
Effects of Various Nitrogen Regimes on the Ability of Rapeseed (Brassica napus L.) to Suppress Littleseed Canarygrass (Phalaris minor Retz.)

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Experiment Design and Data Collection
2.3. Data Analyses
3. Results
3.1. The Impacts of Littleseed Canarygrass on the Growth and Yield of Rapeseed under Various Nitrogen Regimes
3.2. The Inhibitory Effects of Rapeseed on Littleseed Canarygrass under Various Nitrogen Regimes
3.3. Competitive Interactions of Rapeseed and Littleseed Canarygrass
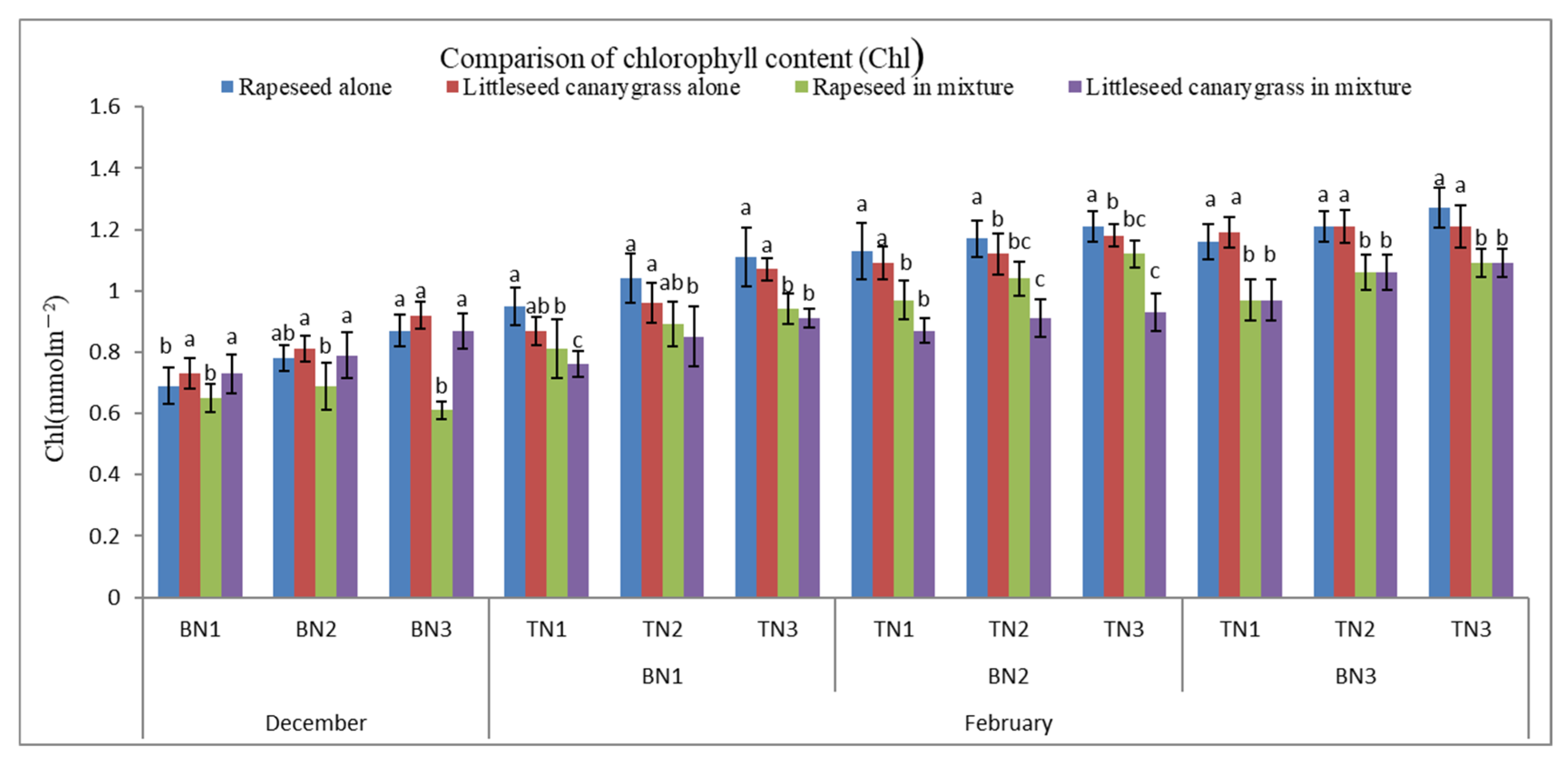
3.4. Photosynthetic Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simberloff, D.; Martin, J.-L.; Genovesi, P.; Maris, V.; Wardle, D.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackburn, T.M.; Essl, F.; Evans, T.; Hulme, P.E.; Jeschke, J.; Kühn, I.; Kumschick, S.; Marková, Z.; Mrugała, A.; Nentwig, W.; et al. A unified classification of alien species based on the magnitude of their environmental impacts. PLoS Biol. 2014, 12, e1001850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sher, A.A.; Marshall, D.L.; Taylor, J.P. Establishment patterns of native Populus and Salix in the presence of invasive nonnative Tamarix. Ecol. Appl. 2002, 12, 760–772. [Google Scholar] [CrossRef]
- Li, W.; Luo, J.; Tian, X.; Chow, W.S.; Sun, Z.; Zhang, T.; Peng, S.; Peng, C. A new strategy for controlling invasive weeds: Selecting valuable native plants to defeat them. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, S.; Xu, G.; Clements, D.R.; Jin, G.; Chen, A.; Zhang, F.; Kato-Noguchi, H. Suppression of the invasive plant mile-a-minute (Mikania micrantha) by local crop sweet potato (Ipomoea batatas) by means of higher growth rate and competition for soil nutrients. BMC Ecol. 2015, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Richardson, D.; Pyšek, P. Plant invasions: Merging the concepts of species invasiveness and community invasibility. Prog. Phys. Geogr. 2006, 30, 409–431. [Google Scholar] [CrossRef]
- Meng, Q.; Yue, S.; Hou, P.; Cui, Z.; Chen, X. Improving yield and nitrogen use efficiency simultaneously for maize and wheat in China: A Review. Pedosphere 2016, 26, 137–147. [Google Scholar] [CrossRef]
- Jinping, W.; Lijia, D.; Weiguo, S. Effects of different nitrogen regimes on competition between Ambrosia artemisiifolia, an invasive species, and two native species, Artemisia annua and Artemisia mongolica. Biodivers. Sci. 2012, 20, 3–11. [Google Scholar] [CrossRef]
