
Alleviation of Sodic Stress in Rice by Exploring the Exopolysaccharide-Producing Sodic-Tolerant Bacteria
 , ,
, ,  ,
,
Abstract
:
1. Introduction
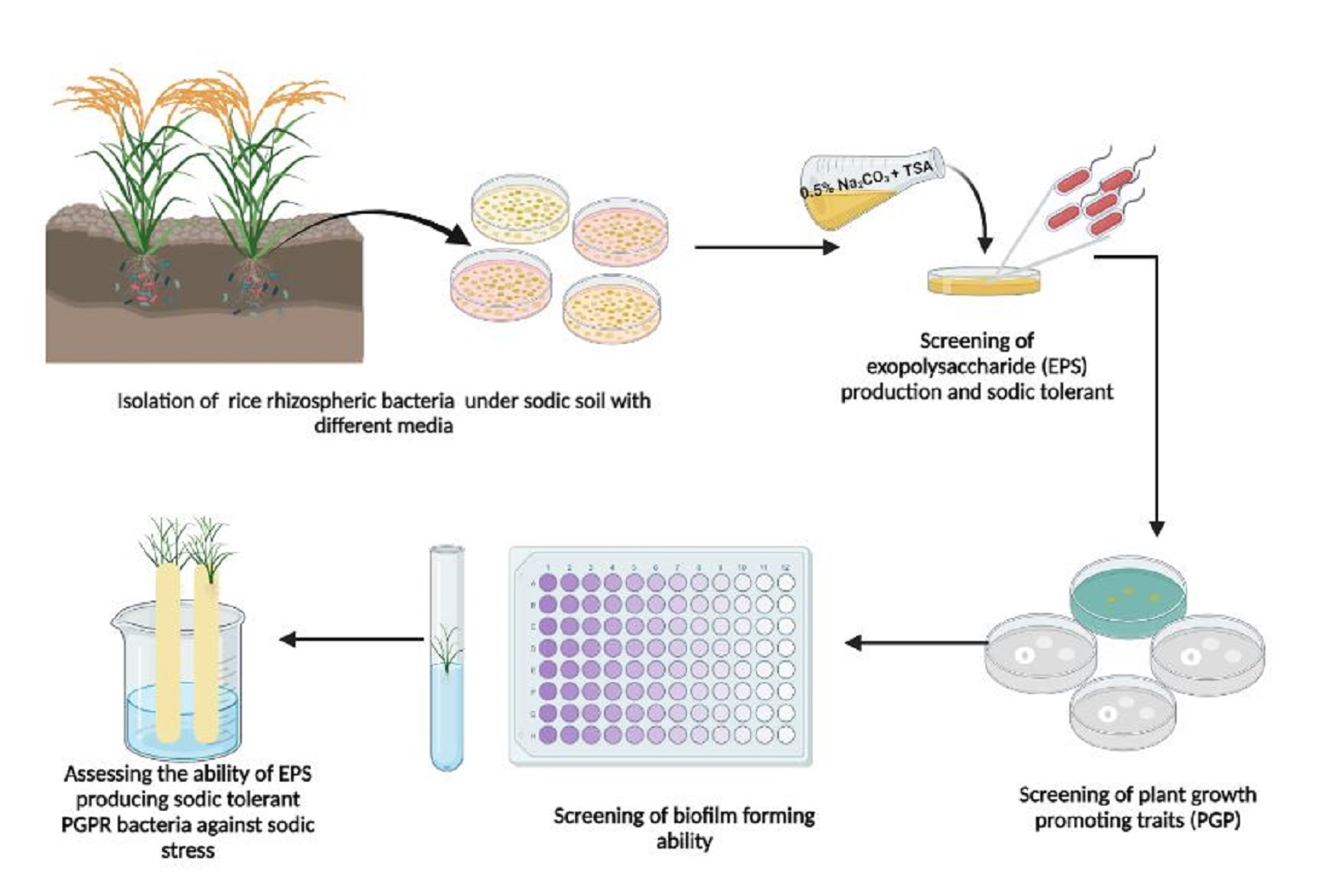
2. Materials and Methods
2.1. Bacteria Isolation
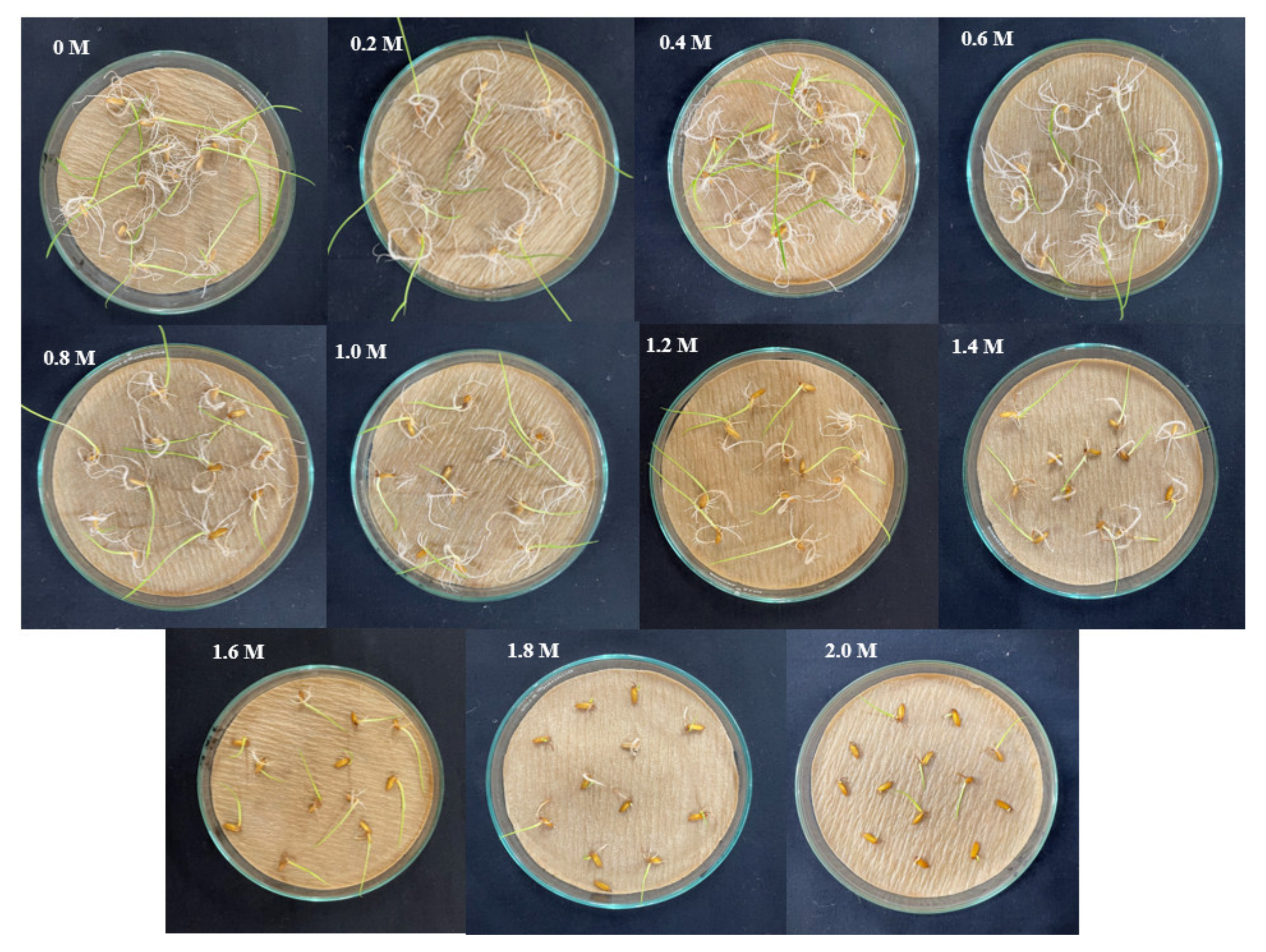
2.2. Standardization of Medium for Inducing Sodicity
2.3. Screening for Exopolysaccharide Production
2.4. DNA Extraction
2.5. Screening of Sodic Tolerance
2.6. Screening for Plant Growth Promoting (PGP) Traits
2.7. Estimation of Osmolytes Production
2.8. Biofilm Formation
2.9. Alleviation of Sodic Stress in Rice
2.10. Bioassay for Biofilm Forming Ability
2.11. Assessing the Intracellular Na+ Concentration in Bacterial Isolates
2.12. Aggregation Formation
2.13. SEM Imaging of Unstressed and Stressed Cultures
2.14. Statistical Analysis
3. Results
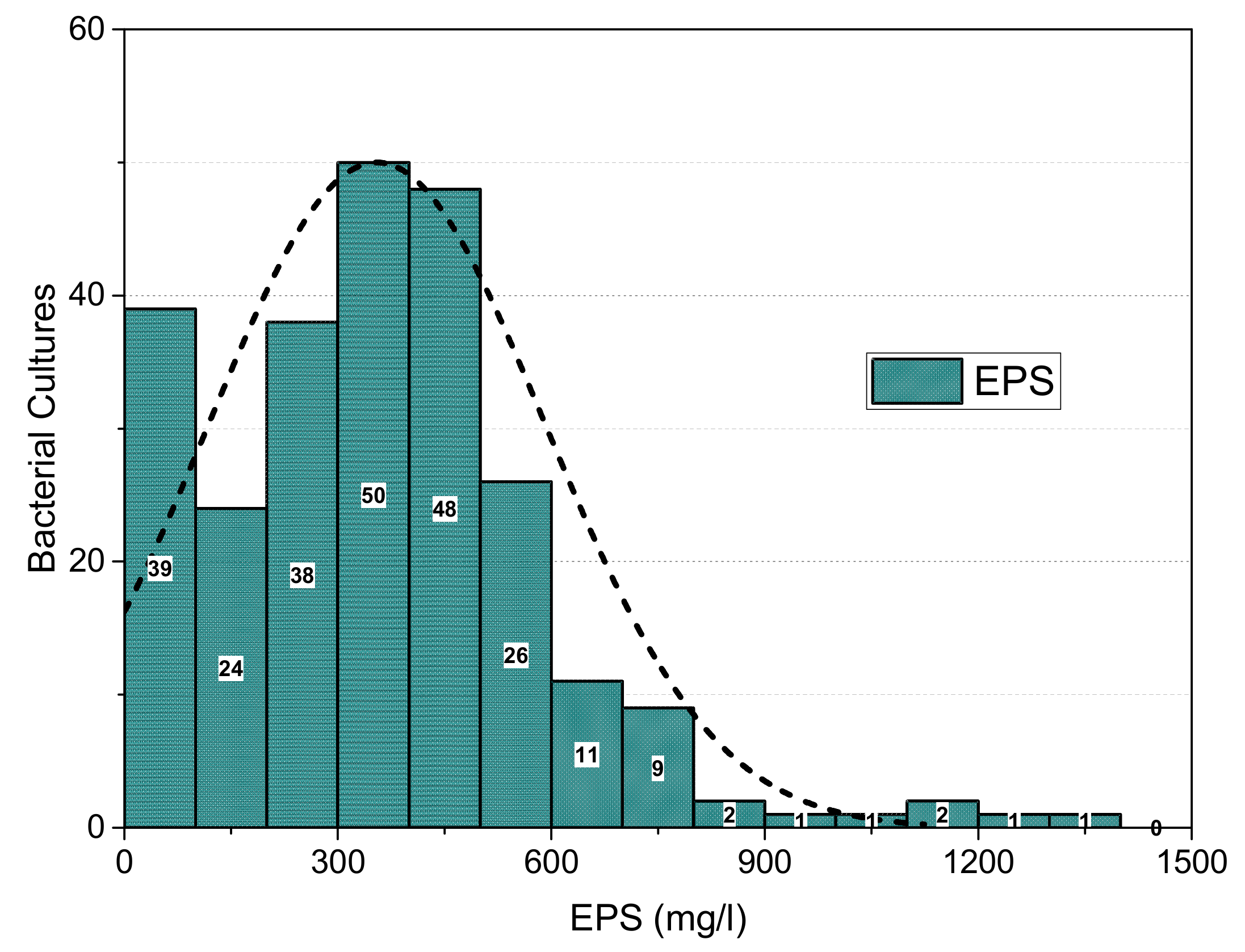
3.1. Isolation and Screening of EPS-Producing Bacteria
3.2. PCR and 16S rRNA Analysis of Bacterial Isolates
3.3. Standardization of Media for Inducing Sodicity
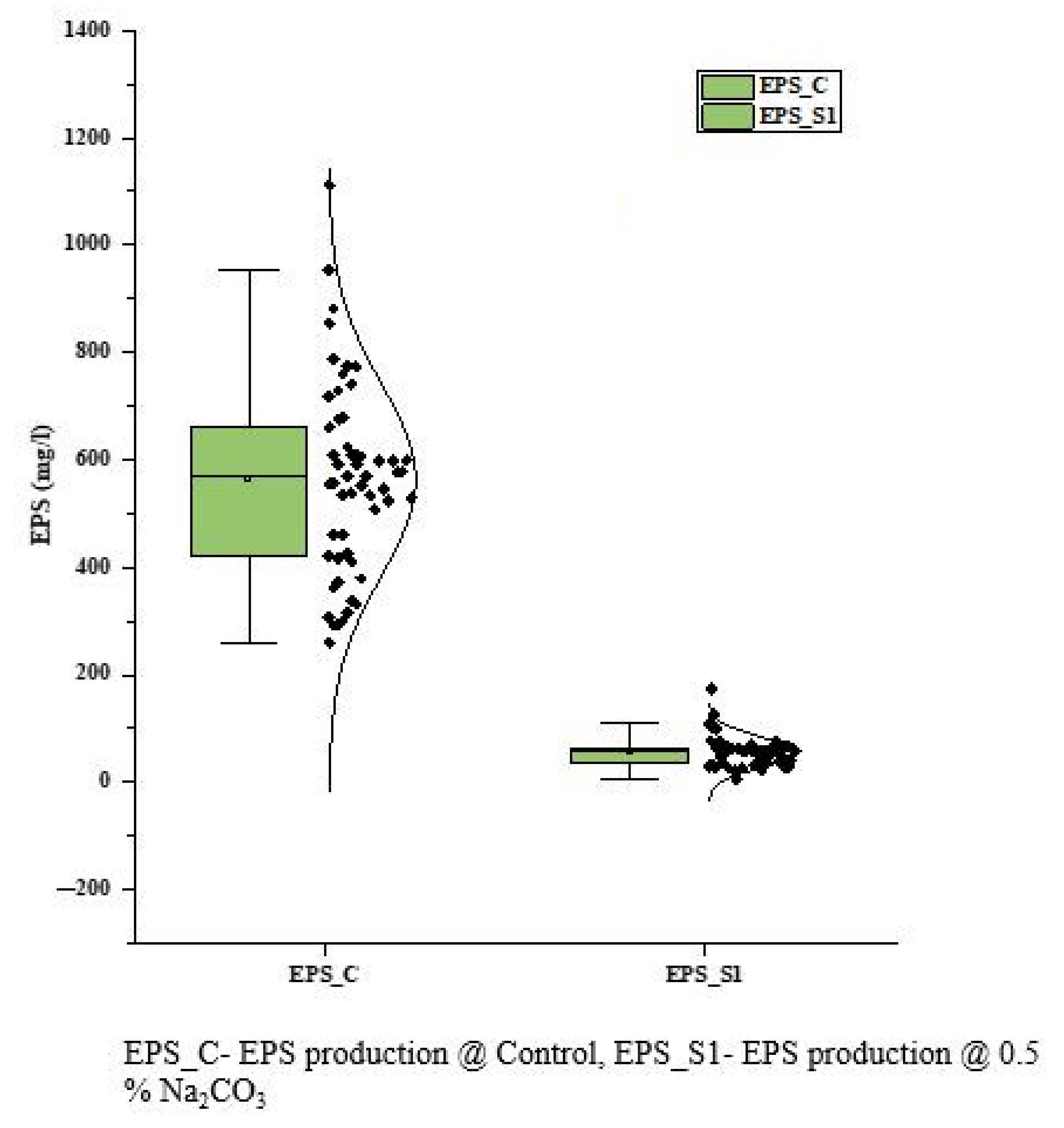
3.4. Screening for Exopolysaccharide Production
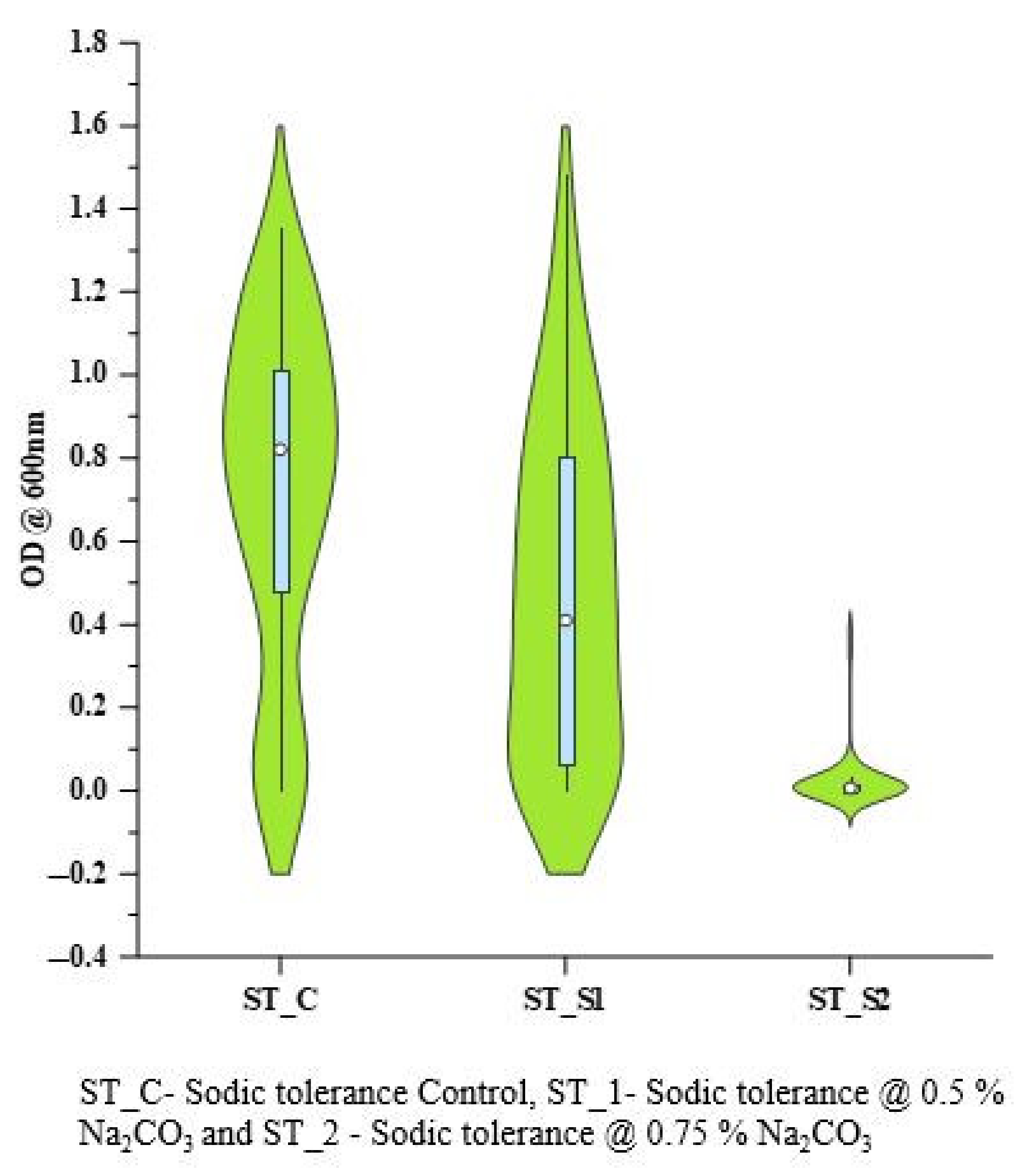
3.5. Screening for Sodicity Tolerance
3.6. Assessing the Plant Growth Promoting Traits and Osmoprotectant Accumulation of Selected EPS-Producing Sodic-Tolerant Bacteria
3.7. Screening the Biofilm-Forming Ability of Bacterial Isolates
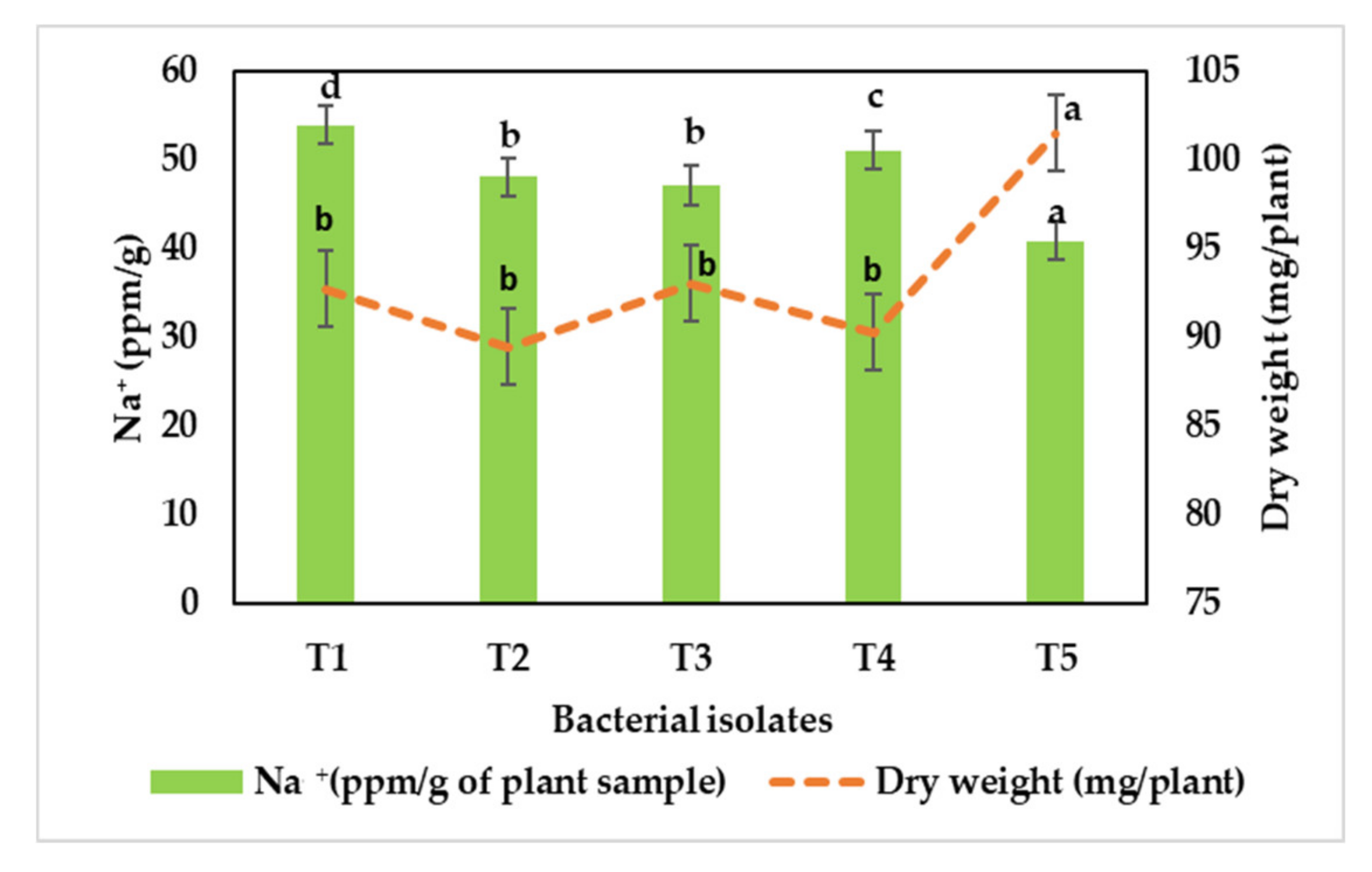
3.8. Intracellular Sodium Accumulation in Bacterial Cells
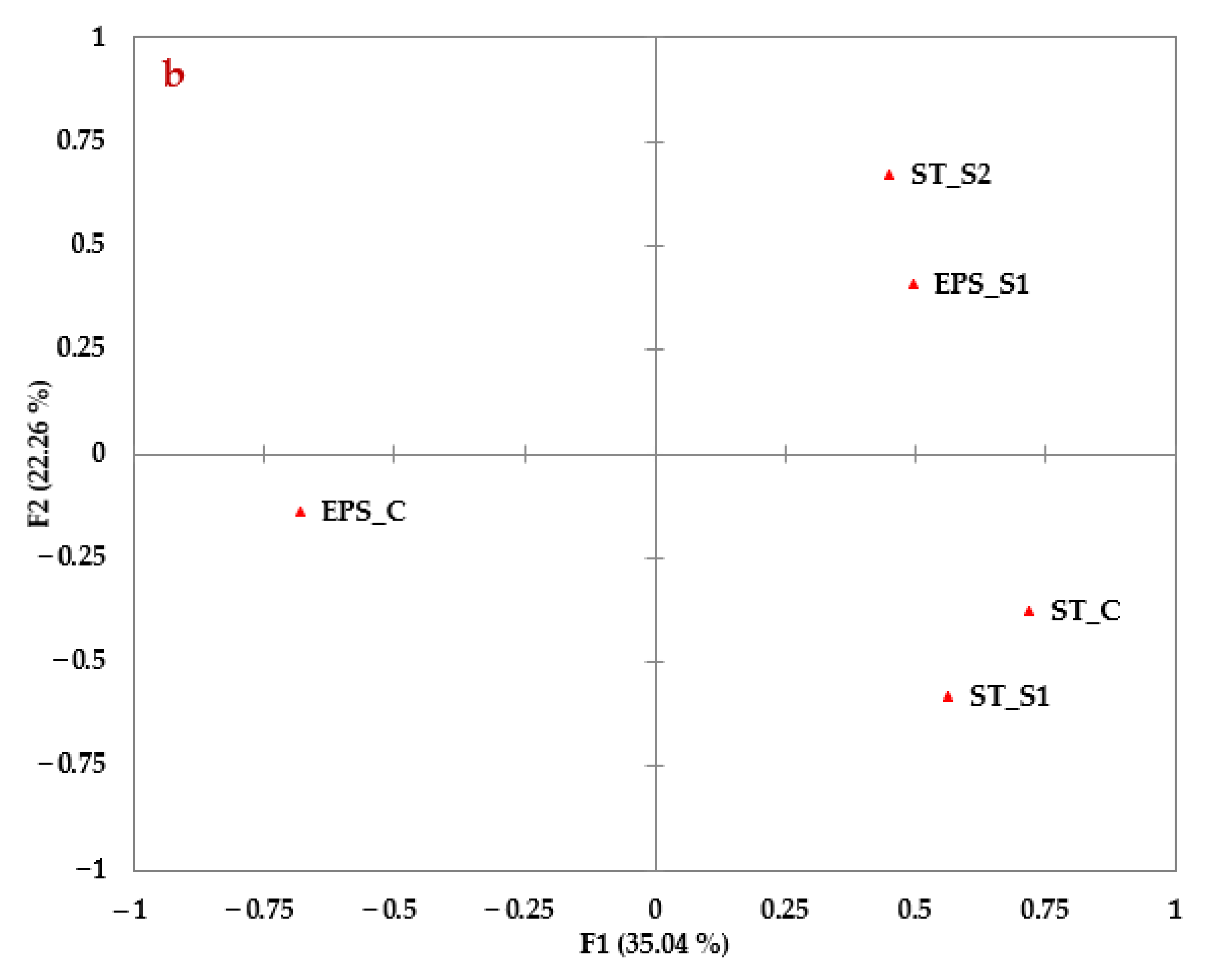
3.9. Aggregation Dispersity Test
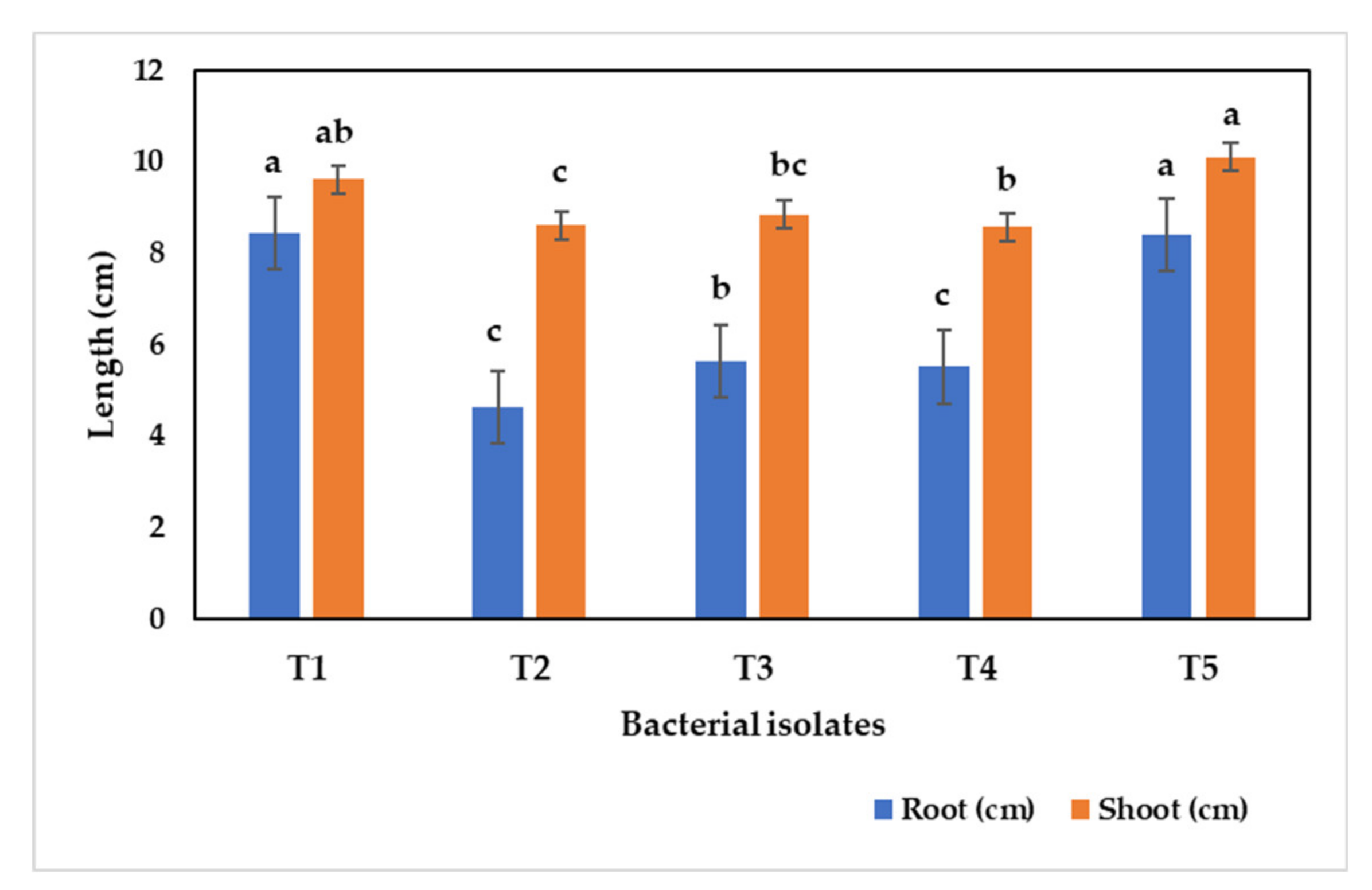
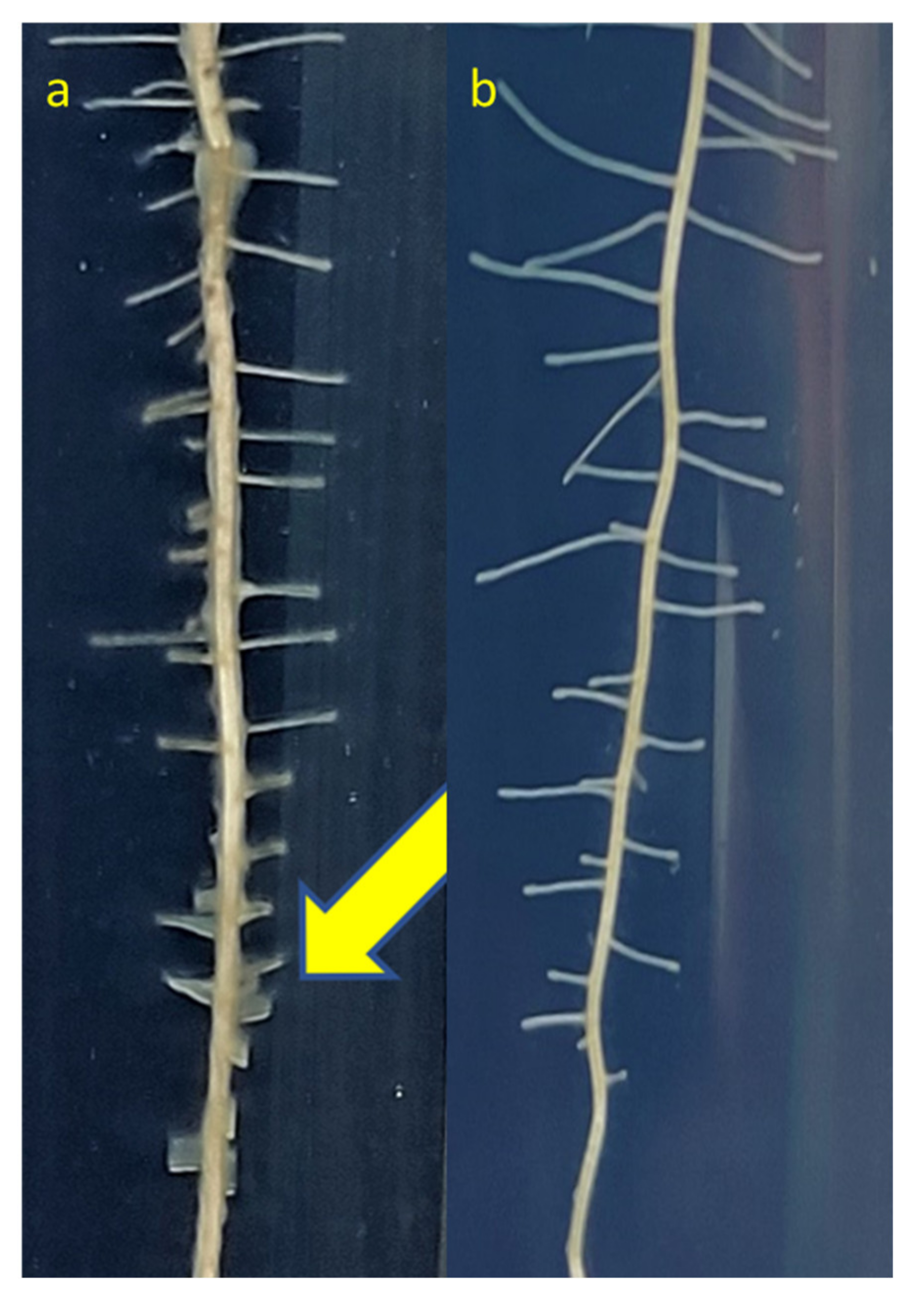
3.10. Alleviation of Sodic Stress
3.11. In Vitro Study on Biofilm Formation
3.12. SEM Image
4. Discussion
4.1. Media Optimization for Isolation of the Sodic Tolerant Bacteria
4.2. Influence of Sodic Stress on PGPtraits
4.3. Alleviation of Sodic Stress under In Vitro Condition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. Status of the World’s Soil Resources (SWSR)–Main Report Rome; FAO: Rome, Italy, 2015. [Google Scholar]
