 cM, 90.442 Mb) | 7.35 | ||
| 2 | 9 (qAL2, qSL2, qSL2d1, qAL2d1, qAL2d, qPN2d, qPV2d1, qREC2 qCC2) | 1 | qSL2d1, qAL2d, qPV2d1, qAL2, qPN2d, qSL2, qREC2 qCC2 | 9.54 | 62.77 | solcap_snp_sl_29658 (56.88 cM, 41.069 Mb) | solcap_snp_sl_49645 (68.27 cM, 43.682 Mb) | 9.96 |
| 2 | qAL2, qSL2, qAL2.2d1, qREC2 | 9.46 | 86.87 | solcap_snp_sl_50166 (82.15cM, 47.751 Mb) | solcap_snp_sl_67138 (91.9cM, 49.894 Mb) | 13.51 | ||
| 3 | 3 (qSL3, qSP3, qPV3d) | 1 | qSL3, qSP3, qPV3d | 5.41 | 78.73 | solcap_snp_sl_62008 (75.64cM, 59.821 Mb) | solcap_snp_sl_100878 (80.69cM, 60.526 Mb) | 16.7 |
| 4 | 3 (qPV4d, qSP4d, qPV4.2d1) | 0 | - | - | - | - | - | - |
| 5 | 1 (qSP5, qFv/Fm5 | 0 | - | - | - | - | - | - |
| 7 | 1 (qSL7) | 0 | - | - | - | - | - | - |
| 8 | 1 (qSL8d1) | 0 | - | - | - | - | - | - |
| 9 | 2 (qAL9d) | 1 | qREC9, qAL9d | 123.34 | 56.99 | solcap_snp_sl_17545 (0 cM, 0.052 Mb) | solcap_snp_sl_63591 (96.67 cM, 68.42 Mb) | 8.92 |
| 11 | 3 (qPV11, qSP11d1, qSL11d1) | 1 | qPV11 | 8.17 | 19.44 | solcap_snp_sl_10611 (14.26cM, 2.024 Mb) | solcap_snp_sl_21767 (24.80 cM, 3.188 Mb) | 36.3 |
| 2 | SP11d1, SL11d1 | 47.05 | 67.46 | solcap_snp_sl_62864 (44cM, 6.119 Mb) | solcap_snp_sl_44931 (92.05cM, 53. 42 Mb) | 14.05 | ||
| 12 | 4 (qSL12d1, qSP12d1, qREC12, qFv/Fm12) | 1 | SP12d1, SL12d1, qREC12 | 21.53 | 28.88 | solcap_snp_sl_45733 (8.19cM, 0.724 Mb) | solcap_snp_sl_41194 (36.4 cM, 3.136 Mb) | 8.66 |
| 2 | qFv/Fm12 | 80.29 | 113.81 | solcap_snp_sl_40924 (22.58cM, 1.917 Mb) | solcap_snp_sl_31342 (99.06 cM, 66.42 Mb) | 8.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayenan, M.A.T.; Danquah, A.; Hanson, P.; Ampomah-Dwamena, C.; Sodedji, F.A.K.; Asante, I.K.; Danquah, E.Y. Accelerating Breeding for Heat Tolerance in Tomato (Solanum lycopersicum L.): An Integrated Approach. Agronomy 2019, 9, 720. https://doi.org/10.3390/agronomy9110720
Ayenan MAT, Danquah A, Hanson P, Ampomah-Dwamena C, Sodedji FAK, Asante IK, Danquah EY. Accelerating Breeding for Heat Tolerance in Tomato (Solanum lycopersicum L.): An Integrated Approach. Agronomy. 2019; 9(11):720. https://doi.org/10.3390/agronomy9110720
Chicago/Turabian StyleAyenan, Mathieu Anatole Tele, Agyemang Danquah, Peter Hanson, Charles Ampomah-Dwamena, Fréjus Ariel Kpêdétin Sodedji, Isaac K. Asante, and Eric Yirenkyi Danquah. 2019. "Accelerating Breeding for Heat Tolerance in Tomato (Solanum lycopersicum L.): An Integrated Approach" Agronomy 9, no. 11: 720. https://doi.org/10.3390/agronomy9110720