
Nitrogen Mineralization, Soil Microbial Biomass and Extracellular Enzyme Activities Regulated by Long-Term N Fertilizer Inputs: A Comparison Study from Upland and Paddy Soils in a Red Soil Region of China

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Experimental Setup
2.2. Sampling and Laboratory Analysis
2.3. Soil Chemical Analyses
2.4. Soil Incubation and N Mineralization Measurements
2.5. Statistical Analysis
3. Results
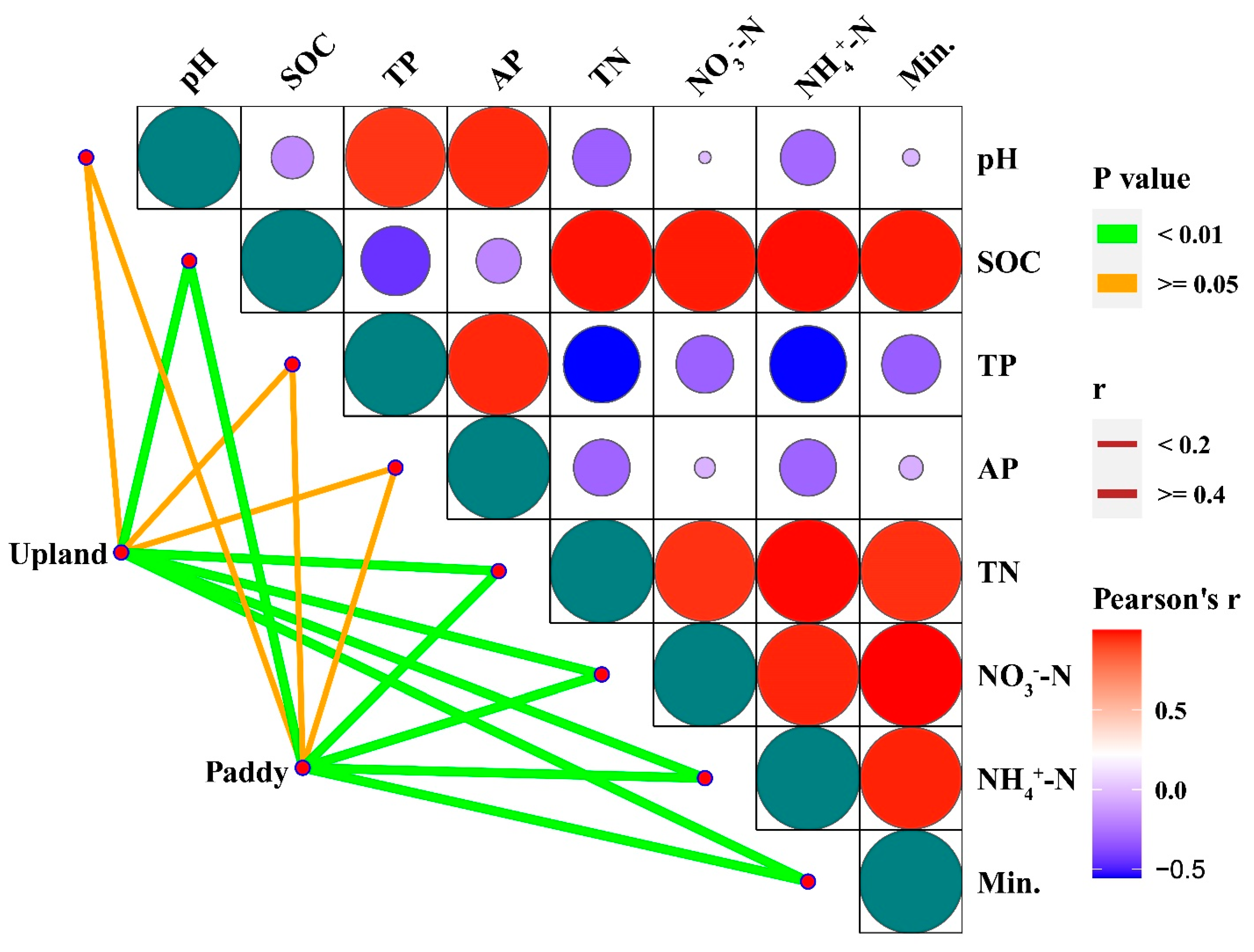
3.1. Soil Chemical Properties and Nutrient Content
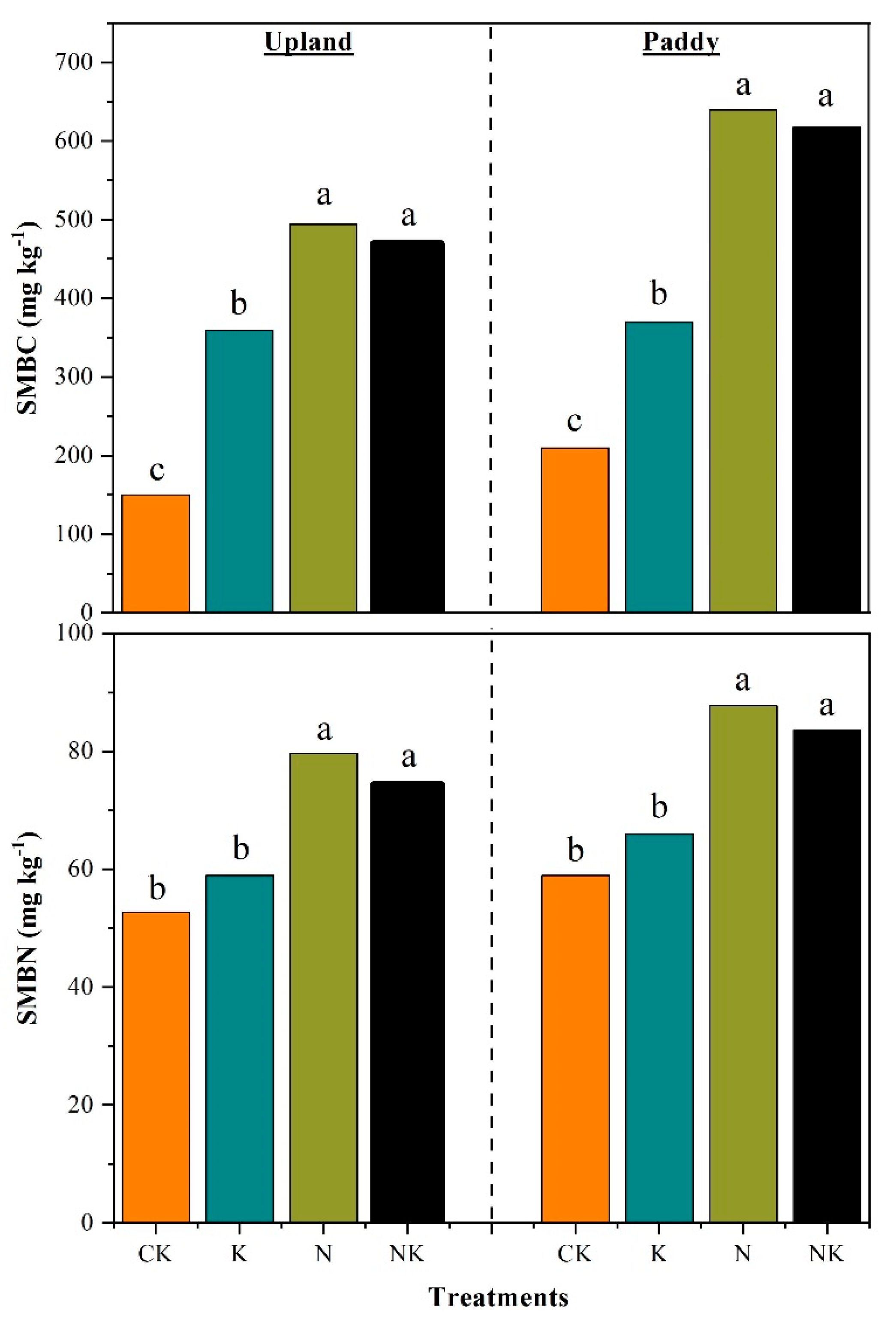
3.2. Responses of Soil Microbial Biomass Nitrogen and Carbon to Nitrogen Additions
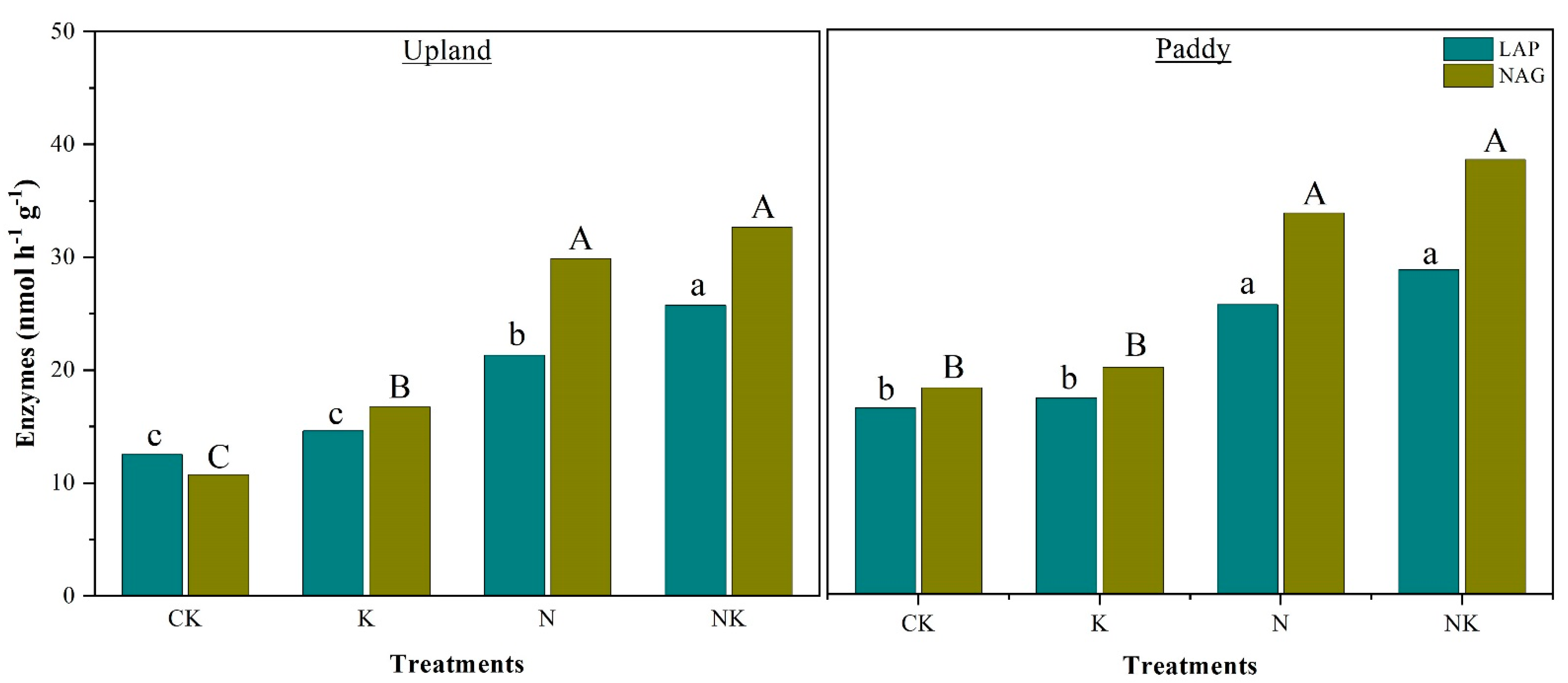
3.3. Soil Enzymatic Activities Associated with Soil N Mineralization
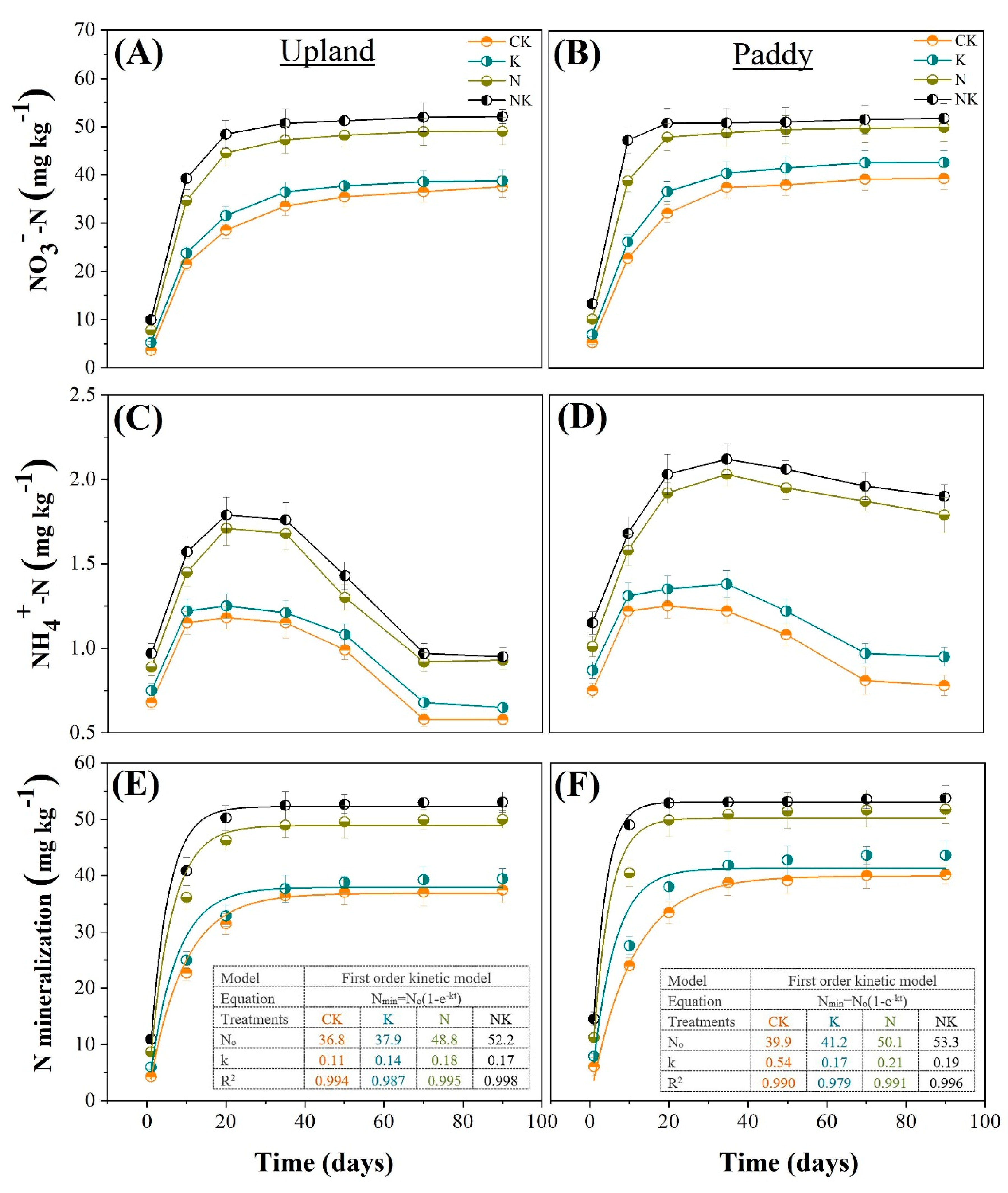
3.4. Soil NO3−-N, NH4+-N Contents during Incubation Period
3.5. N Mineralization Potential and Mineralization Rate Constant (k)
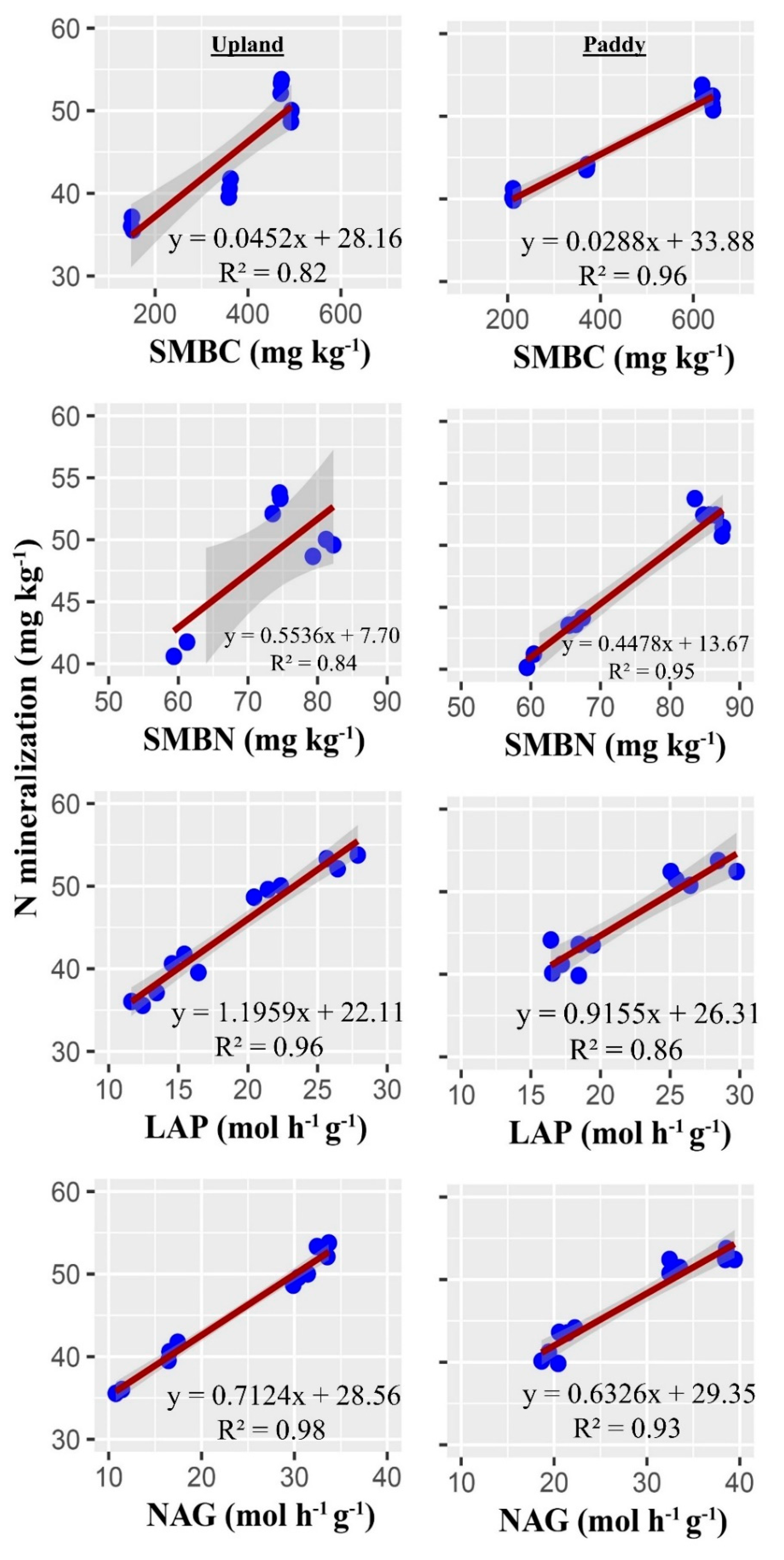
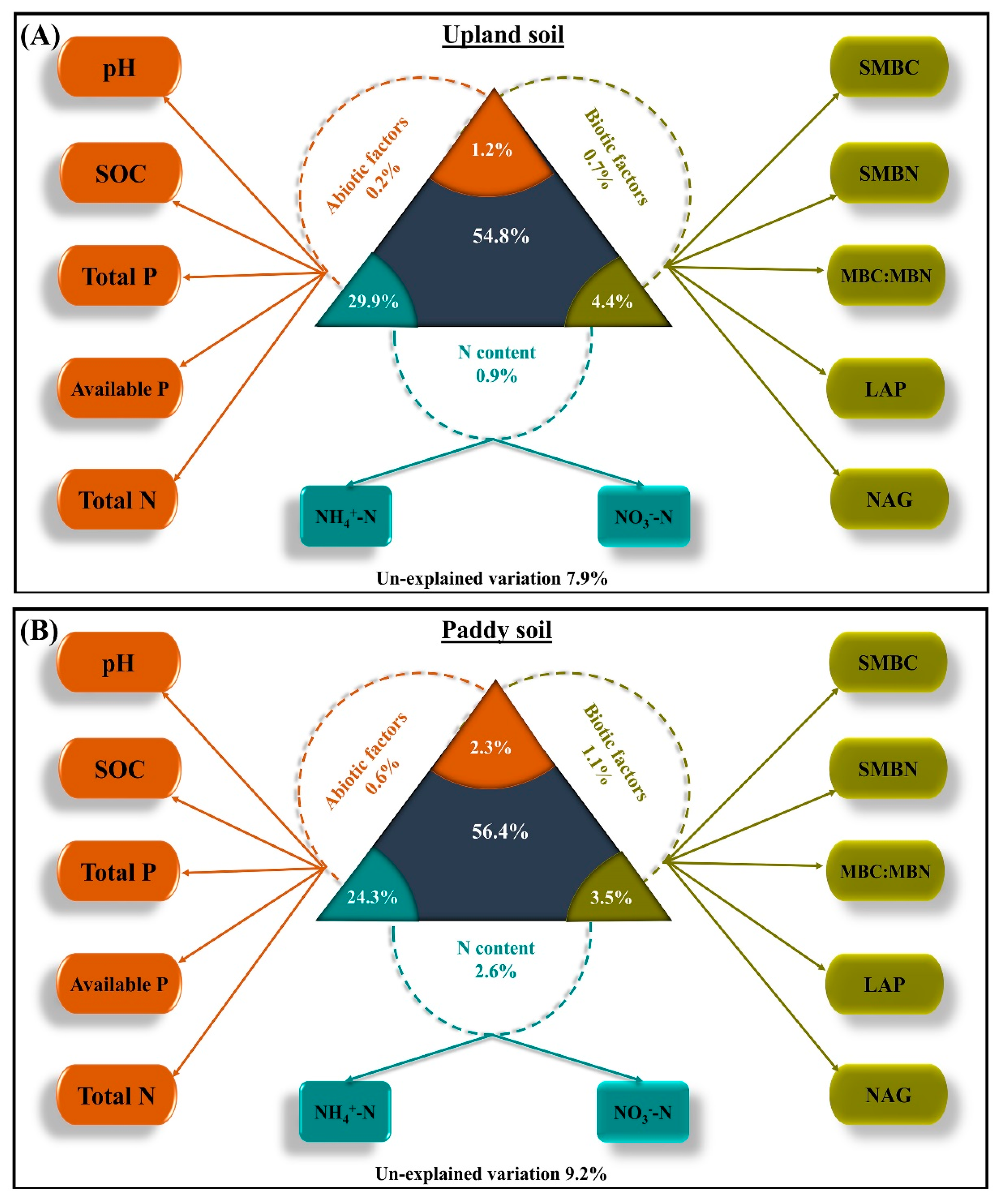
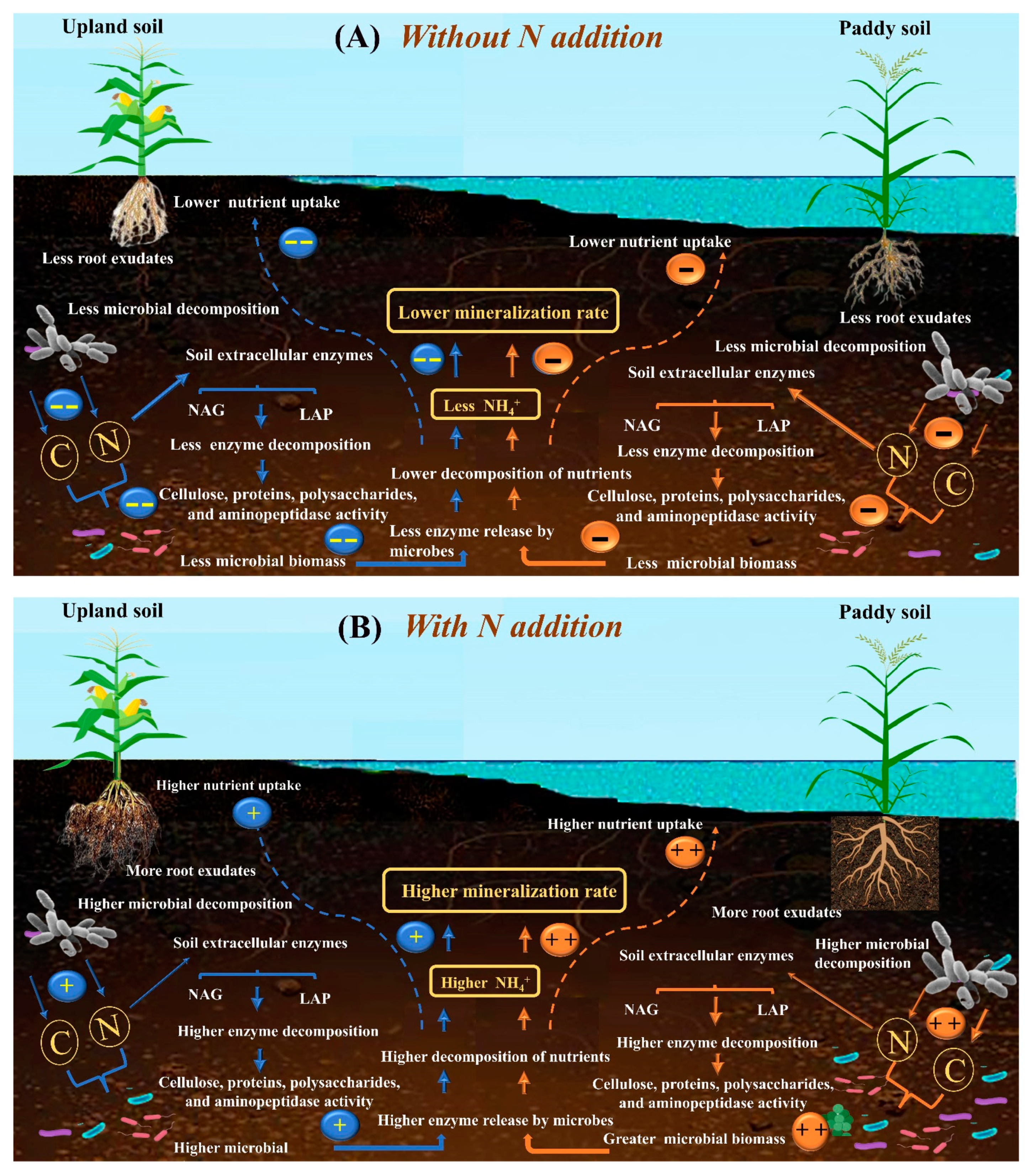
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stanford, G.; Smith, S.J. Nitrogen Mineralization Potentials of Soils. Soil Sci. Soc. Am. J. 1972, 36, 465–472. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, M.; Wang, X.; Huang, Q.; Nie, J.; Li, Z.; Li, S.; Hwang, S.W.; Lee, K.B. Effects of organic amendments on soil carbon sequestration in paddy fields of subtropical China. J. Soils Sediments 2012, 12, 457–470. [Google Scholar] [CrossRef]
- Ali, S.; Dongchu, L.; Jing, H.; Ahmed, W.; Abbas, M.; Qaswar, M.; Anthonio, C.K.; Lu, Z.; Boren, W.; Yongmei, X.; et al. Soil microbial biomass and extracellular enzymes regulate nitrogen mineralization in a wheat-maize cropping system after three decades of fertilization in a Chinese Ferrosol. J. Soils Sediments 2021, 21, 281–294. [Google Scholar] [CrossRef]
- Smith, S.J.; Young, L.B.; Miller, G.E. Evaluation of Soil Nitrogen Mineralization Potentials under Modified Field Conditions. Soil Sci. Soc. Am. J. 1977, 41, 74–76. [Google Scholar] [CrossRef]
- Mikha, M.M.; Rice, C.W.; Benjamin, J.G. Estimating Soil Mineralizable Nitrogen under Different Management Practices. Soil Sci. Soc. Am. J. 2006, 70, 1522–1531. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.; Dong, H.; Zhang, R.; Li, S. Excess molar enthalpies of methylformate + (1-propanol, 2-propanol, 1-butanol, 2-butanol and 1-pentanol) at T = 298.15 K, p = (5.0, 10.0) MPa, and methylformate + 1-propanol at T = 333.15 K, p = 10.0 MPa. Thermochim. Acta 2006, 443, 125–128. [Google Scholar] [CrossRef]
- Bai, J.; Gao, H.; Xiao, R.; Wang, J.; Huang, C. A Review of Soil Nitrogen Mineralization as Affected by Water and Salt in Coastal Wetlands: Issues and Methods. Clean Soil Air Water 2012, 40, 1099–1105. [Google Scholar] [CrossRef]
- Cui, J.; Zhu, Z.; Xu, X.; Liu, S.; Jones, D.L.; Kuzyakov, Y.; Shibistova, O.; Wu, J.; Ge, T. Carbon and nitrogen recycling from microbial necromass to cope with C:N stoichiometric imbalance by priming. Soil Biol. Biochem. 2020, 142, 107720. [Google Scholar] [CrossRef]