- Sharma, D.; Singh, A. Salinity research in India-achievements, challenges and future prospects. Water Energy Int. 2015, 58, 35–45. [Google Scholar]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef]
- Islam, F.; Yasmeen, T.; Arif, M.S.; Ali, S.; Ali, B.; Hameed, S.; Zhou, W. Plant growth promoting bacteria confer salt tolerance in Vigna radiata by up-regulating antioxidant defense and biological soil fertility. Plant Growth Regul. 2016, 80, 23–36. [Google Scholar] [CrossRef]
- Ashraf, M.; Hasnain, S.; Berge, O.; Mahmood, T. Inoculating wheat seedlings with exopolysaccharide-producing bacteria restricts sodium uptake and stimulates plant growth under salt stress. Biol. Fertil. Soils 2004, 40, 157–162. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Jabborova, D.; Berg, G. Synergistic interactions between Bradyrhizobium japonicum and the endophyte Stenotrophomonas rhizophila and their effects on growth, and nodulation of soybean under salt stress. Soil Biol. Biochem. 2016, 405, 35–45. [Google Scholar] [CrossRef]
- Tewari, S.; Arora, K. Talc based exopolysaccharides formulation enhancing growth and production of Hellianthus annuus under saline conditions. Cell. Mol. Biol. 2014, 60, 73–81. [Google Scholar]
- Upadhyay, S.; Singh, J.; Singh, D. Exopolysaccharide-producing plant growth-promoting rhizobacteria under salinity condition. Pedosphere 2011, 21, 214–222. [Google Scholar] [CrossRef]
- Damodaran, T.; Sharma, D.; Mishra, V.; Jha, S.; Kannan, R.; Sah, V.; Rai, R. Isolation of salt tolerant endophyticand rhizospheric bacteria by natural selection and screening for promising plant growth promoting rhizobacteria [PGPR] and growth vigour in Tomato under sodic soil. Afr. J. Microbiol. Res. 2013, 7, 5082–5089. [Google Scholar]
- Tewari, S.; Arora, N.K. Multifunctional exopolysaccharides from Pseudomonas aeruginosa PF23 involved in plant growth stimulation, biocontrol and stress amelioration in sunflower under saline conditions. Curr. Microbiol. 2014, 69, 484–494. [Google Scholar] [CrossRef]
