
Connecting the Lab and the Field: Genome Analysis of Phyllobacterium and Rhizobium Strains and Field Performance on Two Vegetable Crops

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome Analysis
2.2. Quorum Sensing Assays
2.3. Field Experiments
3. Results
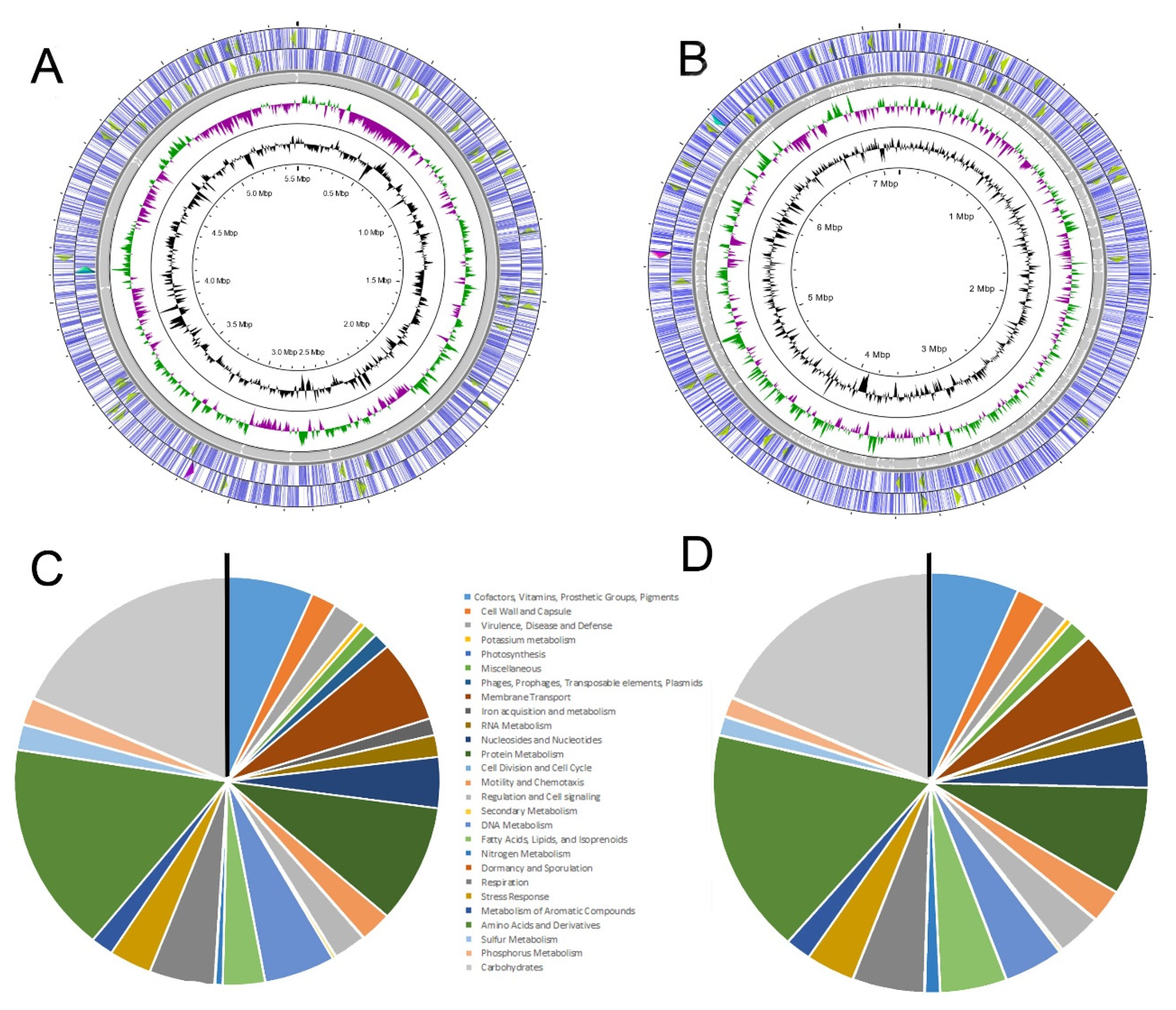
3.1. Genome features
3.2. Analysis of Genes Involved in Quorum Sensing and Plant Colonization
3.3. Analysis of Genes Involved in Plant Growth Promotion
3.4. Field Trials
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.S.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, S.; Kabir, S.; Shabbir, U.; Batool, R. Plant growth promoting rhizobacteria in sustainable agriculture: From theoretical to pragmatic approach. Symbiosis 2019, 78, 115–123. [Google Scholar] [CrossRef]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Kandel, S.L.; Joubert, P.M.; Doty, S.L. Bacterial endophyte colonization and distribution within plants. Microorganisms 2017, 5, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [Green Version]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Omomowo, O.I.; Babalola, O.O. Bacterial and Fungal Endophytes: Tiny Giants with Immense Beneficial Potential for Plant Growth and Sustainable Agricultural Productivity. Microorganisms 2019, 7, 481. [Google Scholar] [CrossRef] [Green Version]
- ALKahtani, M.D.F.; Fouda, A.; Attia, K.A.; Al-Otaibi, F.; Eid, A.M.; Ewais, E.D.; Hijri, M.; St-Arnaud, M.; Hassan, S.D.; Khan, N.; et al. Isolation and Characterization of plant growth promoting endophytic bacteria from desert plants and their application as bioinoculants for sustainable agriculture. Agronomy 2020, 10, 1325. [Google Scholar] [CrossRef]
- Krishnamoorthy, A.; Agarwal, T.; Kotamreddy, J.N.R.; Bhattacharya, R.; Mitra, A.; Maiti, T.K.; Maiti, M.K. Impact of seed-transmitted endophytic bacteria on intra- and inter-cultivar plant growth promotion modulated by certain sets of metabolites in rice crop. Microbiol. Res. 2020, 241, 126582. [Google Scholar] [CrossRef]
- Aeron, A.; Dubey, R.C.; Maheshwari, D.K. Characterization of a plant-growth-promoting non-nodulating endophytic bacterium (Stenotrophomonas maltophilia) from the root nodules of Mucuna utilis var. capitata L. (Safed Kaunch). Can. J. Microbiol. 2020, 66, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Aeron, A.; Maheshwari, D.K.; Meena, V.S. Endophytic bacteria promote growth of the medicinal legume Clitoria ternatea L. by chemotactic activity. Arch. Microbiol. 2020, 202, 1049–1058. [Google Scholar] [CrossRef]
- Dahmani, M.A.; Desrut, A.; Moumen, B.; Verdon, J.; Mermouri, L.; Kacem, M.; Coutos-Thévenot, P.; Kaid-Harche, M.; Bergès, T.; Vriet, C. Unearthing the plant growth-promoting traits of Bacillus megaterium RmBm31, an endophytic bacterium isolated from root nodules of Retama monosperma. Front. Plant Sci. 2020, 11, 124. [Google Scholar] [CrossRef]
- Deng, Z.S.; Kong, Z.Y.; Zhang, B.C.; Zhao, L.F. Insights into non-symbiotic plant growth promotion bacteria associated with nodules of Sphaerophysa salsula growing in northwestern China. Arch. Microbiol. 2020, 202, 399–409. [Google Scholar] [CrossRef]