- Xu, Y.; Ding, F.; Gao, X.; Wang, Y.; Li, M.; Wang, J. Mineralization of plant residues and native soil carbon as affected by soil fertility and residue type. J. Soils Sediments 2019, 19, 1407–1415. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Xu, X. Competition between roots and microorganisms for nitrogen: Mechanisms and ecological relevance. New Phytol. 2013, 198, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Razavi, B.S.; Hu, Y.; Xu, X.; Zhu, Z.; Liu, Y.; Kuzyakov, Y.; Li, Y.; Wu, J.; Ge, T. C/P stoichiometry of dying rice root defines the spatial distribution and dynamics of enzyme activities in root-detritusphere. Biol. Fertil. Soils 2019, 55, 251–263. [Google Scholar] [CrossRef]
- Wessels Perelo, L.; Jimenez, M.; Munch, J.C. Microbial immobilisation and turnover of 15N labelled substrates in two arable soils under field and laboratory conditions. Soil Biol. Biochem. 2006, 38, 912–922. [Google Scholar] [CrossRef]
- Geisseler, D.; Horwath, W.R.; Joergensen, R.G.; Ludwig, B. Pathways of nitrogen utilization by soil microorganisms—A review. Soil Biol. Biochem. 2010, 42, 2058–2067. [Google Scholar] [CrossRef]
- Manzoni, S.; Porporato, A. Soil carbon and nitrogen mineralization: Theory and models across scales. Soil Biol. Biochem. 2009, 41, 1355–1379. [Google Scholar] [CrossRef]
- Li, P.; Yang, Y.; Han, W.; Fang, J. Global patterns of soil microbial nitrogen and phosphorus stoichiometry in forest ecosystems. Glob. Ecol. Biogeogr. 2014, 23, 979–987. [Google Scholar] [CrossRef]
- Zhu, J.; Li, M.; Whelan, M. Phosphorus activators contribute to legacy phosphorus availability in agricultural soils: A review. Sci. Total Environ. 2018, 612, 522–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filip, Z. International approach to assessing soil quality by ecologically-related biological parameters. Agric. Ecosyst. Environ. 2002, 88, 169–174. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Chen, J.; Chen, D.; Xu, Q.; Fuhrmann, J.J.; Li, L.; Pan, G.; Li, Y.; Qin, H.; Liang, C.; Sun, X. Organic carbon quality, composition of main microbial groups, enzyme activities, and temperature sensitivity of soil respiration of an acid paddy soil treated with biochar. Biol. Fertil. Soils 2019, 55, 185–197. [Google Scholar] [CrossRef]
- Ahmed, W.; Qaswar, M.; Jing, H.; Wenjun, D.; Geng, S.; Kailou, L.; Ying, M.; Ao, T.; Mei, S.; Chao, L.; et al. Tillage practices improve rice yield and soil phosphorus fractions in two typical paddy soils. J. Soils Sediments 2020, 20, 850–861. [Google Scholar] [CrossRef]
- Schmidt, I.K.; Michelsen, A.; Jonasson, S. Effects of labile soil carbon on nutrient partitioning between an arctic graminoid and microbes. Oecologia 1997, 112, 557–565. [Google Scholar] [CrossRef]
- Kaschuk, G.; Alberton, O.; Hungria, M. Three decades of soil microbial biomass studies in Brazilian ecosystems: Lessons learned about soil quality and indications for improving sustainability. Soil Biol. Biochem. 2010, 42, 1–13. [Google Scholar] [CrossRef]
- Chantigny, M.H.; Angers, D.A.; Prévost, D.; Simard, R.R.; Chalifour, F.P. Dynamics of soluble organic C and C mineralization in cultivated soils with varying N fertilization. Soil Biol. Biochem. 1999, 31, 543–550. [Google Scholar] [CrossRef]