- Huenneke, L.F.; Hamburg, S.P.; Koide, R.; Mooney, H.A.; Vitousek, P.M. Effects of soil resources on plant invasion and community structure in Californian serpentine grassland. Ecology 1990, 71, 478–491. [Google Scholar] [CrossRef]
- Brooks, M.L. Effects of increased soil nitrogen on the dominance of alien annual plants in the Mojave Desert. J. Appl. Ecol. 2003, 40, 344–353. [Google Scholar] [CrossRef]
- Gonzalez-Andujar, J.L.; Saavedra, M. Spatial distribution of annual grass weed populations in winter cereals. Crop Prot. 2003, 22, 629–633. [Google Scholar] [CrossRef]
- Singh, S.; Kirkwood, R.; Marshall, G. Biology and control of Phalaris minor Retz. (littleseed canarygrass) in wheat. Crop Prot. 1999, 18, 1–16. [Google Scholar] [CrossRef]
- Chhokar, R.S.; Sharma, R.K. Multiple herbicide resistance in littleseed canarygrass (Phalaris minor): A threat to wheat production in India. Weed Biol. Manag. 2008, 8, 112–123. [Google Scholar] [CrossRef]
- Shen, S.C.; Zhang, F.D.; Xu, G.F.; Li, T.L.; Wu, D.; Zhang, Y. Occurrence and infestation of invasive weed in crop field in Yunnan. Southwest China J. Agric. Sci. 2012, 25, 554–561. [Google Scholar]
- Xu, G.F.; Zhang, F.D.; Li, T.L.; Shen, S.C.; Zhang, Y.H. Influence of environmental factors on seed germination of Phalaris paradoxa and Phalaris minor. Acta Bot. Boreali-Occident Sin. 2011, 31, 1458–1465. [Google Scholar] [CrossRef]
- Xu, G.F.; Zhang, Y.; Shen, S.C.; Zhang, F.D.; Li, T.L.; Jin, G.M. Effect of crop species and mixed ratios on morphological plasticity and competitiveness of Phalaris minor Retz. Chin. J. Eco-Agric. 2013, 21, 1507–1514. [Google Scholar] [CrossRef]
- Xu, G.F.; Zhang, F.D.; Li, T.L.; Shan, Q.L.; Zhang, Y.H.; Wu, D. Biological characteristics, influence on growth of wheat and its economical threshold of Phalaris paradoxa L. and Phalaris minor Retz. Sci. Agric. Sin. 2010, 43, 4409–4417. [Google Scholar]
- Khawar, J.; Muhammad, F.; Mubshir, H.; Hafeezur, R.; Muhammad, A.A. Wild oat (Avena fatua L.) and canary grass (Phalaris minor Ritz.) management through allelopathy. J. Plant Prot. Res. 2010, 50, 41–44. [Google Scholar]
- Tang, D.S.; Yang, X.Y.; Li, Y.C.; Xiao, W.X.; Gu, X.J.; Jin, D.J.; Li, R.; Fu, Y. Investigation on the occurrence, damage and control of the invasive weed Phalaris spp. in Yunnan Province. Plant Prot. 2018, 44, 167–169. [Google Scholar]