- Soares, R.; Trejo, J.; Lorite, M.J.; Figueira, E.; Sanjuán, J.; Videira, E.; Castro, I. Diversity, phylogeny and plant growth promotion traits of nodule associated bacteria isolated from Lotus parviflorus. Microorganisms 2020, 8, 499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapia-García, E.Y.; Hernández-Trejo, V.; Guevara-Luna, J.; Rojas-Rojas, F.U.; Arroyo-Herrera, I.; Meza-Radilla, G.; Vásquez-Murrieta, M.S.; Estrada-de Los Santos, P. Plant growth-promoting bacteria isolated from wild legume nodules and nodules of Phaseolus vulgaris L. trap plants in central and southern Mexico. Microbiol. Res. 2020, 239, 126522. [Google Scholar] [CrossRef]
- Tokgöz, S.; Lakshman, D.K.; Ghozlan, M.H.; Pinar, H.; Roberts, D.P.; Mitra, A. Soybean Nodule-Associated Non-Rhizobial Bacteria Inhibit Plant Pathogens and Induce Growth Promotion in Tomato. Plants 2020, 9, 1494. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.A.; Lou, Y.; Hafeez, R.; Li, X.; Hossain, A.; Xie, T.; Lin, L.; Li, B.; Yin, Y.; Yan, J.; et al. Functional analysis and genome mining reveal high potential of biocontrol and plant growth promotion in nodule-inhabiting bacteria within Paenibacillus polymyxa Complex. Front. Microbiol. 2021, 11, 618601. [Google Scholar] [CrossRef]
- Noori, F.; Etesami, H.; Noori, S.; Forouzan, E.; Salehi Jouzani, G.; Malboobi, M.A. Whole genome sequence of Pantoea agglomerans ANP8, a salinity and drought stress-resistant bacterium isolated from alfalfa (Medicago sativa L.) root nodules. Biotechnol. Rep. 2021, 29, e00600. [Google Scholar] [CrossRef]
- Velázquez, E.; Carro, L.; Flores-Félix, J.D.; Martínez-Hidalgo, P.; Menéndez, E.; Ramírez-Bahena, M.H.; Mulas, R.; González-Andrés, F.; Martínez-Molina, E.; Peix, A. The Legume Nodule Microbiome: A Source of Plant Growth-Promoting Bacteria. In Probiotics and Plant Health; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017; pp. 41–70. [Google Scholar]
- Cardoso, P.; Alves, A.; Silveira, P.; Sá, C.; Fidalgo, C.; Freitas, R.; Figueira, E. Bacteria from nodules of wild legume species: Phylogenetic diversity, plant growth promotion abilities and osmotolerance. Sci. Total Environ. 2018, 645, 1094–1102. [Google Scholar] [CrossRef]
- Brígido, C.; Menéndez, E.; Paço, A.; Glick, B.R.; Belo, A.; Félix, M.R.; Oliveira, S.; Carvalho, M. Mediterranean native leguminous plants: A reservoir of endophytic bacteria with potential to enhance chickpea growth under stress conditions. Microorganisms 2019, 7, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ríos-Ruiz, W.F.; Valdez-Nuñez, R.A.; Bedmar, E.J.; Castellano-Hinojosa, A. Utilization of Endophytic Bacteria Isolated from Legume Root Nodules for Plant Growth Promotion. In Field Crops: Sustainable Management by PGPR; Maheshwari, D., Dheeman, S., Eds.; Springer: Cham, Switzerland, 2019; Volume 23, pp. 145–176. [Google Scholar]