- Georg Joergensen, R.; Scheu, S. Response of soil microorganisms to the addition of carbon, nitrogen and phosphorus in a forest Rendzina. Soil Biol. Biochem. 1999, 31, 859–866. [Google Scholar] [CrossRef]
- Padhan, K.; Bhattacharjya, S.; Sahu, A.; Manna, M.C.; Sharma, M.P.; Singh, M.; Wanjari, R.H.; Sharma, R.P.; Sharma, G.K.; Patra, A.K. Soil N transformation as modulated by soil microbes in a 44 years long term fertilizer experiment in a sub-humid to humid Alfisol. Appl. Soil Ecol. 2020, 145, 103355. [Google Scholar] [CrossRef]
- Nannipieri, P.; Trasar-Cepeda, C.; Dick, R.P. Soil enzyme activity: A brief history and biochemistry as a basis for appropriate interpretations and meta-analysis. Biol. Fertil. Soils 2018, 54, 11–19. [Google Scholar] [CrossRef]
- Zhao, F.Z.; Ren, C.J.; Han, X.H.; Yang, G.H.; Wang, J.; Doughty, R. Changes of soil microbial and enzyme activities are linked to soil C, N and P stoichiometry in afforested ecosystems. For. Ecol. Manag. 2018, 427, 289–295. [Google Scholar] [CrossRef]
- Bowles, T.M.; Acosta-Martínez, V.; Calderón, F.; Jackson, L.E. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biol. Biochem. 2014, 68, 252–262. [Google Scholar] [CrossRef]
- Liang, Y.; Li, D.; Lu, X.; Yang, X.; Pan, X.; Mu, H.; Shi, D.; Zhang, B. Soil erosion changes over the past five decades in the red soil region of Southern China. J. Mt. Sci. 2010, 7, 92–99. [Google Scholar] [CrossRef]
- Yan, X.; Zhou, H.; Zhu, Q.H.; Wang, X.F.; Zhang, Y.Z.; Yu, X.C.; Peng, X. Carbon sequestration efficiency in paddy soil and upland soil under long-term fertilization in southern China. Soil Tillage Res. 2013, 130, 42–51. [Google Scholar] [CrossRef]
- Pan, G.; Xu, X.; Smith, P.; Pan, W.; Lal, R. An increase in topsoil SOC stock of China’s croplands between 1985 and 2006 revealed by soil monitoring. Agric. Ecosyst. Environ. 2010, 136, 133–138. [Google Scholar] [CrossRef]
- Yuan, H.; Ge, T.; Zou, S.; Wu, X.; Liu, S.; Zhou, P.; Chen, X.; Brookes, P.; Wu, J. Effect of land use on the abundance and diversity of autotrophic bacteria as measured by ribulose-1,5-biphosphate carboxylase/oxygenase (RubisCO) large subunit gene abundance in soils. Biol. Fertil. Soils 2013, 49, 609–616. [Google Scholar] [CrossRef]
- Sheng, R.; Meng, D.; Wu, M.; Di, H.; Qin, H.; Wei, W. Effect of agricultural land use change on community composition of bacteria and ammonia oxidizers. J. Soils Sediments 2013, 13, 1246–1256. [Google Scholar] [CrossRef]
- Wu, X.; Ge, T.; Wang, W.; Yuan, H.; Wegner, C.E.; Zhu, Z.; Whiteley, A.S.; Wu, J. Cropping systems modulate the rate and magnitude of soil microbial autotrophic CO2 fixation in soil. Front. Microbiol. 2015, 6, 379. [Google Scholar] [CrossRef] [Green Version]
- Ullah, S.; Ai, C.; Huang, S.; Zhang, J.; Jia, L.; Ma, J.; Zhou, W.; He, P. The responses of extracellular enzyme activities and microbial community composition under nitrogen addition in an upland soil. PLoS ONE 2019, 14, e0223026. [Google Scholar] [CrossRef] [Green Version]