- Travlos, I.S.; Chachalis, D. Glyphosate-resistant hairy fleabane (Conyza bonariensis) is reported in Greece. Weed Technol. 2010, 24, 569–573. [Google Scholar] [CrossRef]
- Xu, G.F.; Shen, S.C.; Zhang, Y.H.; Clements, D.R.; Yang, S.S.; Li, J.; Dong, L.Y.; Zhang, F.D. Designing cropping systems to improve the management of the invasive weed Phalaris minor Retz. Agronomy 2019, 9, 809. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.H.; Liu, L.; Fu, M.N. Studies on utilization of Yunnan germplasm resources in rapeseed. J. Anhui Agric. Sci. 2012, 40, 17056–17057. [Google Scholar]
- Lou, Q.F.; Zhang, D.Y.; Huang, J.Z.; Wang, Q.Y.; Liu, T.L.; Zhu, Y.L. Influence of nitrogen on competition between three weed species and bird rape. J. Nanjing Agric. Univ. 2000, 2, 23–26. [Google Scholar]
- Liu, B.; Tao, R.; Lu, J.W.; Li, X.K.; Cong, R.H.; Wu, L.S. On-farm trials of site-specific N management for maximum winter oilseed rape (Brassica napus L.) yield. J. Plant Nutr. 2017, 40, 1300–1311. [Google Scholar] [CrossRef]
- De Wit, C.T. On competition. Versl. Landbouwk. Onderzoek. 1960, 66, 1–82. [Google Scholar]
- Field, C.; Mooney, H.A. The Photosynthesis–Nitrogen Relationship in Wild Plants on the Economy of Plant Form and Function; Givnish, T.J., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 25–55. [Google Scholar]
- Pearcy, R.W.; Ehleringer, J.R.; Mooney, H.A.; Rundel, P.W. Plant Physiological Ecology: Field Methods and Instrumentation; Chapman & Hall: London, UK, 1989. [Google Scholar]
- Fowler, N. Competition and coexistence in a North Carolina grassland: III. mixtures of component species. J. Ecol. 1982, 70, 77–92. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.C.; Xu, G.F.; Li, D.Y.; Jin, G.M.; Liu, S.F.; Clements, D.R.; Yang, Y.S.; Rao, J.; Chen, A.D.; Zhang, F.D.; et al. Ipomoea batatas (sweet potato), a promising replacement control crop for the invasive alien plant Ageratina adenophora (Asteraceae) in China. Manag. Biol. Invasions 2019, 10, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.B. Shoot competition and root competition. J. Appl. Ecol. 1988, 25, 279–296. [Google Scholar] [CrossRef]
- Shivega, W.G.; Aldrich-Wolfe, L. Native plants fare better against an introduced competitor with native microbes and lower nitrogen availability. AoB Plants 2017, 9, plx004. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.-K.; Lee, S.-U.; Kozukue, N.; Levin, C.E.; Friedman, M. Distribution of phenolic compounds and antioxidative activities in parts of sweet potato (Ipomoea batatas L.) plants and in home processed roots. J. Food Compos. Anal. 2011, 24, 29–37. [Google Scholar] [CrossRef]