- Damodaran, T.; Mishra, V.; Jha, S.; Pankaj, U.; Gupta, G.; Gopal, R. Identification of rhizosphere bacterial diversity with promising salt tolerance, PGP traits and their exploitation for seed germination enhancement in sodic soil. Agric. Res. 2019, 8, 36–43. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J. Relevance of microbial extracellular polymeric substances (EPSs)-Part I: Structural and ecological aspects. Water Sci. Technol. 2001, 43, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Neu, T.R.; Lawrence, J.R. In Situ Characterization of Extracellular Polymeric Substances (EPS) in Biofilm Systems. In Microbial Extracellular Polymeric Substances; Wingender, J., Neu, T.R., Flemming, H.C., Eds.; Springer: Berlin, Heidelberg, 1999; pp. 21–47. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Mayer, C.; Korstgens, V.; Borchard, W. Cohesiveness in biofilm matrix. In Community Structure and Co-Operation in Biofilms; Cambridge University Press: Cambridge, UK, 2000; pp. 87–105. [Google Scholar]
- Alami, Y.; Achouak, W.; Marol, C.; Heulin, T. Rhizosphere soil aggregation and plant growth promotion of sunflowers by an exopolysaccharide-producing Rhizobium sp. strain isolated from sunflower roots. Appl. Environ. Microbiol. 2000, 66, 3393–3398. [Google Scholar] [PubMed] [Green Version]
- Shultana, R.; Kee Zuan, A.T.; Yusop, M.R.; Saud, H.M. Characterization of salt-tolerant plant growth-promoting rhizobacteria and the effect on growth and yield of saline-affected rice. PLoS ONE 2020, 15, e0238537. [Google Scholar] [CrossRef]
- Shah, G.; Jan, M.; Afreen, M.; Anees, M.; Rehman, S.; Daud, M.; Malook, I.; Jamil, M. Halophilic bacteria mediated phytoremediation of salt-affected soils cultivated with rice. J. Geochem. Explor. 2017, 174, 59–65. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.t.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192. [Google Scholar] [CrossRef]
- Pikovskaya, R. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Sharon, J.; Hathwaik, L.; Glenn, G.; Imam, S.; Lee, C. Isolation of efficient phosphate solubilizing bacteria capable of enhancing tomato plant growth. J. Soil Sci. Plant Nutr. 2016, 16, 525–536. [Google Scholar] [CrossRef]
