1. Introduction
Conservation agriculture (CA) is a climate-smart strategy that improves soil health [
1]. This strategy involves three interlinked principles of diversified crop rotations, no or little mechanical disturbance of the soil through tillage and the retention of organic residues on the soil surface [
2]. If applied correctly, the aforementioned principles of CA can increase soil organic matter (SOM) and consequently, soil organic carbon (SOC) [
3,
4]. Soil organic matter, which can be estimated from SOC, is considered a key attribute of soil fertility and productivity because of its influence on the physical, chemical, and biological properties of soil [
4]. Earthworms are an important component of soil biota and key in the regulation of organic matter dynamics as well as maintenance of soil structure and fertility. The earthworm’s bioturbation effect facilitates the decomposition of plant litter and other organic substrates and in turn, increases microbial activity, nutrient availability, and plant uptake [
5,
6,
7]. The primary food source for earthworms is organic matter in its various stages of decay [
8]. This suggests that any management practice that improves SOM such as CA practices could result in greater earthworm abundance and diversity. However, there is little understanding of the effect of CA principles on SOM dynamics and consequently, earthworms’ diversity and numbers.
Research suggests that earthworms play a positive role under CA systems. For example, the addition of earthworms into the soil increases vermicompost production due to increased microbial activity and bioturbation [
9]. Vermicompost increases soil’s porosity and water holding capacity by creating vertical and horizontal burrows within the soil, through which oxygen and water can enter, and the emission of carbon dioxide can take place [
10]. Increased water holding capacity is particularly key for crop production under semi-arid environments where rainfall is unreliable. Earthworms are also referred to as “bio-engineers”, often reducing the requirement for tillage. Other advantages include control of soil metal concentrations, aeration, pH, and electrical conductivity [
11]. In CA systems where residue retention is observed, the burrowing activity by earthworms can promote residue decomposition, humus formation, and nutrient cycling [
9]. On the other hand, tilling of soil negatively affects earthworm diversity and density directly through injury caused by cultivation or indirectly due to habitat disruption and reduction in food sources [
12].
Various studies have been done in the localities of the Eastern Cape on the effects of CA on the physicochemical properties of soil [
13,
14]. However, studies that describe the effects of CA principles on earthworms are lacking, especially under the semi-arid environment of the central Eastern Cape of South Africa. Knowledge of the influence of CA practices on earthworms and their relationship with soil organic matter dynamics is imperative for the development of sustainable agroecosystems. Therefore, this study was carried out to investigate the effects of tillage, crop rotations, and residue management on earthworm abundance and species richness in the semi-arid environments of the Alice Jozini ecotope, in central Eastern Cape, South Africa. The study hypothesis was that CA principles such as tillage, crop rotations, and residue management will interact to influence earthworm species richness and abundance in the semi-arid of South Africa.
4. Discussion
Three exotic earthworm genera were observed in the CA trial, which was set on a Haplic Cambisol in Alice within the Raymond Mhlaba municipality. The absence of indigenous earthworm genera in the CA trial site could be attributed to the fact that indigenous earthworms prefer natural undisturbed soils [
24] and areas with a high density of native plant species [
25]. The site at which this study was based has had no indigenous plant species for a long time (>25 years) as it has been used for field crop and forage farming. Furthermore, before trial establishment, the land had been under lucerne, while after 2012, it had been under maize, wheat, or soybean planted in rotations. This continuous cultivation could be responsible for the disappearance of indigenous earthworm species from the trial site.
The higher earthworm density observed under no-till than under CT over the first three seasons (summer, winter, and spring 2015) can be the result of a higher loss of organic matter under CT than under no-till due to accelerated oxidation of organic matter during tillage [
26]. Furthermore, van Capelle et al. [
27] described increased earthworm density under no-till as due to the interacting effects of reduced injuries, decreased exposure to predators at the soil surface, decreased microclimate changes, and increased availability of organic matter, providing a convenient food source in the upper soil layers. A contrasting trend was observed during the summer 2016–2017 season, where earthworm species density was greater under CT than under no-till for all three earthworm species. Ernst and Emmerling [
12] observed a similar trend and suggested that the abundance of earthworms under CT in some situations is likely to be related to lower soil bulk density within the tillage zone and increased transport of organic matter below the soil surface as a result of the plough effect.