- Schjoerring, J.K.; Bock, J.G.H.; Gammelvind, L.; Jensen, C.R.; Mogensen, V.O. Nitrogen incorporation and remobilization in different shoot components of field-grown winter oilseed rape (Brassica napus L.) as affected by rate of nitrogen application and irrigation. Plant Soil 1995, 177, 255–264. [Google Scholar] [CrossRef]
- Lambers, H.; Poorter, H. Inherent variation in growth rate between higher plants: A search for physiological causes and ecological consequences. Adv. Ecol. Res. 1992, 23, 187–261. [Google Scholar] [CrossRef]
- Weigelt, A.; Jolliffe, P. Indices of plant competition. J. Ecol. 2003, 91, 707–720. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Treatments | Total Aboveground Biomass of Rapeseed (g·plant−1) | Rapeseed Yield (kg·ha−1) | Yield Reduction Rates of Rapeseed (%) | |||
|---|---|---|---|---|---|---|
| BN | TN | Monoculture | Mixed Culture | Monoculture | Mixed Culture | |
| BN1 | TN1 | 57.1 ± 0.45 I(a) | 36.6 ± 0.73 G(b) | 1405.3 ± 40.1 I(a) | 891.5 ± 29.6 G(b) | 36.34 ± 3.16 B |
| TN2 | 65.4 ± 0.91 H(a) | 46.4 ± 0.77 F(b) | 1678.5 ± 23.5 H(a) | 1101.5 ± 17.2 F(b) | 34.35 ± 1.16 B | |
| TN3 | 72.7 ± 0.54 G(a) | 54.1 ± 0.97 E(b) | 1843.5 ± 16.4 G(a) | 1228.8 ± 26.2 E(b) | 33.30 ± 2.04 B | |
| BN2 | TN1 | 78.9 ± 1.25 F(a) | 59.6 ± 0.59 D(b) | 2256.3 ± 19.9 F(a) | 1651.3 ± 32.5 C(b) | 26.79 ± 1.71 C |
| TN2 | 87.1 ± 0.46 E(a) | 70.7 ± 0.94 B(b) | 2471.4 ± 18.2 E(a) | 1874.8 ± 18.5 B(b) | 24.15 ± 0.30 C | |
| TN3 | 92.6 ± 0.97 D(a) | 76.9 ± 0.81 A(b) | 2623.3 ± 33.4 D(a) | 2021.3 ± 35.5 A(b) | 22.95 ± 0.91 C | |
| BN3 | TN1 | 96.8 ± 0.70 C(a) | 52.9 ± 0.78 E(b) | 2768.1 ± 21.8 C(a) | 1256.2 ± 23.9 E(b) | 54.60 ± 1.22 A |
| TN2 | 102.9 ± 1.09 B(a) | 63.4 ± 0.99 C(b) | 2864.2 ± 13.7 B(a) | 1318.1 ± 33.8 E(b) | 53.97 ± 1.28 A | |
| TN3 | 109.7 ± 1.00 A(a) | 75.6 ± 1.04 A(b) | 3024.4 ± 42.7 A(a) | 1434.2 ± 38.8 D(b) | 52.57 ± 1.25 A | |
| Treatments | Total Aboveground Biomass of Littleseed Canarygrass (g·plant−1) | Seed Number of Littleseed Canarygrass | Control Efficacy (%) | |||
|---|---|---|---|---|---|---|
| BN | TN | Monoculture | Mixed Culture | Monoculture | Mixed Culture | |
| BN1 | TN1 | 33.6 ± 0.79 I(a) | 17.4 ± 0.71 C(b) | 4171.3 ± 35.6 I(a) | 2094.3 ± 40.5 D(b) | 49.8 ± 0.73 F |
| TN2 | 36.9 ± 0.60 H(a) | 18.5 ± 0.63 C(b) | 4491.3 ± 34.4 H(a) | 1952.8 ± 41.8 E(b) | 56.5 ± 1.18 E | |