- Anandham, R.; Sridar, R.; Nalayini, P.; Poonguzhali, S.; Madhaiyan, M. Potential for plant growth promotion in groundnut (Arachis hypogaea L.) cv. ALR-2 by co-inoculation of sulfur-oxidizing bacteria and Rhizobium. Microbiol. Res. 2007, 162, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Qurashi, A.W.; Sabri, A.N. Osmolyte accumulation in moderately halophilic bacteria improves salt tolerance of chickpea. Pak. J. Bot. 2013, 45, 1011–1016. [Google Scholar]
- Bates, S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter dish biofilm formation assay. Vis. Exp. 2011, 47, 2437. [Google Scholar] [CrossRef] [PubMed]
- Balasundararajan, V.; Dananjeyan, B. Occurrence of diversified N-acyl homoserine lactone mediated biofilm-forming bacteria in rice rhizoplane. J. Basic Microbiol. 2019, 59, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Omuto, C.T.; Vargas, R.R.; El Mobarak, A.M.; Mohamed, N.; Viatkin, K.Y.Y. Mapping of Salt-Affected Soils: Technical Manua; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Ibarra-Villarreal, A.L.; Gándara-Ledezma, A.; Godoy-Flores, A.D.; Herrera-Sepúlveda, A.; Díaz-Rodríguez, A.M.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Salt-tolerant Bacillus species as a promising strategy to mitigate the salinity stress in wheat (Triticum turgidum subsp. durum). J. Arid Environ. 2021, 186, 104399. [Google Scholar] [CrossRef]
- Rousk, J.; Elyaagubi, F.K.; Jones, D.L.; Godbold, D.L. Bacterial salt tolerance is unrelated to soil salinity across an arid agroecosystem salinity gradient. Soil Biol. Biochem. 2011, 43, 1881–1887. [Google Scholar] [CrossRef]
- Soleimani, R.; Alikhani, H.; Towfighi, H.; Khavazi, K.; Pourbabaee, A. Isolated bacteria from saline–sodic soils alter the response of wheat under high adsorbed sodium and salt stress. Int. J. Environ. Sci. Technol. 2017, 14, 143–150. [Google Scholar] [CrossRef]
- Kailasan, N.S.; Vamanrao, V.B. Isolation and characterization of Ralstonia pickettii-A novel phosphate solubilizing bacterium from Pomegranate rhizosphere from Western India. Int. J. Life Sci. Biotechnol. Pharma Res. 2015, 4, 1. [Google Scholar]