- Treseder, K.K. Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies. Ecol. Lett. 2008, 11, 1111–1120. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Padre, A.T.; Sato, C.; Shiono, H.; Hattori, S.; Kajihara, A.; Aoyama, M.; Tawaraya, K.; Kumagai, K. Changes in the soil C and N contents, C decomposition and N mineralization potentials in a rice paddy after long-term application of inorganic fertilizers and organic matter. Soil Sci. Plant Nutr. 2016, 62, 212–219. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zhang, W.; Gurmesa, G.A.; Zhu, X.; Li, D.; Mo, J. Phosphorus addition affects soil nitrogen dynamics in a nitrogen-saturated and two nitrogen-limited forests. Eur. J. Soil Sci. 2017, 68, 472–479. [Google Scholar] [CrossRef]
- Chen, F.S.; Niklas, K.J.; Liu, Y.; Fang, X.M.; Wan, S.Z.; Wang, H. Nitrogen and phosphorus additions alter nutrient dynamics but not resorption efficiencies of Chinese fir leaves and twigs differing in age. Tree Physiol. 2015, 35, 1106–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbin, J.D.; Avis, P.G.; Wilbur, R.B. The role of phosphorus availability in the response of soil nitrogen cycling, understory vegetation and arbuscular mycorrhizal inoculum potential to elevated nitrogen inputs. Water Air Soil Pollut. 2003, 147, 141–162. [Google Scholar] [CrossRef]
- Homeier, J.; Hertel, D.; Camenzind, T.; Cumbicus, N.L.; Maraun, M.; Martinson, G.O.; Poma, L.N.; Rillig, M.C.; Sandmann, D.; Scheu, S.; et al. Tropical Andean Forests Are Highly Susceptible to Nutrient Inputs-Rapid Effects of Experimental N and P Addition to an Ecuadorian Montane Forest. PLoS ONE 2012, 7, e47128. [Google Scholar] [CrossRef] [PubMed]
- Martinson, G.O.; Corre, M.D.; Veldkamp, E. Responses of nitrous oxide fluxes and soil nitrogen cycling to nutrient additions in montane forests along an elevation gradient in southern Ecuador. Biogeochemistry 2013, 112, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ge, T.; Zhu, Z.; Liu, S.; Luo, Y.; Li, Y.; Wang, P.; Gavrichkova, O.; Xu, X.; Wang, J.; et al. Carbon input and allocation by rice into paddy soils: A review. Soil Biol. Biochem. 2019, 133, 97–107. [Google Scholar] [CrossRef]
- Page, A.L.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis Part 2: Chemical and Mineralogical Properties. Agronomy Monograph No. 9 ASA-SSSA Madison, Wisconsin; Soil Sci. Soc. Am. Inc.: Madison, WI, USA, 1982; p. 1159. [Google Scholar]
- Black, C.A. Methods of Soil Analysis; Mongraph 9; American Society of Agronomy: Madison, WI, USA, 1965. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Sugawara, E.; Nikaido, H. Properties of AdeABC and AdeIJK efflux systems of Acinetobacter baumannii compared with those of the AcrAB-TolC system of Escherichia coli. Antimicrob. Agents Chemother. 2014, 58, 7250–7257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Wu, J.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation-extraction-an automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Jenkinson, D.S. Determination of microbial biomass carbon and nitrogen in soil. Adv. Nitrogen Cycl. Agric. Ecosyst. 1988, 368–386. [Google Scholar]
- DeForest, J.L. The influence of time, storage temperature, and substrate age on potential soil enzyme activity in acidic forest soils using MUB-linked substrates and l-DOPA. Soil Biol. Biochem. 2009, 41, 1180–1186. [Google Scholar] [CrossRef]