- Paterson, J.; Jahanshah, G.; Li, Y.; Wang, Q.; Mehnaz, S.; Gross, H. The contribution of genome mining strategies to the understanding of active principles of PGPR strains. FEMS Microbiol. Ecol. 2017, 93, 1–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, A.; Salas Gonzalez, I.; Mittelviefhaus, M.; Clingenpeel, S.; Herrera Paredes, S.; Miao, J.; Wang, K.; Devescovi, G.; Stillman, K.; Monteiro, F.; et al. Genomic features of bacterial adaptation to plants. Nat. Genet. 2018, 50, 138–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruto, M.; Prigent-Combaret, C.; Muller, D.; Moënne-Loccoz, Y. Analysis of genes contributing to plant-beneficial functions in plant growth-promoting rhizobacteria and related Proteobacteria. Sci. Rep. 2015, 4, 6261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, F.X.; Hernandez, A.G.; Glick, B.R.; Rossi, M.J. The extreme plant-growth-promoting properties of Pantoea phytobeneficialis MSR2 revealed by functional and genomic analysis. Environ. Microbiol. 2020, 22, 1341–1355. [Google Scholar] [CrossRef]
- Jiménez-Gómez, A.; Saati-Santamaría, Z.; Kostovcik, M.; Rivas, R.; Velázquez, E.; Mateos, P.F.; Menéndez, E.; García-Fraile, P. Selection of the Root Endophyte Pseudomonas brassicacearum CDVBN10 as Plant Growth Promoter for Brassica napus L. Crops. Agronomy 2020, 10, 1788. [Google Scholar] [CrossRef]
- Flores, A.; Diaz-Zamora, J.T.; Orozco-Mosqueda, M.C.; Chávez, A.; de los Santos-Villalobos, S.; Valencia-Cantero, E.; Santoyo, G. Bridging genomics and field research: Draft genome sequence of Bacillus thuringiensis CR71, an endophytic bacterium that promotes plant growth and fruit yield in Cucumis sativus L. 3 Biotech 2020, 10, 220. [Google Scholar] [CrossRef]
- Pastor-Bueis, R.; Sánchez-Cañizares, C.; James, E.K.; González-Andrés, F. Formulation of a highly effective inoculant for common bean based on an autochthonous elite strain of Rhizobium leguminosarum bv. phaseoli, and genomic-based insights into its agronomic performance. Front. Microbiol. 2019, 10, 2724. [Google Scholar] [CrossRef]
- Flores-Félix, J.D.; Carro, L.; Velázquez, E.; Valverde, Á.; Cerda-Castillo, E.; García-Fraile, P.; Rivas, R. Phyllobacterium endophyticum sp. nov., isolated from nodules of Phaseolus vulgaris. Int. J. Syst. Evol. Microbiol. 2013, 63, 821–826. [Google Scholar] [CrossRef] [Green Version]
- Flores-Félix, J.D.; Sánchez-Juanes, F.; García-Fraile, P.; Valverde, A.; Mateos, P.F.; Gónzalez-Buitrago, J.M.; Velázquez, E.; Rivas, R. Phaseolus vulgaris is nodulated by the symbiovar viciae of several genospecies of Rhizobium laguerreae complex in a Spanish region where Lens culinaris is the traditionally cultivated legume. Syst. Appl. Microbiol. 2019, 42, 240–247. [Google Scholar] [CrossRef]
- Flores-Félix, J.D.; Menéndez, E.; Rivera, L.P.; Marcos-García, M.; Martínez-Hidalgo, P.; Mateos, P.F.; Martínez-Molina, E.; Velázquez, E.; García-Fraile, P.; Rivas, R. Use of Rhizobium leguminosarum as a potential biofertilizer for Lactuca sativa and Daucus carota crops. J. Plant Nutr. Soil Sci. 2013, 176, 876–882. [Google Scholar] [CrossRef]