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef]
- Sekhar, V.C.; Nampoothiri, K.M.; Mohan, A.J.; Nair, N.R.; Bhaskar, T.; Pandey, A. Microbial degradation of high impact polystyrene (HIPS), an e-plastic with decabromodiphenyl oxide and antimony trioxide. J. Hazard. Mater. 2016, 318, 347–354. [Google Scholar] [CrossRef]
- Shabbir, S.; Faheem, M.; Ali, N.; Kerr, P.G.; Wang, L.-F.; Kuppusamy, S.; Li, Y. Periphytic biofilm: An innovative approach for biodegradation of microplastics. Sci. Total Environ. 2020, 717, 137064. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.G.; Hinduja, M.; Sujitha, K.; Rajan, N.N.; Dharani, G. Biodegradation of polystyrene by deep-sea Bacillus paralicheniformis G1 and genome analysis. Sci. Total Environ. 2021, 774, 145002. [Google Scholar] [CrossRef]
- Dunlap, C.A.; Kwon, S.-W.; Rooney, A.P.; Kim, S.-J. Bacillus paralicheniformis sp. nov., isolated from fermented soybean paste. Int. J. Syst. Evol. Microbiol. 2015, 65, 3487–3492. [Google Scholar] [CrossRef]
- Kearns, D.B.; Chu, F.; Branda, S.S.; Kolter, R.; Losick, R. A master regulator for biofilm formation by Bacillus subtilis. Mol. Microbiol. 2005, 55, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Dubnau, E.J.; Carabetta, V.J.; Tanner, A.W.; Miras, M.; Diethmaier, C.; Dubnau, D. A protein complex supports the production of Spo0A-P and plays additional roles for biofilms and the K-state in Bacillus subtilis. Mol. Microbiol. 2016, 101, 606–624. [Google Scholar] [CrossRef]
- Ventosa, A.; Nieto, J.J.; Oren, A. Biology of moderately halophilic aerobic bacteria. Microbiol. Mol. Biol. Rev. 1998, 62, 504–544. [Google Scholar] [CrossRef]
- Arora, S.; Patel, P.N.; Vanza, M.J.; Rao, G. Isolation and characterization of endophytic bacteria colonizing halophyte and other salt tolerant plant species from coastal Gujarat. Afr. J. Microbiol. Res. 2014, 8, 1779–1788. [Google Scholar]
- Arora, S.; Vanza, M.J.; Mehta, R.; Bhuva, C.; Patel, P.N. Halophilic microbes for bio-remediation of salt affected soils. Afr. J. Microbiol. Res. 2014, 8, 3070–3078. [Google Scholar]