- Peng, X.; Wang, W. Stoichiometry of soil extracellular enzyme activity along a climatic transect in temperate grasslands of northern China. Soil Biol. Biochem. 2016, 98, 74–84. [Google Scholar] [CrossRef]
- Zhang, Q.; Miao, F.; Wang, Z.; Shen, Y.; Wang, G. Effects of long-term fertilization management practices on soil microbial biomass in China’s cropland: A meta-analysis. Agron. J. 2017, 109, 1183–1195. [Google Scholar] [CrossRef]
- Tian, J.; Lu, S.; Fan, M.; Li, X.; Kuzyakov, Y. Integrated management systems and N fertilization: Effect on soil organic matter in rice-rapeseed rotation. Plant Soil 2013, 372, 53–63. [Google Scholar] [CrossRef]
- Wei, W.; Yan, Y.; Cao, J.; Christie, P.; Zhang, F.; Fan, M. Effects of combined application of organic amendments and fertilizers on crop yield and soil organic matter: An integrated analysis of long-term experiments. Agric. Ecosyst. Environ. 2016, 225, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Ye, G.; Lin, Y.; Liu, D.; Chen, Z.; Luo, J.; Bolan, N.; Fan, J.; Ding, W. Long-term application of manure over plant residues mitigates acidification, builds soil organic carbon and shifts prokaryotic diversity in acidic Ultisols. Appl. Soil Ecol. 2019, 133, 24–33. [Google Scholar] [CrossRef]
- DeForest, J.L.; Zak, D.R.; Pregitzer, K.S.; Burton, A.J. Atmospheric nitrate deposition and the microbial degradation of cellobiose and vanillin in a northern hardwood forest. Soil Biol. Biochem. 2004, 36, 965–971. [Google Scholar] [CrossRef]
- Hyvönen, R.; Persson, T.; Andersson, S.; Olsson, B.; Ågren, G.I.; Linder, S. Impact of long-term nitrogen addition on carbon stocks in trees and soils in northern Europe. Biogeochemistry 2008, 89, 121–137. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Burton, A.J.; Zak, D.R.; Talhelm, A.F. Simulated chronic nitrogen deposition increases carbon storage in Northern Temperate forests. Glob. Chang. Biol. 2008, 14, 142–153. [Google Scholar] [CrossRef]
- Waldrop, M.P.; Firestone, M.K. Altered utilization patterns of young and old soil C by microorganisms caused by temperature shifts and N additions. Biogeochemistry 2004, 67, 235–248. [Google Scholar] [CrossRef]
- Zhang, J.; Qin, J.; Yao, W.; Bi, L.; Lai, T.; Yu, X. Effect of long-term application of manure and mineral fertilizers on nitrogen mineralization and microbial biomass in paddy soil during rice growth stages. Plant Soil Environ. 2009, 55, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Six, J.; Conant, R.T.; Paul, E.A.; Paustian, K. Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils. Plant Soil 2002, 241, 155–176. [Google Scholar] [CrossRef]
- Zhou, P.; Pan, G.X.; Spaccini, R.; Piccolo, A. Molecular changes in particulate organic matter (POM) in a typical Chinese paddy soil under different long-term fertilizer treatments. Eur. J. Soil Sci. 2010, 61, 231–242. [Google Scholar] [CrossRef]
- Wang, W.J.; Baldock, J.A.; Dalal, R.C.; Moody, P.W. Decomposition dynamics of plant materials in relation to nitrogen availability and biochemistry determined by NMR and wet-chemical analysis. Soil Biol. Biochem. 2004, 36, 2045–2058. [Google Scholar] [CrossRef]