- Flores-Félix, J.D.; Silva, L.R.; Rivera, L.P.; Marcos-García, M.; García-Fraile, P.; Martínez-Molina, E.; Mateos, P.F.; Velázquez, E.; Andrade, P.; Rivas, R. Plants probiotics as a tool to produce highly functional fruits: The case of Phyllobacterium and vitamin C in strawberries. PLoS ONE 2015, 10, e0122281. [Google Scholar] [CrossRef]
- Celador-Lera, L.; Flores-Félix, J.D.; Marcos-García, M.; Rivera, L.P.; Martinez-Hidalgo, P.; Menéndez, E.; Martinez-Molina, E.; Mateos, P.F.; Velázquez, E.; Rivas, R. Aplicación de bacterias endófiticas de vegetales cómo potenciales inoculantes de cultivos de zanahoria y lechuga. In Proceedings of the IV Congreso de Microbiología Industrial y Biotecnología Microbiana, Salamanca, Spain, 14–16 November 2012. [Google Scholar]
- Flores-Félix, J.D.; Menéndez, E.; Marcos-García, M.; Mateos, P.F.; Martínez-Molina, E.; Velázquez, M.E.; Rivas, R. PGPR-based biofertilizers increase carrot production. New Biotechnol. 2016, 33, 430–431. [Google Scholar] [CrossRef]
- Beringer, J.E. R factors transfer in Rhizobium leguminosarum. J. Gen. Microbiol. 1974, 84, 188–198. [Google Scholar] [CrossRef] [Green Version]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Haft, D.H.; DiCuccio, M.; Badretdin, A.; Brover, V.; Chetvernin, V.; O’Neill, K.; Li, W.; Chitsaz, F.; Derbyshire, M.K.; Gonzales, N.R.; et al. RefSeq: An update on prokaryotic genome annotation and curation. Nucleic Acids Res. 2018, 46, D851–D860. [Google Scholar] [CrossRef] [PubMed]
- Aramaki, T.; Blanc-Mathieu, R.; Endo, H.; Ohkubo, K.; Kanehisa, M.; Goto, S.; Ogata, H. KofamKOALA: KEGG ortholog assignment based on profile HMM and adaptive score threshold. Bioinformatics 2019, 19, 2251–2252. [Google Scholar] [CrossRef] [Green Version]
- Grant, J.R.; Stothard, P. The CGView Server: A comparative genomics tool for circular genomes. Nucleic Acids Res. 2008, 36, W181–W184. [Google Scholar] [CrossRef]
- Petkau, A.; Stuart-Edwards, M.; Stothard, P.; Van Domselaar, G. Interactive microbial genome visualization with GView. Bioinformatics 2010, 26, 3125–3126. [Google Scholar] [CrossRef]
- Vincent, J.M. The cultivation, isolation and maintenance of Rhizobia. In A Manual for the Practical Study of the Root-Nodule Bacteria. IBP Handbook 15; Black Well Scientific Publications: Oxford, UK, 1970; pp. 1–13. [Google Scholar]
- Marenda, M.; Zanardo, M.; Trovato, A.; Seno, F.; Squartini, A. Modeling quorum sensing trade-offs between bacterial cell density and system extension from open boundaries. Sci. Rep. 2016, 6, 39142. [Google Scholar] [CrossRef]
- Augimeri, R.V.; Varley, A.J.; Strap, J.L. Establishing a role for bacterial cellulose in environmental interactions: Lessons learned from diverse biofilm-producing Proteobacteria. Front. Microbiol. 2015, 6, 1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohan, R.; Benton, M.; Dangelmaier, E.; Fu, Z.; Chandra Sekhar, A. Quorum Sensing and biofilm formation in pathogenic and mutualistic plant-bacterial interactions. In Implication of Quorum Sensing System in Biofilm Formation and Virulence; Veera Bramhachari, P., Ed.; Springer: Singapore, 2018; pp. 133–160. [Google Scholar]