- Sandhya, V.; Shaik, Z.A.; Grover, M.; Reddy, G.; Venkateswarlu, B. Alleviation of drought stress effects in sunflower seedlings by the exopolysaccharides producing Pseudomonas putida strain GAP-P45. Biol. Fertil. Soils 2009, 46, 17–26. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | IAA (mg/L) | Zn Solubilization (%) | Siderophore Production | |
|---|---|---|---|---|
| Without Stress | With Stress | |||
| Rhodococcus pyridinivorans L3C9N | 2.5 ± 0.3 ef | 2.1 ± 0.4 a | − | + |
| Bacillus velezensis L2C3L | 1.4 ± 0.2 i | 0.6 ± 0.1 hi | − | + |
| Cytobacillus firmus L1C4L | 8.8 ± 0.3 a | 1.2 ± 0.1 cd | − | − |
| Bacillus paralicheniformis L1C5L | 2.9 ± 0.2 cd | 0.3 ± 0.1 jk | 260 | + |
| Ralstonia pickettii L4C6L | 0.7 ± 0.0 jk | 0.9 ± 0.2 fg | 520 | − |
| Arthrobacter sp. L5C8L | 2.6 ± 0.3 efg | 0.0 ± 0.0 l | − | − |
| Niallia circulans L2C9L | 1.9 ± 0.0 hi | 0.5 ± 0.1 i | − | + |
| Staphylococcus sp. L2C2T | 0.7 ± 0.0 jk | 0.1 ± 0.0 kl | − | − |
| Burkholderia territorii L2C6T | 0.7 ± 0.0 jk | 0.4 ± 0.1 ij | 188 | + |
| Stenotrophomonas maltophilia L2C7T | 1.9 ± 0.1 hi | 1.4 ± 0.1 b | − | + |
| Bacillus stercoris L3C2T | 1.5 ± 0.2 hi | 1.3 ± 0.2 bc | − | + |
| Bacillus sp. L3C3T | 0.9 ± 0.1 j | 1.1 ± 0.1 cd | − | − |
| Bacillus cabrialesii L1C5T | 0.5 ± 0.1 k | 0.8 ± 0.1 gh | − | + |
| Bacillus safensis L5C13T | 0.7 ± 0.0 jk | 0.8 ± 0.1 gh | 500 | − |
| Pseudomonas sp. L5C14T | 3.7 ± 0.1 b | 0.9 ± 0.0 fg | 186 | + |
| Burkholderia cepacian L4C7T | 2.2 ± 0.1 g | 1.0 ± 0.1 de | 300 | + |
| Bacillus rugosus L1C7T | 3.0 ± 0.2 cd | 0.6 ± 0.1 i | 140 | + |
| Bacillus tequilensis L3C6T | 1.7 ± 0.1 hi | 1.0 ± 0.2 de | − | + |
| Kocuria sp. L2C2R | 3.1 ± 0.2 c | 0.2 ± 0.0 jk | − | − |
| Franconibacter helveticus L2C1L2 | 0.9 ± 0.1 j | 1.1 ± 0.1 cd | − | − |
| Bacillus paramycoides L4C1L2 | 2.7 ± 0.3 de | 0.6 ± 0.1 hi | − | − |
| Bacillus zanthoxyli L2C2L2 | 2.6 ± 0.2 ef | 1.2 ± 0.0 cd | − | − |
| Acinetobacter sp. L5C6L2 | 2.3 ± 0.0 fg | 0.4 ± 0.1 ij | − | − |
| Bacillus coreaensis L1C1T2 | 2.5 ± 0.3 efg | 0.6 ± 0.0 hi | − | + |
| Achromobacter sp. L1C9T2 | 0.0 ± 0.0 l | 0.9 ± 0.0 ef | 300 | − |
| Bacillus altitudinis L3C3T2 | 2.3 ± 0.0 g | 1.0 ± 0.0 de | 600 | + |