| TN3 | 41.0 ± 0.64 G(a) | 19.7 ± 0.75 C(b) | 4808.5 ± 66.4 G(a) | 1864.5 ± 67.1 E(b) | 61.2 ± 0.88 D | |
| BN2 | TN1 | 43.7 ± 0.78 F(a) | 17.6 ± 0.88 C(b) | 5227.8 ± 74.4 F(a) | 1732.8 ± 27.7 F(b) | 66.9 ± 0.63 C |
| TN2 | 47.0 ± 0.61 E(a) | 18.8 ± 0.60 C(b) | 5557.3 ± 92.5 E(a) | 1301.0 ± 29.5 G(b) | 76.6 ± 0.56 B | |
| TN3 | 49.4 ± 0.76 D(a) | 19.3 ± 0.92 C(b) | 5989.8 ± 103.4 D(a) | 1211.5 ± 33.9 G(b) | 79.8 ± 0.63 A | |
| BN3 | TN1 | 53.7 ± 0.53 C(a) | 35.3 ± 1.06 B(b) | 6223.3 ± 82.1 C(a) | 3314.8 ± 34.9 C(b) | 46.7 ± 0.73 G |
| TN2 | 59.0 ± 0.72 B(a) | 38.7 ± 0.63 A(b) | 6558.1 ± 66.1 B(a) | 3623.1 ± 30.18 B(b) | 45.2 ± 0.35 G | |
| TN3 | 62.3 ± 0.69 A(a) | 40.8 ± 0.86 A(b) | 6979.5 ± 133.9 A(a) | 3852.5 ± 52.0 A(b) | 44.8 ± 1.06 G | |
| BN | TN | Rapeseed RY | Littleseed Canarygrass RY | CB Index for Rapeseed |
|---|---|---|---|---|
| BN1 | TN1 | 0.641 ± 0.016 d** | 0.517 ± 0.009 b** | 0.216 ± 0.008 e** |
| TN2 | 0.709 ± 0.002 c** | 0.501 ± 0.012 bc** | 0.348 ± 0.015 d** | |
| TN3 | 0.744 ± 0.008 b** | 0.479 ± 0.014 c** | 0.440 ± 0.019 c** | |
| BN2 | TN1 | 0.756 ± 0.005 b** | 0.401 ± 0.013 d** | 0.635 ± 0.037 b** |
| TN2 | 0.812 ± 0.009 a** | 0.399 ± 0.008 d** | 0.710 ± 0.009 a** | |
| TN3 | 0.831 ± 0.004 a** | 0.391 ± 0.013 d** | 0.755 ± 0.031 a** | |
| BN3 | TN1 | 0.547 ± 0.007 e** | 0.658 ± 0.014 a** | −0.186 ± 0.012 h** |
| TN2 | 0.616 ± 0.007 d** | 0.655 ± 0.006 a** | −0.061 ± 0.005 g** | |
| TN3 | 0.690 ± 0.016 c** | 0.654 ± 0.008 a** | 0.052 ± 0.016 f** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, G.; Shen, S.; Zhang, Y.; Clements, D.R.; Yang, S.; Wen, L.; Zhang, F.; Dong, L. Effects of Various Nitrogen Regimes on the Ability of Rapeseed (Brassica napus L.) to Suppress Littleseed Canarygrass (Phalaris minor Retz.). Agronomy 2022, 12, 713. https://doi.org/10.3390/agronomy12030713
Xu G, Shen S, Zhang Y, Clements DR, Yang S, Wen L, Zhang F, Dong L. Effects of Various Nitrogen Regimes on the Ability of Rapeseed (Brassica napus L.) to Suppress Littleseed Canarygrass (Phalaris minor Retz.). Agronomy. 2022; 12(3):713. https://doi.org/10.3390/agronomy12030713
Chicago/Turabian StyleXu, Gaofeng, Shicai Shen, Yun Zhang, David Roy Clements, Shaosong Yang, Lina Wen, Fudou Zhang, and Liyao Dong. 2022. "Effects of Various Nitrogen Regimes on the Ability of Rapeseed (Brassica napus L.) to Suppress Littleseed Canarygrass (Phalaris minor Retz.)" Agronomy 12, no. 3: 713. https://doi.org/10.3390/agronomy12030713