- Römling, U.; Galperin, M.Y. Bacterial cellulose biosynthesis: Diversity of operons, subunits, products, and functions. Trends Microbiol. 2015, 23, 545–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ormeño-Orrillo, E.; Menna, P.; Almeida, L.G.; Ollero, F.J.; Nicolás, M.F.; Pains Rodrigues, E.; Shigueyoshi Nakatani, A.; Silva Batista, J.S.; Oliveira Chueire, L.M.; Souza, R.C.; et al. Genomic basis of broad host range and environmental adaptability of Rhizobium tropici CIAT 899 and Rhizobium sp. PRF 81 which are used in inoculants for common bean (Phaseolus vulgaris L.). BMC Genom. 2012, 13, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robledo, M.; Rivera, L.; Jiménez-Zurdo, J.I.; Rivas, R.; Dazzo, F.; Velázquez, E.; Martínez-Molina, E.; Hirsch, A.M.; Mateos, P.F. Role of Rhizobium endoglucanase CelC2 in cellulose biosynthesis and biofilm formation on plant roots and abiotic surfaces. Microb. Cell Factories 2012, 11, 125. [Google Scholar] [CrossRef] [Green Version]
- Downie, J.; González, J. Cell-to-Cell communication in rhizobia: Quorum Sensing and Plant Signaling. In Chemical Communication among Bacteria; Winans, S., Bassler, B., Eds.; ASM Press: Washington, DC, USA, 2008; pp. 213–232. [Google Scholar]
- Calatrava-Morales, N.; McIntosh, M.; Soto, M.J. Regulation mediated by N-Acyl homoserine lactone quorum sensing signals in the Rhizobium-Legume symbiosis. Genes 2018, 9, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, J.; Zhou, L.; Wu, G.; Zhao, Y.; Jiang, T.; Liu, F. The AHL Quorum-sensing System negatively regulates growth and autolysis in Lysobacter brunescens. Front. Microbiol. 2019, 10, 2748. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen–phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, H.; Fraga, R.; Gonzalez, T.; Bashan, Y. Genetics of phosphate solubilization and its potential applications for improving plant growth-promoting bacteria. Plant Soil 2006, 287, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd Allah, E.F.; Hashem, A. Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar] [CrossRef]
- Duca, D.R.; Glick, B.R. Indole-3-acetic acid biosynthesis and its regulation in plant-associated bacteria. Appl. Microbiol. Biotechnol. 2020, 104, 8607–8619. [Google Scholar] [CrossRef] [PubMed]
- Duca, D.; Lorv, J.; Patten, C.L.; Rose, D.; Glick, B.R. Indole-3-acetic acid in plant–microbe interactions. Antonie Leeuwenhoek 2014, 106, 85–125. [Google Scholar] [CrossRef] [PubMed]
- Carson, K.C.; Meyer, J.M.; Dilworth, M.J. Hydroxamate siderophores of root nodule bacteria. Soil Biol. Biochem. 2000, 32, 11–21. [Google Scholar] [CrossRef]
- Sahin, U.; Ekinci, M.; Kiziloglu, F.M.; Yildirim, E.; Turan, M.; Kotan, R.; Ors, S. Ameliorative effects of plant growth promoting bacteria on water-yield relationships, growth, and nutrient uptake of lettuce plants under different irrigation levels. Hortscience 2015, 50, 1379–1386. [Google Scholar] [CrossRef] [Green Version]