| Bacteria | Proline (mg/L) | Trehalose (mg/L) | Glycine Betaine (mg/L) | |||
|---|---|---|---|---|---|---|
| Without Stress | With Stress | Without Stress | With Stress | Without Stress | With Stress | |
| Rhodococcus pyridinivorans L3C9N | 39.8 ± 5.1 bc | 8.2 ± 1.3 cd | 161 ± 22 b | 92 ± 15 d | 232 ± 34 de | 113 ± 01 k |
| Bacillus velezensis L2C3L | 16.4 ± 1.9 ij | 7.2 ± 0.8 de | 106 ± 16 ef | 52 ± 09 fg | 248 ± 39 cd | 124 ± 30 jk |
| Cytobacillus firmus L1C4L | 39.6 ± 1.7 bc | 4.8 ± 0.6 ij | 131 ± 13 bc | 16 ± 01 lm | 209 ± 24 de | 163 ± 11 hi |
| Bacillus paralicheniformis L1C5L | 29.2 ± 7.1 ef | 9.8 ± 1.9 bc | 156 ± 30 bc | 111 ± 23 c | 307 ± 58 bc | 139 ± 26 ij |
| Ralstonia pickettii L4C6L | 35.8 ± 7.1 cd | 5.5 ± 1.0 fg | 156 ± 40 bc | 14 ± 3.5 lm | 266 ± 66 cd | 155 ± 33 hi |
| Arthrobacter sp. L5C8L | 37.8 ± 2.0 bc | 7.4 ± 0.6 de | 75 ± 04 hi | 14 ± 01 lm | 263 ± 31 cd | 273 ± 50 b |
| Niallia circulans L2C9L | 43.8 ± 7.3 bc | 7.0 ± 1.1 de | 88 ± 15 gh | 42 ± 05 hi | 190 ± 18 ef | 374 ± 40 a |
| Staphylococcus sp. L2C2T | 47.4 ± 0.3 ab | 7.8 ± 0.3 de | 107 ± 04 ef | 65 ± 02 ef | 256 ± 07 cd | 225 ± 08 cd |
| Burkholderia territorii L2C6T | 40.1 ± 5.6 bc | 5.7 ± 0.7 fg | 122 ± 19 de | 52 ± 09 fg | 280 ± 50 bcd | 229 ± 26 cd |
| Stenotrophomonas maltophilia L2C7T | 47.3 ± 3.2 ab | 6.4 ± 0.3 de | 65 ± 06 kl | 51 ± 03 fg | 238 ± 17 cd | 226 ± 19 cd |
| Bacillus stercoris L3C2T | 38.1 ± 6.3 bc | 6.7 ± 1.0 de | 73 ± 11 jk | 61 ± 09 ef | 337 ± 15 b | 214 ± 01 ef |
| Bacillus sp. L3C3T | 46.4 ± 5.0 ab | 7.1 ± 0.6 de | 121 ± 11 de | 60 ± 03 fg | 254 ± 42 cd | 276 ± 42 b |
| Bacillus cabrialesii L1C5T | 46.5 ± 8.4 ab | 5.1 ± 8.4 hi | 225 ± 08 a | 35 ± 03 jk | 277 ± 50 bc | 262 ± 22 bc |
| Bacillus safensis L5C13T | 53.7 ± 7.7 a | 12.8 ± 7.7 a | 100 ± 08 ef | 26 ± 03 kl | 445 ± 77 a | 222 ± 33 cd |
| Pseudomonas sp. L5C14T | 35.5 ± 1.3 cd | 5.2 ± 1.3 hi | 56 ± 01 l | 41 ± 02 ij | 229 ± 06 de | 131 ± 01 jk |
| Burkholderia cepacian L4C7T | 29.3 ± 3.1 ef | 4.0 ± 3.1 jk | 124 ± 04 de | 55 ± 01 g | 234 ± 20 de | 111 ± 08 k |