- Dinesh, R.; Srinivasan, V.; Hamza, S.; Manjusha, A.; Kumar, P.S. Short-term effects of nutrient management regimes on biochemical and microbial properties in soils under rainfed ginger (Zingiber officinale Rosc.). Geoderma 2012, 173–174, 192–198. [Google Scholar] [CrossRef]
- Kaur, T.; Brar, B.S.; Dhillon, N.S. Soil organic matter dynamics as affected by long-term use of organic and inorganic fertilizers under maize-wheat cropping system. Nutr. Cycl. Agroecosyst. 2008, 81, 59–69. [Google Scholar] [CrossRef]
- Geisseler, D.; Scow, K.M. Long-term effects of mineral fertilizers on soil microorganisms—A review. Soil Biol. Biochem. 2014, 75, 54–63. [Google Scholar] [CrossRef]
- Singh, J.S.; Gupta, V.K. Soil microbial biomass: A key soil driver in management of ecosystem functioning. Sci. Total Environ. 2018, 634, 497–500. [Google Scholar] [CrossRef]
- Dijkstra, F.A.; Hobbie, S.E.; Reich, P.B.; Knops, J.M.H. Divergent effects of elevated CO2, N fertilization, and plant diversity on soil C and N dynamics in a grassland field experiment. Plant Soil 2005, 272, 41–52. [Google Scholar] [CrossRef]
- Quan, Z.; Huang, B.; Lu, C.; Shi, Y.; Chen, X.; Zhang, H.; Fang, Y. The fate of fertilizer nitrogen in a high nitrate accumulated agricultural soil. Sci. Rep. 2016, 5, 21539. [Google Scholar] [CrossRef] [PubMed]
- Mooshammer, M.; Wanek, W.; Zechmeister-Boltenstern, S.; Richter, A. Stoichiometric imbalances between terrestrial decomposer communities and their resources: Mechanisms and implications of microbial adaptations to their resources. Front. Microbiol. 2014, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G. Leucine aminopeptidase, β-glucosidase and alkaline phosphatase activity rates and their significance in nutrient cycles in some coastal Mediterranean sites. Mar. Drugs 2010, 8, 916–940. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, B.A. Enzyme activities as a component of soil biodiversity: A review. Pedobiologia 2005, 49, 637–644. [Google Scholar] [CrossRef]
- Stone, M.M.; Weiss, M.S.; Goodale, C.L.; Adams, M.B.; Fernandez, I.J.; German, D.P.; Allison, S.D. Temperature sensitivity of soil enzyme kinetics under N-fertilization in two temperate forests. Glob. Chang. Biol. 2012, 18, 1173–1184. [Google Scholar] [CrossRef]
- Sharma, V.; Mir, S.H.; Sharma, A. Nitrogen Mineralization as Influenced by Different Organic Manures in an Inceptisol in the Foothill Himalayas. Commun. Soil Sci. Plant Anal. 2016, 47, 194–202. [Google Scholar] [CrossRef]
- Benedetti, A.; Sebastiani, G. Determination of potentially mineralizable nitrogen in agricultural soil. Biol. Fertil. Soils 1996, 21, 114–120. [Google Scholar] [CrossRef]
- Savin, M.C.; Görres, J.H.; Neher, D.A.; Amador, J.A. Uncoupling of carbon and nitrogen mineralization: Role of microbivorous nematodes. Soil Biol. Biochem. 2001, 33, 1463–1472. [Google Scholar] [CrossRef]
- Frampton, C.M.; Zaman, M.; Di, H.J.; Cameron, K.C. Gross nitrogen mineralization and nitrification rates and their relationships to enzyme activities and the soil microbial biomass in soils treated with dairy shed effluent and ammonium fertilizer at different water potentials. Biol. Fertil. Soils 1999, 29, 178–186. [Google Scholar] [CrossRef]