- Cipriano, M.A.; Lupatini, M.; Lopes-Santos, L.; da Silva, M.J.; Roesch, L.F.; Destéfano, S.A.; Freitas, S.S.; Kuramae, E.E. Lettuce and rhizosphere microbiome responses to growth promoting Pseudomonas species under field conditions. FEMS Microbiol. Ecol. 2016, 92, fiw197. [Google Scholar] [CrossRef] [Green Version]
- Venancio, W.S.; Marcolino Gomes, J.; Shigueyoshi Nakatani, A.; Hungria, M.; Silva Araujo, R. Lettuce production under reduced levels of N-fertilizer in the presence of plant growth-promoting Bacillus spp. bacteria. J. Pure Appl. Microbiol. 2019, 13, 1941–1952. [Google Scholar] [CrossRef] [Green Version]
- Vetrano, F.; Miceli, C.; Angileri, V.; Frangipane, B.; Moncada, A.; Miceli, A. Effect of bacterial inoculum and fertigation management on nursery and field production of lettuce plants. Agronomy 2020, 10, 1477. [Google Scholar] [CrossRef]


{kind=link}
| Location | Remondo | Ciudad Rodrigo | |
|---|---|---|---|
| Crop | Carrot | Lettuce | |
| Latitude | 41°20′00.9″ N | 40°35′02.6″ N | |
| Longitude | 4°29′30.7″ W | 6°31′56.1″ W | |
| Texture (%) | Sand | 93.1 | 30.6 |
| Silt | 2.3 | 50.3 | |
| Clay | 4.6 | 19.1 | |
| pH 1:2 | (soil: water) | 7.0 | 7.6 |
| Electric conductivity | (dS/m) | 0.15 | 0.25 |
| Organic matter | (%) | 0.11 | 5.44 |
| Total nitrogen * | (%) | 0.01 | 0.35 |
| Ratio C/N | 4.3 | 9.0 | |
| Lime | (%) | 0.29 | 3.64 |
| P-Olsen | (mg kg−1) | 33.0 | 280.0 |
| K | (cmol (+) kg−1) | 0.25 | 1.36 |
| Ca | (cmol (+) kg−1) | 4.10 | 23.6 |
| Mg | (cmol (+) kg−1) | 1.09 | 5.11 |
| Na | (cmol (+) kg−1) | 0.69 | 0.13 |
| Location | Month | Temperatures (°C) * | Monthly Rainfall (mm) | ||||
|---|---|---|---|---|---|---|---|
| Hmax (°C) | Havg (°C) | Tavg (°C) | Lavg (°C) | Lmin (°C) | |||
| Remondo (Segovia station) (Carrot crop) | May | 23.0 | 16.0 | 10.3 | 5.1 | 0.4 | 69.8 |
| June | 31.0 | 23.9 | 16.7 | 9.5 | 3.3 | 7.0 | |
| July | 34.6 | 31.3 | 23.6 | 15.9 | 9.8 | 9.2 | |
| August | 35.8 | 30.3 | 22.4 | 14.4 | 8.8 | 0.4 | |
| September | 31.9 | 25.5 | 18.9 | 12.2 | 5.8 | 27.4 | |
| October | 25.4 | 19.6 | 14.4 | 9.2 | 0.7 | 32.4 | |
| Ciudad Rodrigo (Saelices el Chico station) (Lettuce crop) | June | 34.0 | 26.5 | 18.5 | 10.5 | 8.8 | 18.2 |
| July | 38.5 | 33.3 | 23.9 | 14.5 | 8.7 | 0.6 | |
| August | 38.9 | 32.6 | 23.2 | 13.9 | 8.7 | 0.0 | |
| September | 34.2 | 28.0 | 20.1 | 12.1 | 7.5 | 119 | |
| Genome Data | 1 | 2 |
|---|---|---|
| DDBJ/EMBL/GenBank accession number | VSZT00000000 | VSZV00000000 |
| Sequence size (bp) | 5,505,652 | 7,182,427 |
| Number of contigs | 15 | 356 |
| GC content (%) | 58.30 | 57.20 |
| Shortest contig size | 322 | 252 |
| Longest contig size | 2,119,450 | 557,230 |
| N50 value | 332,658 | 437,109 |
| L50 value | 5 | 74 |
| Number of coding sequences | 5152 | 7108 |
| Number of RNAs (tRNAs, rRNAs, others RNAs) | 52 (45, 3, 4) | 53 (44, 5, 4) |
| Crop/Treatment | Biomass per Plant (g) | Expected Yield (t ha−1) | |||
|---|---|---|---|---|---|