| Bacillus rugosus L1C7T | 32.6 ± 7.7 de | 5.4 ± 7.7 gh | 72 ± 08 jk | 41 ± 01 ij | 168 ± 39 gh | 172 ± 40 gh |
| Bacillus tequilensis L3C6T | 35.9 ± 6.8 cd | 13.4 ± 6.8 a | 126 ± 07 cd | 50 ± 15 gh | 161 ± 43 hi | 142 ± 35 ij |
| Kocuria sp. L2C2R | 31.8 ± 3.4 de | 10.3 ± 3.4 b | 62 ± 03 kl | 59 ± 01 fg | 232 ± 12 de | 159 ± 12 hi |
| Franconibacter helveticus L2C1L2 | 35.0 ± 5.6 cd | 3.4 ± 5.9 k | 65 ± 06 l | 32 ± 02 jk | 128 ± 15 i | 164 ± 25 hi |
| Bacillus paramycoides L4C1L2 | 26.6 ± 2.7 gh | 3.3 ± 2.7 k | 62 ± 03 kl | 132.8 ± 03 b | 261 ± 25 cd | 210 ± 15 ef |
| Bacillus zanthoxyli L2C2L2 | 10.5 ± 0.2 j | 5.8 ± 0.2 fg | 89 ± 0.2 gh | 91.1 ± 02 d | 181 ± 05 fg | 151 ± 2.9 ij |
| Acinetobacter sp. L5C6L2 | 23.8 ± 4.2 hi | 5.9 ± 4.2 ef | 102 ± 04 ef | 74.1 ± 02 ef | 203 ± 34 ef | 127 ± 18 jk |
| Bacillus coreaensis L1C1T2 | 28.8 ± 1.2 fg | 1.3 ± 1.2 l | 90 ± 01 fg | 33.8 ± 01 jk | 165 ± 10 gh | 180 ± 09 fg |
| Achromobacter sp. L1C9T2 | 40.6 ± 1.7 bc | 3.4 ± 1.7 k | 140 ± 02 bc | 56.7 ± 02 fg | 190 ± 06 ef | 200 ± 01 ef |
| Bacillus altitudinis L3C3T2 | 36.3 ± 1.1 cd | 6.9 ± 0.1 de | 143 ± 02 bc | 149.1 ± 02 a | 241 ± 42 cd | 213 ± 30 ef |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gunasekaran, Y.; Thiyageshwari, S.; Ariyan, M.; Roy Choudhury, A.; Park, J.-H.; Selvi, D.; Chithra, L.; Anandham, R. Alleviation of Sodic Stress in Rice by Exploring the Exopolysaccharide-Producing Sodic-Tolerant Bacteria. Agriculture 2022, 12, 1451. https://doi.org/10.3390/agriculture12091451
Gunasekaran Y, Thiyageshwari S, Ariyan M, Roy Choudhury A, Park J-H, Selvi D, Chithra L, Anandham R. Alleviation of Sodic Stress in Rice by Exploring the Exopolysaccharide-Producing Sodic-Tolerant Bacteria. Agriculture. 2022; 12(9):1451. https://doi.org/10.3390/agriculture12091451
Chicago/Turabian StyleGunasekaran, Yazhini, Subramaniam Thiyageshwari, Manikandan Ariyan, Aritra Roy Choudhury, Jung-Ho Park, Duraisamy Selvi, Lakshmanan Chithra, and Rangasamy Anandham. 2022. "Alleviation of Sodic Stress in Rice by Exploring the Exopolysaccharide-Producing Sodic-Tolerant Bacteria" Agriculture 12, no. 9: 1451. https://doi.org/10.3390/agriculture12091451