Farming systems that include the use of diversified crop rotations have been reported to positively impact earthworm density [
28,
29]. The observed improved earthworm densities and biomass in rotations that had a winter crop are consistent with the findings of Schmidt et al. [
30]. According to Schmidt et al. [
30], crop rotations involving cereal and legume crops provided food and favorable nutrition, which resulted in an increase in earthworm density even during the winter season. The findings of Schimidt et al. [
30] could in part explain the greatest values of earthworm ash-free dry weight observed in the MWS in all the seasons. While the MFS had both cereal and legume crops in the rotation, the fallow period in between rendered the earthworms inactive as it appears that even under no-till (where conditions were assumed favorable), the earthworm dry weight and densities were low. The work by Schimidt et al. [
30] mirrored the present study in that earthworm data in both experiments were collected after 3 years of legume–cereal rotations under no-till and CT tillage treatments.
The positive response of earthworms to residue retention irrespective of tillage management can be due to increased moisture availability. According to Du Toit [
31] and Riley et al. [
3], the short-term influence of residue retention is the improvement in soil moisture availability, while the long-term benefit is the improvement in organic matter content. Improved soil moisture under residue retention treatments favors earthworms’ survival and ensures a stable growth of earthworms during the seasons [
32]. This input of crop residues is favorable for earthworm populations in terms of quantity, nutritional quality, and continuity of earthworm life cycles throughout the year. This could explain the improved earthworm numbers starting from the spring to the summer 2016–2017 season under the MWM and MWS rotations that had the winter wheat crop.
The overall observed variability of earthworm densities and diversity during the four different sampling seasons could have been the result of the differences in moisture and rainfall patterns over the seasons. For instance, during the summer 2015–2016 season, there were observably high temperatures coupled with very little effective rainfall, even in the previous months before sampling. This could be the reason why the lowest earthworm density was observed in the summer 2015–2016 season. The reason for such a trend is seasonal mortality because of high temperature, which prevailed in the months before the sampling of earthworms. The situation changed in summer 2016–2017 when the amount of rainfall received in this season surpassed that which was received in the preceding seasons, creating conditions that were more conducive for earthworm growth and development. The findings of the present study are consistent with findings by Smetak et al. [
33] who found that harsh environmental conditions not only limit reproduction but also the survival of adult earthworms from year to year.
The highest MBC was observed under the MWM and MWS and treatments with MFM and MFS receiving the least MBC, indicating that diversified crop rotations favored greater microbial activity. As observed earlier, these rotations also supported a greater earthworm abundance and diversity due to their ability to improve SOC. Therefore, the observed increase in microbial diversity could be attributed to the known effect of earthworms to breakdown and macerate organic matter and making it a better substrate for microbial growth. The correlations between the earthworm data and abiotic factors observed soil pH, soil moisture, SOC, and MBC as the major determinant factors of earthworm abundance measures. Dominguez and Edwards (1997) reported that environmental conditions are most likely to affect earthworm diversity through several mechanisms. A similar trend was identified by Ganihar [
34], where it was observed that natural factors such as temperature, moisture, and organic matter are also important parameters affecting the diversity of earthworms.