| Fresh | Dry | ||||
| Carrot (Roots) | |||||
| Uninoculated control | 61.82 (±2.63) | b | 6.40 (±0.25) | b | 31.5 |
| PEPV16 | 75.03 (±3.39) | a | 7.57 (±0.32) | a | 38.3 |
| PEPV15 | 79.02 (±2.79) | a | 8.01 (±0.24) | a | 40.3 |
| Mean square | 4376.998 | 37.335 | |||
| F-value and significance | 9.288 | *** | 9.280 | *** | |
| Lettuce (shoots) | |||||
| Uninoculated control | 1289 (±42) | b | 65.4 (±2.1) | b | 25.8 |
| PEPV15 | 1432 (±44) | a | 74.8 (±2.6) | a | 28.6 |
| PEPV16 | 1507 (±25) | a | 82.8 (±2.3) | a | 30.1 |
| Mean square | 331,459.370 | 2048.907 | |||
| F-value and significance | 8.496 | *** | 13.695 | *** | |
| N (g/100g) | P (mg/kg) | K (mg/kg) | Mg (mg/kg) | Ca (mg/kg) | Fe (mg/kg) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Carrot (roots) | ||||||||||||
| Uninoculated control | 1.04 (±0.044) | a | 2127 (±91) | b | 24,547 (±923) | b | 1058 (±46) | c | 2742 (±154) | b | 116 (±14) | a |
| PEPV15 | 1.10 (±0.008) | a | 2529 (±27) | a | 30,886 (±645) | a | 1244 (±17) | b | 3325 (±42) | a | 78 (±5) | a |
| PEPV16 | 1.12 (±0.062) | a | 2839 (±111) | a | 27,909 (±966) | ab | 1414 (±48) | a | 3248 (±116) | a | 116 (±11) | a |
| Mean Square | 0.007 | 509,536.343 | 40,234,943.659 | 127,293.063 | 401,076.976 | 1967.711 | ||||||
| F value and significance | 0.840 | ns | 17.957 | *** | 13.705 | ** | 19.920 | *** | 7.761 | * | 4.128 | ns |
| Lettuce (shoots) | ||||||||||||
| Uninoculated control | 3.39 (±0.030) | a | 3842 (±149) | a | 50,659 (±2025) | a | 2988 (±94) | ab | 9604 (±397) | b | 642 (±67) | a |
| PEPV15 | 3.43 (±0.059) | a | 4238 (±168) | a | 52,011 (±2210) | a | 2877 (±49) | b | 9949 (±123) | b | 766 (±106) | a |
| PEPV16 | 3.62 (±0.083) | a | 4243 (±113) | a | 54,865 (±1597) | a | 3385 (±140) | a | 12,576 (±446) | a | 1034 (±179) | a |
| Mean Square | 0.058 | 212,160.780 | 18,436,322.965 | 285,011.620 | 10,572,050.235 | 160,557.838 | ||||||
| F value and significance | 3.862 | ns | 2.515 | ns | 1.199 | ns | 6.883 | ** | 21.334 | *** | 2.517 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores-Félix, J.D.; Velázquez, E.; Martínez-Molina, E.; González-Andrés, F.; Squartini, A.; Rivas, R. Connecting the Lab and the Field: Genome Analysis of Phyllobacterium and Rhizobium Strains and Field Performance on Two Vegetable Crops. Agronomy 2021, 11, 1124. https://doi.org/10.3390/agronomy11061124
Flores-Félix JD, Velázquez E, Martínez-Molina E, González-Andrés F, Squartini A, Rivas R. Connecting the Lab and the Field: Genome Analysis of Phyllobacterium and Rhizobium Strains and Field Performance on Two Vegetable Crops. Agronomy. 2021; 11(6):1124. https://doi.org/10.3390/agronomy11061124
Chicago/Turabian StyleFlores-Félix, José David, Encarna Velázquez, Eustoquio Martínez-Molina, Fernando González-Andrés, Andrea Squartini, and Raúl Rivas. 2021. "Connecting the Lab and the Field: Genome Analysis of Phyllobacterium and Rhizobium Strains and Field Performance on Two Vegetable Crops" Agronomy 11, no. 6: 1124. https://doi.org/10.3390/agronomy11061124