- Schloter, M.; Nannipieri, P.; Sørensen, S.J.; van Elsas, J.D. Microbial indicators for soil quality. Biol. Fertil. Soils 2018, 54, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, S.; Nayak, A.K.; Kumar, A.; Tripathi, R.; Shahid, M.; Bhattacharyya, P.; Raja, R.; Panda, B.B. Carbon and nitrogen mineralization kinetics in soil of rice-rice system under long term application of chemical fertilizers and farmyard manure. Eur. J. Soil Biol. 2013, 58, 113–121. [Google Scholar] [CrossRef]
- Zaman, M.; Matsushima, M.; Chang, S.X.; Inubushi, K.; Nguyen, L.; Goto, S.; Kaneko, F.; Yoneyama, T. Nitrogen mineralization, N2O production and soil microbiological properties as affected by long-term applications of sewage sludge composts. Biol. Fertil. Soils 2004, 40, 101–109. [Google Scholar] [CrossRef]
- Sharifi, M.; Zebarth, B.J.; Miller, J.J.; Burton, D.L.; Grant, C.A. Soil nitrogen mineralization in a soil with long-term history of fresh and composted manure containing straw or wood-chip bedding. Nutr. Cycl. Agroecosyst. 2014, 99, 63–78. [Google Scholar] [CrossRef]
- Zaman, M.; Di, H.J.; Cameron, K.C. A field study of gross rates of N mineralization and nitrification and their relationships to microbial biomass and enzyme activities in soils treated with dairy effluent and ammonium fertilizer. Soil Use Manag. 1999, 15, 188–194. [Google Scholar] [CrossRef]
- Bengtsson, G.; Bengtson, P.; Månsson, K.F. Gross nitrogen mineralization-, immobilization-, and nitrification rates as a function of soil C/N ratio and microbial activity. Soil Biol. Biochem. 2003, 35, 143–154. [Google Scholar] [CrossRef]
- Tung, P.G.A.; Yusoff, M.K.; Majid, N.M.; Joo, G.K.; Huang, G.H. Effect of N and K fertilizers on nutrient leaching and groundwater quality under mature oil palm in Sabah during the monsoon period. Am. J. Appl. Sci. 2009. [Google Scholar] [CrossRef]
- Zhang, F.; Niu, J.; Zhang, W.; Chen, X.; Li, C.; Yuan, L.; Xie, J. Potassium nutrition of crops under varied regimes of nitrogen supply. Plant Soil 2010, 335, 21–34. [Google Scholar] [CrossRef]
- Zhu, Z.L.; Chen, D.L. Nitrogen fertilizer use in China—Contributions to food production, impacts on the environment and best management strategies. Nutr. Cycl. Agroecosyst. 2002, 63, 117–127. [Google Scholar] [CrossRef]
- Sahrawat, K.L. Fertility and organic matter in submerged rice soils. Curr. Sci. 2005, 88, 735–739. [Google Scholar]
- Chen, H.; Li, D.; Xiao, K.; Wang, K. Soil microbial processes and resource limitation in karst and non-karst forests. Funct. Ecol. 2018, 32, 1400–1409. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, Y.; Gao, D.; Wang, X.; Liu, W.; Deng, J.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Ecoenzymatic stoichiometry and nutrient dynamics along a revegetation chronosequence in the soils of abandoned land and Robinia pseudoacacia plantation on the Loess Plateau, China. Soil Biol. Biochem. 2019, 134, 1–14. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, Y.; Chen, Y.; Zhang, J.; Li, H.; Wang, L.; Chen, Q. Short-term warming shifts microbial nutrient limitation without changing the bacterial community structure in an alpine timberline of the eastern Tibetan Plateau. Geoderma 2020, 360, 113985. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Treatments | Fertilization | N Status | |