5. Conclusions
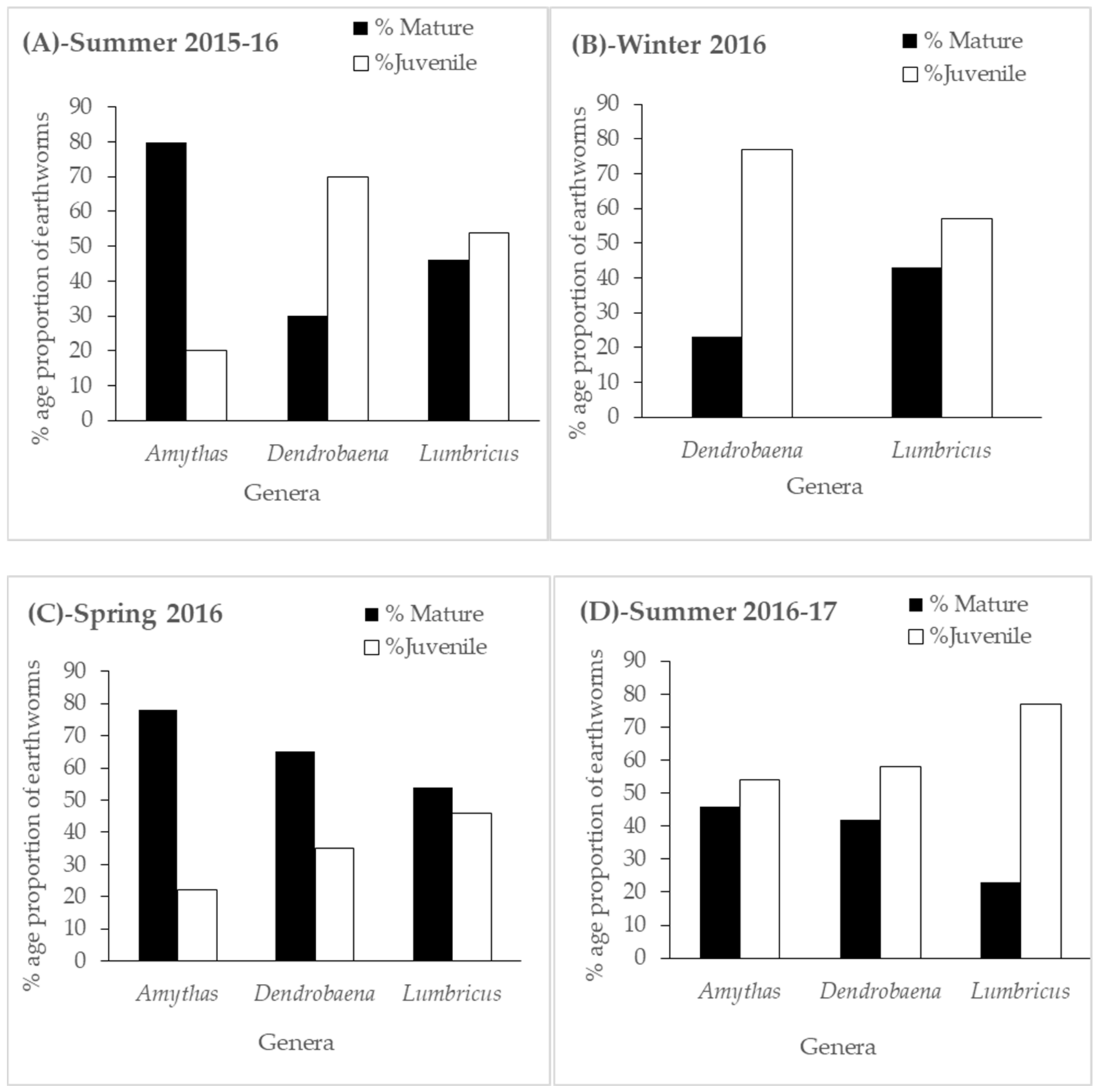
Three exotic earthworm genera were observed from the trial and these were Amynthas, Lumbricus, and Dendrobaena. Tillage, crop rotations, and residue management had a significant influence on earthworm diversity and density. No-till consistently favored earthworm abundance and diversity during the first three sampling seasons, while a contrasting trend was observed during the last sampling period of the summer 2016–2017 season. Moreover, rotations, which included cereal and legume crops together with residue retention, positively influenced earthworm diversity and density irrespective of tillage management. Earthworm parameters measured in the study significantly correlated with pH, SOC, MBC, and soil moisture, suggesting that these abiotic and biotic factors are the explanatory variables for earthworm growth and development.




{kind=link}