|---|---|---|---|---|
| N | K | |||
| Upland | CK | 0 | 0 | Without N |
| K | 0 | 60 | Without N | |
| N | 60 | 0 | With N | |
| NK | 60 | 60 | With N | |
| Paddy | CK | 0 | 0 | Without N |
| K | 0 | 75 | Without N | |
| N | 90 | 0 | With N | |
| NK | 90 | 75 | With N | |
| Enzyme | Abb. | Substrate | EC | Substrate Conc. (nM) | Inc. Time |
|---|---|---|---|---|---|
| Leucine-aminopeptidase | LAP | L-Leucine-7-amido-4-methylcoumarin hydrochloride | 3.4.11.1 | 400 | 45 min |
| N-acetyl-b-D-glucosaminidase | NAG | 4-Methylumbelliferyl N-acetyl-b-D-glucosaminide | 3.2.1.30 | 300 | 4 h |
| Sites | Treatments | pH | SOC (g kg−1) | Total P (g kg−1) | Averaged Total P (g kg−1) | Available P (mg kg−1) | Total N (g kg−1) |
|---|---|---|---|---|---|---|---|
| Upland | CK | 4.91 ± 0.16 | 8.54 ± 0.23 f | 1.64 ± 0.02 | 1.08A | 10.7 ± 1.08 | 0.10 ± 0.12 d |
| Paddy | CK | 5.22 ± 0.17 | 13.3 ± 1.73 b | 0.51 ± 0.02 | 6.71 ± 0.65 | 1.54 ± 0.04 b | |
| Upland | K | 4.93 ± 0.26 | 9.76 ± 1.24 e | 1.54 ± 0.01 | 1.02A | 9.74 ± 0.84 | 0.16 ± 0.07 d |
| Paddy | K | 5.25 ± 0.24 | 14.2 ± 0.98 b | 0.50 ± 0.06 | 6.79 ± 0.33 | 1.55 ± 0.07 b | |
| Upland | N | 4.27 ± 0.12 | 12.9 ± 1.11 cd | 1.38 ± 0.03 | 0.92B | 9.37 ± 0.22 | 1.25 ± 0.02 c |
| Paddy | N | 5.13 ± 0.20 | 20.8 ± 1.02 a | 0.46 ± 0.02 | 6.44 ± 0.37 | 2.18 ± 0.02 a | |
| Upland | NK | 4.34 ± 0.13 | 12.6 ± 2.05 d | 1.39 ± 0.02 | 0.92B | 9.46 ± 0.76 | 1.28 ± 0.03 c |
| Paddy | NK | 5.15 ± 0.16 | 20.5 ± 0.75 a | 0.45 ± 0.06 | 6.42 ± 0.88 | 2.14 ± 0.08 a | |
| ANOVA | |||||||
| Soil type | ns | *** | *** | *** | *** | ||
| Fertilization | ns | *** | *** | ns | *** | ||
| S × F | ns | *** | Ns | ns | *** | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, S.; Liu, K.; Ahmed, W.; Jing, H.; Qaswar, M.; Kofi Anthonio, C.; Maitlo, A.A.; Lu, Z.; Liu, L.; Zhang, H. Nitrogen Mineralization, Soil Microbial Biomass and Extracellular Enzyme Activities Regulated by Long-Term N Fertilizer Inputs: A Comparison Study from Upland and Paddy Soils in a Red Soil Region of China. Agronomy 2021, 11, 2057. https://doi.org/10.3390/agronomy11102057
Ali S, Liu K, Ahmed W, Jing H, Qaswar M, Kofi Anthonio C, Maitlo AA, Lu Z, Liu L, Zhang H. Nitrogen Mineralization, Soil Microbial Biomass and Extracellular Enzyme Activities Regulated by Long-Term N Fertilizer Inputs: A Comparison Study from Upland and Paddy Soils in a Red Soil Region of China. Agronomy. 2021; 11(10):2057. https://doi.org/10.3390/agronomy11102057
Chicago/Turabian StyleAli, Sehrish, Kailou Liu, Waqas Ahmed, Huang Jing, Muhammad Qaswar, Christian Kofi Anthonio, Ali Akbar Maitlo, Zhang Lu, Lisheng Liu, and Huimin Zhang. 2021. "Nitrogen Mineralization, Soil Microbial Biomass and Extracellular Enzyme Activities Regulated by Long-Term N Fertilizer Inputs: A Comparison Study from Upland and Paddy Soils in a Red Soil Region of China" Agronomy 11, no. 10: 2057. https://doi.org/10.3390/agronomy11102057